
Biometal Dyshomeostasis in Olfactory Mucosa of Alzheimer’s Disease Patients

 , , , ,
, , , , 
 and
and
Abstract
:1. Introduction
2. Results
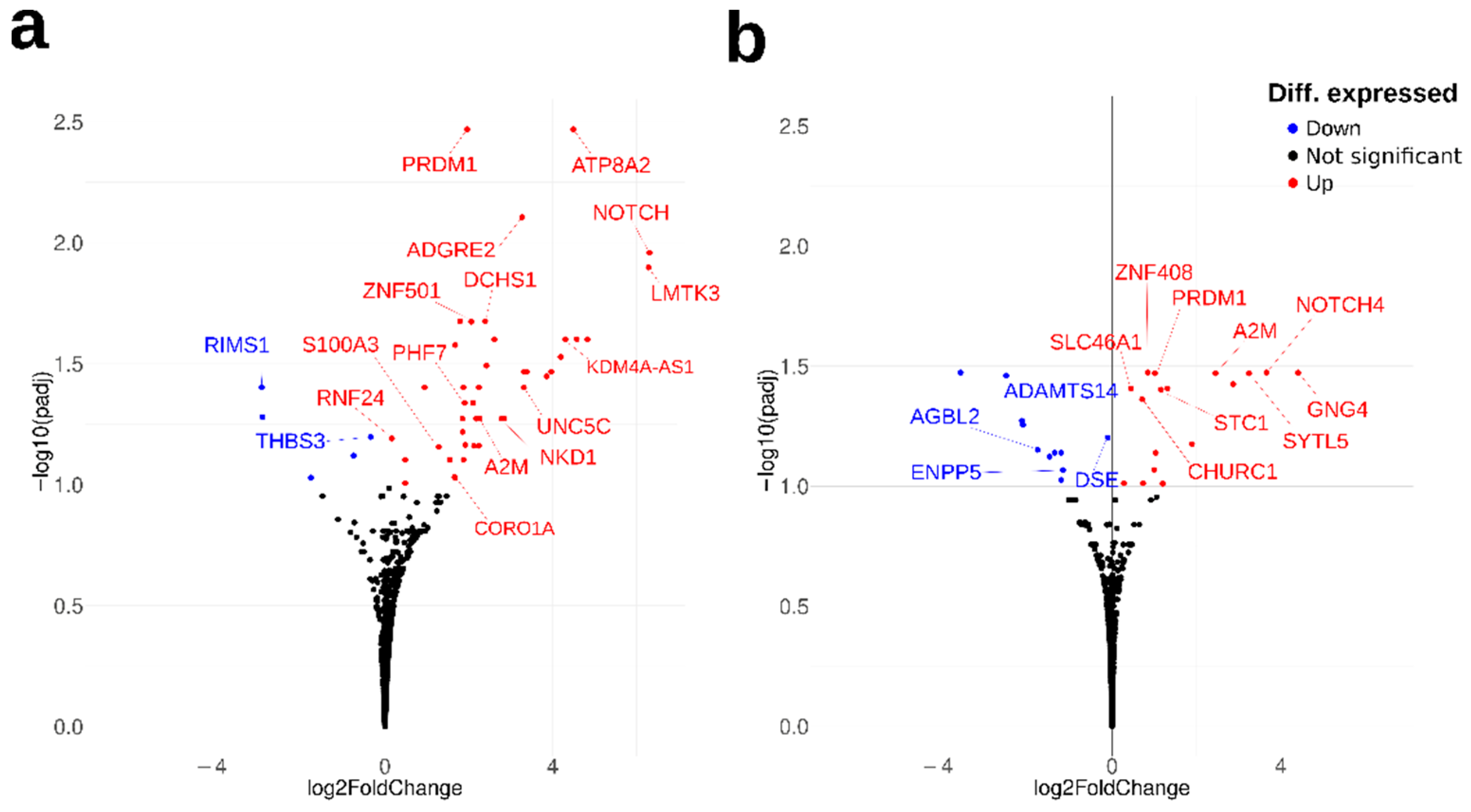
2.1. Transcriptomic Analysis of Metal-Related Gene Alterations in AD OM Cells
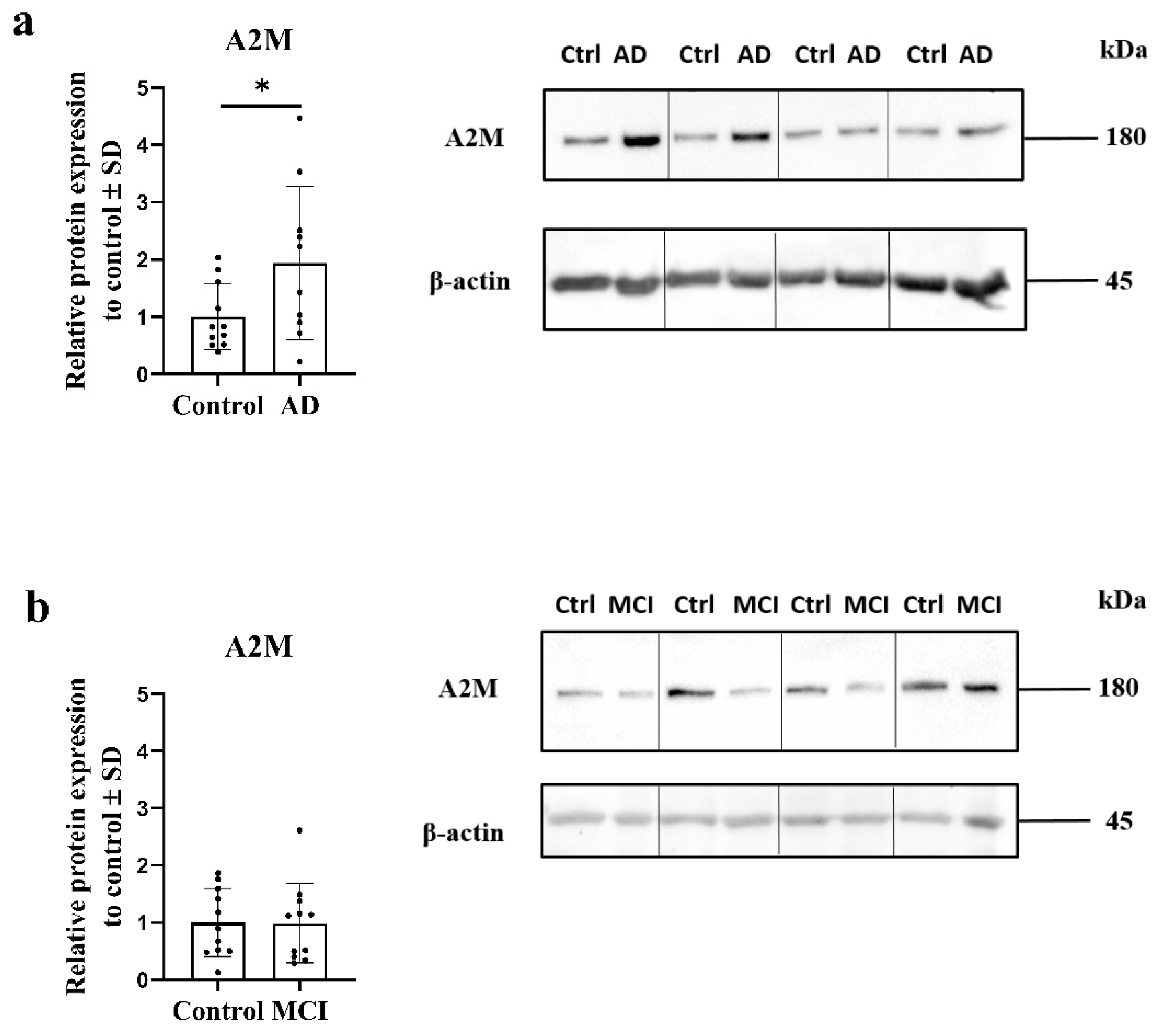
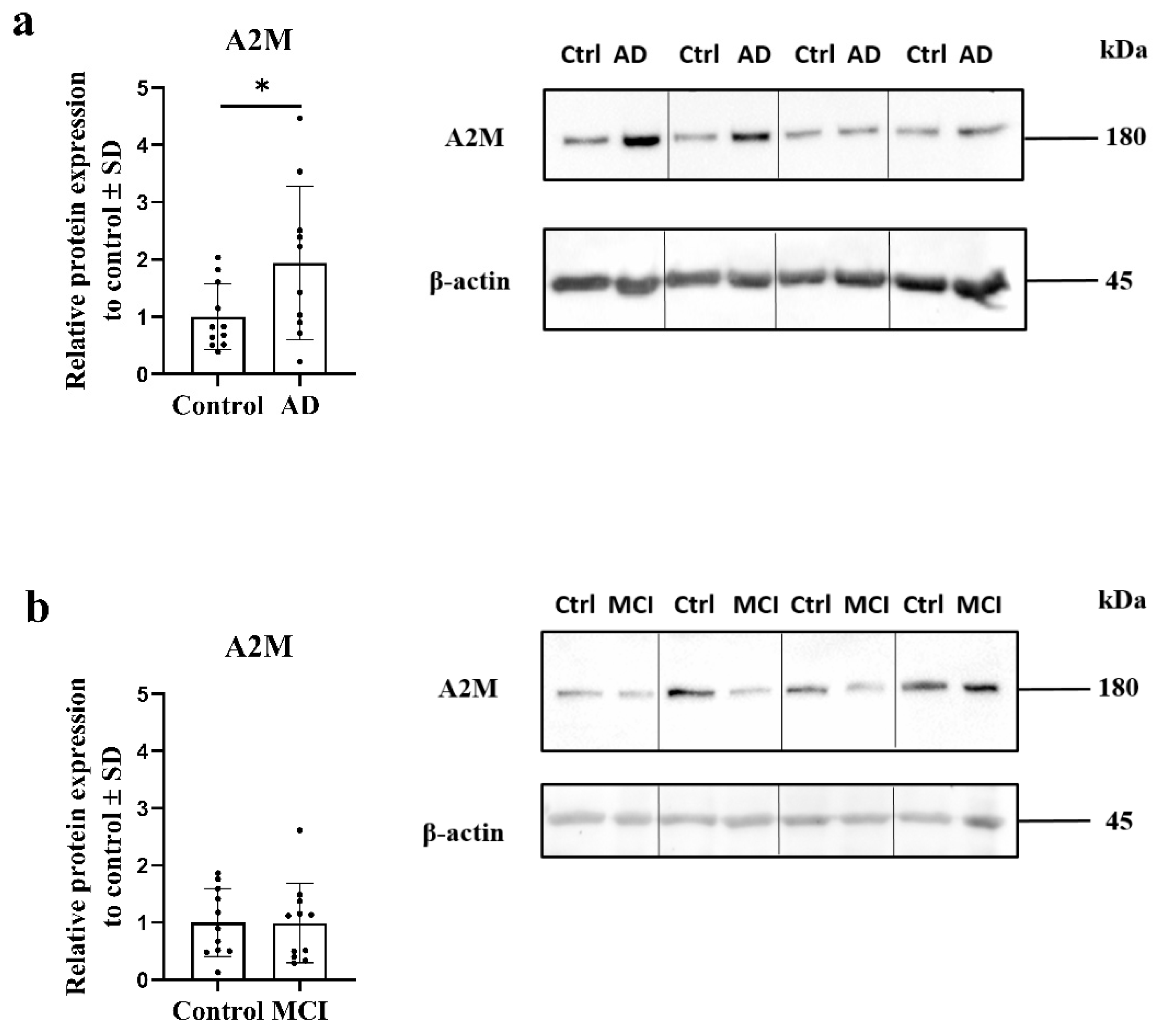
2.2. Expression of Alpha-2-Macroglobulin Is Altered in AD OM Cells
2.3. Elemental Content in AD OM Cells
3. Discussion
4. Materials and Methods
4.1. Patients and Culture of OM Cells
4.2. RNA Extraction and RNA Sequencing
4.3. RNA Sequencing Data Processing and Analysis
4.4. Protein Extraction and Western Blotting
4.5. Metal Quantitation via ICP-MS
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Study Group | n | Age * | Sex | Other Diseases |
|---|---|---|---|---|
| Controls | 10 | 71.1 ± 3.8 | Females 70.0% Males 30.0% | Cardiovascular 100.0% Thyroid 30.0% Respiratory 10.0% Autoimmune 20.0% Diabetes 0.0% |
| AD | 12 | 68.3 ± 7.4 | Females 50.0% Males 50.0% | Cardiovascular 66.7% Thyroid 8.3% Respiratory 0.0% Autoimmune 0.0% Diabetes 33.3% |
| MCI | 11 | 70.9 ± 6.4 | Females 45.5% Males 54.5% | Cardiovascular 63.6% Thyroid 9.1% Respiratory 9.1% Autoimmune 27.3% Diabetes 18.2% |
| Study Group | n | Age * | Sex |
|---|---|---|---|
| Controls | 11 | 69.1 ± 3.2 | Females 63.6% |
| Males 36.4% | |||
| AD | 10 | 68.7 ± 7.7 | Females 60.0% |
| Males 40.0% |
References
- Scheltens, P.; de Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s Disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s Disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Jack, C.R.; Knopman, D.S.; Jagust, W.J.; Shaw, L.M.; Aisen, P.S.; Weiner, M.W.; Petersen, R.C.; Trojanowski, J.Q. Hypothetical Model of Dynamic Biomarkers of the Alzheimer’s Pathological Cascade. Lancet Neurol. 2010, 9, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, H.R.; Bates, K.A.; Rodrigues, M.; Taddei, K.; Laws, S.M.; Lautenschlager, N.T.; Dhaliwal, S.S.; Johnston, A.N.B.; MacKay-Sim, A.; Gandy, S.; et al. Olfactory Dysfunction Is Associated with Subjective Memory Complaints in Community-Dwelling Elderly Individuals. J. Alzheimers Dis. 2009, 17, 135–142. [Google Scholar] [CrossRef]
- Sohrabi, H.R.; Bates, K.A.; Weinborn, M.G.; Johnston, A.N.B.; Bahramian, A.; Taddei, K.; Laws, S.M.; Rodrigues, M.; Morici, M.; Howard, M.; et al. Olfactory Discrimination Predicts Cognitive Decline among Community-Dwelling Older Adults. Transl. Psychiatry 2012, 2, e118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.J.; Shin, I.S.; Lee, J.E. Olfactory Function in Mild Cognitive Impairment and Alzheimer’s Disease: A Meta-Analysis. Laryngoscope 2019, 129, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Lampinen, R.; Feroze Fazaludeen, M.; Avesani, S.; Örd, T.; Penttilä, E.; Lehtola, J.-M.; Saari, T.; Hannonen, S.; Saveleva, L.; Kaartinen, E.; et al. Single-Cell RNA-Seq Analysis of Olfactory Mucosal Cells of Alzheimer’s Disease Patients. Cells 2022, 11, 676. [Google Scholar] [CrossRef] [PubMed]
