
Molecular Cytogenetic and Physiological Characterization of a Novel Wheat-Rye T1RS.1BL Translocation Line from Secale cereal L. Weining with Resistance to Stripe Rust and Functional “Stay Green” Trait

 and
and
Abstract
:1. Introduction
2. Results
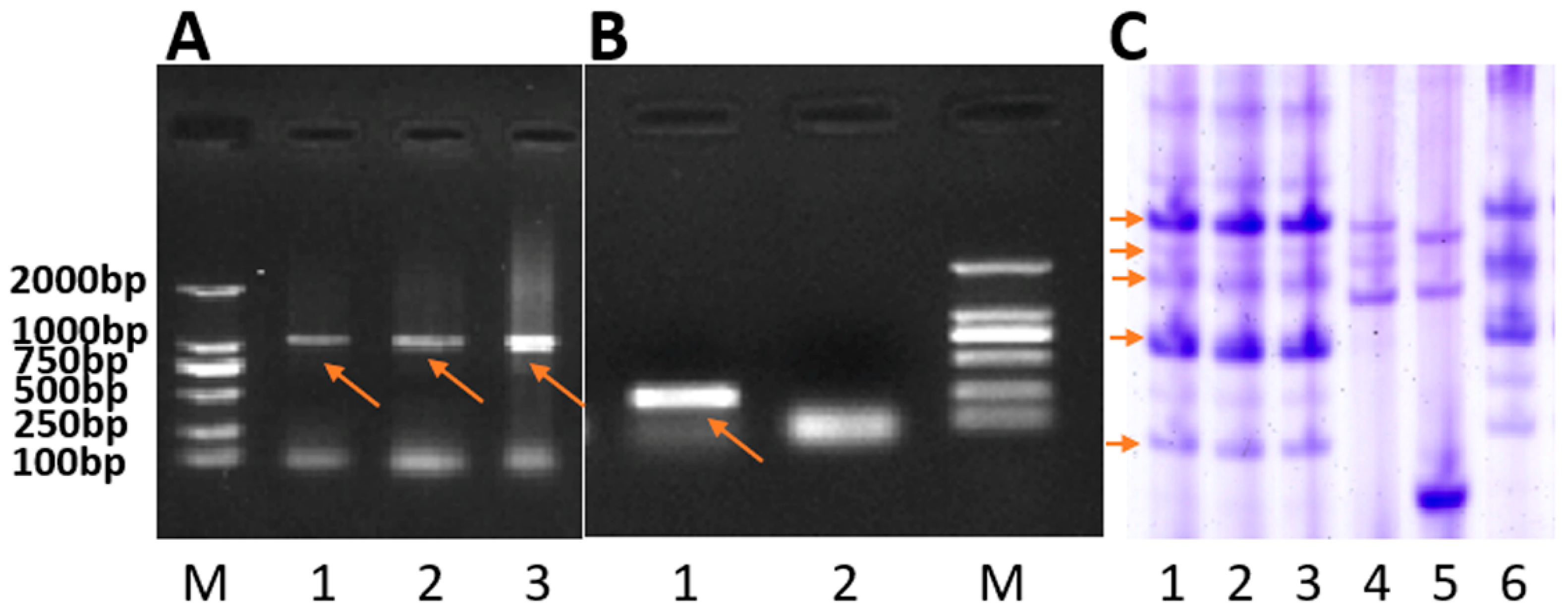
2.1. Development of the New T1RS.1BL Translocation Line RT843-5
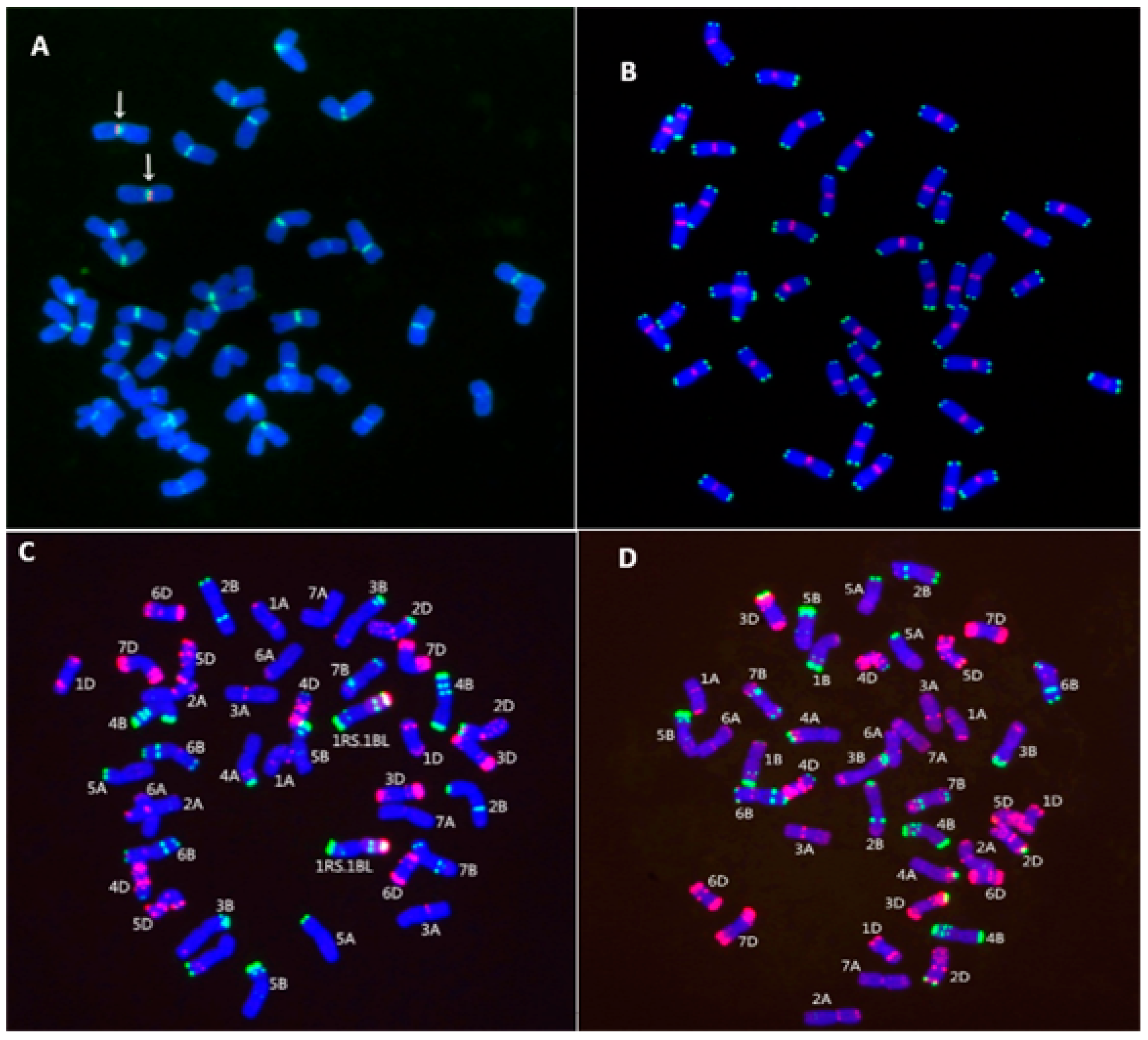
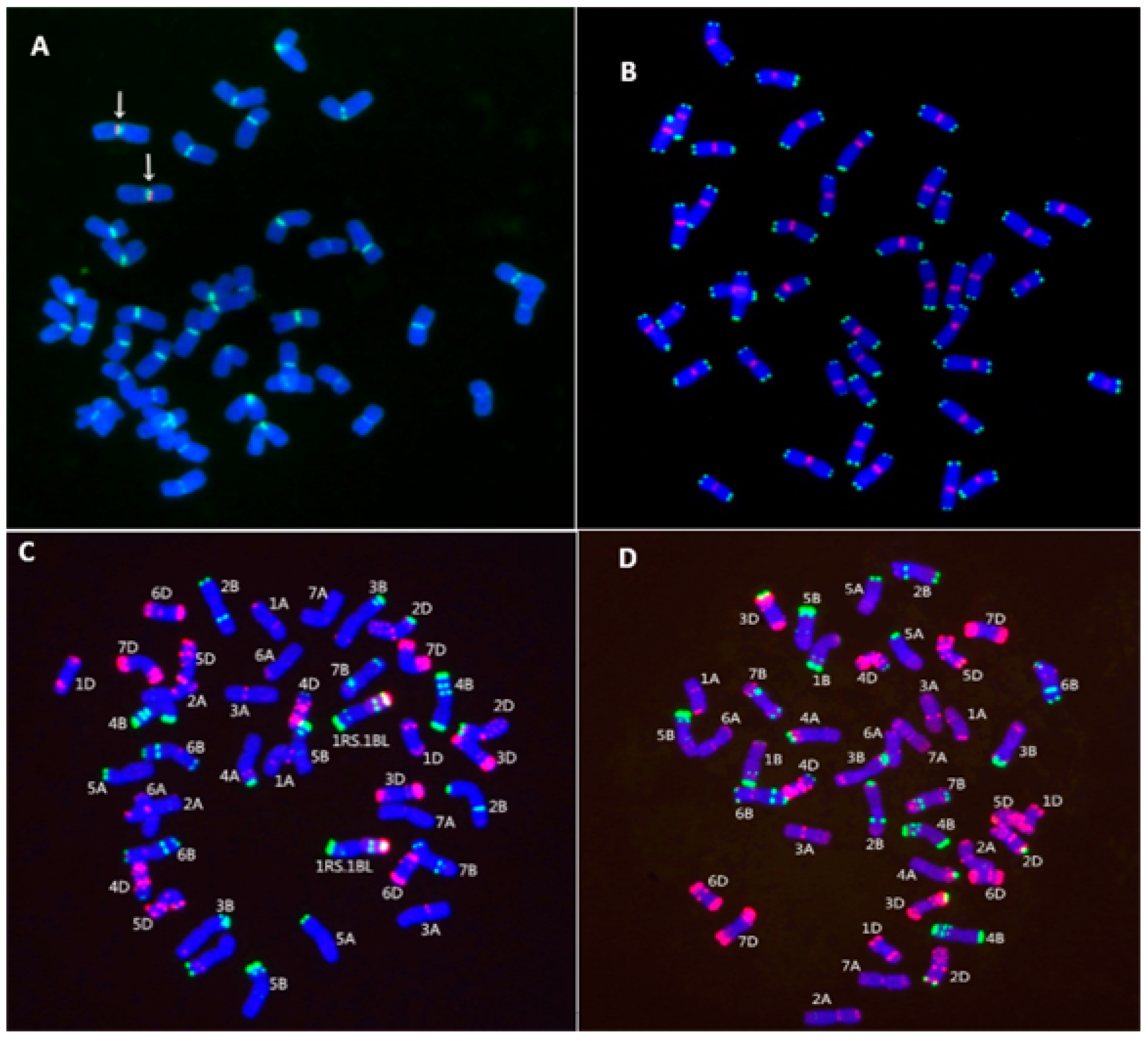
2.2. Chromosome Identification
2.3. Analysis of Resistance to Stripe Rust
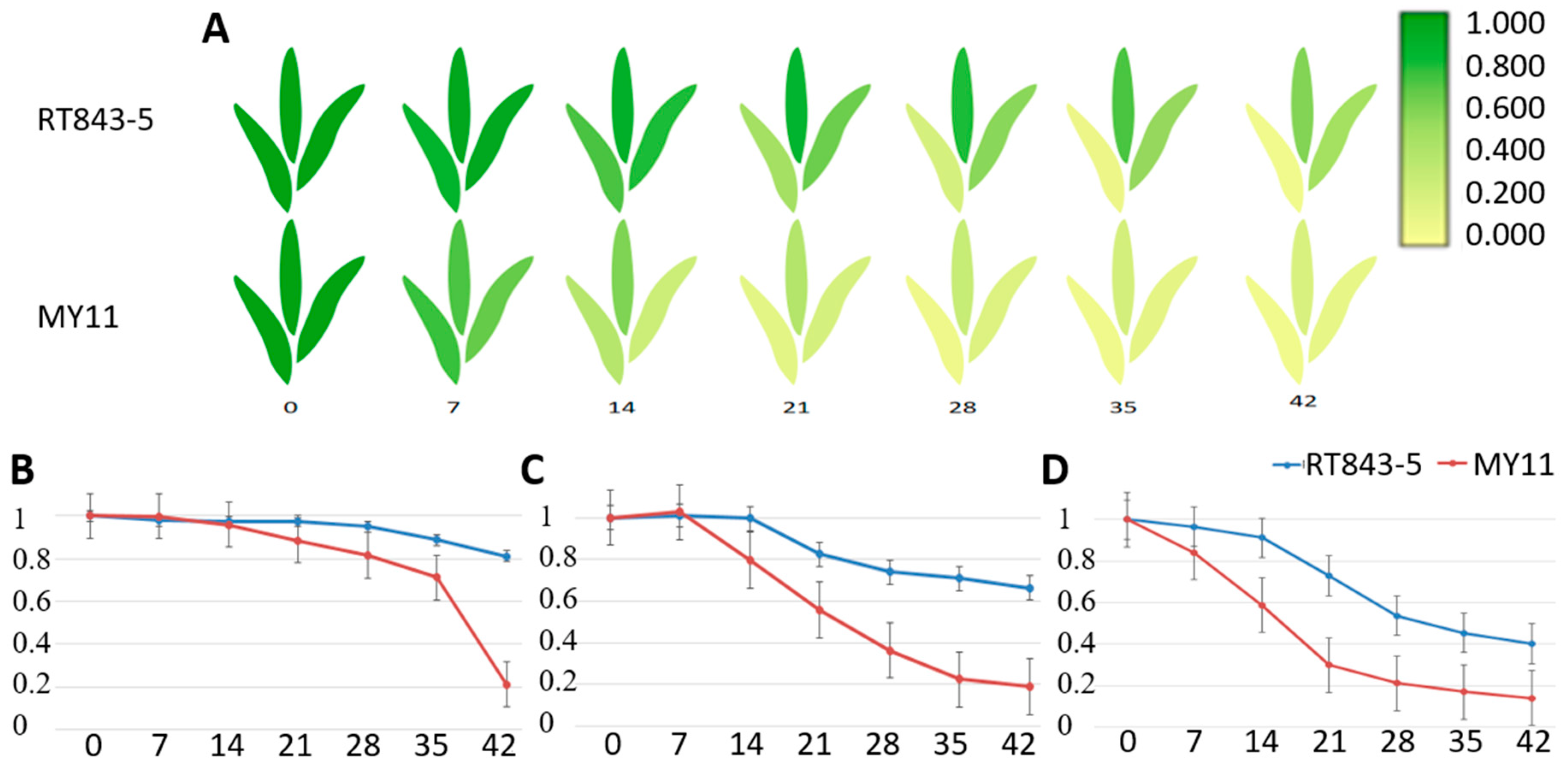
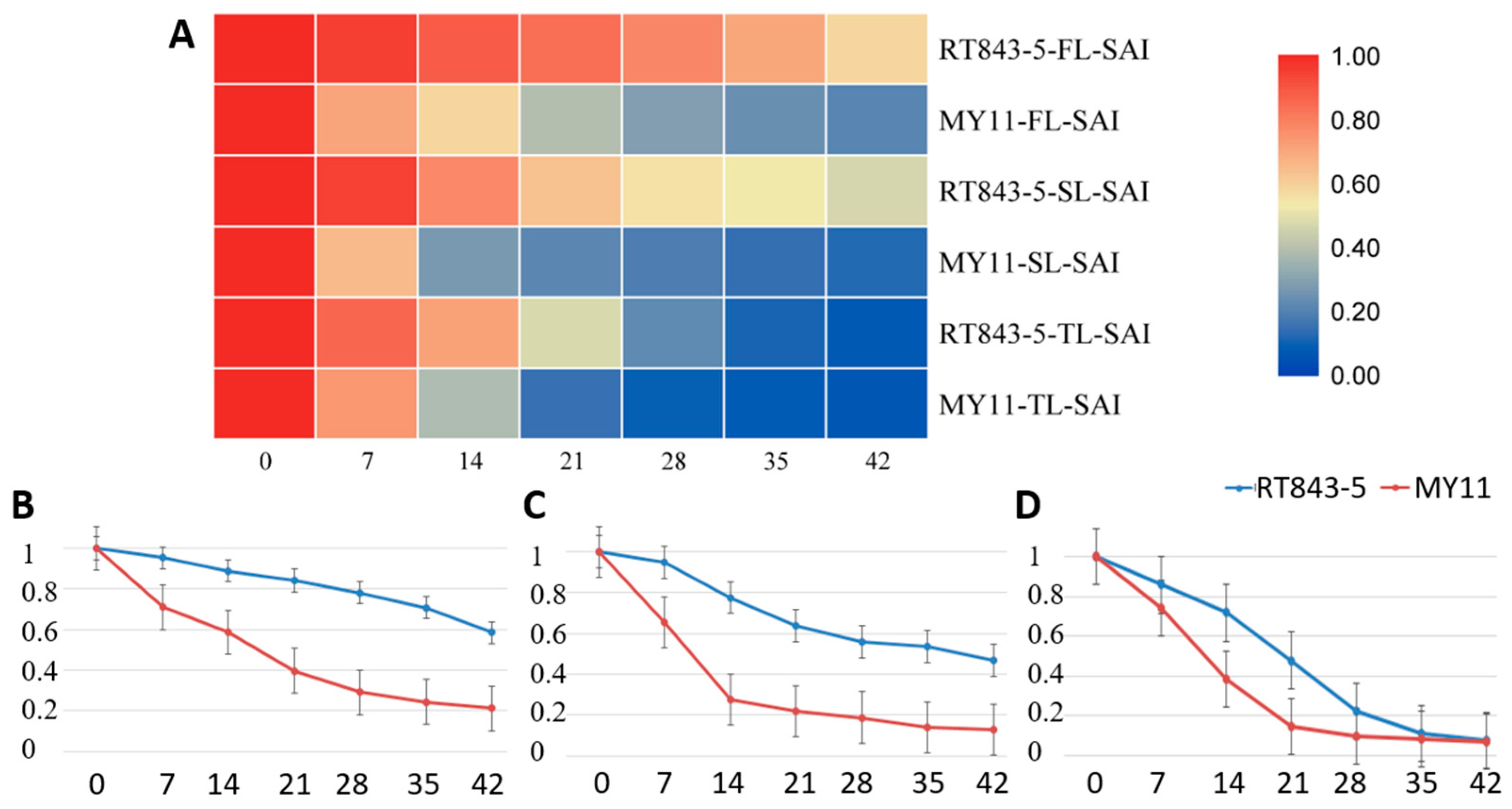
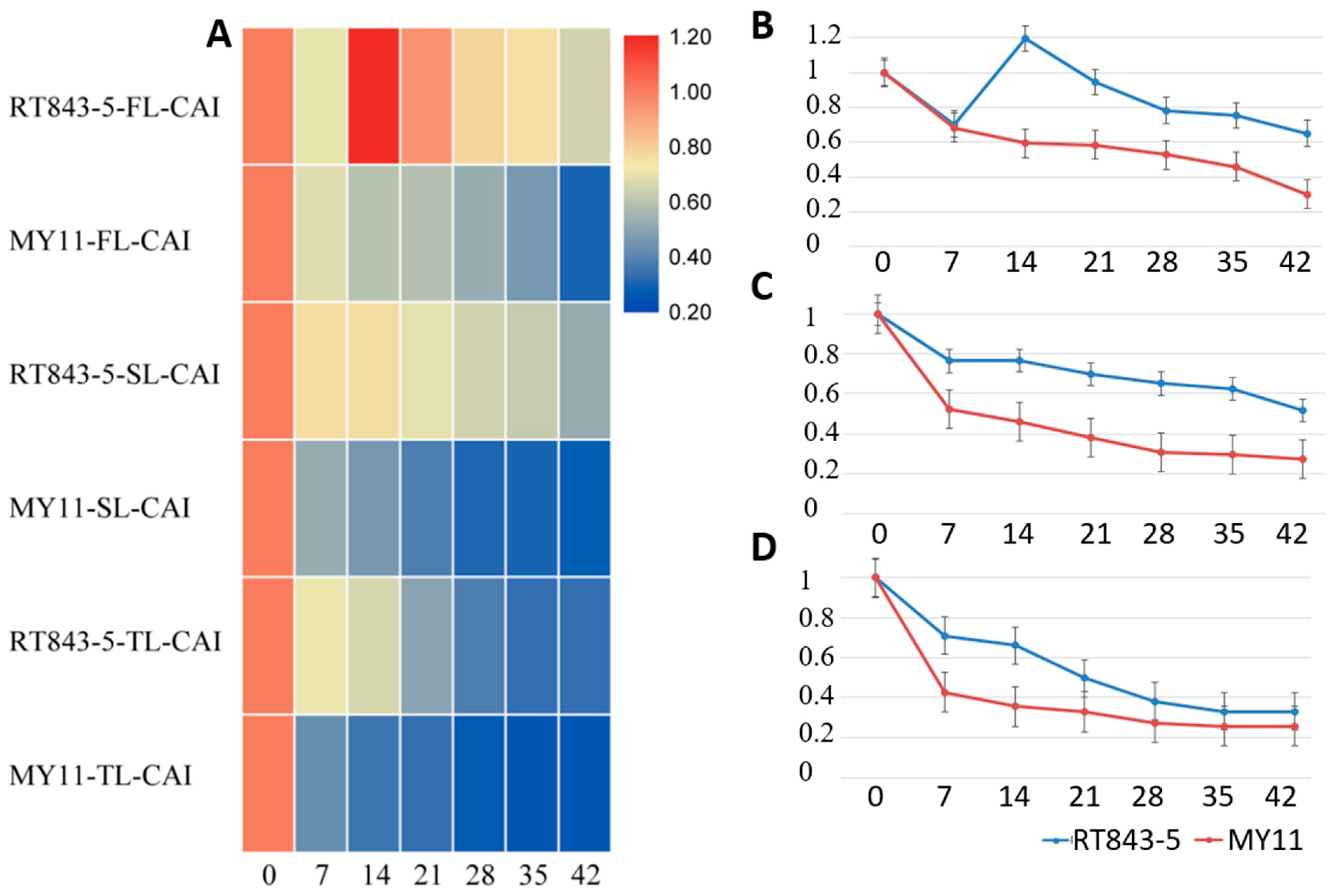
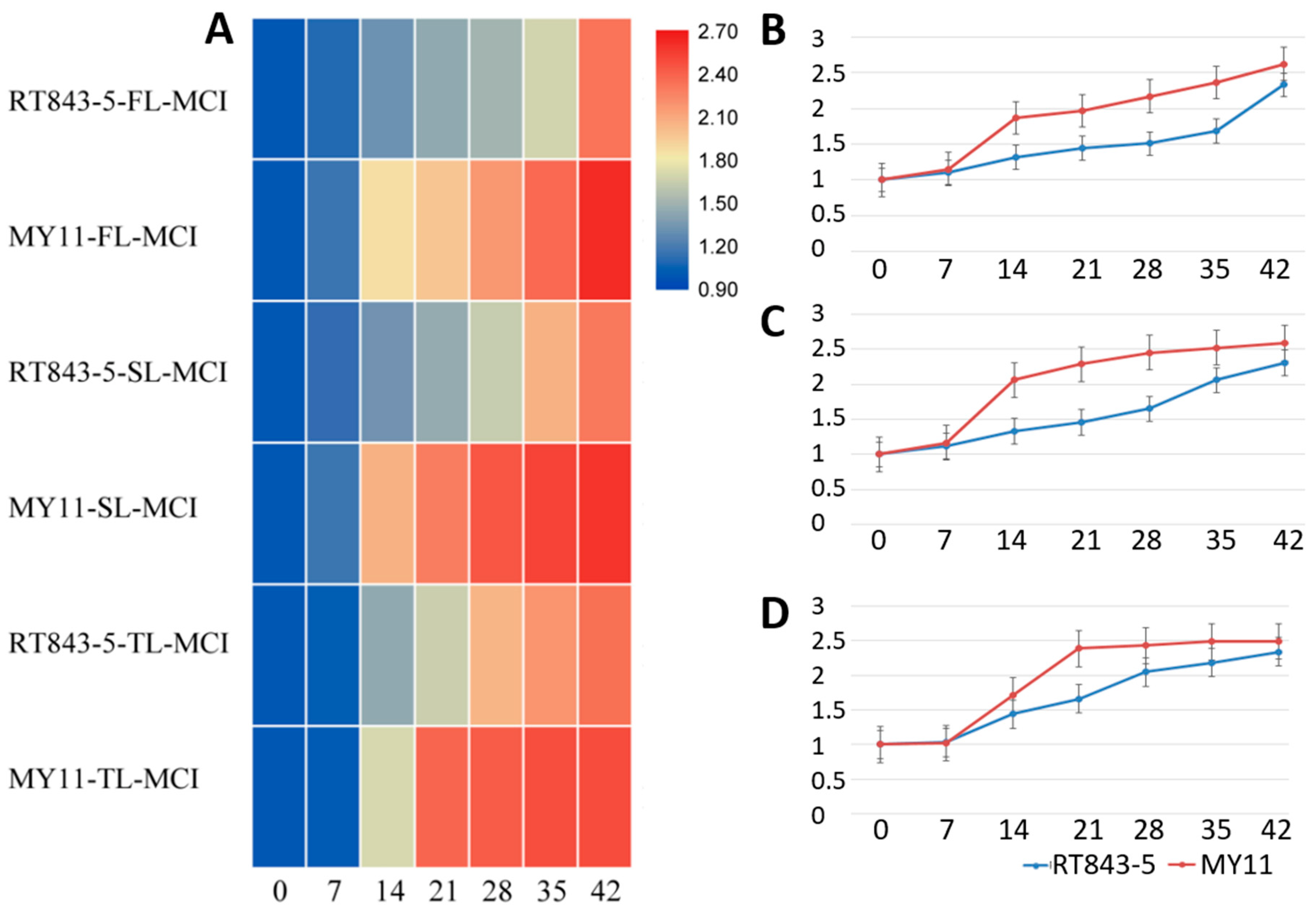
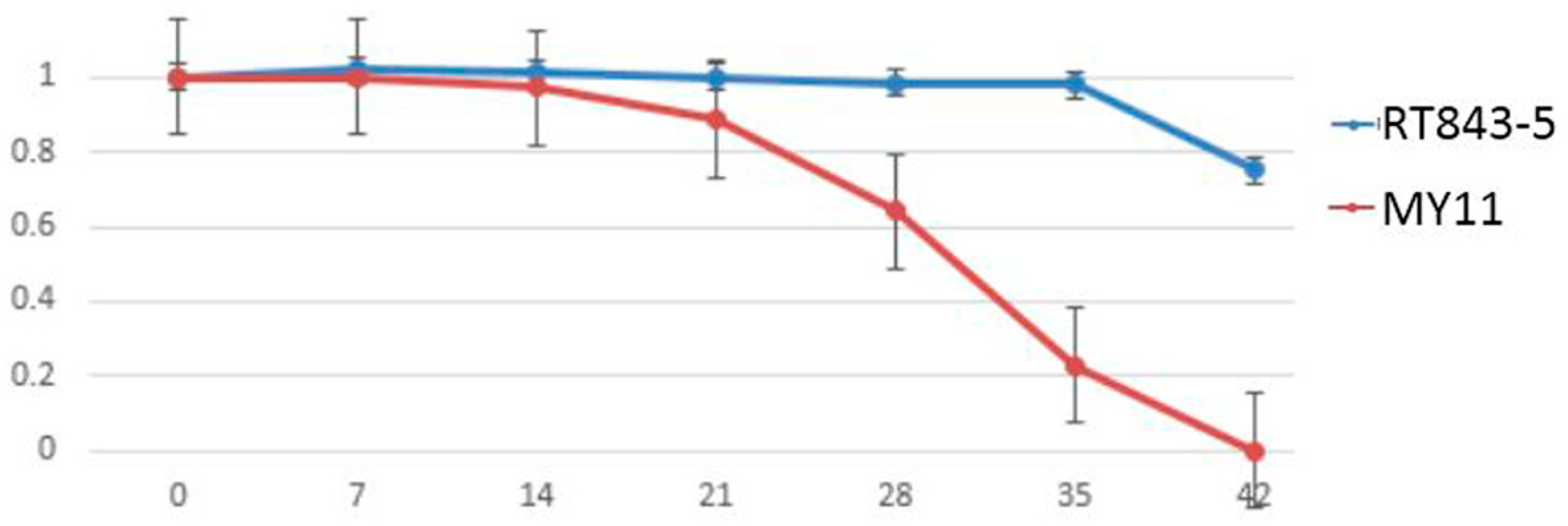
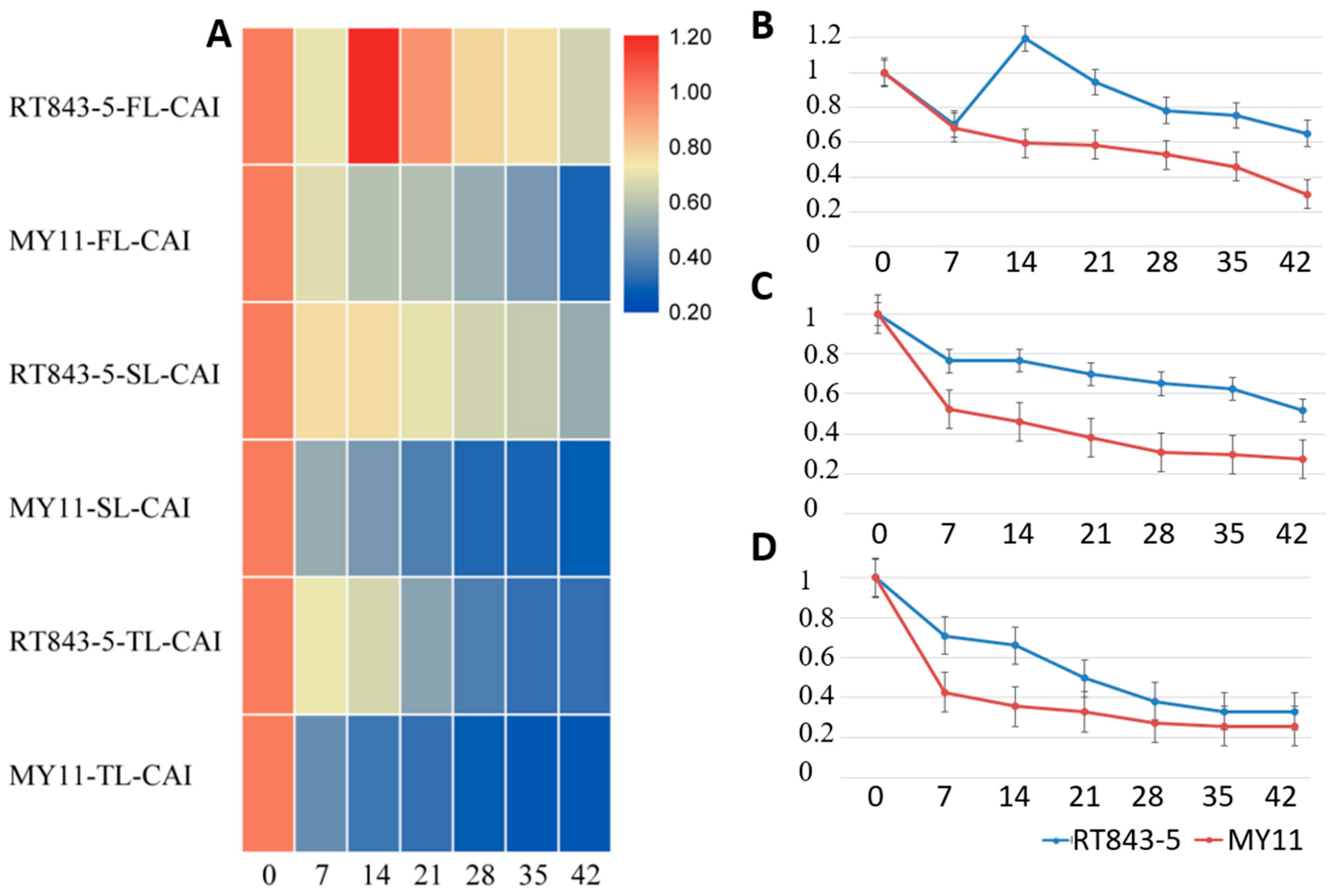
2.4. Measurement and Analysis of Functional Stay-Green Traits
2.5. Agronomic Traits of RT843-5
3. Discussion
3.1. New 1RS Chromosome Arm with a Resistance Gene to Pst Originating from Weining Rye
3.2. Functional Stay-Green Trait of RT843-5
4. Materials and Methods
4.1. Plant Materials
4.2. Cytogenetic and Molecular Analyses
4.3. Stripe Rust Tests
4.4. Physiological Measurement of the Stay-Green Trait
4.5. Field Experiments
4.6. Index Calculation and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ren, T.-H.; Chen, F.; Yan, B.-J.; Zhang, H.-Q.; Ren, Z.-L. Genetic diversity of wheat-rye 1BL.1RS translocation lines derived from different wheat and rye sources. Euphytica 2012, 183, 133–146. [Google Scholar] [CrossRef]
- Molnár-Láng, M.; Cseh, A.; Szakács, É.; Molnár, I. Development of a wheat genotype combining the recessive crossability alleles kr1kr1kr2kr2 and the 1BL.1RS translocation, for the rapid enrichment of 1RS with new allelic variation. Theor. Appl. Genet. 2010, 120, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chaudhary, H.K.; Kapoor, C.; Manoj, N.V.; Singh, K.; Sood, V.K. Molecular cytogenetic analysis of novel wheat-rye translocation lines and their characterization for drought tolerance and yellow rust resistance. Cereal Res. Comm. 2021, 1–11. [Google Scholar] [CrossRef]
- Liu, J.; Chang, Z.; Zhang, X.; Yang, Z.; Li, X.; Jia, J.; Zhan, H.; Guo, H.; Wang, J. Putative Thinopyrum intermedium-derived stripe rust resistance gene Yr50 maps on wheat chromosome arm 4BL. Theor. Appl. Genet. 2013, 126, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Jia, J.; Zhang, X.; Li, X.; Yang, Z.; Ma, J.; Guo, H.; Zhan, H.; Qiao, L.; Chang, Z. Molecular mapping of the stripe rust resistance gene Yr69 on wheat chromosome 2AS. Plant Dis. 2016, 100, 1717–1724. [Google Scholar] [CrossRef] [Green Version]
