
Dexamethasone Attenuates the Expression of MMP-13 in Chondrocytes through MKP-1

 and
and
Abstract
1. Introduction
2. Results
2.1. Dexamethasone Attenuates the Expression of Catabolic Enzymes in Chondrocytes from MKP-1 WT Mice, and This Effect Is Diminished in MKP-1 KO Mice
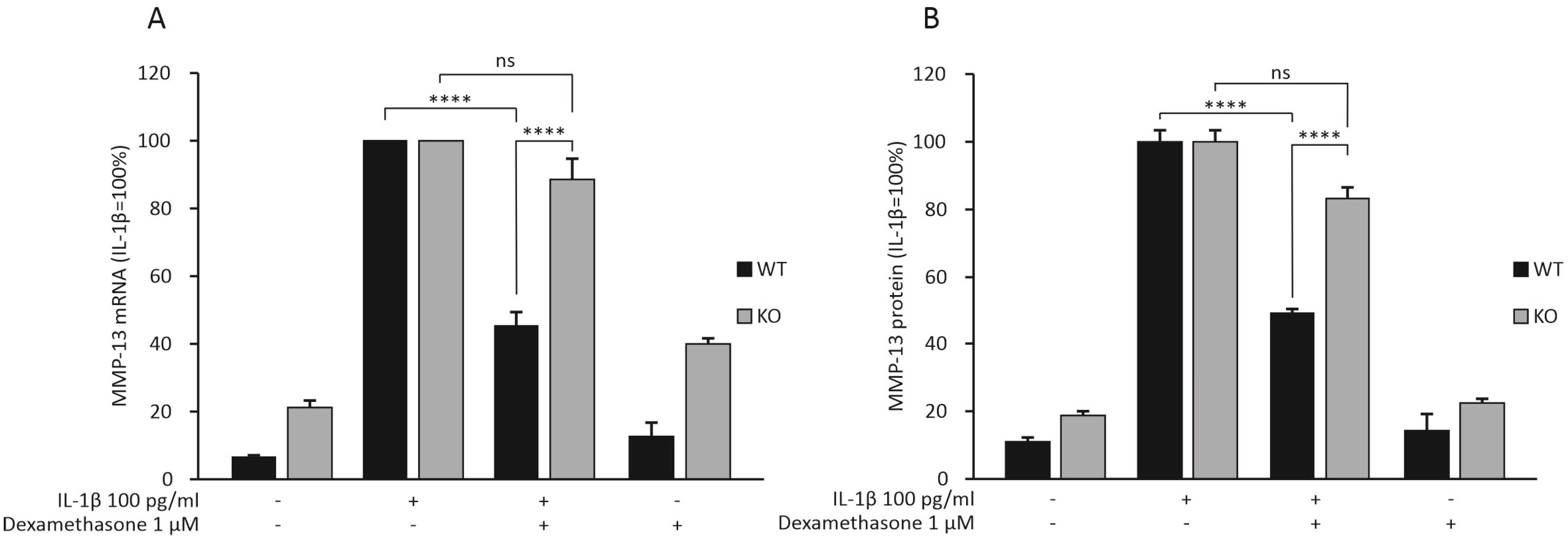