- Fasae, K.D.; Abolaji, A.O.; Faloye, T.R.; Odunsi, A.Y.; Oyetayo, B.O.; Enya, J.I.; Rotimi, J.A.; Akinyemi, R.O.; Whitworth, A.J.; Aschner, M. Metallobiology and Therapeutic Chelation of Biometals (Copper, Zinc and Iron) in Alzheimer’s Disease: Limitations, and Current and Future Perspectives. J. Trace Elem. Med. Biol. 2021, 67, 126779. [Google Scholar] [CrossRef]
- Samudralwar, D.L.; Diprete, C.C.; Ni, B.F.; Ehmann, W.D.; Markesbery, W.R. Elemental Imbalances in the Olfactory Pathway in Alzheimer’s Disease. J. Neurol. Sci. 1995, 130, 139–145. [Google Scholar] [CrossRef]
- Ono, S.I.; Cherian, G.M. Regional Distribution of Metallothionein, Zinc, and Copper in the Brain of Different Strains of Rats. Biol. Trace Elem. Res. 1999, 69, 151–159. [Google Scholar] [CrossRef]
- Sastre, M.; Ritchie, C.W.; Hajji, N. Metal Ions in Alzheimer’s Disease Brain. JSM Alzheimers Dis. Related Dementia 2015, 2, 1014. [Google Scholar]
- Tamano, H.; Takeda, A. Age-Dependent Modification of Intracellular Zn2+ Buffering in the Hippocampus and Its Impact. Biol. Pharm. Bull. 2019, 42, 1070–1075. [Google Scholar] [CrossRef] [Green Version]
- Zaręba, N.; Kepinska, M. The Function of Transthyretin Complexes with Metallothionein in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 9003. [Google Scholar] [CrossRef]
- Wood, J.A.; Wood, P.L.; Ryan, R.; Graff-Radford, N.R.; Pilapil, C.; Robitaille, Y.; Quirion, R. Cytokine Indices in Alzheimer’s Temporal Cortex: No Changes in Mature IL-1 Beta or IL-1RA but Increases in the Associated Acute Phase Proteins IL-6, Alpha 2-Macroglobulin and C-Reactive Protein. Brain Res. 1993, 629, 245–252. [Google Scholar] [CrossRef]
- Kiddle, S.J.; Sattlecker, M.; Proitsi, P.; Simmons, A.; Westman, E.; Bazenet, C.; Nelson, S.K.; Williams, S.; Hodges, A.; Johnston, C.; et al. Candidate Blood Proteome Markers of Alzheimer’s Disease Onset and Progression: A Systematic Review and Replication Study. J. Alzheimers Dis. 2014, 38, 515–531. [Google Scholar] [CrossRef] [Green Version]
- Ulgen, E.; Ozisik, O.; Sezerman, O.U. PathfindR: An R Package for Comprehensive Identification of Enriched Pathways in Omics Data through Active Subnetworks. Front. Genet. 2019, 10, 858. [Google Scholar] [CrossRef]
