
Different Contribution of Monocyte- and Platelet-Derived Microvesicles to Endothelial Behavior

 ,
,
Abstract
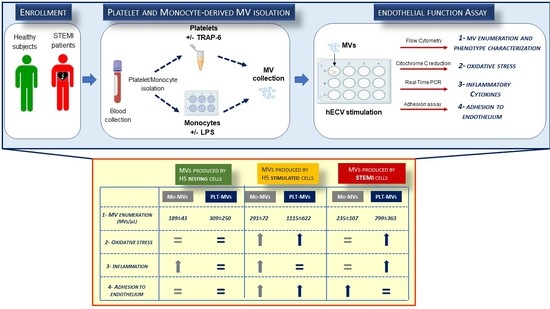
:
1. Introduction
2. Results
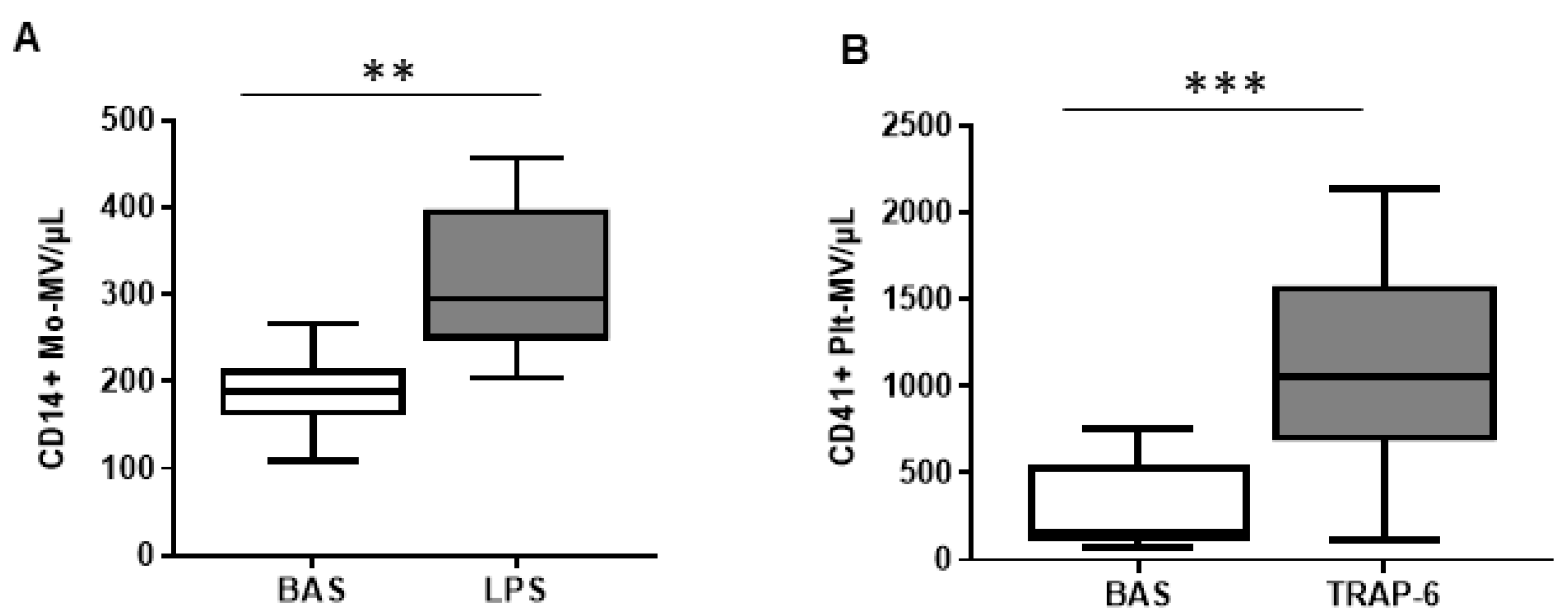
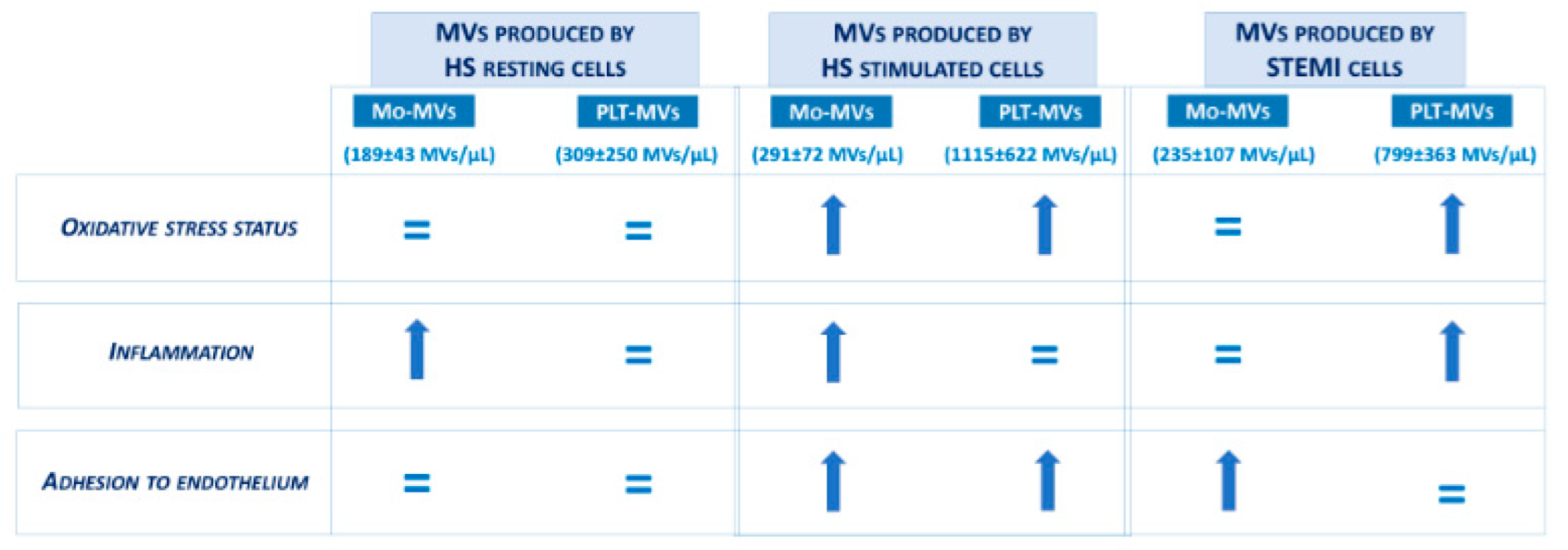
2.1. The In Vitro Ability to Generate MVs Is Higher in Platelets Than in Monocytes of Healthy Subjects
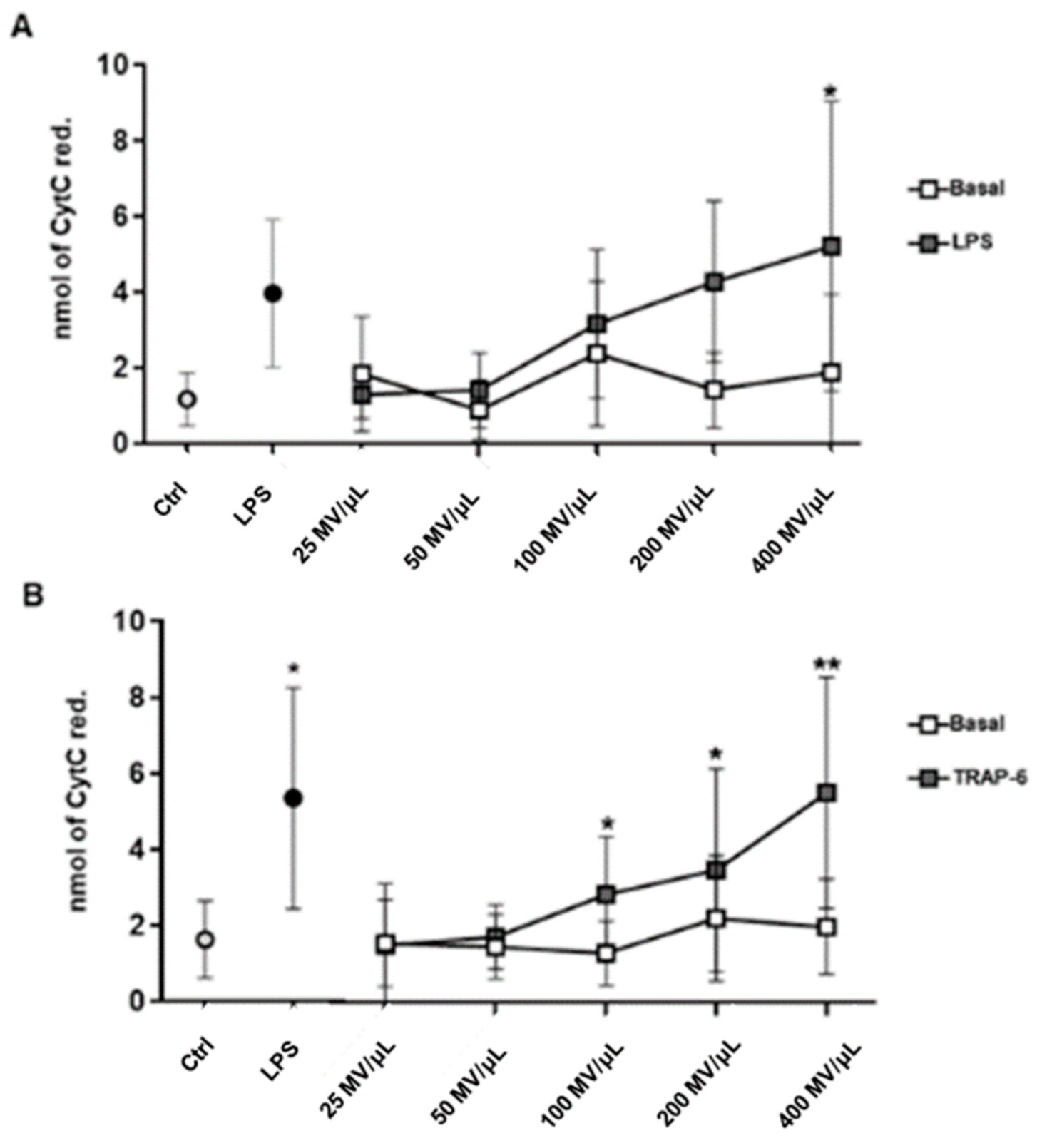
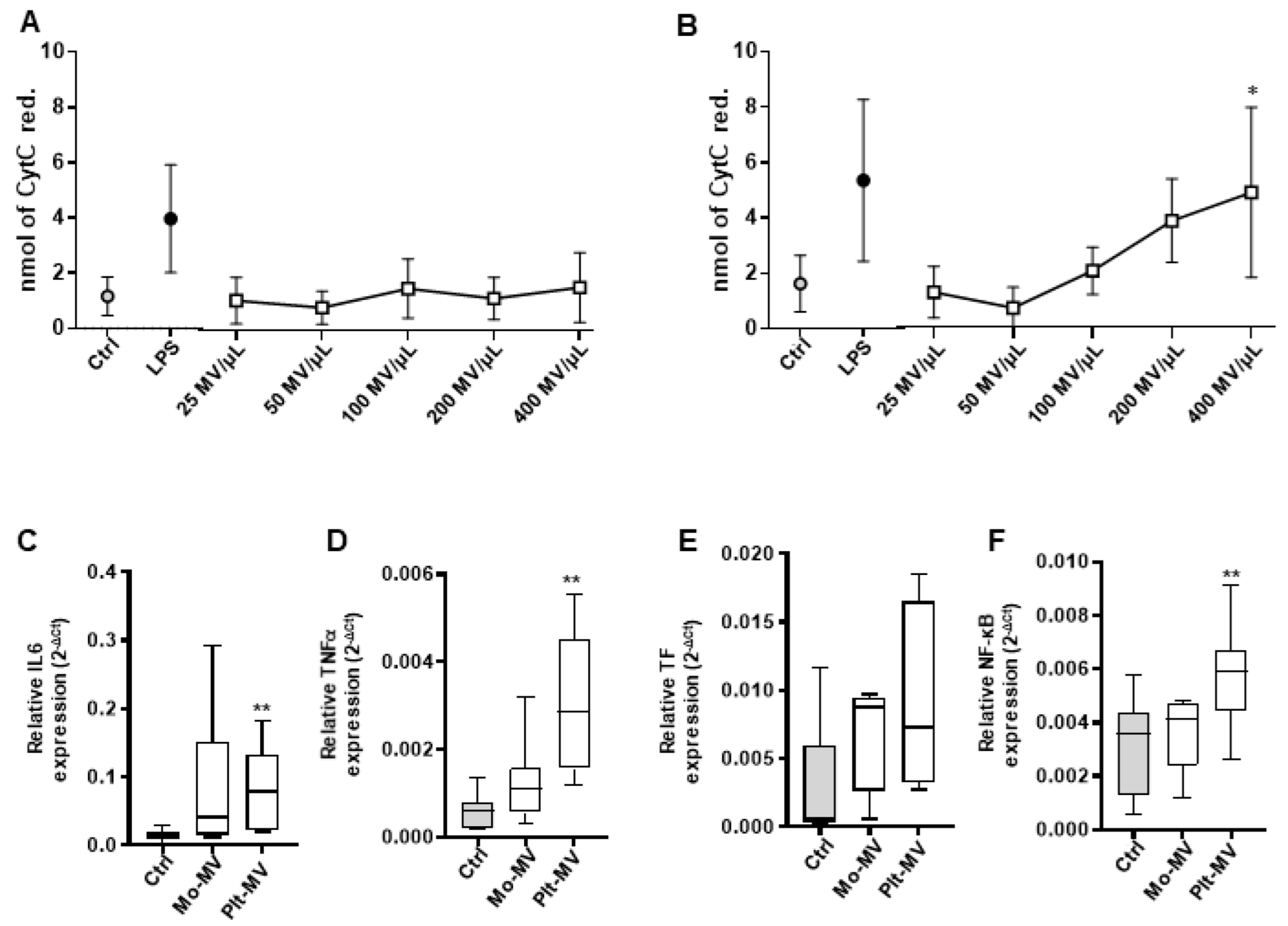
2.2. MVs Released by Activated Monocytes and Platelets Induce Oxidative Stress in Endothelial Cells
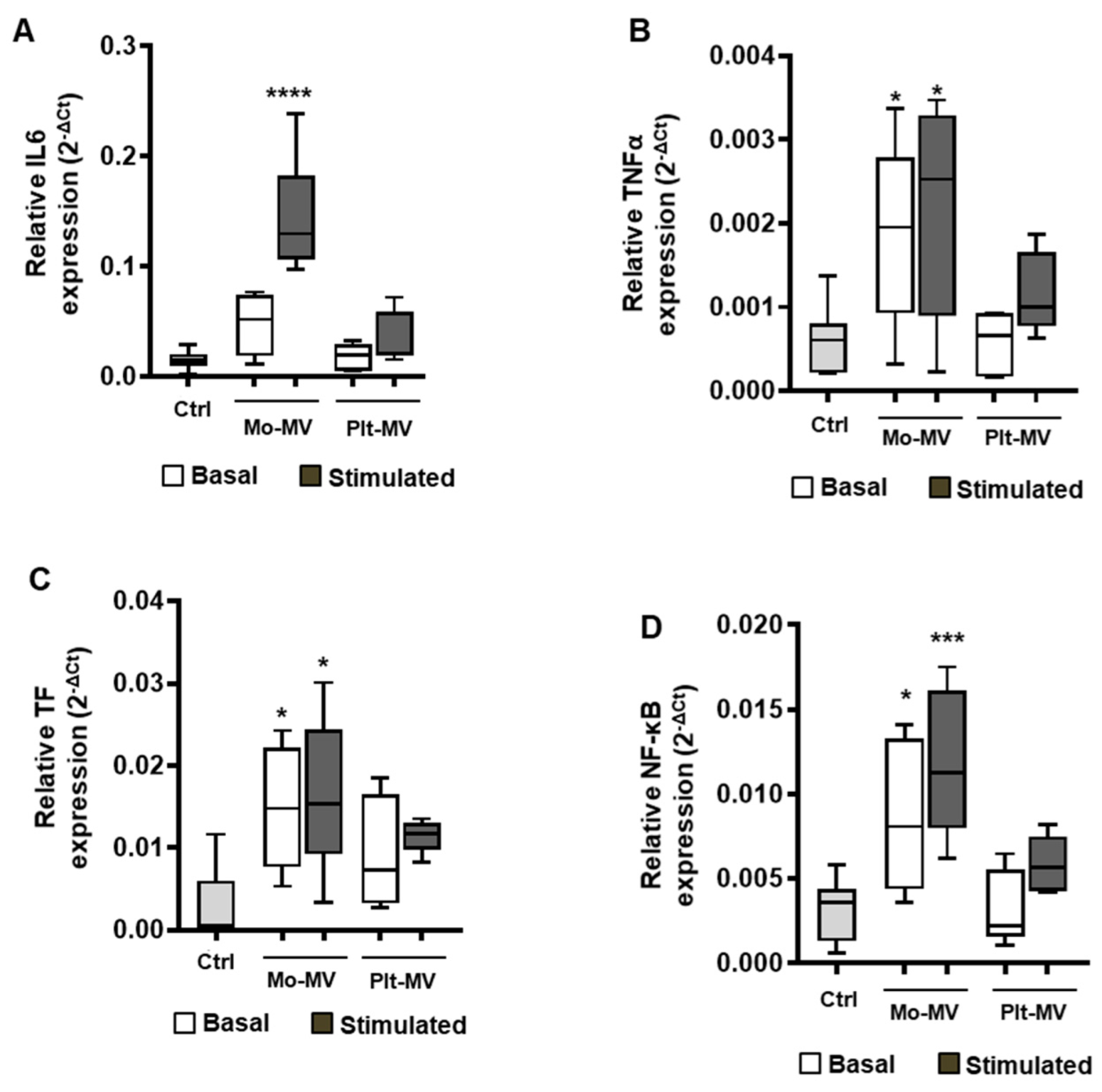
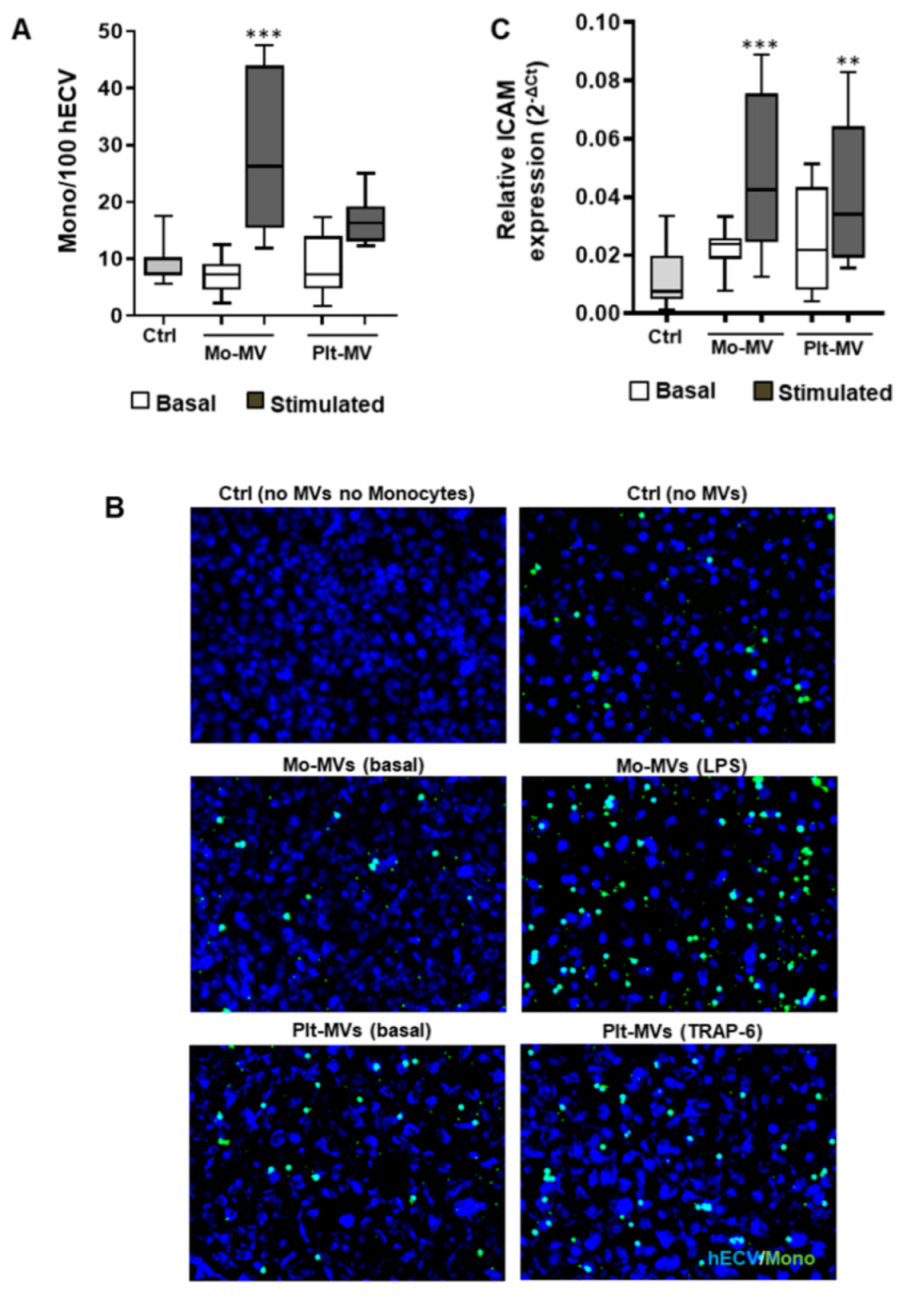
2.3. Monocyte-MVs More Than Platelet-MVs Induce the Expression of Inflammation Markers in Endothelial Cells
2.4. Both Monocyte- and Platelet-Derived MVs Favor Monocyte Adhesiveness to Endothelial Cells
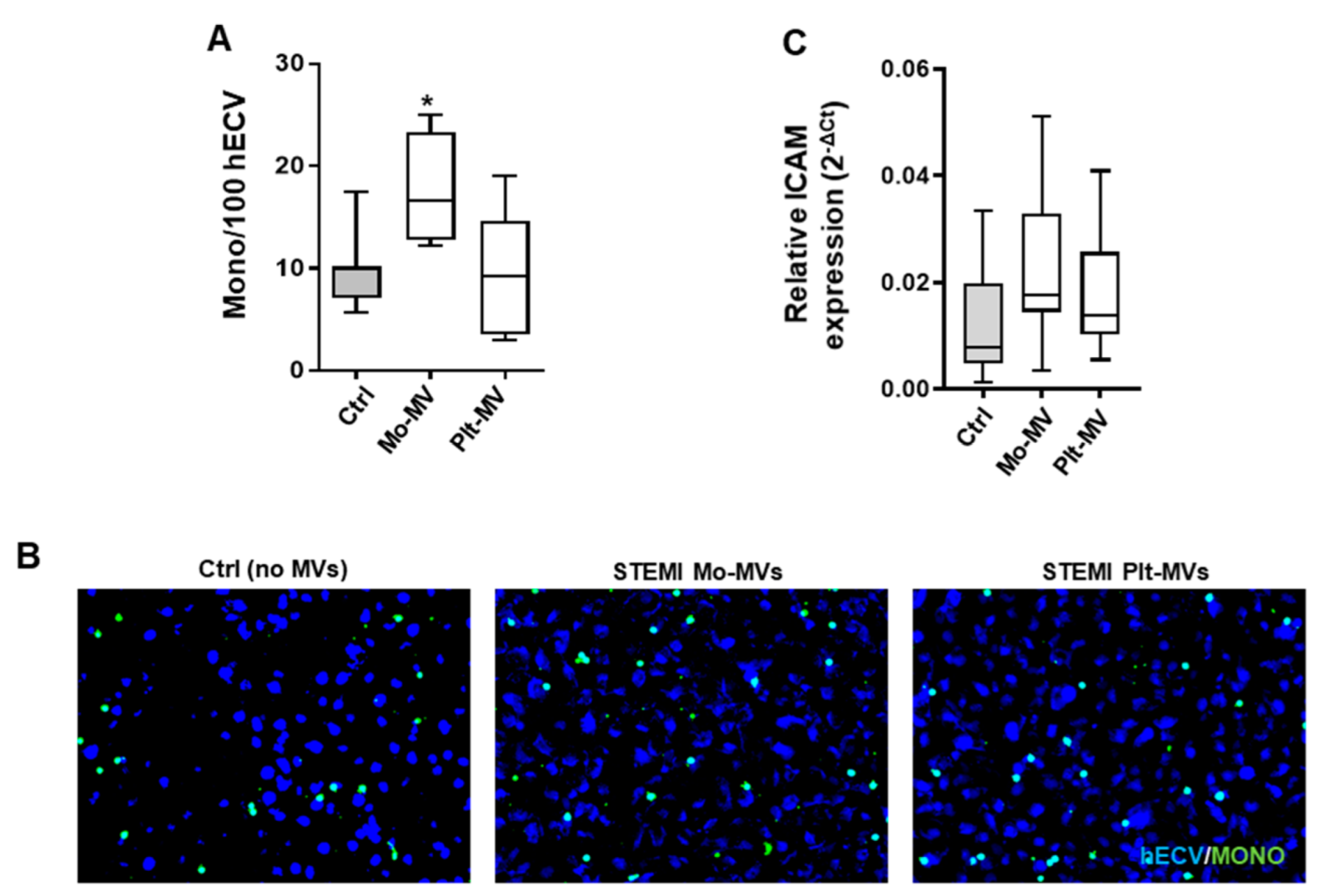
2.5. The Amount of MVs Spontaneously Released by Cells from STEMI Patients Is Comparable to That of Stimulated Cells from Healthy Subjects
2.6. STEMI Platelet-Derived MVs Induce Oxidative Stress and Inflammatory Phenotype on Endothelium While Monocyte-Derived MVs Affect Only Adhesion Properties
2.7. A Peculiar Surface Antigenic Profile Characterizes MVs Released from STEMI Cells
3. Discussion
4. Study Limitations
5. Materials and Methods
5.1. Patients and Healthy Subjects’ Enrollment
5.2. Cells Isolation
5.3. Microvesicles Production
5.4. Flow Cytometry
5.5. Cell Culture
5.6. Quantitative Real-Time PCR
5.7. Superoxide Anion (O2−) Production
5.8. Adhesion Assay
5.9. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Berumen Sanchez, G.; Bunn, K.E.; Pua, H.H.; Rafat, M. Extracellular vesicles: Mediators of intercellular communication in tissue injury and disease. Cell Commun. Signal. 2021, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Amabile, N.; Cheng, S.; Renard, J.M.; Larson, M.G.; Ghorbani, A.; McCabe, E.; Griffin, G.; Guerin, C.; Ho, J.E.; Shaw, S.Y.; et al. Association of circulating endothelial microparticles with cardiometabolic risk factors in the Framingham Heart Study. Eur. Heart J. 2014, 35, 2972–2979. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, F.; Darmon, P.; Hugel, B.; Combes, V.; Sanmarco, M.; Velut, J.G.; Arnoux, D.; Charpiot, P.; Freyssinet, J.M.; Oliver, C.; et al. Type 1 and type 2 diabetic patients display different patterns of cellular microparticles. Diabetes 2002, 51, 2840–2845. [Google Scholar] [CrossRef] [Green Version]
