
Streptozotocin-Induced Diabetes Causes Changes in Serotonin-Positive Neurons in the Small Intestine in Pig Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
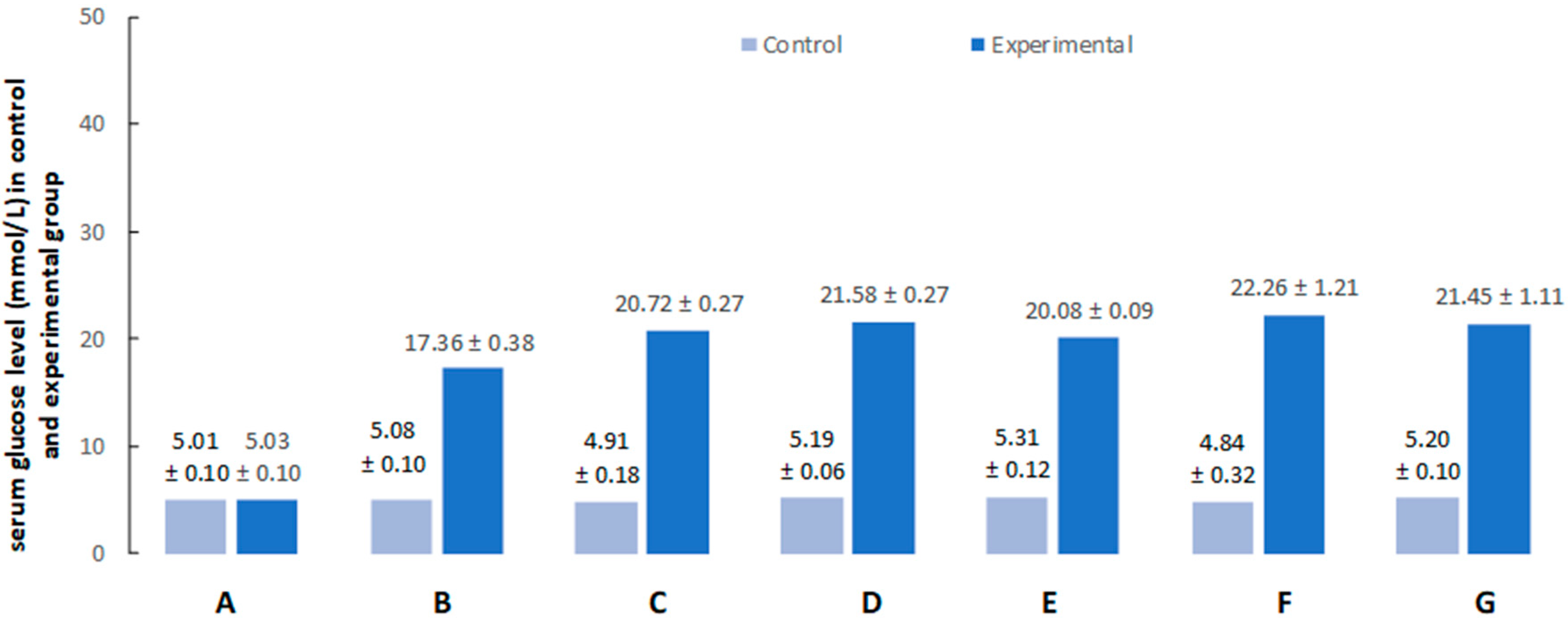
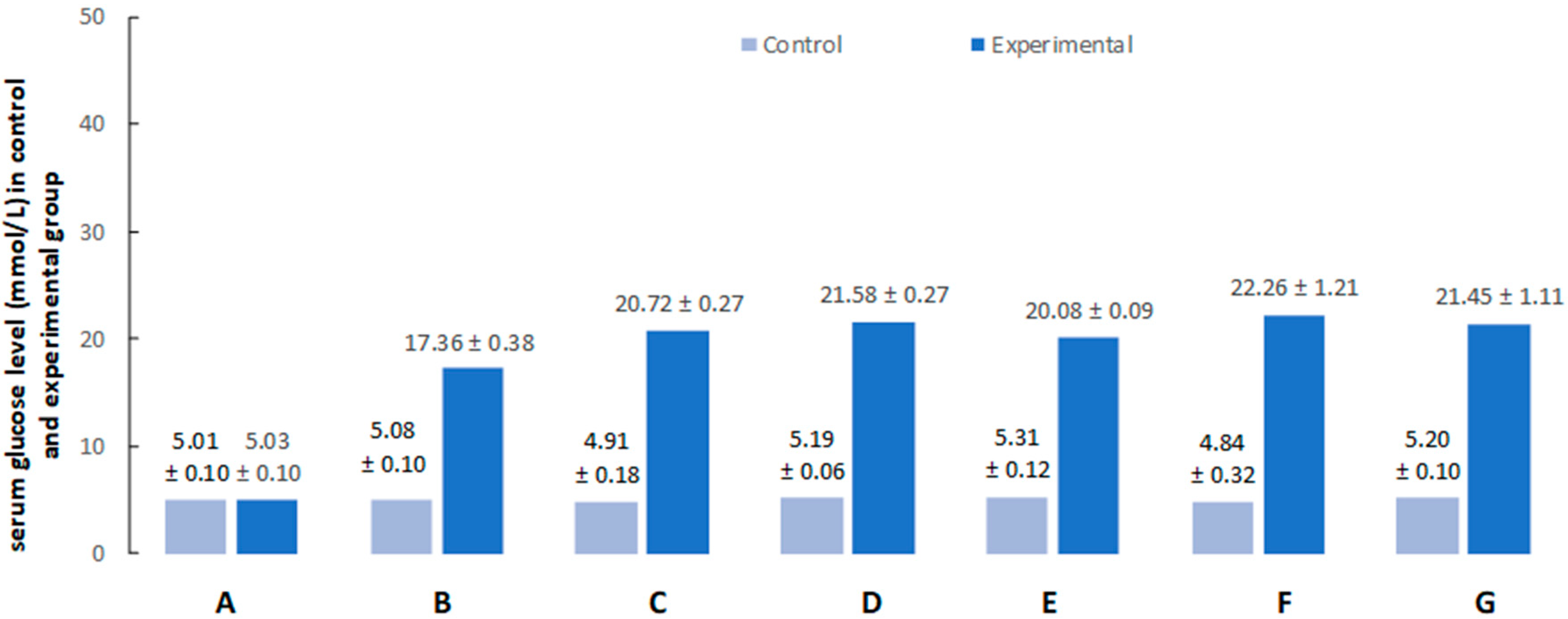
2.1. Hyperglycemia
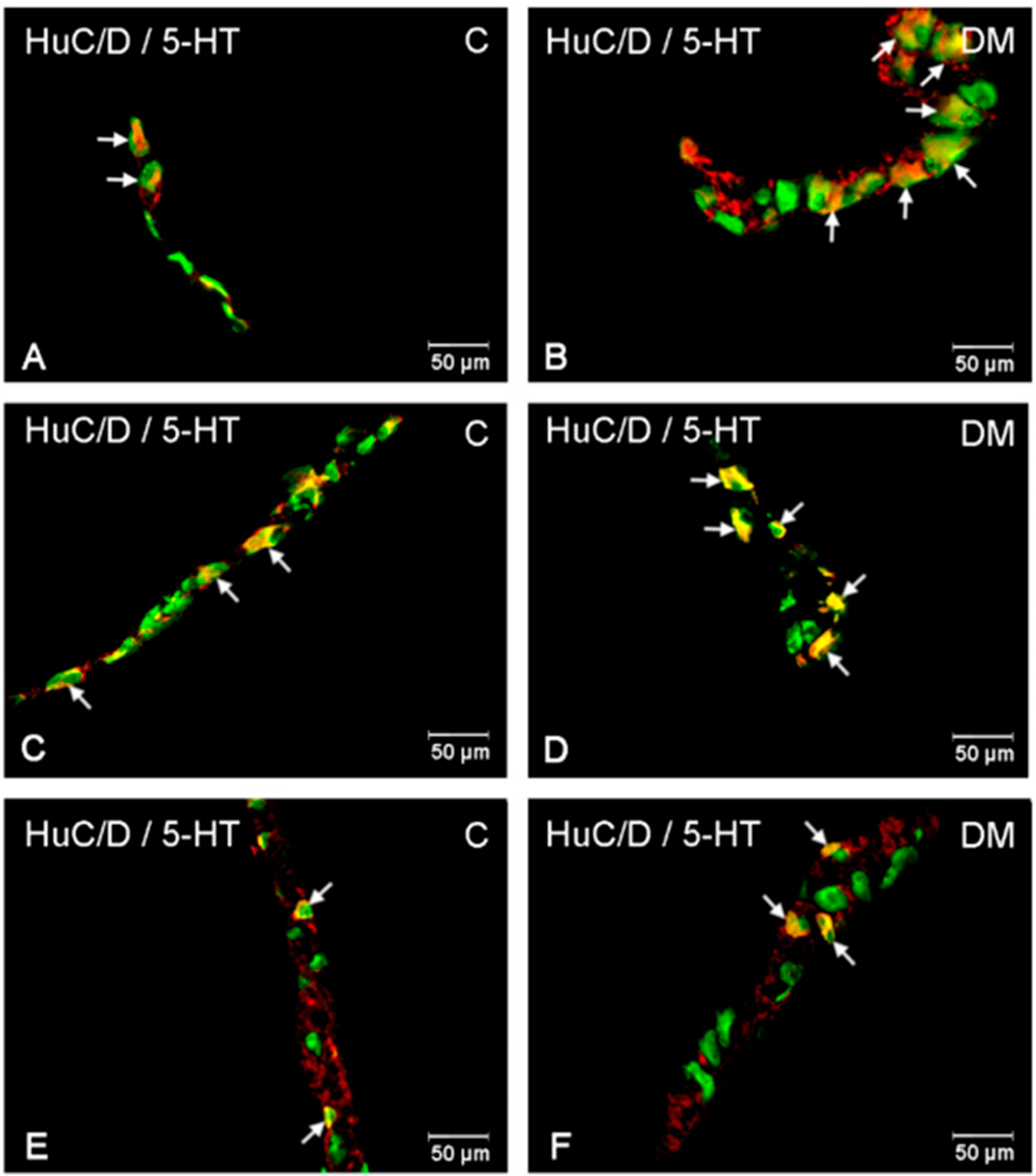
2.2. Immunofluorescence
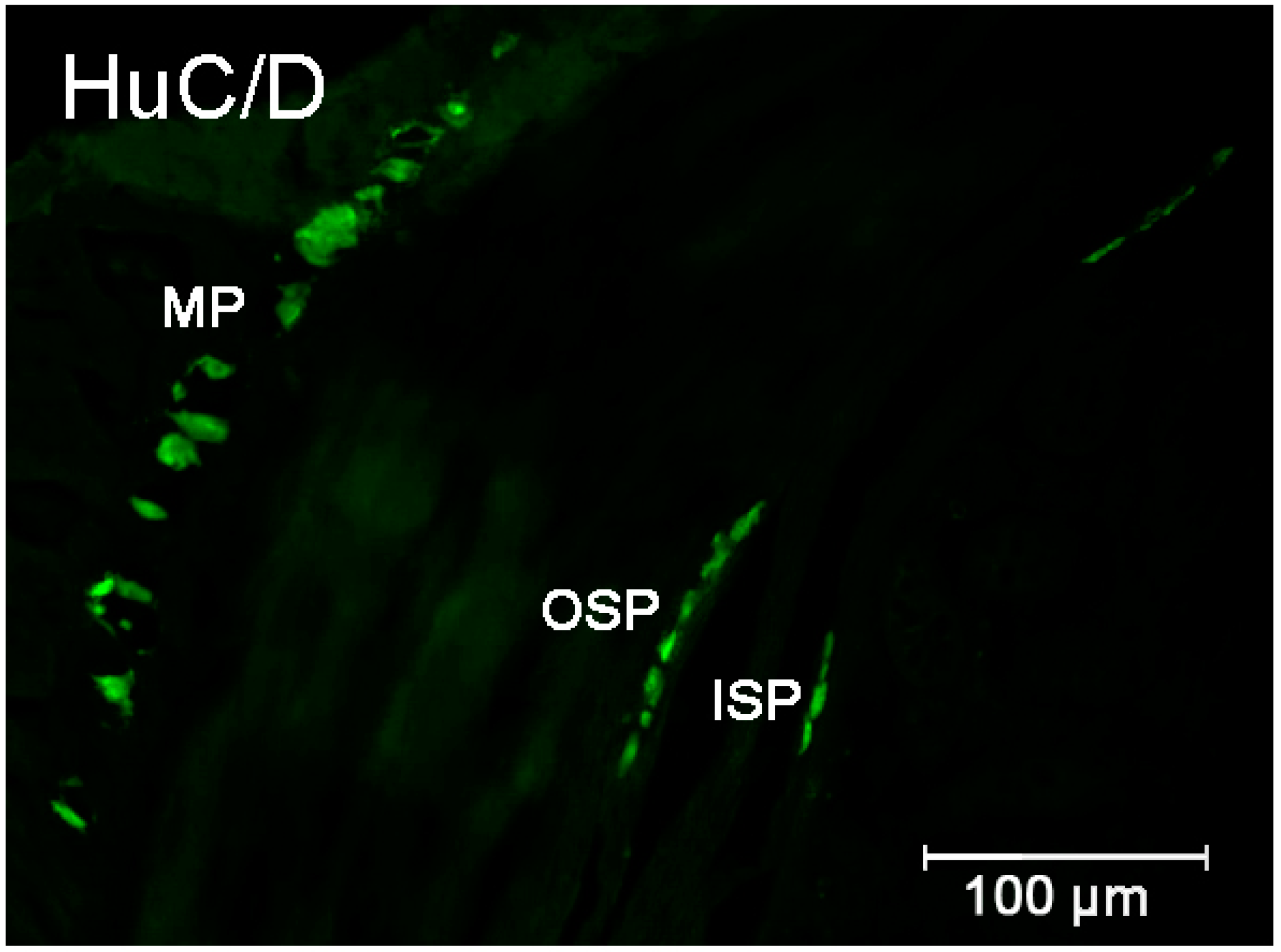
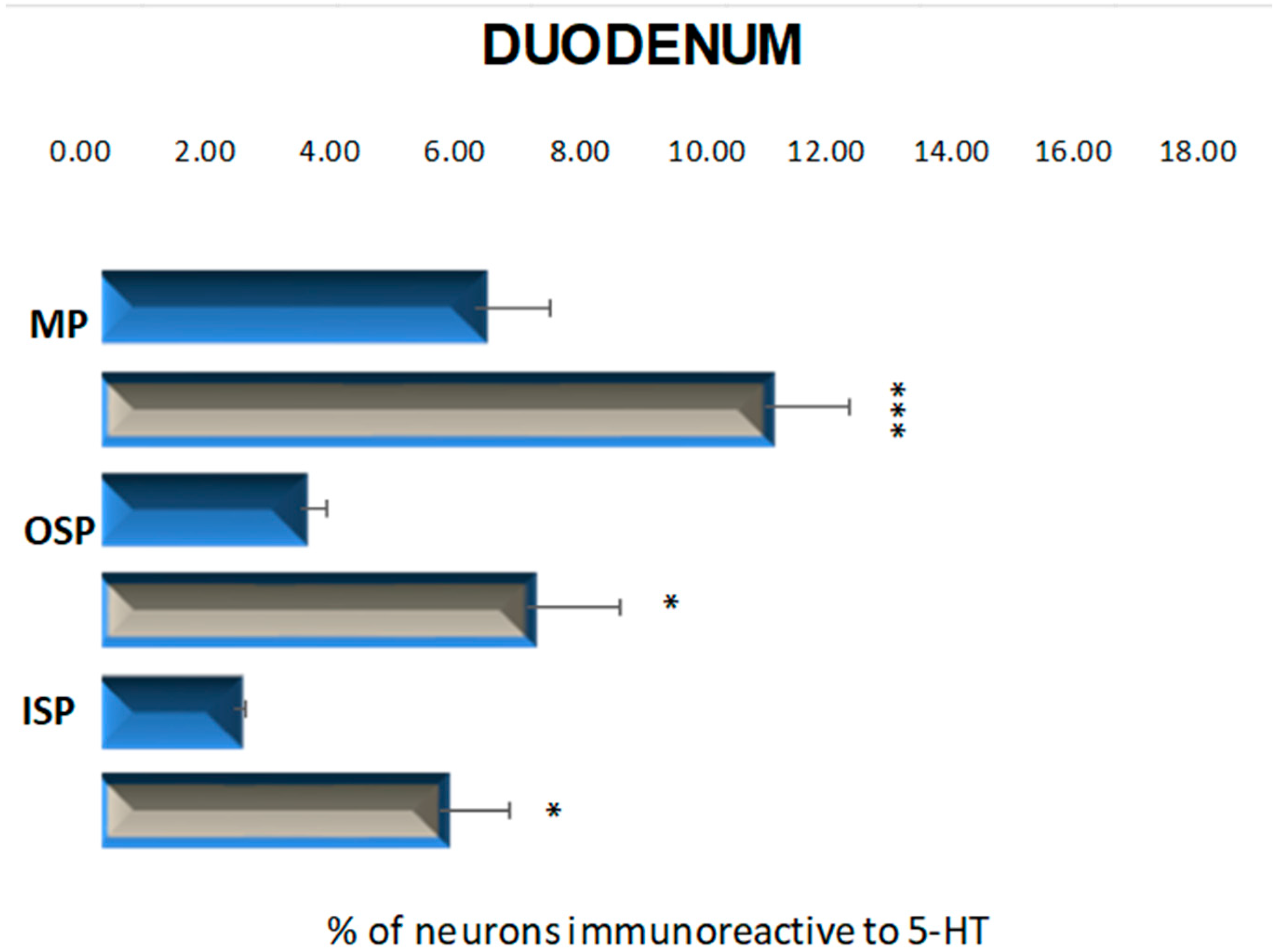
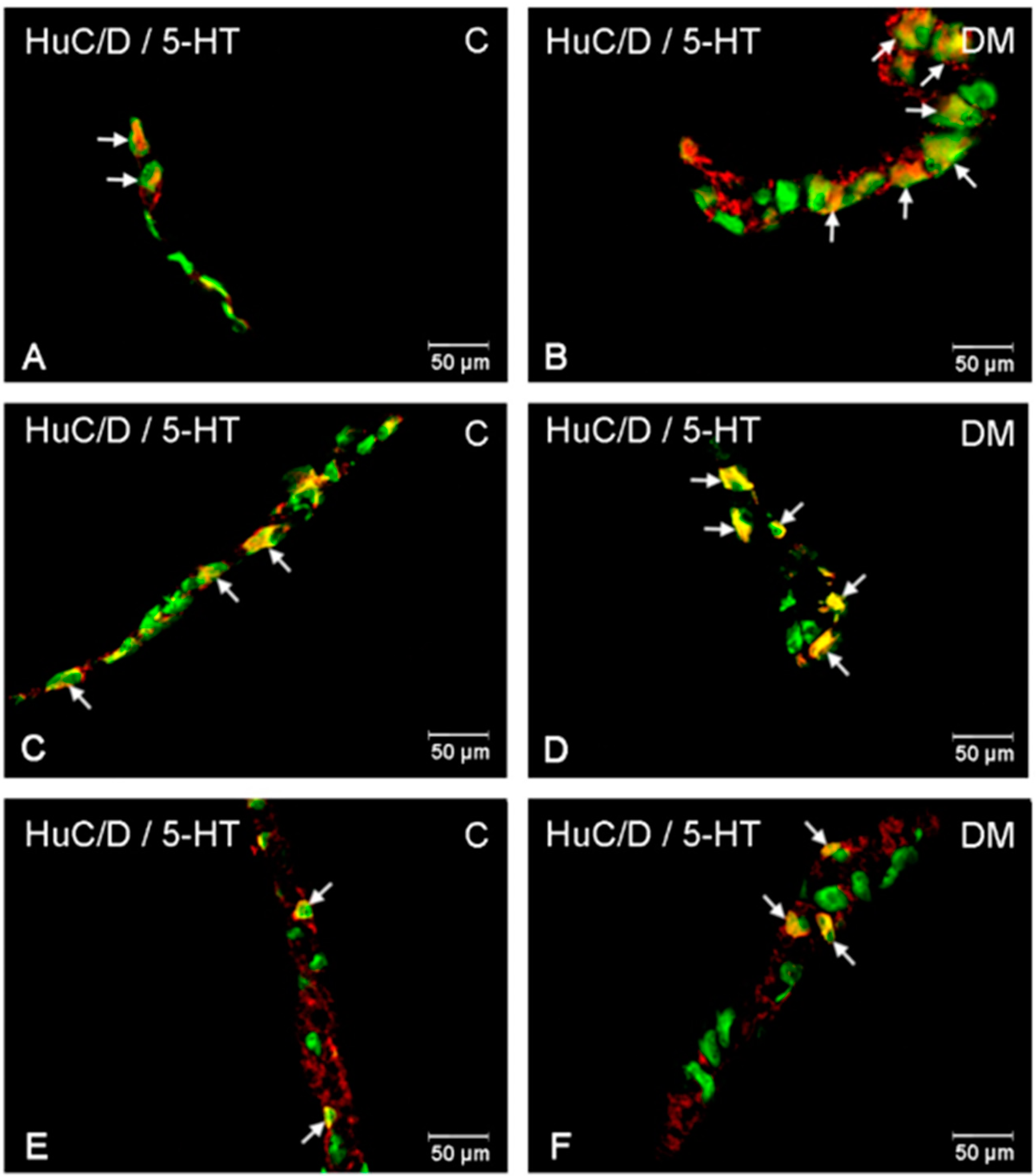
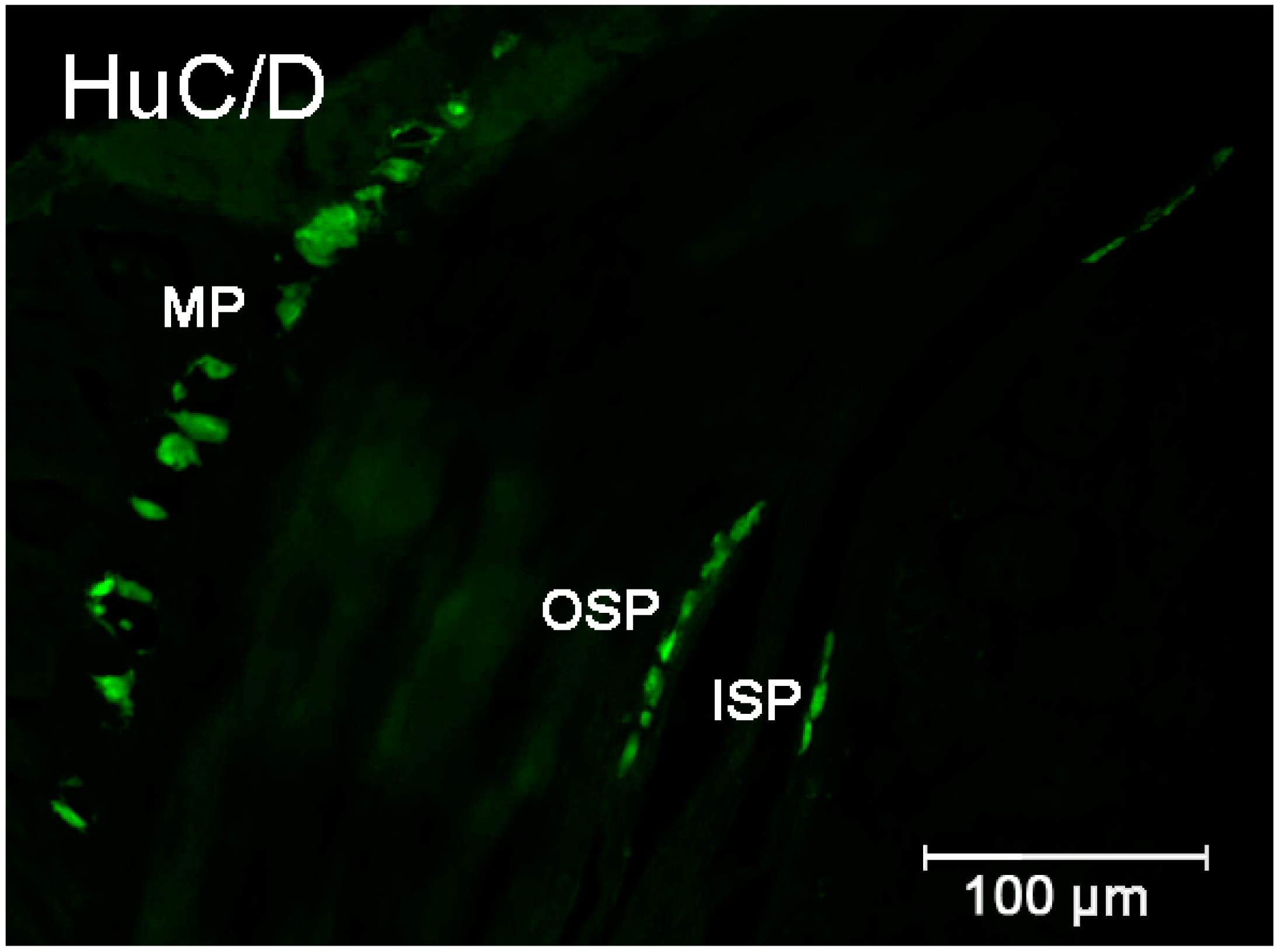
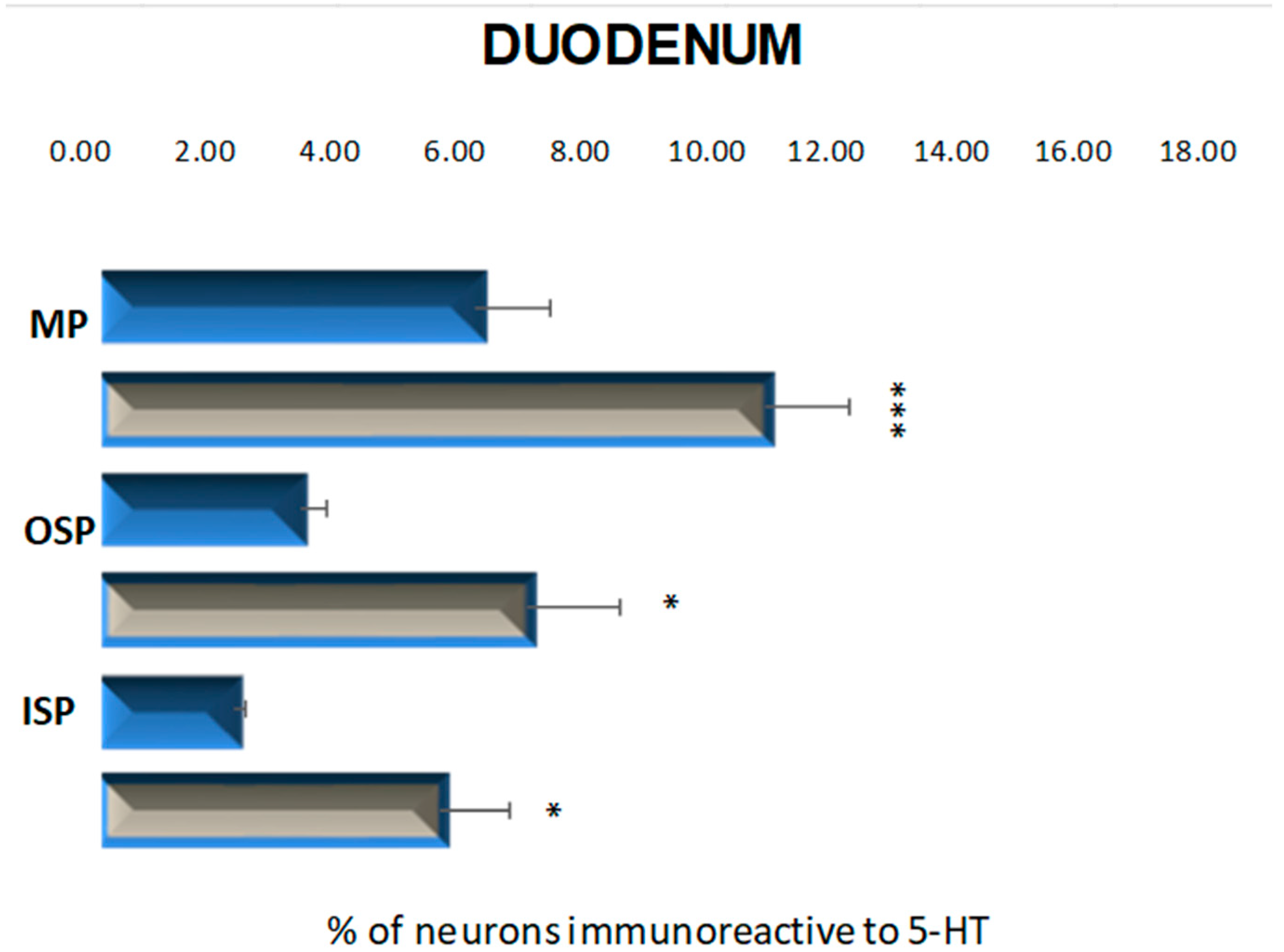
2.2.1. Duodenum
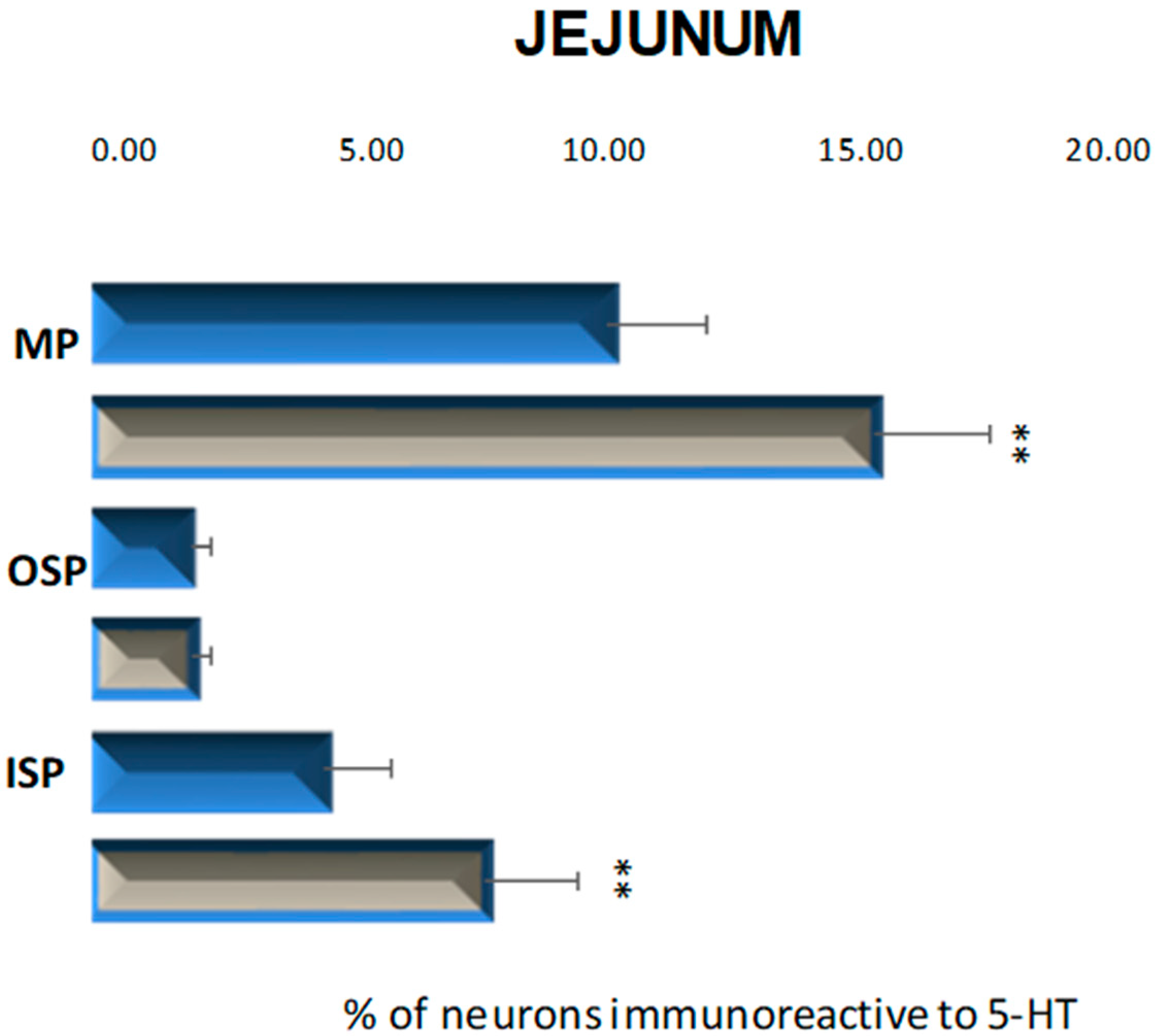
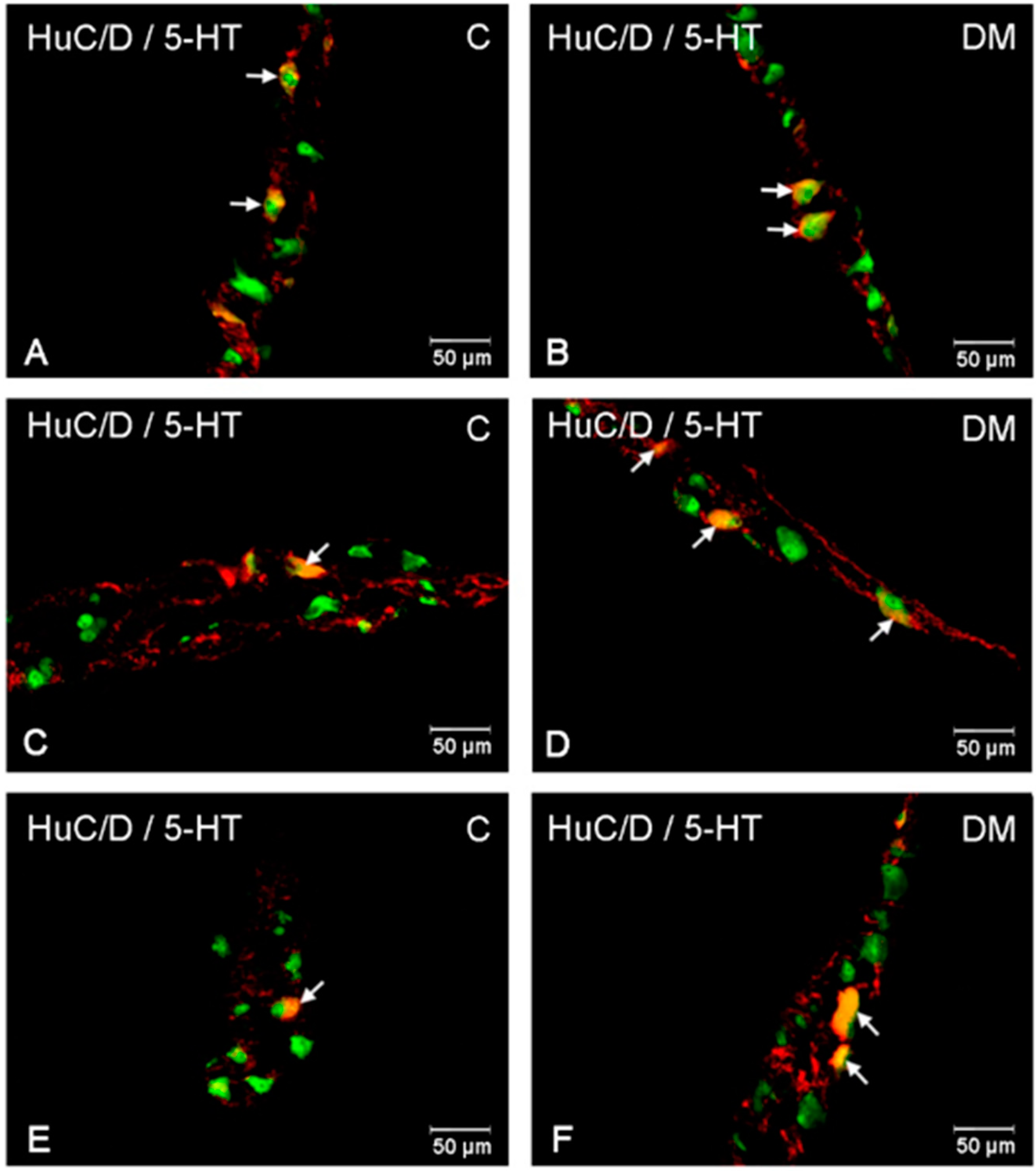
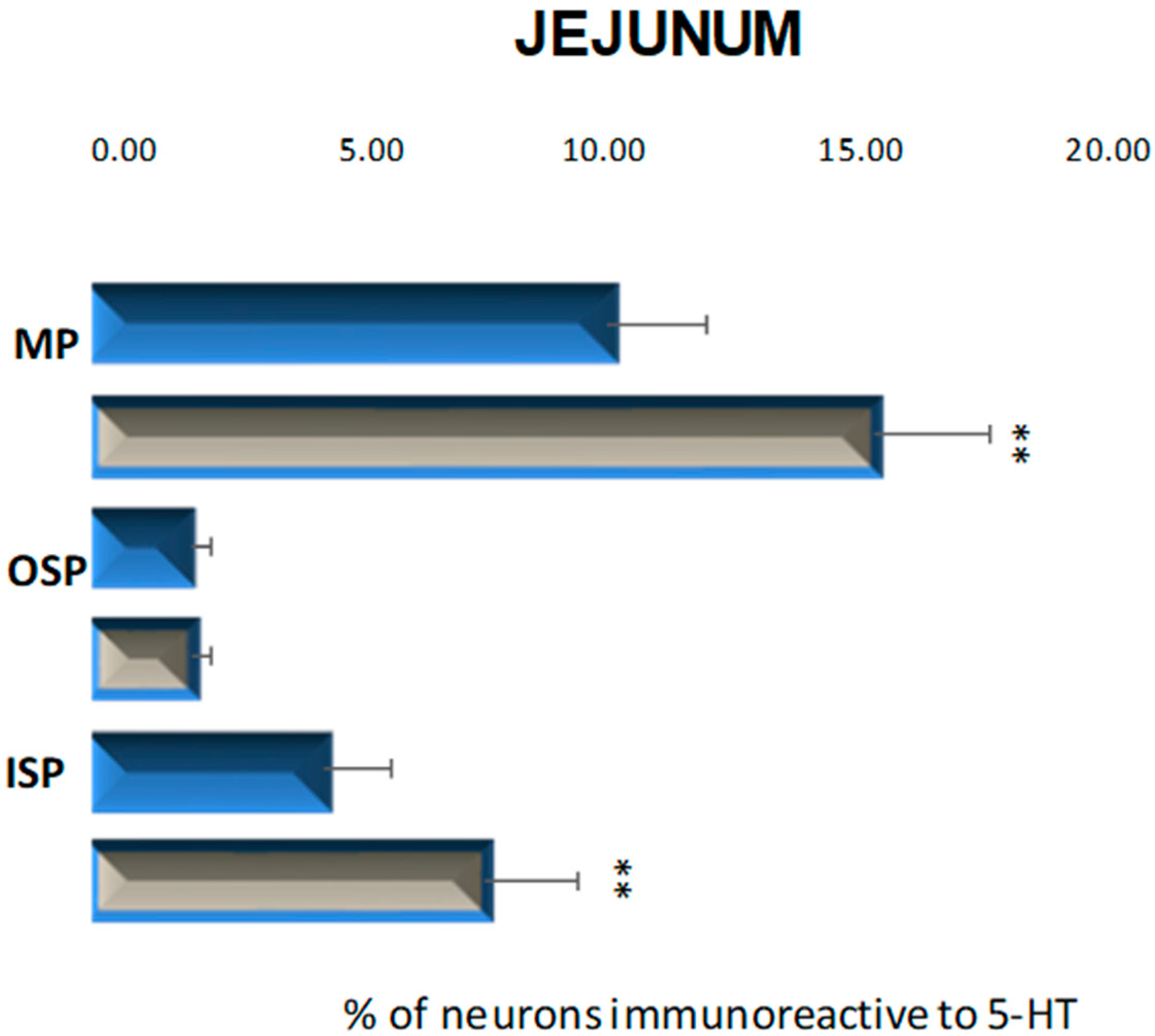
2.2.2. Jejunum
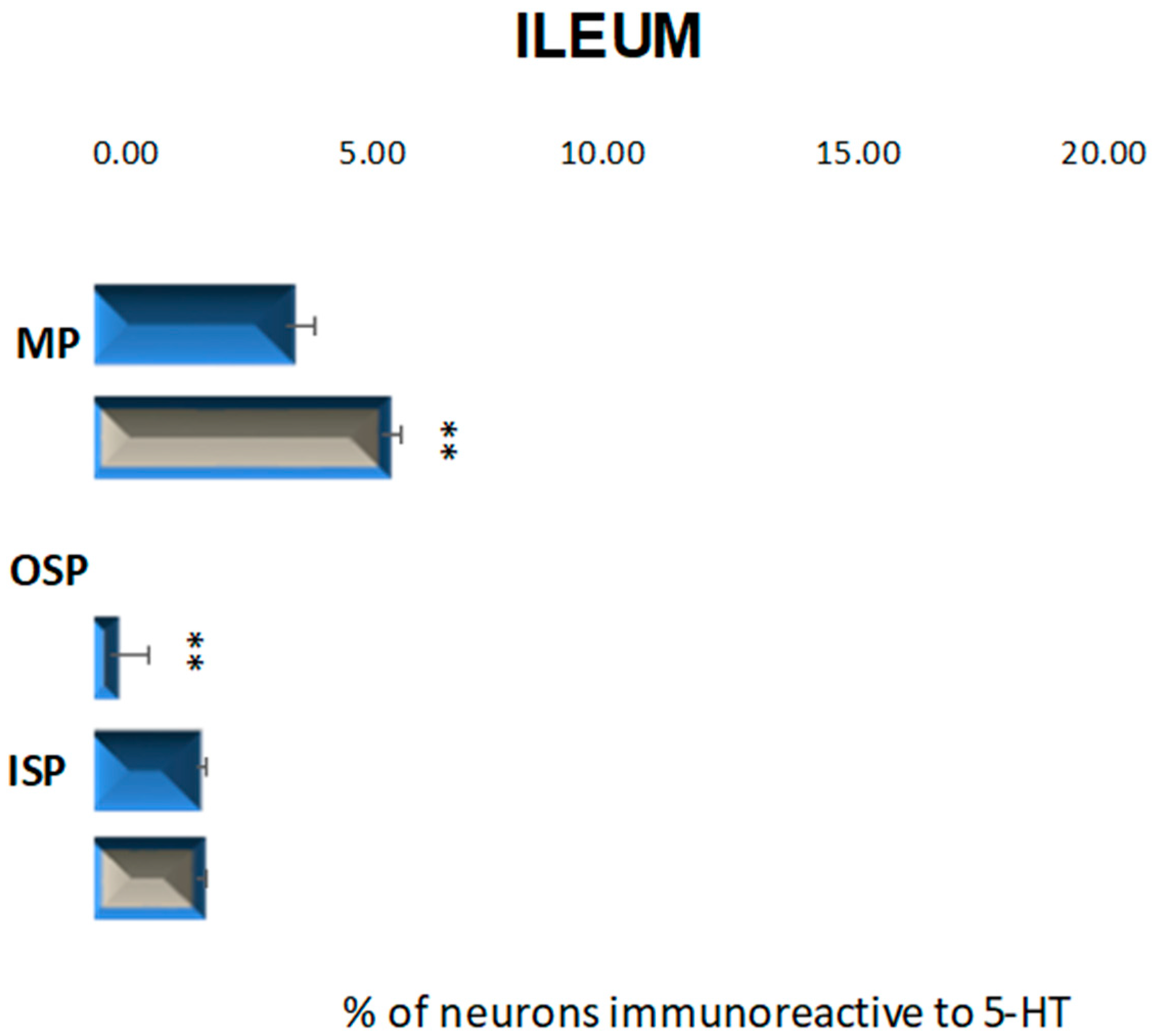
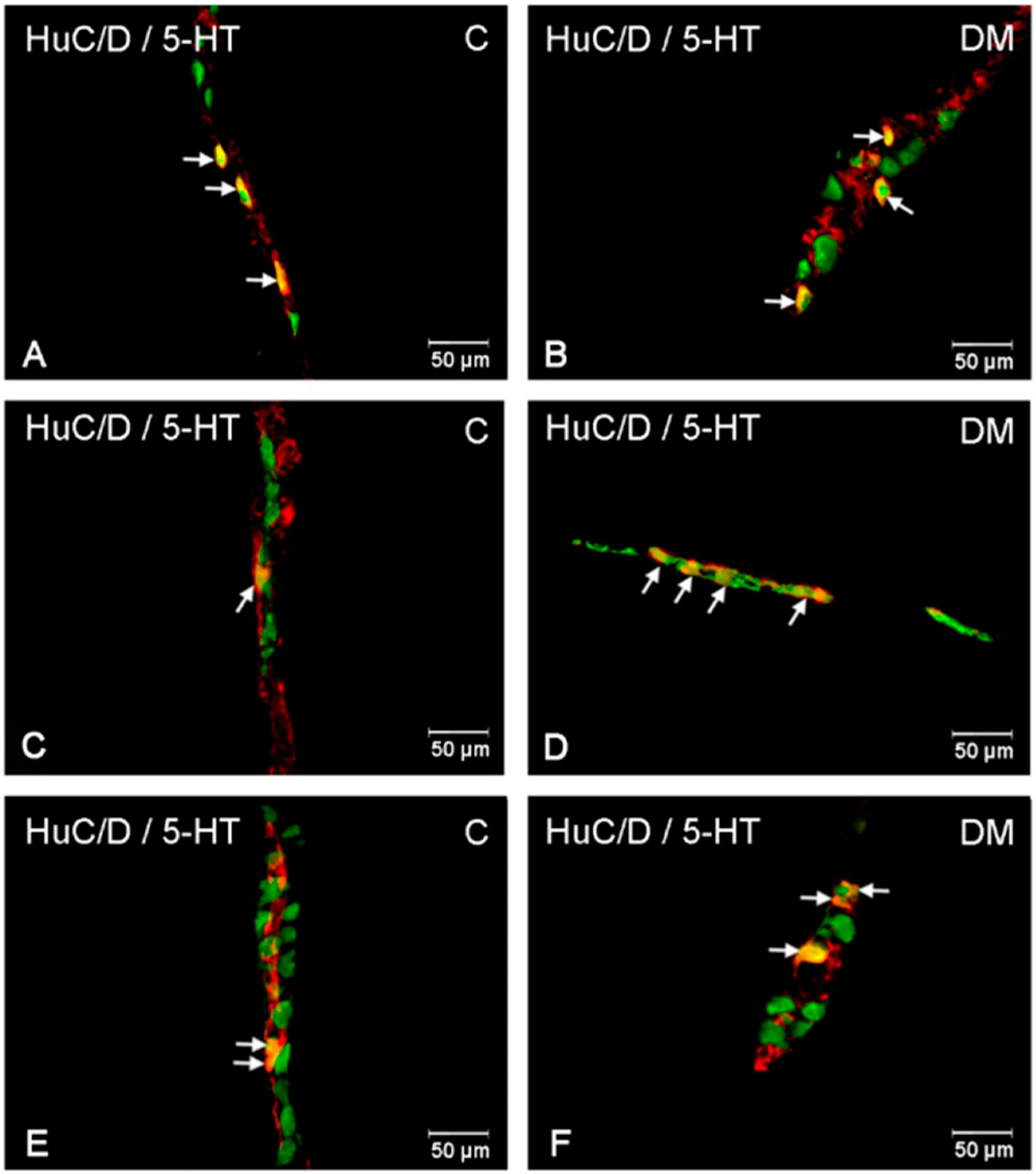
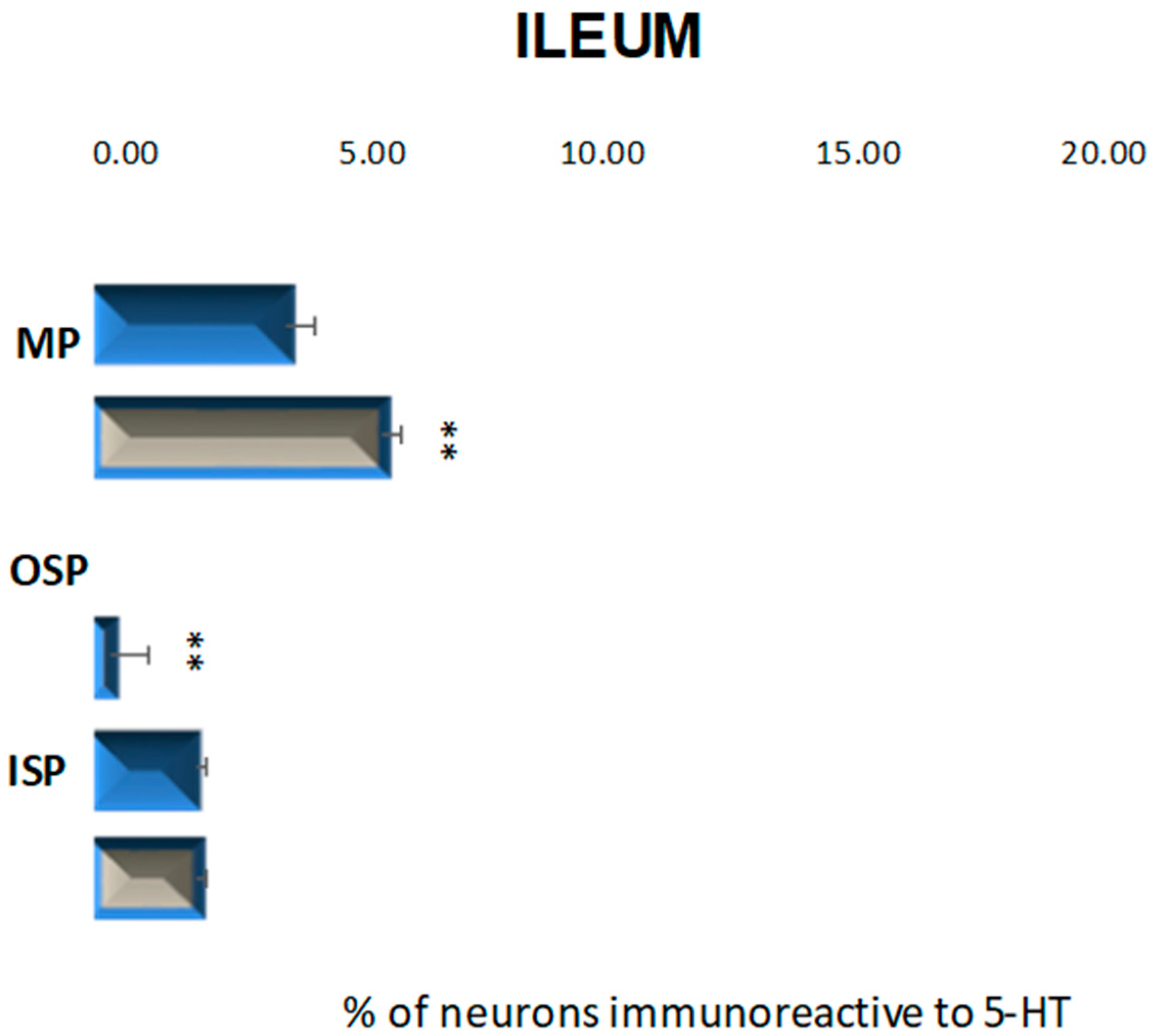
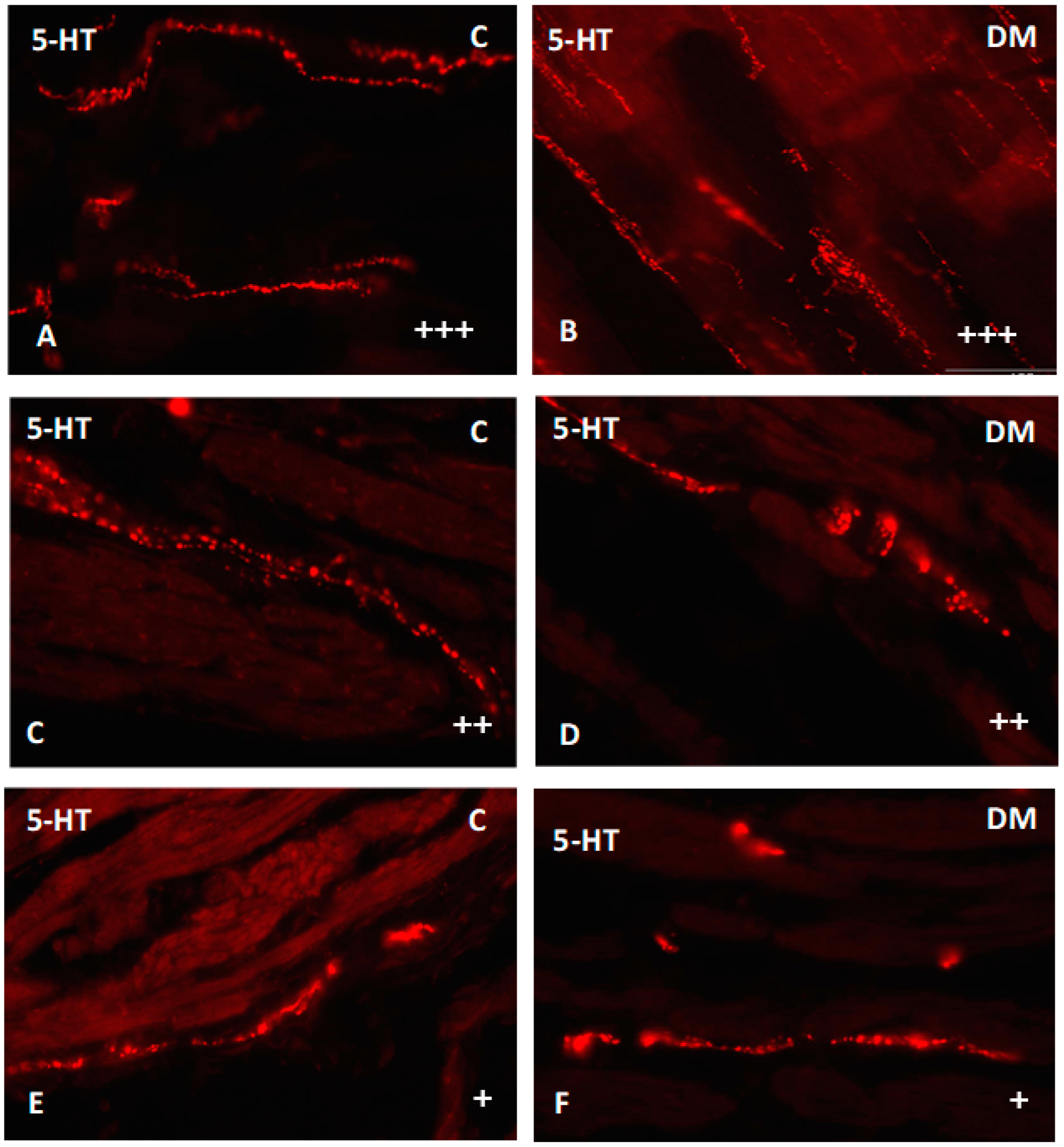
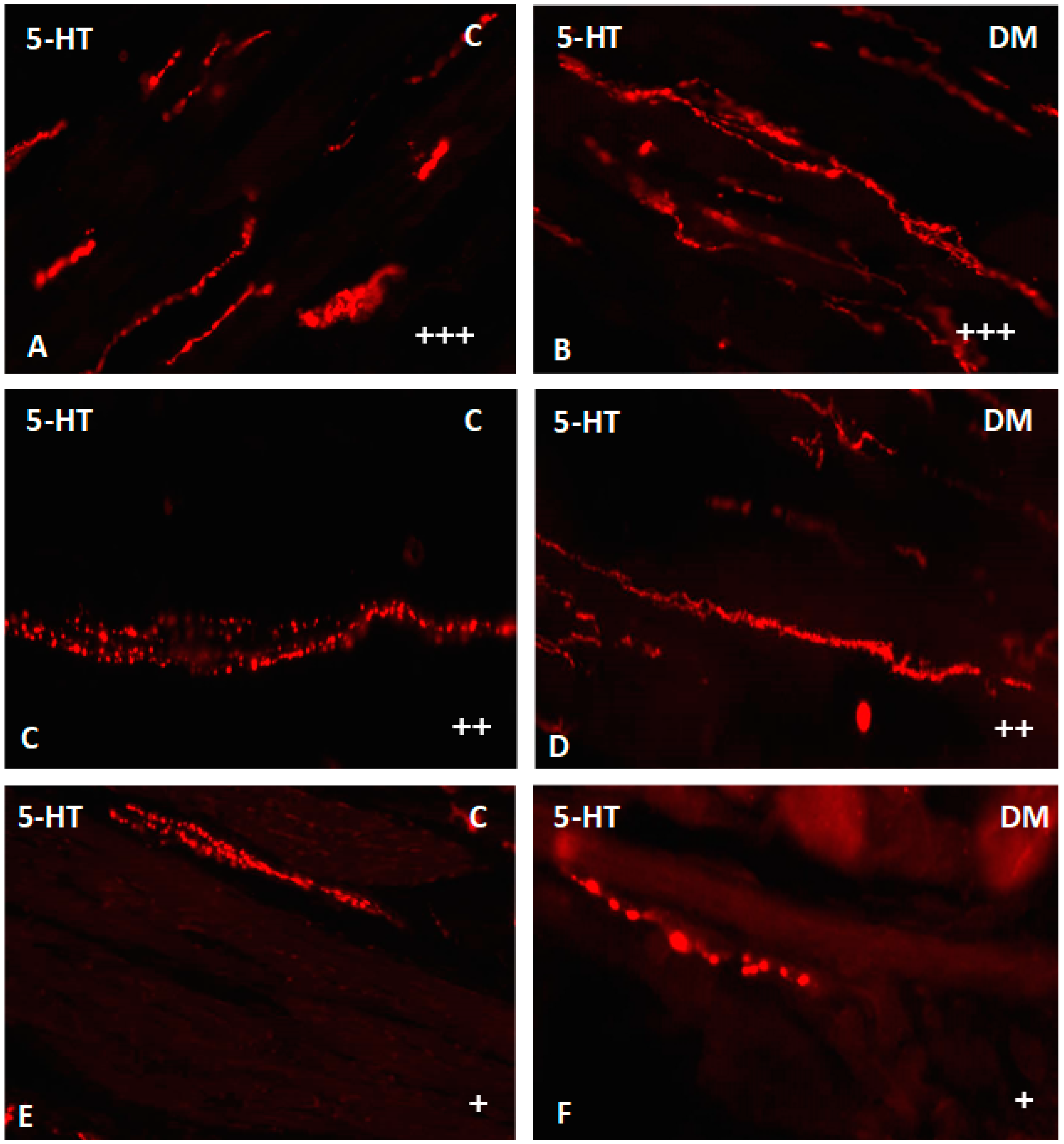
2.2.3. Ileum
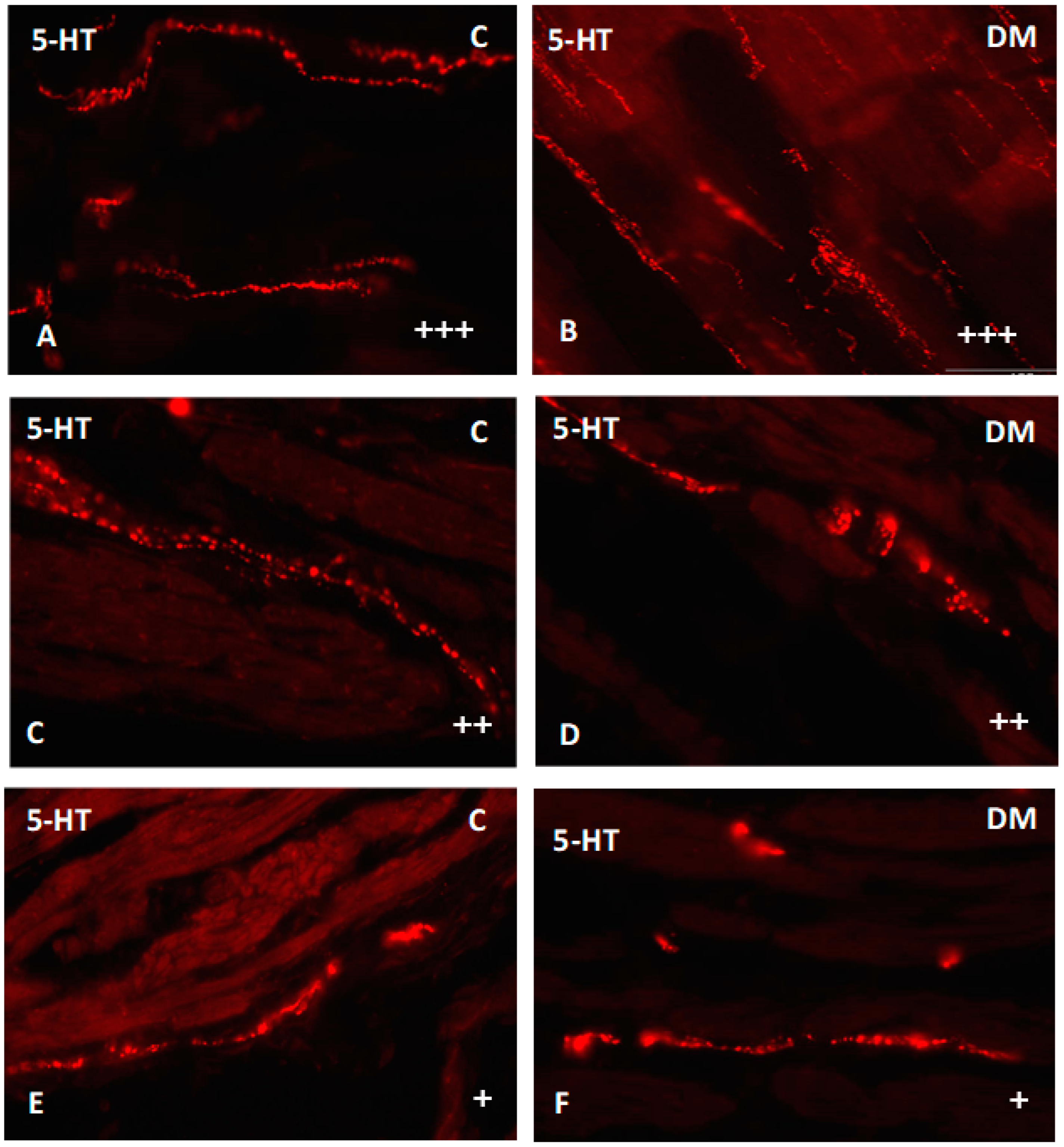
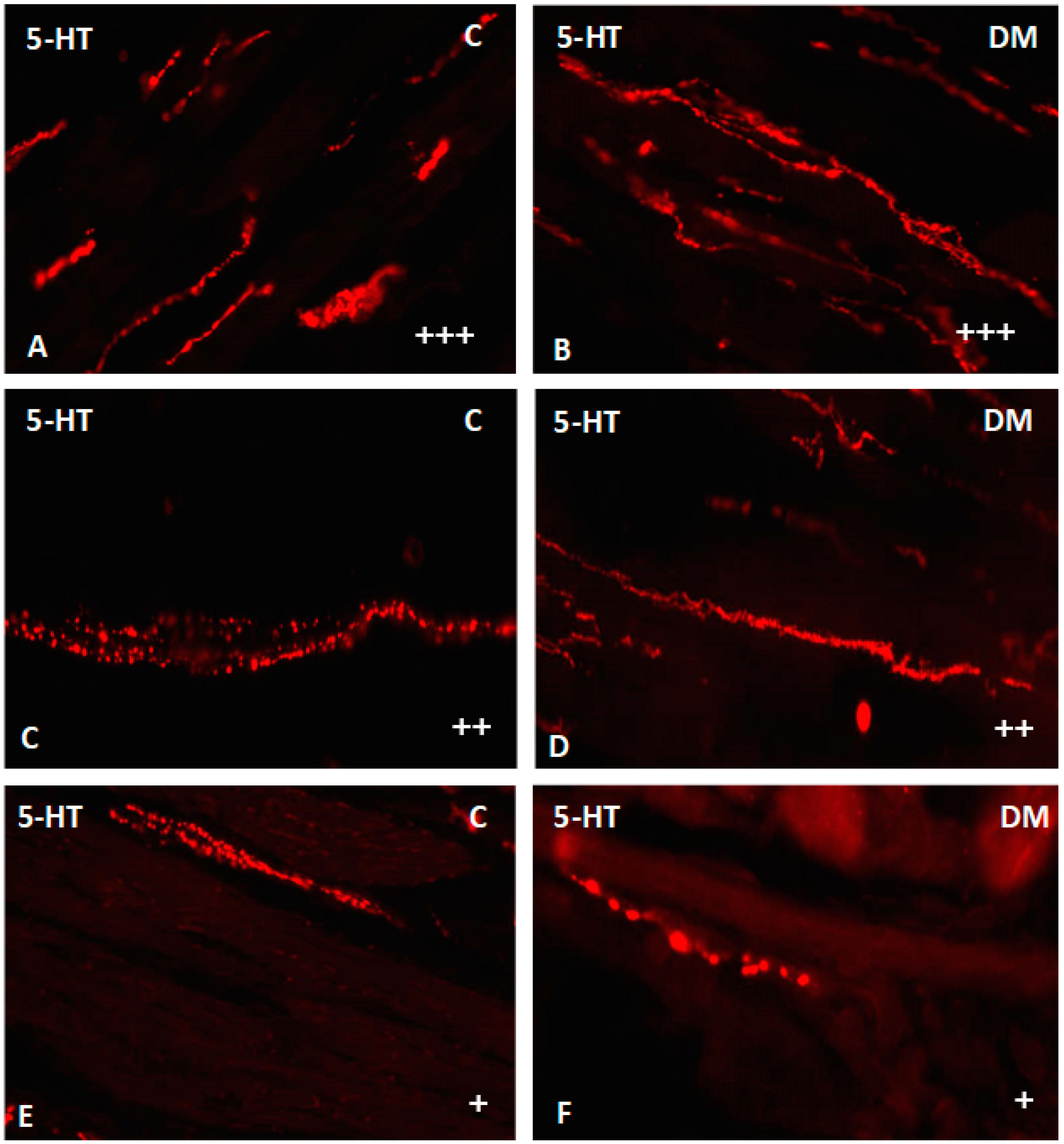
2.2.4. Nerve Fibers
3. Discussion
4. Materials and Methods
Counting of the Nerve Structures and Statistical Evaluation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamlin, K.E.; Fischer, F.E. The synthesis of 5-hydroxytryptamine. J. Am. Chem. Soc. 1951, 73, 5007. [Google Scholar] [CrossRef]