- Shibata, N.; Ohnuma, T.; Higashi, S.; Higashi, M.; Usui, C.; Ohkubo, T.; Watanabe, T.; Kawashima, R.; Kitajima, A.; Ueki, A.; et al. Genetic Association between Notch4 Polymorphisms and Alzheimer’s Disease in the Japanese Population. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 350–351. [Google Scholar] [CrossRef]
- Kapoor, A.; Nation, D.A. Role of Notch Signaling in Neurovascular Aging and Alzheimer’s Disease. Semin. Cell Dev. Biol. 2021, 116, 90–97. [Google Scholar] [CrossRef]
- Seyfried, N.T.; Dammer, E.B.; Swarup, V.; Geschwind, D.H.; Lah, J.J.; Levey, A.I. A Multi-Network Approach Identifies Protein-Specific Co-Expression in Asymptomatic and Symptomatic Alzheimer’s Disease. Cell Syst. 2017, 4, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Beak, S.H.; Charidimou, A.; Song, M. Discovering New Genes in the Pathways of Common Sporadic Neurodegenerative Diseases: A Bioinformatics Approach. J. Alzheimers Dis. 2016, 51, 293–312. [Google Scholar] [CrossRef]
- Hondius, D.C.; van Nierop, P.; Li, K.W.; Hoozemans, J.J.M.; van der Schors, R.C.; van Haastert, E.S.; van der Vies, S.M.; Rozemuller, A.J.M.; Smit, A.B. Profiling the Human Hippocampal Proteome at All Pathologic Stages of Alzheimer’s Disease. Alzheimers Dement. 2016, 12, 654–668. [Google Scholar] [CrossRef]
- Huat, T.J.; Camats-Perna, J.; Newcombe, E.A.; Valmas, N.; Kitazawa, M.; Medeiros, R. Metal Toxicity Links to Alzheimer’s Disease and Neuroinflammation. J. Mol. Biol. 2019, 431, 1843–1868. [Google Scholar] [CrossRef]
- Burnet, F.M. A Possible Role of Zinc in the Pathology of Dementia. Lancet 1981, 317, 186–188. [Google Scholar] [CrossRef]
- Witt, B.; Schaumlöffel, D.; Schwerdtle, T. Subcellular Localization of Copper-Cellular Bioimaging with Focus on Neurological Disorders. Int. J. Mol. Sci. 2020, 21, 2341. [Google Scholar] [CrossRef] [Green Version]
- Lindeque, J.Z.; Levanets, O.; Louw, R.; van der Westhuizen, F.H. The Involvement of Metallothioneins in Mitochondrial Function and Disease. Curr. Protein Pept. Sci. 2010, 11, 292–309. [Google Scholar] [CrossRef] [Green Version]
- Huiliang, Z.; Mengzhe, Y.; Xiaochuan, W.; Hui, W.; Min, D.; Mengqi, W.; Jianzhi, W.; Zhongshan, C.; Caixia, P.; Rong, L. Zinc Induces Reactive Astrogliosis through ERK-Dependent Activation of Stat3 and Promotes Synaptic Degeneration. J. Neurochem. 2021, 159, 1016–1027. [Google Scholar] [CrossRef]
- Lengyel, I.; Flinn, J.M.; Peto, T.; Linkous, D.H.; Cano, K.; Bird, A.C.; Lanzirotti, A.; Frederickson, C.J.; van Kuijk, F.J.G.M. High Concentration of Zinc in Sub-Retinal Pigment Epithelial Deposits. Exp. Eye Res. 2007, 84, 772–780. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Salminen, A.; Haapasalo, A.; Soininen, H.; Hiltunen, M. Age-Related Macular Degeneration (AMD): Alzheimer’s Disease in the Eye? J. Alzheimers Dis. 2011, 24, 615–631. [Google Scholar] [CrossRef] [Green Version]