2.2. Dexamethasone Attenuates MMP-13 Expression in IL-1β-Stimulated Chondrocytes from WT Mice, but Not in Chondrocytes from MKP-1 KO Mice
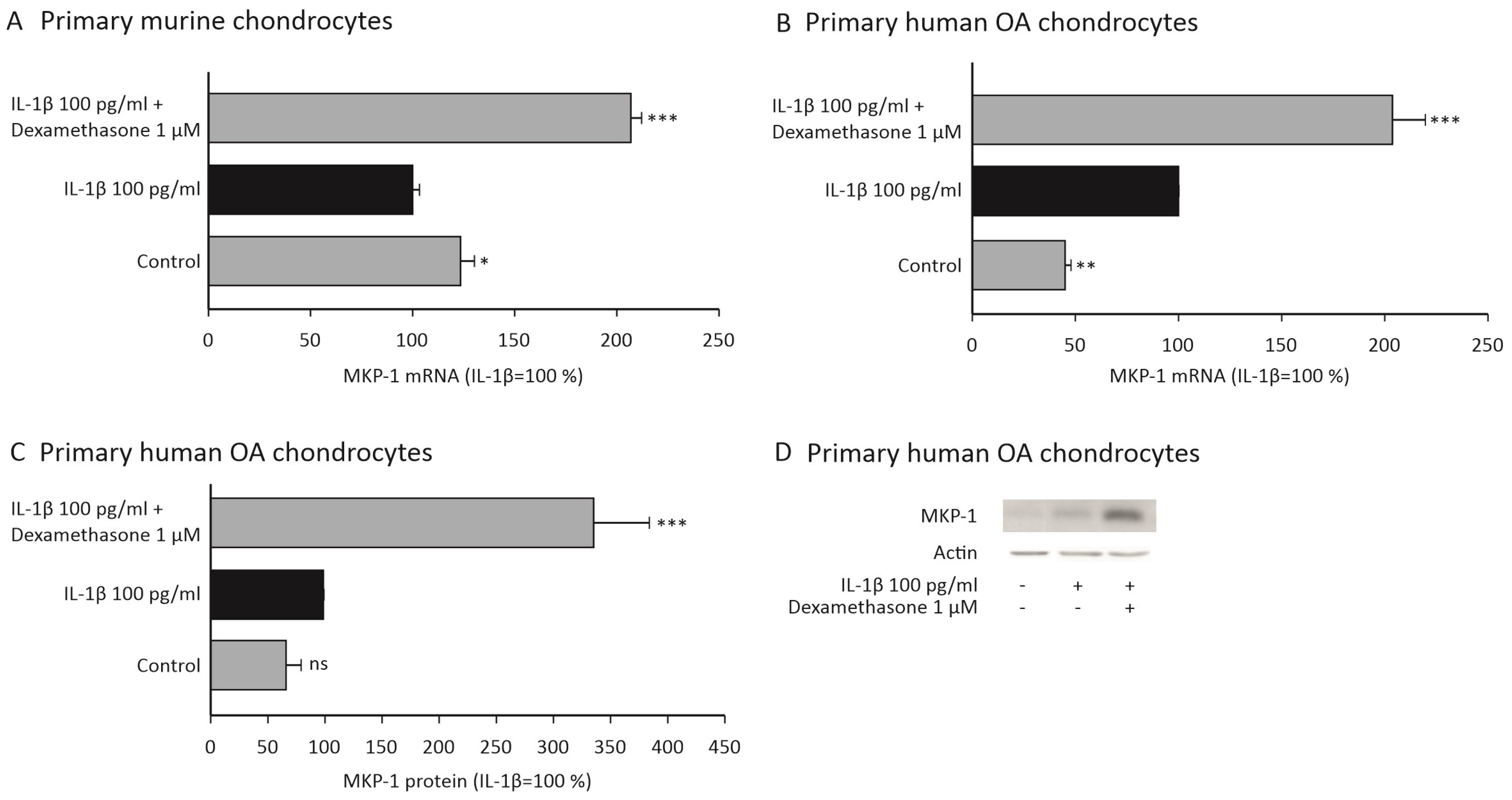
2.3. Dexamethasone Increases the Expression of MKP-1 in Chondrocytes
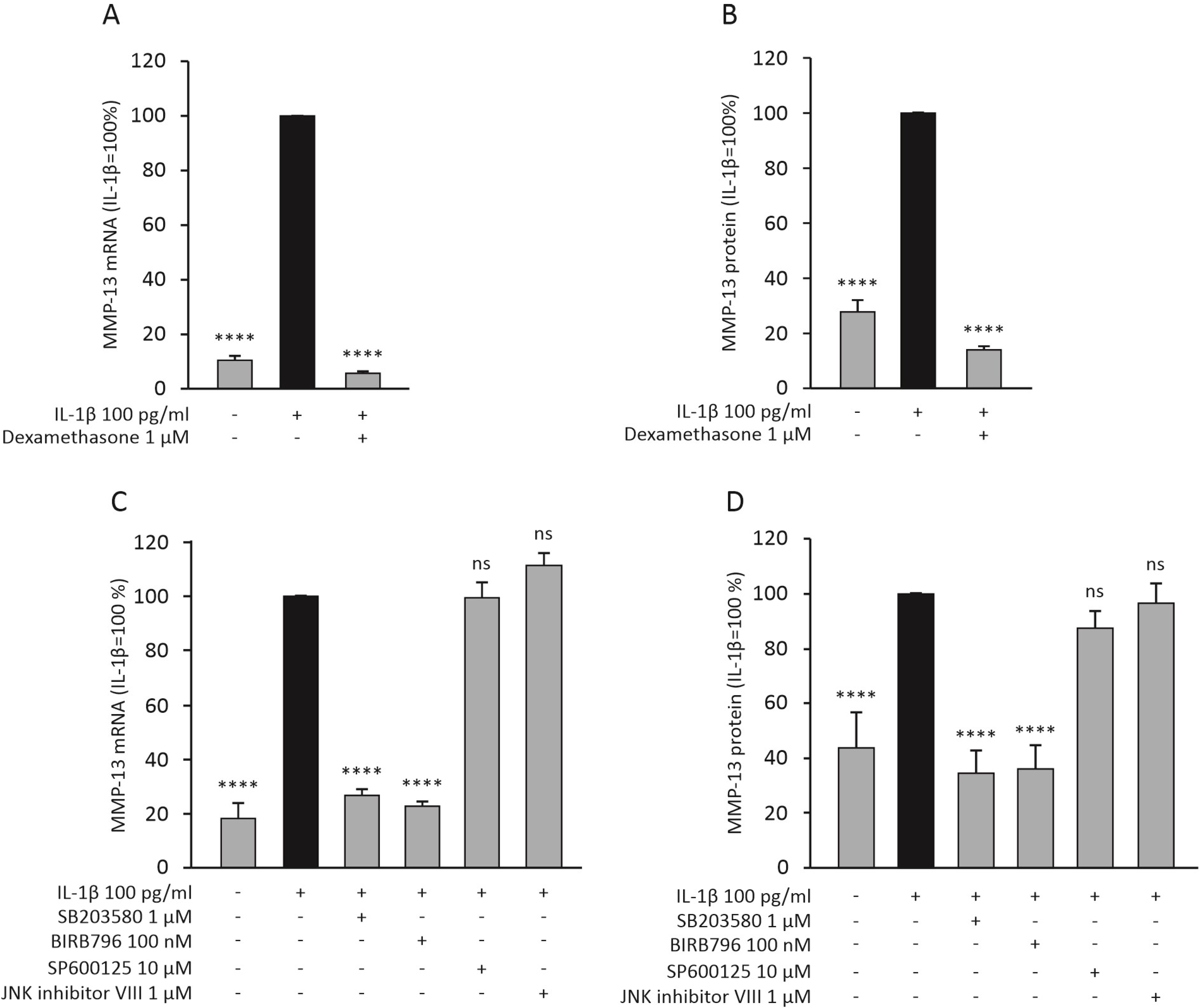
2.4. Dexamethasone and p38 MAP Kinase Inhibitors Attenuate the Expression of MMP-13 in Primary Human OA Chondrocytes