- Erspamer, V.; Asero, B. Identification of enteramine, the specific hormone of the enterochromaffin cell system, as 5-hydroxytryptamine. Nature 1951, 169, 800. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Drakontides, A.B.; Ross, L.L. Serotonin: Synthesis and release from the myenteric plexus of the mouse intestine. Science 1965, 149, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Camilleri, M. Serotonin: A mediator of the brain-gut connection. Am. J. Gastroenterol. 2000, 95, 2698–2709. [Google Scholar] [CrossRef] [PubMed]
- Banskota, S.; Ghia, J.E.; Khan, W.I. Serotonin in the gut: Blessing or a curse. Biochimie 2019, 161, 56–64. [Google Scholar] [CrossRef]
- Koopman, N.; Katsavelis, D.; Hove, A.S.T.; Brul, S.; de Jonge, W.J.; Seppen, J. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef]
- Liu, N.; Sun, S.; Wang, P.; Sun, Y.; Hu, Q.; Wang, X. The Mechanism of Secretion and Metabolism of Gut-Derived 5-Hydroxytryptamine. Int. J. Mol. Sci. 2021, 22, 7931. [Google Scholar] [CrossRef]
- Gershon, M.D. Serotonin receptors and transporters-roles in normal and abnormal gastrointestinal motility. Aliment. Pharmacol. Ther. 2004, 20, 3–14. [Google Scholar] [CrossRef]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 2002, 71, 533–554. [Google Scholar] [CrossRef]