- LaFerla, F.M. Calcium Dyshomeostasis and Intracellular Signalling in Alzheimer’s Disease. Nat. Rev. Neurosci. 2002, 3, 862–872. [Google Scholar] [CrossRef]
- Block, E.; Batista, V.S.; Matsunami, H.; Zhuang, H.; Ahmed, L. The Role of Metals in Mammalian Olfaction of Low Molecular Weight Organosulfur Compounds. Nat. Prod. Rep. 2017, 34, 529–557. [Google Scholar] [CrossRef] [Green Version]
- Sunderman, F.W., Jr. Nasal Toxicity, Carcinogenicity, and Olfactory Uptake of Metals. Ann. Clin. Lab. Sci. 2001, 31, 3–24. [Google Scholar] [PubMed]
- McBride, K.; Slotnick, B.; Margolis, F.L. Does Intranasal Application of Zinc Sulfate Produce Anosmia in the Mouse? An Olfactometric and Anatomical Study. Chem. Senses 2003, 28, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, T.M.; Smith, W.M. The Bradford Hill Criteria and Zinc-Induced Anosmia: A Causality Analysis. Arch. Otolaryngol. Head Neck Surg. 2010, 136, 673–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitcroft, K.L.; Ezzat, M.; Cuevas, M.; Andrews, P.; Hummel, T. The Effect of Intranasal Sodium Citrate on Olfaction in Post-Infectious Loss: Results from a Prospective, Placebo-Controlled Trial in 49 Patients. Clin. Otolaryngol. 2017, 42, 557–563. [Google Scholar] [CrossRef]
- Bush, A.I.; Pettingell, W.H.; Multhaup, G.; Paradis, M.D.; Vonsattel, J.P.; Gusella, J.F.; Beyreuther, K.; Masters, C.L.; Tanzi, R.E. Rapid Induction of Alzheimer A Beta Amyloid Formation by Zinc. Science 1994, 265, 1464–1467. [Google Scholar] [CrossRef]
- Wang, L.; Yin, Y.L.; Liu, X.Z.; Shen, P.; Zheng, Y.G.; Lan, X.R.; Lu, C.B.; Wang, J.Z. Current Understanding of Metal Ions in the Pathogenesis of Alzheimer’s Disease. Transl. Neurodegener. 2020, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Wang, T.; Zheng, W.; Zhao, B.L.; Danscher, G.; Chen, Y.H.; Wang, Z.Y. Zinc Overload Enhances APP Cleavage and Aβ Deposition in the Alzheimer Mouse Brain. PLoS ONE 2010, 5, e15349. [Google Scholar] [CrossRef]
- Ryan, K.C.; Ashkavand, Z.; Norman, K.R. The Role of Mitochondrial Calcium Homeostasis in Alzheimer’s and Related Diseases. Int. J. Mol. Sci. 2020, 21, 9153. [Google Scholar] [CrossRef]
- Tooyama, I.; Kawamata, T.; Akiyama, H.; Moestrup, S.K.; Gliemann, J.; McGeer, P.L. Immunohistochemical Study of Alpha 2 Macroglobulin Receptor in Alzheimer and Control Postmortem Human Brain. Mol. Chem. Neuropathol. 1993, 18, 153–160. [Google Scholar] [CrossRef]