- Koga, H.; Sugiyama, S.; Kugiyama, K.; Fukushima, H.; Watanabe, K.; Sakamoto, T.; Yoshimura, M.; Jinnouchi, H.; Ogawa, H. Elevated levels of remnant lipoproteins are associated with plasma platelet microparticles in patients with type-2 diabetes mellitus without obstructive coronary artery disease. Eur. Heart J. 2006, 27, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, R.A.; Jy, W.; Jimenez, J.J.; Mauro, L.M.; Horstman, L.L.; Valle, M.; Aime, G.; Ahn, Y.S. Effects of severe hypertension on endothelial and platelet microparticles. Hypertension 2003, 41, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, C.; Gudi, K.; Krause, A.; Sackrowitz, R.; Harvey, B.G.; Strulovici-Barel, Y.; Mezey, J.G.; Crystal, R.G. Circulating endothelial microparticles as a measure of early lung destruction in cigarette smokers. Am. J. Respir. Crit. Care Med. 2011, 184, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Baron, M.; Boulanger, C.M.; Staels, B.; Tailleux, A. Cell-derived microparticles in atherosclerosis: Biomarkers and targets for pharmacological modulation? J. Cell Mol. Med. 2012, 16, 1365–1376. [Google Scholar] [CrossRef]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA expression in circulating microvesicles predicts cardiovascular events in patients with coronary artery disease. J. Am. Heart Assoc. 2014, 3, e001249. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, G.; Liu, M.L. Microvesicles as Emerging Biomarkers and Therapeutic Targets in Cardiometabolic Diseases. Genom. Proteom. Bioinform. 2018, 16, 50–62. [Google Scholar] [CrossRef]
- Camera, M.; Brambilla, M.; Canzano, P.; Cavallotti, L.; Parolari, A.; Tedesco, C.C.; Zara, C.; Rossetti, L.; Tremoli, E. Association of Microvesicles With Graft Patency in Patients Undergoing CABG Surgery. J. Am. Coll. Cardiol. 2020, 75, 2819–2832. [Google Scholar] [CrossRef]
- Diamant, M.; Tushuizen, M.E.; Sturk, A.; Nieuwland, R. Cellular microparticles: New players in the field of vascular disease? Eur. J. Clin. Investig. 2004, 34, 392–401. [Google Scholar] [CrossRef]