- Annaházi, A.; Berger, T.E.; Demir, I.E.; Zeller, F.; Müller, M.; Anneser, M.; Skerra, A.; Michel, K.; Schemann, M. Metabotropic 5-HT receptor-mediated effects in the human submucous plexus. Neurogastroenterol Motil. 2022, 19, e14380. [Google Scholar] [CrossRef] [PubMed]
- Jonnakuty, C.; Gragnoli, C. What do we know about serotonin? J. Cell. Physiol. 2008, 217, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Seuwen, K.; Pouyssegur, J. Serotonin as a growth factor. Biochem. Pharmacol. 1999, 39, 985–990. [Google Scholar] [CrossRef]
- Fanburg, B.L.; Lee, S.L. A new role for an old molecule: Serotonin as a mitogen. Am. J. Physiol. 1997, 272, L795–L806. [Google Scholar] [CrossRef]
- Coates, M.D.; Tekin, I.; Vrana, K.E.; Mawe, G.M. Review article: The many potential roles of intestinal serotonin (5-hydroxytryptamine, 5-HT) signalling in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2017, 46, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin, Germany, 2014; Volume 817, pp. 39–71. [Google Scholar]
- Furness, J.B. The organisation of the autonomic nervous system: Peripheral connections. Auton. Neurosci. 2006, 130, 1–5. [Google Scholar] [CrossRef]
- Schemann, M.; Neunlist, M. The human enteric nervous system. Neurogastroenterol. Motil. 2004, 16, 55–59. [Google Scholar] [CrossRef]