- Du, Y.; Ni, B.; Glinn, M.; Dodel, R.C.; Bales, K.R.; Zhang, Z.; Hyslop, P.A.; Paul, S.M. Alpha2-Macroglobulin as a Beta-Amyloid Peptide-Binding Plasma Protein. J. Neurochem. 1997, 69, 299–305. [Google Scholar] [CrossRef]
- Narita, M.; Holtzman, D.M.; Schwartz, A.L.; Bu, G. Alpha2-Macroglobulin Complexes with and Mediates the Endocytosis of Beta-Amyloid Peptide via Cell Surface Low-Density Lipoprotein Receptor-Related Protein. J. Neurochem. 1997, 69, 1904–1911. [Google Scholar] [CrossRef]
- Blacker, D.; Wilcox, M.A.; Laird, N.M.; Rodes, L.; Horvath, S.M.; Go, R.C.P.; Perry, R.; Watson, B.; Bassett, S.S.; McInnis, M.G.; et al. Alpha-2 Macroglobulin Is Genetically Associated with Alzheimer Disease. Nat. Genet. 1998, 19, 357–360. [Google Scholar] [CrossRef]
- Westwood, S.; Leoni, E.; Hye, A.; Lynham, S.; Khondoker, M.R.; Ashton, N.J.; Kiddle, S.J.; Baird, A.L.; Sainz-Fuertes, R.; Leung, R.; et al. Blood-Based Biomarker Candidates of Cerebral Amyloid Using PiB PET in Non-Demented Elderly. J. Alzheimers Dis. 2016, 52, 561–572. [Google Scholar] [CrossRef]
- Eke, C.S.; Jammeh, E.; Li, X.; Carroll, C.; Pearson, S.; Ifeachor, E. Early Detection of Alzheimer’s Disease with Blood Plasma Proteins Using Support Vector Machines. IEEE J. Biomed. Health Inform. 2021, 25, 218–226. [Google Scholar] [CrossRef]
- Poller, W.; Barth, J.; Voss, B. Detection of an Alteration of the Alpha 2-Macroglobulin Gene in a Patient with Chronic Lung Disease and Serum Alpha 2-Macroglobulin Deficiency. Hum. Genet. 1989, 83, 93–96. [Google Scholar] [CrossRef]
- Ho, A.S.; Cheng, C.C.; Lee, S.C.; Liu, M.L.; Lee, J.Y.; Wang, W.M.; Wang, C.C. Novel Biomarkers Predict Liver Fibrosis in Hepatitis C Patients: Alpha 2 Macroglobulin, Vitamin D Binding Protein and Apolipoprotein AI. J. Biomed. Sci. 2010, 17, 58. [Google Scholar] [CrossRef] [Green Version]
- Tripodi, A.; Chantarangkul, V.; de Stefano, V.; Mannucci, P. Alpha(2)-Macroglobulin Levels Are High in Adult Patients with Congenital Antithrombin Deficiency. Thromb. Res. 2000, 98, 117–122. [Google Scholar] [CrossRef]
- Kanoh, Y.; Ohtani, N.; Mashiko, T.; Ohtani, S.; Nishikawa, T.; Egawa, S.; Baba, S.; Ohtani, H. Levels of Alpha 2 Macroglobulin Can Predict Bone Metastases in Prostate Cancer. Anticancer Res. 2001, 21, 551–556. [Google Scholar]
- El Haj, M.; Gandolphe, M.C.; Gallouj, K.; Kapogiannis, D.; Antoine, P. From Nose to Memory: The Involuntary Nature of Odor-Evoked Autobiographical Memories in Alzheimer’s Disease. Chem. Senses 2018, 43, 27–34. [Google Scholar] [CrossRef] [Green Version]
- O’Bryant, S.E.; Lacritz, L.H.; Hall, J.; Waring, S.C.; Chan, W.; Khodr, Z.G.; Massman, P.J.; Hobson, V.; Cullum, C.M. Validation of the New Interpretive Guidelines for the Clinical Dementia Rating Scale Sum of Boxes Score in the National Alzheimer’s Coordinating Center Database. Arch. Neurol. 2010, 67, 746–749. [Google Scholar] [CrossRef]