- Bayles, R.A.; Flath, K.; Mogens, S.H.; Vallavieillepope, C.D. Breakdown of the Yr17 resistance to yellow rust of wheat in northern Europe. Agronomie 2000, 20, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Marais, G.F.; Mccallum, B.; Snyman, J.E.; Pretorius, Z.A.; Marais, A.S. Leaf rust and stripe rust resistance genes Lr54 and Yr37 transferred to wheat from Aegilops kotschyi. Plant Breed. 2005, 124, 538–541. [Google Scholar] [CrossRef]
- Marais, G.F.; Badenhorst, P.E.; Eksteen, A.; Pretorius, Z.A. Reduction of Aegilops sharonensis chromatin associated with resistance genes Lr56 and Yr38 in wheat. Euphytica 2010, 171, 15–22. [Google Scholar] [CrossRef]
- Vasu, K.; Shilpa, S.; Bikram, S.G. Molecular genetic description of the cryptic wheat–Aegilops geniculata, introgression carrying rust resistance genes, Lr57, and, Yr40, using wheat ESTs and synteny with rice. Genome 2009, 52, 1025–1036. [Google Scholar]
- Cao, Z.; Deng, Z.; Wang, M.; Wang, X.; Jing, J.; Zhang, X. Inheritance and molecular mapping of an alien stripe-rust resistance gene from a wheat-Psathyrostachys huashanica translocation line. Plant Sci. 2008, 174, 544–549. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, J.; He, F.; Ma, H.; Wang, H. Molecular cytogenetic, identification of a wheat (Triticum aestivum)-American dune grass (Leymus mollis) translocation line resistant to stripe rust. Genet. Mol. Res. 2012, 11, 3198–3206. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, R.; Korzun, V. About the origin of 1RS.1BL wheat–rye chromosome translocations from Germany. Plant Breed. 1997, 116, 537–540. [Google Scholar] [CrossRef]
- Rabinovich, S.V. Importance of wheat–rye translocations for breeding modern cultivars of Triticum aestivum L. Euphytica 1998, 100, 323–340. [Google Scholar] [CrossRef]
- Ren, T.-H.; Yang, Z.-J.; Yan, B.-J.; Zhang, H.-Q.; Fu, S.-L.; Ren, Z.-L. Development and characterization of a new 1BL.1RS translocation line with resistance to stripe rust and powdery mildew of wheat. Euphytica 2009, 169, 207–213. [Google Scholar] [CrossRef]
- Mago, R.; Miah, H.; Lawrence, G.J.; Wellings, C.R.; Spielmeyer, W.; Bariana, H.S.; McIntosh, R.A.; Pryor, A.J.; Ellis, J.G. High-resolution mapping and mutation analysis separate the rust resistance genes Sr31, Lr26 and Yr9 on the short arm of rye chromosome 1. Theor. Appl. Genet. 2005, 112, 41–50. [Google Scholar] [CrossRef]