- Vajen, T.; Mause, S.F.; Koenen, R.R. Microvesicles from platelets: Novel drivers of vascular inflammation. Thromb. Haemost. 2015, 114, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Mezouar, S.; Mege, D.; Darbousset, R.; Farge, D.; Debourdeau, P.; Dignat-George, F.; Panicot-Dubois, L.; Dubois, C. Involvement of platelet-derived microparticles in tumor progression and thrombosis. Semin. Oncol. 2014, 41, 346–358. [Google Scholar] [CrossRef]
- Cloutier, N.; Tan, S.; Boudreau, L.H.; Cramb, C.; Subbaiah, R.; Lahey, L.; Albert, A.; Shnayder, R.; Gobezie, R.; Nigrovic, P.A.; et al. The exposure of autoantigens by microparticles underlies the formation of potent inflammatory components: The microparticle-associated immune complexes. EMBO Mol. Med. 2013, 5, 235–249. [Google Scholar] [CrossRef]
- Brill, A.; Dashevsky, O.; Rivo, J.; Gozal, Y.; Varon, D. Platelet-derived microparticles induce angiogenesis and stimulate post-ischemic revascularization. Cardiovasc. Res. 2005, 67, 30–38. [Google Scholar] [CrossRef]
- Agouni, A.; Lagrue-Lak-Hal, A.H.; Ducluzeau, P.H.; Mostefai, H.A.; Draunet-Busson, C.; Leftheriotis, G.; Heymes, C.; Martinez, M.C.; Andriantsitohaina, R. Endothelial dysfunction caused by circulating microparticles from patients with metabolic syndrome. Am. J. Pathol. 2008, 173, 1210–1219. [Google Scholar] [CrossRef] [Green Version]
- Barry, O.P.; Pratico, D.; Savani, R.C.; FitzGerald, G.A. Modulation of monocyte-endothelial cell interactions by platelet microparticles. J. Clin. Investig. 1998, 102, 136–144. [Google Scholar] [CrossRef]
- Del Conde, I.; Shrimpton, C.N.; Thiagarajan, P.; Lopez, J.A. Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood 2005, 106, 1604–1611. [Google Scholar] [CrossRef]
- Li, M.; Yu, D.; Williams, K.J.; Liu, M.L. Tobacco smoke induces the generation of procoagulant microvesicles from human monocytes/macrophages. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1818–1824. [Google Scholar] [CrossRef]
- Mayr, M.; Grainger, D.; Mayr, U.; Leroyer, A.S.; Leseche, G.; Sidibe, A.; Herbin, O.; Yin, X.; Gomes, A.; Madhu, B.; et al. Proteomics, metabolomics, and immunomics on microparticles derived from human atherosclerotic plaques. Circ. Cardiovasc. Genet. 2009, 2, 379–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardelli, C.; Amoruso, A.; Federici Canova, D.; Fresu, L.; Balbo, P.; Neri, T.; Celi, A.; Brunelleschi, S. Autocrine activation of human monocyte/macrophages by monocyte-derived microparticles and modulation by PPARgamma ligands. Br. J. Pharmacol. 