- Destexhe, A.; Marder, E. Plasticity in single neuron and circuit computations. Nature 2004, 431, 789–795. [Google Scholar] [CrossRef]
- Forbes, J.M.; Cooper, M.E. Mechanisms of diabetic complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Chandrasekharan, B.; Srinivasan, S. Diabetes and the enteric nervous system. Neurogastroenterol. Motil. 2007, 19, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Bennet, H.; Balhuizen, A.; Medina, A.; Nitert, M.D.; Laakso, E.O.; Essen, S.; Spegel, P.; Storm, P.; Krus, U.; Wierup, N.; et al. Altered serotonin (5-HT) 1D and 2A receptor expression may contribute to defective insulin and glucagon secretion in human type 2 diabetes. Peptides 2015, 71, 113–120. [Google Scholar] [CrossRef] [PubMed]
- De Deurwaerdère, P.; Di Giovanni, G. Serotonin in Health and Disease. Int. J. Mol. Sci. 2020, 21, 3500. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.C.; Cheng, H.T.; Stables, C.L.; Smith, A.L.; Feldman, E.L. Diabetic neuropathy: Clinical manifestations and current treatments. Lancet Neurol. 2012, 11, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.C.; Motta, M.E.F.A. Mechanisms and factors associated with gastrointestinal symptoms in patients with diabetes mellitus. J. Pediatr. 