- Kumlay, A.M.; Baenziger, P.S.; Gill, K.S.; Shelton, D.R.; Graybosch, R.A.; Lukaszewski, A.J.; Wesenberg, D.M. Understanding the effect of rye chromatin in bread wheat. Crop Sci. 2003, 43, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Howell, T.; Hale, I.; Jankuloski, L.; Bonafede, M.; Gilbert, M.; Dubcovsky, J. Mapping a region within the 1RS.1BL translocation in common wheat affecting grain yield and canopy water status. Theor. Appl. Genet. 2014, 127, 2695–2709. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Tang, Z.; Fu, S.; Yan, B.; Tan, F.; Ren, Z.; Li, Z. Molecular cytogenetic characterization of novel wheat-rye T1RS.1BL translocation lines with high resistance to diseases and great agronomic traits. Front. Plant Sci. 2017, 8, 799. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.X.; Chen, X.M.; Line, R.F.; Leung, H.; Wellings, C.R. Development of resistance gene analog polymorphism markers for the Yr9 gene resistance to wheat stripe rust. Genome 2001, 44, 509–516. [Google Scholar] [CrossRef]
- Gong, W.; Han, R.; Ren, T.; Wang, C.; Yang, Z.; Yan, M.; Luo, P.; Liu, A.; Li, H.; Liu, C.; et al. Molecular detection of 1RS/1BL translocation and stripe rust resistance gene Yr41 in 1293 wheat cultivars (lines). Shandong Agri. Sci. 2020, 52, 1–6. [Google Scholar]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, H.; Ougham, H.J. The stay-green trait. J. Exp. Bot. 2014, 65, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Crasta, O.R.; Xu, W.W.; Rosenow, D.T.; Mullet, J.; Nguyen, H.T. Mapping of post-flowering drought resistance traits in grain sorghum: Association between QTLs influencing premature senescence and maturity. Mol. Gen. Genet. 1999, 262, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.-W.; Lee, Y.-J.; Koh, H.-J.; Lee, B.-M.; Nam, Y.-W.; Paek, N.-C. Isolation, characterization, and mapping of the stay green mutant in rice. Theor. Appl. Genet. 2002, 104, 526–532. [Google Scholar] [CrossRef]
- Jiang, G.-H.; He, Y.-Q.; Xu, C.-G.; Li, X.-H.; Zhang, Q. The genetic basis of stay-green in rice analyzed in a population of doubled haploid lines derived from an indica by japonica cross. Theor. Appl. Genet. 2004, 108, 688–698. [Google Scholar] [CrossRef]