- Welsh, K.A.; Butters, N.; Hughes, J.P.; Mohs, R.C.; Heyman, A. Detection and Staging of Dementia in Alzheimer’s Disease. Use of the Neuropsychological Measures Developed for the Consortium to Establish a Registry for Alzheimer’s Disease. Arch. Neurol. 1992, 49, 448–452. [Google Scholar] [CrossRef]
- Chandler, M.J.; Lacritz, L.H.; Hynan, L.S.; Barnard, H.D.; Allen, G.; Deschner, M.; Weiner, M.F.; Cullum, C.M. A Total Score for the CERAD Neuropsychological Battery. Neurology 2005, 65, 102–106. [Google Scholar] [CrossRef]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The Diagnosis of Dementia Due to Alzheimer’s Disease: Recommendations from the National Institute on Aging-Alzheimer’s Association Workgroups on Diagnostic Guidelines for Alzheimer’s Disease. Alzheimers Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Chew, S.; Lampinen, R.; Saveleva, L.; Korhonen, P.; Mikhailov, N.; Grubman, A.; Grubman, A.; Grubman, A.; Polo, J.M.; Polo, J.M.; et al. Urban Air Particulate Matter Induces Mitochondrial Dysfunction in Human Olfactory Mucosal Cells. Part. Fibre Toxicol. 2020, 17, 18. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Frankish, A.; Diekhans, M.; Jungreis, I.; Lagarde, J.; Loveland, J.E.; Mudge, J.M.; Sisu, C.; Wright, J.C.; Armstrong, J.; Barnes, I.; et al. GENCODE 2021. Nucleic Acids Res. 2021, 49, D916–D923. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread Aligner: Fast, Accurate and Scalable Read Mapping by Seed-and-Vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating High-Throughput Genomic Analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Stephens, M. False Discovery Rates: A New Deal. Biostatistics 2017, 18, 275. [Google Scholar] [CrossRef] [Green Version]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Carbon, S.; Douglass, E.; Good, B.M.; Unni, D.R.; Harris, N.L.; Mungall, C.J.; Basu, S.; Chisholm, R.L.; Dodson, R.J.; Hartline, E.; et al. The Gene Ontology Resource: Enriching a GOld Mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar]
- Giurgiu, M.; Reinhard, J.; Brauner, B.; Dunger-Kaltenbach, I.; Fobo, G.; Frishman, G.; Montrone, C.; Ruepp, A. CORUM: The Comprehensive Resource of Mammalian Protein Complexes-2019. Nucleic Acids Res. 2019, 47, D559–D563. [Google Scholar] [CrossRef] [Green Version]
- Kanninen, K.M.; Grubman, A.; Meyerowitz, J.; Duncan, C.; Tan, J.L.; Parker, S.J.; Crouch, P.J.; Paterson, B.M.; Hickey, J.L.; Donnelly, P.S.; et al. Increased Zinc and Manganese in Parallel with Neurodegeneration, Synaptic Protein Changes and Activation of Akt/GSK3 Signaling in Ovine CLN6 Neuronal Ceroid Lipofuscinosis. PLoS ONE 2013, 8, e58644. [Google Scholar] [CrossRef] [Green Version]