2012, 88, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, E.; Braun-Reichhart, C.; Streckel, E.; Renner, S. Genetically engineered pig models for diabetes research. Transgenic Res. 2014, 23, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.O.; Rolin, B. Use of the Goettingen Minipig as a Model of Diabetes with Special Focus on Type 1 Diabetes Research. ILAR 2004, 45, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulc, M.; Palus, K.; Zielonka, Ł.; Gajecka, M.; Całka, J. Changes in expression of inhibitory substances in the intramural neurons of the stomach following streptozotocin- induced diabetes in the pig. World J. Gastroenterol. 2017, 23, 6088–6099. [Google Scholar] [CrossRef]
- Atkinson, W.; Lockhart, S.; Whorwell, P.; Keevil, B.; Houghton, L. Altered 5-hydroxytryptamine signaling in patients with constipation- and diarrhea-predominant irritable bowel syndrome. Gastroenterology 2006, 130, 34–43. [Google Scholar] [CrossRef]
- Xie, Z.; Chang, C.; Zhou, Z. Molecular mechanisms in autoimmune type 1 diabetes: A critical review. Clin. Rev. Allergy Immunol. 2014, 47, 174–192. [Google Scholar] [CrossRef]
- Takahara, H.; Fujimura, M.; Taniguchi, S.; Hayashi, N.; Nakamura, T.; Fujimiya, M. Changes in serotonin levels and 5-HT receptor activity in duodenum of streptozotocin-diabetic rats. Am. J. Physiol. Liver Physiol. 2001, 281, G798–G808. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.B.; Witte, A.B. The role of serotonin in intestinal luminal sensing and secretion. Acta Physiol. 2008, 193, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Naruse, S.; Kitagawa, M.; Ishiguro, H.; Yoshikawa, T.; Ko, S.B.; Yamamoto, A.; Hamada, H.; Hayakawa, T. 5-hydroxytryptamine strongly inhibits fluid secretion in Guinea pig pancreatic duct cells. J. Clin. Investig. 2001, 108, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.G.; E Foxx-Orenstein, A.; Grider, J.R. Propulsion in Guinea pig colon induced by 5-hydroxytryptamine (HT) via 5-HT4 and 5-HT3 receptors. J. Pharmacol. Exp. Ther. 1999, 288, 93–97. [Google Scholar] [PubMed]
- Kendig, D.M.; Grider, J.R. Serotonin and colonic motility. Neuro Gastroenterol. Motil. 2015, 27, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Israelyan, N.; Del Colle, A.; Li, Z.; Park, Y.; Xing, A.; Jacobsen, J.P.R.; Luna, R.A.; Jensen, D.D.; Madra, M.; Saurman, V.; et al. Effects of Serotonin and Slow-Release 5-Hydroxytryptophan on Gastrointestinal Motility in a Mouse Model of Depression. Gastroenterology 2019, 157, 507–521. [Google Scholar] [CrossRef]