- Luquez, V.M.; Guiamet, J.J. The stay green mutations d1 and d2 increase water stress susceptibility in soybeans. J. Exp. Bot. 2002, 53, 1421–1428. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, J.; Deng, D.; Ding, H.; Bian, Y.; Yin, Z.; Wu, Y.; Zhou, B.; Zhao, Y. A comprehensive meta-analysis of plant morphology, yield, stay-green, and virus disease resistance QTL in maize (Zea mays L.). Planta 2016, 243, 459–471. [Google Scholar] [CrossRef]
- Coupe, S.A.; Watson, L.M.; Ryan, D.J.; Pinkney, T.; Eason, J.R. Molecular analysis of programmed cell death during senescence in Arabidopsis thaliana and Brassica oleracea: Cloning broccoli LSD1, Bax inhibitor and serine palmitoyltransferase homologues. J. Exp. Bot. 2004, 55, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Brownhill, E.; Pourtau, N. Mechanisms of the light-dependent induction of cell death in tobacco plants with delayed senescence. J. Exp. Bot. 2005, 56, 2897–2905. [Google Scholar] [CrossRef]
- Huang, Y.-J.; To, K.-Y.; Yap, M.-N.; Chiang, W.-J.; Suen, D.-F.; Chen, S.-C. Cloning and characterization of leaf senescence up-regulated genes in sweet potato. Physiol. Plant. 2001, 113, 384–391. [Google Scholar] [CrossRef]
- Hafsi, M.; Mechmeche, W.; Bouamama, L.; Djekoune, A.; Zaharieva, M.; Monneveux, P. Flag leaf senescence, as evaluated by numerical image analysis, and its relationship with yield under drought in durum wheat. J. Agron. Crop Sci. 2000, 185, 275–280. [Google Scholar] [CrossRef]
- Spano, G.; Di Fonzo, N.; Perrotta, C.; Platani, C.; Ronga, G.; Lawlor, D.W.; Napier, J.A.; Shewry, P.R. Physiological characterization of ‘stay green’ mutants in durum wheat. J. Exp. Bot. 2003, 54, 1415–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liang, Y.; Hu, X.; Wang, X.; Tan, F.; Zhang, H.; Ren, Z.; Luo, P. Physiological characterization of ‘stay green’ wheat cultivars during grain filling stage under field growing conditions. Acta Physiol. Plant. 2010, 32, 875–882. [Google Scholar] [CrossRef]
- Wang, W.; Hao, Q.; Wang, W.; Li, Q.; Chen, F.; Ni, F.; Wang, Y.; Fu, D.; Wu, J.; Wang, W. The involvement of cytokinin and nitrogen metabolism in delayed flag leaf senescence in a wheat stay-green mutant, tasg1. Plant Sci. 2019, 278, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.K.; Kumari, M.; Singh, V.P.; Reddy, C.M.; Kumar, S.; Rane, J.; Chand, R. Stay green trait: Variation, inheritance and its association with spot blotch resistance in spring wheat (Triticum aestivum L.). Euphytica 2007, 153, 59–71. [Google Scholar] [CrossRef]