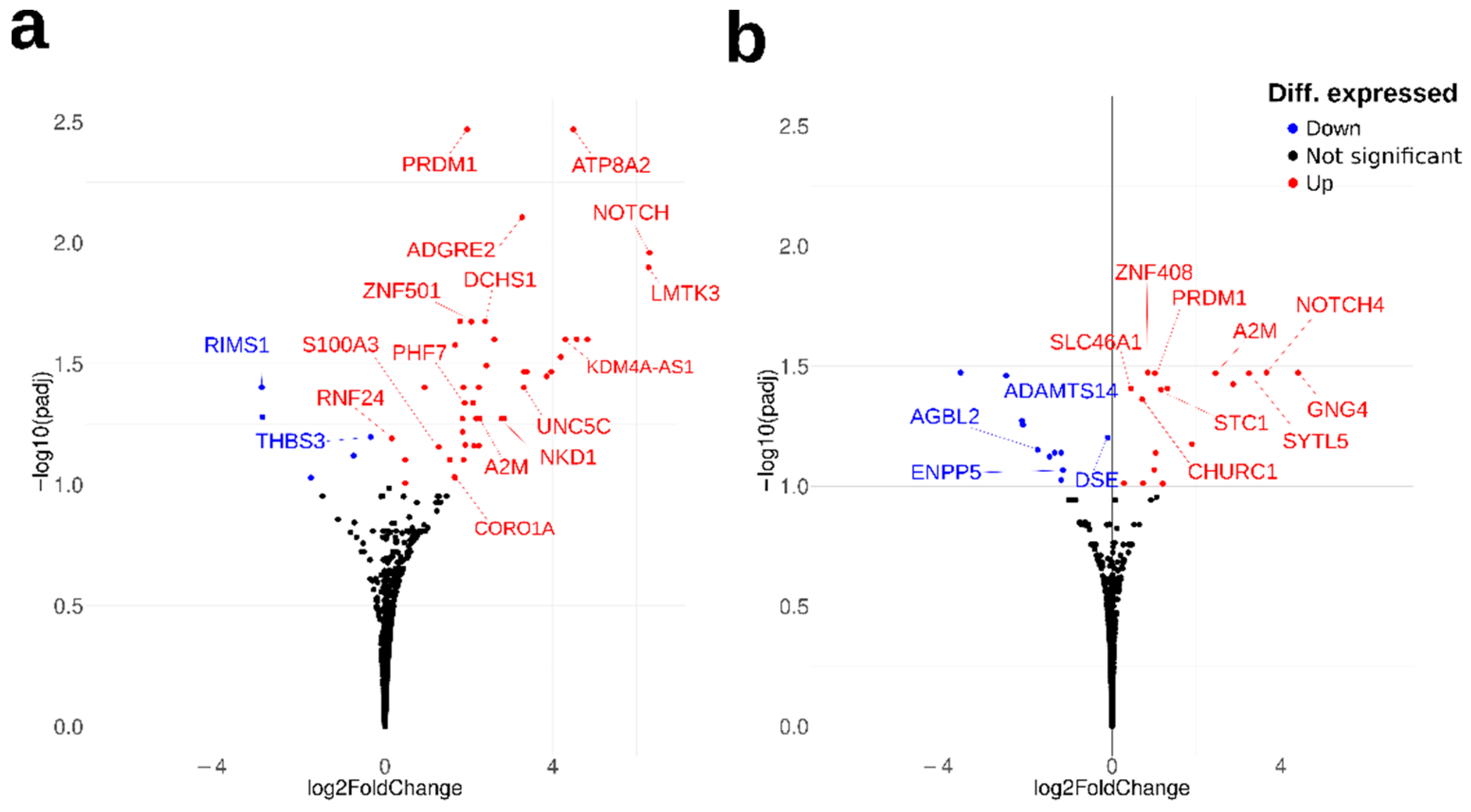
| Gene Symbol | Gene Name | Log2FC | p-Value | padj-Value | Metal-Binding/Metal-Related Function 1 |
|---|---|---|---|---|---|
| NOTCH4 | notch receptor 4 | 6.31 | 2.72 × 10−6 | 0.0110 | metal-binding; Ca |
| LMTK3 | lemur tyrosine kinase 3 | 6.29 | 3.90 × 10−6- | 0.0127 | metal-binding; Mg |
| ATP8A2 | ATPase phospholipid transporting 8A2 | 4.49 | 4.19 × 10−7 | 0.0034 | metal-binding; Mg |
| KDM4A-AS1 | KDM4A antisense RNA 1 | 4.30 | 1.85 × 10−5 | 0.0251 | for KDM4A: metal-binding; Fe, Zn |
| ADGRE2 | adhesion G protein-coupled receptor E2 | 3.27 | 1.45 × 10−6 | 0.0079 | metal-binding; Ca |
| NKD1 | NKD inhibitor of WNT signaling pathway 1 | 2.78 | 9.71 × 10−5 | 0.0533 | metal-binding; Ca |
| DCHS1 | dachsous cadherin-related 1 | 2.39 | 8.83 × 10−6 | 0.0211 | metal-binding; Ca |
| A2M | alpha-2-macroglobulin | 2.26 | 1.02 × 10−4 | 0.0533 | Ca-dependent protein binding |
| ZNF501 | zinc finger protein 501 | 2.06 | 1.05 × 10−5 | 0.0212 | metal-binding; Zn |
| PRDM1 | PR/SET domain 1 | 1.96 | 3.17 × 10−7 | 0.0034 | metal-binding; Zn |
| PLS1 | plastin 1 | 1.92 | 1.52 × 10−4 | 0.0686 | metal-binding; Ca |
| PHF7 | PHD finger protein 7 | 1.90 | 7.37 × 10−5 | 0.0460 | metal-binding; Zn |
| CORO1A | coronin 1A | 1.67 | 2.66 × 10−4 | 0.0938 | calcium ion transport |
| S100A3 | S100 calcium binding protein A3 | 1.28 | 1.68 × 10−4 | 0.0698 | metal-binding; Ca, Zn |
| RNF24 | ring finger protein 24 | 0.17 | 1.39 × 10−4 | 0.0644 | metal-binding; Zn |
| THBS3 | thrombospondin 3 | −0.34 | 1.33 × 10−4 | 0.0635 | calcium ion binding |
| RIMS1 | regulating synaptic membrane exocytosis 1 | −2.94 | 5.49 × 10−5 | 0.0397 | metal-binding; Zn |
| Gene Symbol | Gene Name | Log2FC | p-Value | padj Value | Metal-Binding/Metal-Related Function 1 |
|---|---|---|---|---|---|
| GNG4 | G protein subunit gamma 4 | 4.42 | 2.71 × 10−6 | 0.0337 | pathway, Ca |
| NOTCH4 | notch receptor 4 | 3.66 | 8.31 × 10−6 | 0.0337 | metal-binding; Ca |
| SYTL5 | synaptotagmin like 5 | 3.25 | 1.27 × 10−5 | 0.0339 | metal-binding; Zn |
| A2M | alpha-2-macroglobulin | 2.45 | 1.47 × 10−5 | 0.0339 | Ca-dependent protein binding |
| STC1 | stanniocalcin 1 | 1.16 | 2.94 × 10−5 | 0.0397 | Ca homeostasis |
| PRDM1 | PR/SET domain | 1.02 | 1.07 × 10−5 | 0.0339 | metal-binding; Zn |
| HTR2B | 5-hydroxytryptamine receptor 2B | 1.00 | 1.24 × 10−4 | 0.0856 | Ca-mediated signaling |
| ZNF408 | zinc finger protein 408 | 0.85 | 6.13 × 10−6 | 0.0337 | metal-binding; Zn |