- Lesurtel, M.; Soll, C.; Graf, R.; Clavien, P.A. Role of serotonin in the hepato-gastrointestinal tract: An old molecule for new perspectives. Cell Mol. Life. Sci. 2008, 65, 940–952. [Google Scholar] [CrossRef]
- Jones, L.A.; Sun, E.W.; Martin, A.M.; Keating, D.J. The ever-changing roles of serotonin. Int. J. Biochem. Cell Biol. 2020, 125, 105776. [Google Scholar] [CrossRef]
- Siriwardena, A.; Kellum, J.M. A 5-HT2 receptor mediates serotonin-induced electrolyte transport in rat left colon. J. Surg. Res. 1993, 55, 323–329. [Google Scholar] [CrossRef]
- Gatopoulou, A.; Papanas, N.; Maltezos, E. Diabetic gastrointestinal autonomic neuropathy: Current status and new achievements for everyday clinical practice. Eur. J. Intern. Med. 2012, 23, 499–505. [Google Scholar] [CrossRef]
- Gonkowski, S. Bisphenol A (BPA)-Induced Changes in the Number of Serotonin-Positive Cells in the Mucosal Layer of Porcine Small Intestine-the Preliminary Studies. Int. J. Mol. Sci. 2020, 21, 1079. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 44, 129–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. Mechanisms of disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Matsuzawa, D.; Ishii, D.; Tomizawa, H.; Sajiki, J.; Shimizu, E. Perinatal exposure to bisphenol A enhances contextual fear memory and acts the serotoninergic system in juvenile female mice. Horm. Behav. 