- Sun, P.; Wu, Q.; Xu, B.; Chang, S.; Miao, H.; Jing, Z. Progress in research on STAY-GREEN genes in plants. Plant Physiol. J. 2015, 51, 1017–1023. [Google Scholar]
- Li, G.; Wang, L.; Yang, J.; He, H.; Jin, H.; Li, X.; Ren, T.; Ren, Z.; Li, F.; Han, X.; et al. A high-quality genome assembly highlights rye genomic characteristics and agronomically important genes. Nat. Genet. 2021, 53, 574. [Google Scholar] [CrossRef]
- Ren, T.; Ren, Z.; Yang, M.; Yan, B.; Tan, F.; Fu, S.; Tang, Z.; Li, Z. Novel source of 1RS from Baili rye conferred high resistance to diseases and enhanced yield traits to common wheat. Mol. Breed. 2018, 38, 101. [Google Scholar] [CrossRef]
- Anderson, G.R.; Papa, D.; Peng, J.; Tahir, M.; Lapitan, N.L.V. Genetic mapping of Dn7, a rye gene conferring resistance to the Russian wheat aphid in wheat. Theor. Appl. Genet. 2003, 107, 1297–1303. [Google Scholar] [CrossRef]
- Han, G.; Liu, S.; Wang, J.; Jin, Y.; Zhou, Y.; Luo, Q.; Liu, H.; Zhao, H.; An, D. Identification of an elite wheat-rye T1RS·1BL translocation line conferring high resistance to powdery mildew and stripe rust. Plant Dis. 2020, 104, 2940–2948. [Google Scholar] [CrossRef]
- Weightman, R.M.; Millar, S.; Alava, J.; Foulkes, M.J.; Fish, L.; Snape, J.W. Effects of drought and the presence of the 1bl/1rs translocation on grain vitreosity, hardness and protein content in winter wheat. J. Cereal Sci. 2008, 47, 457–468. [Google Scholar] [CrossRef]
- Ren, T.H.; Chen, F.; Zou, Y.T.; Jia, Y.H.; Zhang, H.Q.; Yan, B.J.; Ren, Z.L. Evolutionary trends of microsatellites during the speciation process and phylogenetic relationships within the genus Secale. Genome 2011, 54, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Mater, Y.; Baenziger, S.; Gill, K.; Graybosch, R.; Whitcher, L.; Baker, C.; Specht, J.; Dweikat, I. Linkage mapping of powdery mildew and greenbug resistance genes on recombinant 1RS from ‘Amigo’ and ‘Kavkaz’ wheat–rye translocations of chromosome 1RS.1AL. Genome 2004, 47, 292–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Éva, S.; Kitti, S.P.; Balázs, K.; Annamária, S.; László, I.; Marianna, R.; Gyula, V.; István, M.; Márta, M. 1RS arm of Secale cereanum ‘Kriszta’ confers resistance to stripe rust, improved yield components and high arabinoxylan content in wheat. Sci. Rep. 2020, 10, 1792. [Google Scholar]