| CHURC1 | churchill domain containing 1 | 0.72 | 3.49 × 10−5 | 0.0435 | metal-binding; Zn |
| SLC46A1 | solute carrier family 46 member 1 | 0.45 | 2.66 × 10−5 | 0.0392 | iron homeostasis |
| DSE | dermatan sulfate epimerase | −0.10 | 6.20 × 10−5 | 0.0628 | metal-binding, Mn |
| ENPP5 | ectonucleotide pyrophosphatase/phosphodiesterase family member 5 | −1.17 | 1.27 × 10−4 | 0.0856 | metal-binding; Zn |
| ADAMTS14 | ADAM metallopeptidase with thrombospondin type 1 motif 14 | −1.21 | 9.42 × 10−5 | 0.0727 | metal-binding; Zn |
| AGBL2 | AGBL carboxypeptidase 2 | −1.77 | 7.85 × 10−5 | 0.0706 | metal-binding; Zn |
| Metal | Controls Mean ± SEM † | AD Mean ± SEM † | Unpaired t-Test (Two-Tailed) p-Value |
|---|---|---|---|
| 23Na | 7436 ± 450.7 | 11,600 ± 1005 | 0.0020 * |
| 24Mg | 13.29 ± 1.627 | 16.16 ± 1.553 | 0.2252 |
| 31P | 543.4 ± 45.32 | 629.9 ± 37.82 | 0.1716 |
| 39K | 6.645 ± 0.7977 | 7.674 ± 0.5793 | 0.3181 |
| 43Ca | 7.299 ± 0.754 | 10.02 ± 1.008 | 0.0416 * |
| 55Mn | 0.1171 ± 0.01115 | 0.1437 ± 0.01158 | 0.1153 |
| 56Fe | 1.718 ± 0.1663 | 2.047 ± 0.2251 | 0.2499 |
| 63Cu | 0.1836 ± 0.02722 | 0.3156 ± 0.08026 | 0.1218 |
| 66Zn | 1.899 ± 0.1417 | 2.385 ± 0.1570 | 0.0340 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lampinen, R.; Górová, V.; Avesani, S.; Liddell, J.R.; Penttilä, E.; Závodná, T.; Krejčík, Z.; Lehtola, J.-M.; Saari, T.; Kalapudas, J.; et al. Biometal Dyshomeostasis in Olfactory Mucosa of Alzheimer’s Disease Patients. Int. J. Mol. Sci. 2022, 23, 4123. https://doi.org/10.3390/ijms23084123
Lampinen R, Górová V, Avesani S, Liddell JR, Penttilä E, Závodná T, Krejčík Z, Lehtola J-M, Saari T, Kalapudas J, et al. Biometal Dyshomeostasis in Olfactory Mucosa of Alzheimer’s Disease Patients. International Journal of Molecular Sciences. 2022; 23(8):4123. https://doi.org/10.3390/ijms23084123
Chicago/Turabian StyleLampinen, Riikka, Veronika Górová, Simone Avesani, Jeffrey R. Liddell, Elina Penttilä, Táňa Závodná, Zdeněk Krejčík, Juha-Matti Lehtola, Toni Saari, Juho Kalapudas, and et al. 2022. "Biometal Dyshomeostasis in Olfactory Mucosa of Alzheimer’s Disease Patients" International Journal of Molecular Sciences 23, no. 8: 4123. https://doi.org/10.3390/ijms23084123
APA StyleLampinen, R., Górová, V., Avesani, S., Liddell, J. R., Penttilä, E., Závodná, T., Krejčík, Z., Lehtola, J.-M., Saari, T., Kalapudas, J., Hannonen, S., Löppönen, H., Topinka, J., Koivisto, A. M., White, A. R., Giugno, R., & Kanninen, K. M. (2022). Biometal Dyshomeostasis in Olfactory Mucosa of Alzheimer’s Disease Patients. International Journal of Molecular Sciences, 23(8), 4123. https://doi.org/10.3390/ijms23084123