2013, 63, 709–716. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulc, M.; Palus, K.; Całka, J.; Kosacka, J.; Nowicki, M. Streptozotocin-Induced Diabetes Causes Changes in Serotonin-Positive Neurons in the Small Intestine in Pig Model. Int. J. Mol. Sci. 2022, 23, 4564. https://doi.org/10.3390/ijms23094564
Bulc M, Palus K, Całka J, Kosacka J, Nowicki M. Streptozotocin-Induced Diabetes Causes Changes in Serotonin-Positive Neurons in the Small Intestine in Pig Model. International Journal of Molecular Sciences. 2022; 23(9):4564. https://doi.org/10.3390/ijms23094564
Chicago/Turabian StyleBulc, Michał, Katarzyna Palus, Jarosław Całka, Joanna Kosacka, and Marcin Nowicki. 2022. "Streptozotocin-Induced Diabetes Causes Changes in Serotonin-Positive Neurons in the Small Intestine in Pig Model" International Journal of Molecular Sciences 23, no. 9: 4564. https://doi.org/10.3390/ijms23094564
APA StyleBulc, M., Palus, K., Całka, J., Kosacka, J., & Nowicki, M. (2022). Streptozotocin-Induced Diabetes Causes Changes in Serotonin-Positive Neurons in the Small Intestine in Pig Model. International Journal of Molecular Sciences, 23(9), 4564. https://doi.org/10.3390/ijms23094564