2012, 165, 716–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerri, C.; Chimenti, D.; Conti, I.; Neri, T.; Paggiaro, P.; Celi, A. Monocyte/macrophage-derived microparticles up-regulate inflammatory mediator synthesis by human airway epithelial cells. J. Immunol. 2006, 177, 1975–1980. [Google Scholar] [CrossRef] [Green Version]
- Rautou, P.E.; Leroyer, A.S.; Ramkhelawon, B.; Devue, C.; Duflaut, D.; Vion, A.C.; Nalbone, G.; Castier, Y.; Leseche, G.; Lehoux, S.; et al. Microparticles from human atherosclerotic plaques promote endothelial ICAM-1-dependent monocyte adhesion and transendothelial migration. Circ. Res. 2011, 108, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.G.; Williams, J.C.; Davis, B.K.; Jacobson, K.; Doerschuk, C.M.; Ting, J.P.; Mackman, N. Monocytic microparticles activate endothelial cells in an IL-1beta-dependent manner. Blood 2011, 118, 2366–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N.G.; Smith, C.W.; Entman, M.L. The inflammatory response in myocardial infarction. Cardiovasc. Res. 2002, 53, 31–47. [Google Scholar] [CrossRef]
- Mulvihill, N.T.; Foley, J.B. Inflammation in acute coronary syndromes. Heart 2002, 87, 201–204. [Google Scholar] [CrossRef]
- Canzano, P.; Brambilla, M.; Porro, B.; Cosentino, N.; Tortorici, E.; Vicini, S.; Poggio, P.; Cascella, A.; Pengo, M.F.; Veglia, F.; et al. Platelet and Endothelial Activation as Potential Mechanisms Behind the Thrombotic Complications of COVID-19 Patients. JACC Basic Transl. Sci. 2021, 6, 202–218. [Google Scholar] [CrossRef]
- Brambilla, M.; Camera, M.; Colnago, D.; Marenzi, G.; De Metrio, M.; Giesen, P.L.; Balduini, A.; Veglia, F.; Gertow, K.; Biglioli, P.; et al. Tissue factor in patients with acute coronary syndromes: Expression in platelets, leukocytes, and platelet-leukocyte aggregates. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Packard, R.R.; Libby, P. Inflammation in atherosclerosis: From vascular biology to biomarker discovery and risk prediction. Clin. Chem. 2008, 54, 24–38. [Google Scholar] [CrossRef] [Green Version]