- Yoshida, S.; Cock, J.H. Growth performance of an improved rice variety in the tropics. Int. Rice Comm. Newsl. 1971, 20, 1–15. [Google Scholar]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Monk, L.S.; Fagerstedt, K.V.; Crawford, R.M. Oxygen toxicity and superoxide dismutase as an antioxidant in physiological stress. Physiol. Plant. 1989, 76, 456–459. [Google Scholar] [CrossRef]
- Mishra, N.P.; Mishra, R.K.; Singhal, G.S. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol. 1993, 102, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Luo, P.; Ren, Z.; Wu, X.; Zhang, H.; Zhang, H.; Feng, J. Structural and biochemical mechanism responsible for the stay-green phenotype in common wheat. Chin. Sci. Bull. 2006, 51, 2595–2603. [Google Scholar] [CrossRef]
- Hodges, D.M.; Delong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbitruic acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Ren, T.; Sun, Z.; Ren, Z.; Tan, F.; Luo, P.; Tang, Z.; Fu, S.; Li, Z. Molecular and cytogenetic characterization of a wheat-rye 7BS.7RL translocation line with resistance to stripe rust, powdery mildew and Fusarium head blight. Phytopathology 2020, 110, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; He, M.; Sun, Z.; Tan, F.; Luo, P.; Tang, Z.; Fu, S.; Yan, B.; Ren, Z.; Li, Z. The Polymorphisms of oligonucleotide probes in wheat cultivars determined by ND-FISH. Molecules 2019, 24, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Tang, Z.; Duan, Q.; Tang, S.; Fu, S. Using the 6RLKu minichromosome of rye (Secale cereale L.) to create wheat-rye 6D/6RLKu small segment translocation lines with powdery mildew resistance. Int. J. Mol. Sci. 2018, 19, 3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure from small quantities of fresh leaf tissues. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Chai, J.F.; Zhou, R.H.; Jia, J.Z.; Liu, X. Development and application of a new codominant PCR marker for detecting 1BL.1RS wheat–rye chromosome translocations. Plant Breed. 2006, 125, 302–304. [Google Scholar] [CrossRef]
- Zhang, L.; He, Z.; Lu, M.; Pang, B.; Zhang, X.; Xia, L.; Ellison, F. Identification of 1BL/1RS translocation via multiplex PCR, markers of Glu-B3, Gli-B1 and SEC-1b in common wheat. Sci. Agric. Sin. 2003, 36, 1566–1570. [Google Scholar]
- Weng, D.; Xu, S.; Lin, R.; Wan, A.; Li, J.; Wu, L. Microsatellite marker linked with stripe rust resistant gene Yr9 in wheat. Acta Gene. Sin. 2005, 32, 937–941. [Google Scholar]