- Kavsak, P.A.; Ko, D.T.; Newman, A.M.; Palomaki, G.E.; Lustig, V.; MacRae, A.R.; Jaffe, A.S. Risk stratification for heart failure and death in an acute coronary syndrome population using inflammatory cytokines and N-terminal pro-brain natriuretic peptide. Clin. Chem. 2007, 53, 2112–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartford, M.; Wiklund, O.; Mattsson Hulten, L.; Persson, A.; Karlsson, T.; Herlitz, J.; Caidahl, K. C-reactive protein, interleukin-6, secretory phospholipase A2 group IIA and intercellular adhesion molecule-1 in the prediction of late outcome events after acute coronary syndromes. J. Intern. Med. 2007, 262, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Velasco, M.; Gonzalez-Ramos, S.; Bosca, L. Involvement of monocytes/macrophages as key factors in the development and progression of cardiovascular diseases. Biochem. J. 2014, 458, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Gerhardt, T.; Ley, K. Monocyte trafficking across the vessel wall. Cardiovasc. Res. 2015, 107, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Thakur, M.; Mishra, M.; Yadav, M.; Vibhuti, R.; Menon, A.M.; Nagda, G.; Dwivedi, V.P.; Dakal, T.C.; Yadav, V. Gene regulation of intracellular adhesion molecule-1 (ICAM-1): A molecule with multiple functions. Immunol. Lett. 2021, 240, 123–136. [Google Scholar] [CrossRef]
- Kratofil, R.M.; Kubes, P.; Deniset, J.F. Monocyte Conversion During Inflammation and Injury. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Slomka, A.; Urban, S.K.; Lukacs-Kornek, V.; Zekanowska, E.; Kornek, M. Large Extracellular Vesicles: Have We Found the Holy Grail of Inflammation? Front. Immunol. 2018, 9, 2723. [Google Scholar] [CrossRef]
- Dalli, J.; Montero-Melendez, T.; Norling, L.V.; Yin, X.; Hinds, C.; Haskard, D.; Mayr, M.; Perretti, M. Heterogeneity in neutrophil microparticles reveals distinct proteome and functional properties. Mol. Cell. Proteom. 2013, 12, 2205–2219. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Norling, L.V.; Montero-Melendez, T.; Federici Canova, D.; Lashin, H.; Pavlov, A.M.; Sukhorukov, G.B.; Hinds, C.J.; Perretti, M. Microparticle alpha-2-macroglobulin enhances pro-resolving responses and promotes survival in sepsis. EMBO Mol. Med. 2014, 6, 27–42. [Google Scholar] [CrossRef]
- Headland, S.E.; Norling, L.V. The resolution of inflammation: Principles and challenges. Semin. Immunol. 2015, 27, 149–160. [Google Scholar] [CrossRef]
- Khandagale, A.; Lindahl, B.; Lind, S.B.; Shevchenko, G.; Siegbahn, A.; Christersson, C. Plasma-derived extracellular vesicles from myocardial infarction patients inhibits tumor necrosis factor-alpha induced cardiac cell death. Curr. Res. Transl. Med. 2021, 70, 103323. [Google Scholar] [CrossRef] [PubMed]
- Oggero, S.; de Gaetano, M.; Marcone, S.; Fitzsimons, S.; Pinto, A.L.; Ikramova, D.; Barry, M.; Burke, D.; Montero-Melendez, T.; Cooper, D.; et al. Extracellular vesicles from monocyte/platelet aggregates modulate human atherosclerotic plaque reactivity. J. Extracell. Vesicles 2021, 10, 12084. [Google Scholar] [CrossRef] [PubMed]
- Stepien, E.; Stankiewicz, E.; Zalewski, J.; Godlewski, J.; Zmudka, K.; Wybranska, I. Number of microparticles generated during acute myocardial infarction and stable angina correlates with platelet activation. Arch. Med. Res. 2012, 43, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Suades, R.; Fuentes, E.; Palomo, I.; Padro, T. Role of Platelet-Derived Microvesicles As Crosstalk Mediators in Atherothrombosis and Future Pharmacology Targets: A Link between Inflammation, Atherosclerosis, and Thrombosis. Front. Pharmacol. 2016, 7, 293. [Google Scholar] [CrossRef] [Green Version]
- Terrisse, A.D.; Puech, N.; Allart, S.; Gourdy, P.; Xuereb, J.M.; Payrastre, B.; Sie, P. Internalization of microparticles by endothelial cells promotes platelet/endothelial cell interaction under flow. J. Thromb. Haemost. 2010, 8, 2810–2819. [Google Scholar] [CrossRef]
- Mause, S.F.; von Hundelshausen, P.; Zernecke, A.; Koenen, R.R.; Weber, C. Platelet microparticles: A transcellular delivery system for RANTES promoting monocyte recruitment on endothelium. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1512–1518. [Google Scholar] [CrossRef] [Green Version]
- Gasa, N.; Meiring, M. Microparticles: A link to increased thrombin generation. Blood Coagul. Fibrinolysis 2021, 32, 204–208. [Google Scholar] [CrossRef]
- Andrikopoulos, P.; Kieswich, J.; Harwood, S.M.; Baba, A.; Matsuda, T.; Barbeau, O.; Jones, K.; Eccles, S.A.; Yaqoob, M.M. Endothelial Angiogenesis and Barrier Function in Response to Thrombin Require Ca2+ Influx through the Na+/Ca2+ Exchanger. J. Biol. Chem. 2015, 290, 18412–18428. [Google Scholar] [CrossRef] [Green Version]