- Mago, R.; Spielmeyer, W.; Lawrence, G.J.; Lagudah, E.S.; Ellis, J.G.; Pryor, A. Identification and mapping of molecular markers linked to rust resistance genes located on chromosome 1RS of rye using wheat-rye translocation lines. Theor. Appl. Genet. 2002, 104, 1317–1324. [Google Scholar] [CrossRef]
- Ren, T.; Jiang, Q.; Sun, Z.; Zhao, L.; Peng, W.; Ren, Z.; Tan, F.; Luo, P.; Li, Z. Development and molecular cytogenetic characterization of novel primary wheat-rye 1RS.1BL translocation lines from multiple rye sources with resistance to stripe rust. Plant Dis. 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Wan, A.; Zhao, Z.; Chen, X.; He, Z.; Jin, S.; Jia, Q.; Yao, G.; Yang, J.; Wang, B.; Li, G. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddling, J.; Gelang-Alfredsson, J.; Piikki, k.; Pleijel, H. Evaluating the relationship between leaf chlorophyll concentration and SPAD-502 chlorophyll meter readings. Photosynth. Res. 2007, 91, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Behera, R.K.; Mishra, P.C.; Choudhury, N.K. High irradiance and water stress induce alterations in pigment composition and chloroplast activities of primary wheat leaves. J. Plant Physiol. 2002, 159, 967–973. [Google Scholar] [CrossRef]
- Kraus, T.E.; Fletcher, R.A. Paclotutrazol protects wheat seedling from heat and paraquat injury. Is detoxification of active oxygen involved? Plant Cell Physiol. 1994, 35, 45–52. [Google Scholar]
- Li, J.; Deng, L.; Jin, J.; Yan, Y. Optimization of grains-rice extrusion process based on flavonoids retention and gelatinization degree. Food Mach. 2014, 30, 190–195. [Google Scholar]
- Huang, Y.; Li, D.; Wang, L.; Yong, C.; Sun, E.; Jin, H.; Huang, H. Decreased enzyme activities, ammonification rate and ammonifiers contribute to higher nitrogen retention in hyperthermophilic pretreatment composting. Bioresour. Technol. 2019, 272, 521–528. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Translocation Lines and Controls | Chromosome Type | Pst Pathotypes and Isolates | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CYR32 | CYR33 | CYR34 | SY3 | SY4 | SY5 | SY7 | HY8 | In the Field | ||
| RT843-5 | T1RS.1BL | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| CN11 | T1RS.1BL | 6 | 6 | 8 | 8 | 8 | 0 | 0 | 6 | 7 |
| MY11 | 1B | 8 | 8 | 8 | 8 | 8 | 8 | 6 | 0 | 9 |
| Cross | Pst Mixture | ||||
|---|---|---|---|---|---|