- Suades, R.; Padro, T.; Crespo, J.; Ramaiola, I.; Martin-Yuste, V.; Sabate, M.; Sans-Rosello, J.; Sionis, A.; Badimon, L. Circulating microparticle signature in coronary and peripheral blood of ST elevation myocardial infarction patients in relation to pain-to-PCI elapsed time. Int. J. Cardiol. 2016, 202, 378–387. [Google Scholar] [CrossRef]
- Serebruany, V.L.; Malinin, A.I.; Ziai, W.; Pokov, A.N.; Bhatt, D.L.; Alberts, M.J.; Hanley, D.F. Effects of clopidogrel and aspirin in combination versus aspirin alone on platelet activati.ion and major receptor expression in patients after recent ischemic stroke: For the Plavix Use for Treatment of Stroke (PLUTO-Stroke) trial. Stroke 2005, 36, 2289–2292. [Google Scholar] [CrossRef] [Green Version]
- Taus, F.; Meneguzzi, A.; Castelli, M.; Minuz, P. Platelet-Derived Extracellular Vesicles as Target of Antiplatelet Agents. What Is the Evidence? Front. Pharmacol. 2019, 10, 1256. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Shan, X.F.; Wang, X.L.; Dong, W.W.; Li, Z.; Liu, X.H.; Zhang, W.; Xing, K.; Chang, F.J. Endothelial damage effects of circulating microparticles from patients with stable angina are reduced by aspirin through ERK/p38 MAPKs pathways. Cardiovasc. Ther. 2017, 35, e12273. [Google Scholar] [CrossRef]
- Franca, C.N.; Pinheiro, L.F.; Izar, M.C.; Brunialti, M.K.; Salomao, R.; Bianco, H.T.; Kasmas, S.H.; Barbosa, S.P.; de Nucci, G.; Fonseca, F.A. Endothelial progenitor cell mobilization and platelet microparticle release are influenced by clopidogrel plasma levels in stable coronary artery disease. Circ. J. 2012, 76, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Jastrzebska, M.; Marcinowska, Z.; Oledzki, S.; Chelstowski, K.; Siennicka, A.; Klysz, M.; Clark, J.S. Variable gender-dependent platelet responses to combined antiplatelet therapy in patients with stable coronary-artery disease. J. Physiol. Pharmacol. 2018, 69, 595–605. [Google Scholar] [CrossRef]
- Lubsczyk, B.; Kollars, M.; Hron, G.; Kyrle, P.A.; Weltermann, A.; Gartner, V. Low dose acetylsalicylic acid and shedding of microparticles in vivo in humans. Eur. J. Clin. Investig. 2010, 40, 477–482. [Google Scholar] [CrossRef]
- Bulut, D.; Becker, V.; Mugge, A. Acetylsalicylate reduces endothelial and platelet-derived microparticles in patients with coronary artery disease. Can. J. Physiol. Pharmacol. 2011, 89, 239–244. [Google Scholar] [CrossRef]
- Talmon, M.; Rossi, S.; Pastore, A.; Cattaneo, C.I.; Brunelleschi, S.; Fresu, L.G. Vortioxetine exerts anti-inflammatory and immunomodulatory effects on human monocytes/macrophages. Br. J. Pharmacol. 2018, 175, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Signorelli, S.S.; Conti, G.O.; Fiore, M.; Elfio, M.G.; Cristaldi, A.; Nicolosi, I.; Zuccarello, P.; Zanoli, L.; Gaudio, A.; Di Raimondo, D.; et al. Inter-Relationship between Platelet-Derived Microparticles and Oxidative Stress in Patients with Venous Thromboembolism. Antioxidants 2020, 9, 1217. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STEMI Patients (n = 7) | |
|---|---|
| Men, n (%) | 3 (43) |
| Age, years | 63 ± 13 |
| Hypertension, n (%) | 6 (86) |
| Diabetes, n (%) | 3 (43) |
| Dyslipidemia, n (%) | 5 (71) |
| Pharmacological therapy | |
| Aspirin, n (%) | 7 (100) |
| P2Y12 inhibitors, n (%) | 7 (100) |
| Calcium antagonists, n (%) | 2 (29) |
| ACE inhibitors, n (%) | 4 (57) |
| Statins, n (%) | 5 (71) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brambilla, M.; Talmon, M.; Canzano, P.; Fresu, L.G.; Brunelleschi, S.; Tremoli, E.; Camera, M. Different Contribution of Monocyte- and Platelet-Derived Microvesicles to Endothelial Behavior. Int. J. Mol. Sci. 2022, 23, 4811. https://doi.org/10.3390/ijms23094811
Brambilla M, Talmon M, Canzano P, Fresu LG, Brunelleschi S, Tremoli E, Camera M. Different Contribution of Monocyte- and Platelet-Derived Microvesicles to Endothelial Behavior. International Journal of Molecular Sciences. 2022; 23(9):4811. https://doi.org/10.3390/ijms23094811
Chicago/Turabian StyleBrambilla, Marta, Maria Talmon, Paola Canzano, Luigia G. Fresu, Sandra Brunelleschi, Elena Tremoli, and Marina Camera. 2022. "Different Contribution of Monocyte- and Platelet-Derived Microvesicles to Endothelial Behavior" International Journal of Molecular Sciences 23, no. 9: 4811. https://doi.org/10.3390/ijms23094811
APA StyleBrambilla, M., Talmon, M., Canzano, P., Fresu, L. G., Brunelleschi, S., Tremoli, E., & Camera, M. (2022). Different Contribution of Monocyte- and Platelet-Derived Microvesicles to Endothelial Behavior. International Journal of Molecular Sciences, 23(9), 4811. https://doi.org/10.3390/ijms23094811