| Resistant | Segregation | Susceptible | Ratio | χ2 | |
| (RT843 × MY11) Testcross | 217 | - | 201 | 1:1 | 0.62 |
| (RT843 × MY11) F2 | 224 | 67 | 3:1 | 0.60 | |
| (RT843 × MY11) F3 | 64 | 160 | 67 | 1:2:1 | 2.95 |
| Lines | Yield (kg/ha) | NS (m-2) | KN (per Spike) | TKW (g) | AGB (kg/ha) | HI (%) |
|---|---|---|---|---|---|---|
| MY11 | 5271.3 ± 113.1a | 321.7 ± 10.3a | 43.3 ± 2.2a | 44.4 ± 0.4a | 13385.5 ± 1084.8ab | 46.6 ± 0.2a |
| RT843-5 | 5980 ± 109.4b | 307.2 ± 6.1a | 40.7 ± 0.7a | 47.8 ± 0.4b | 12901.7 ± 203.3a | 46.4 ± 0.2a |
| Molecular Markers | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| ω-Sec-P | accttcctcatctttgtcct | ccgatgcctataccactact |
| Gil-B1 | gcagacctgtgtcattggtc | gatatagtggcagcaggatacg |
| Xgwm582 | aagcactacgaaaatatgac | tcttaaggggtgttatcata |
| Iag95 | ctctgtggatagttacttgatcga | cctagaacatgcatggctgttaca |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Jiang, Q.; Fan, T.; Zhao, L.; Ren, Z.; Tan, F.; Luo, P.; Ren, T. Molecular Cytogenetic and Physiological Characterization of a Novel Wheat-Rye T1RS.1BL Translocation Line from Secale cereal L. Weining with Resistance to Stripe Rust and Functional “Stay Green” Trait. Int. J. Mol. Sci. 2022, 23, 4626. https://doi.org/10.3390/ijms23094626
Li Z, Jiang Q, Fan T, Zhao L, Ren Z, Tan F, Luo P, Ren T. Molecular Cytogenetic and Physiological Characterization of a Novel Wheat-Rye T1RS.1BL Translocation Line from Secale cereal L. Weining with Resistance to Stripe Rust and Functional “Stay Green” Trait. International Journal of Molecular Sciences. 2022; 23(9):4626. https://doi.org/10.3390/ijms23094626
Chicago/Turabian StyleLi, Zhi, Qing Jiang, Tao Fan, Liqi Zhao, Zhenglong Ren, Feiquan Tan, Peigao Luo, and Tianheng Ren. 2022. "Molecular Cytogenetic and Physiological Characterization of a Novel Wheat-Rye T1RS.1BL Translocation Line from Secale cereal L. Weining with Resistance to Stripe Rust and Functional “Stay Green” Trait" International Journal of Molecular Sciences 23, no. 9: 4626. https://doi.org/10.3390/ijms23094626
APA StyleLi, Z., Jiang, Q., Fan, T., Zhao, L., Ren, Z., Tan, F., Luo, P., & Ren, T. (2022). Molecular Cytogenetic and Physiological Characterization of a Novel Wheat-Rye T1RS.1BL Translocation Line from Secale cereal L. Weining with Resistance to Stripe Rust and Functional “Stay Green” Trait. International Journal of Molecular Sciences, 23(9), 4626. https://doi.org/10.3390/ijms23094626