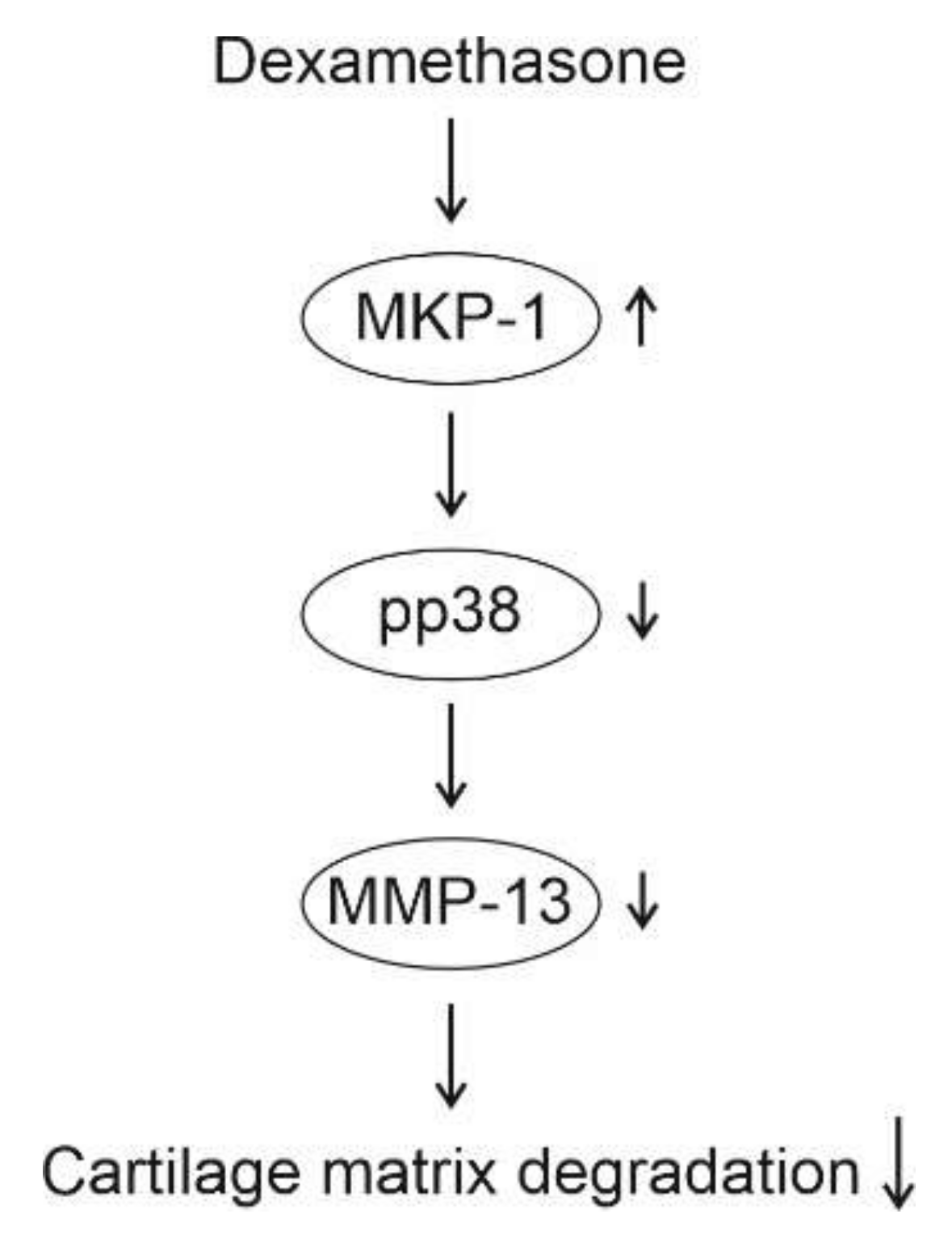
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Chondrocyte Isolation and Culture
4.3. Human OA Chondrocyte Isolation and Culture
4.4. RNA Isolation and Sample Preparation
4.5. Next-Generation Sequencing and Data Analysis
4.6. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.7. Enzyme-Linked Immunosorbent Assay
4.8. Western Blot
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| ELISA | enzyme-linked immunosorbent assay |
| FC | fold change |
| FDR | false discovery rate |
| IL | interleukin |
| KO | knockout |
| MAP kinase | mitogen-activated protein kinase |
| MKP-1 | mitogen-activated protein kinase phosphatase-1 |
| MMP | matrix metalloproteinase |
| OA | osteoarthritis |
| qRT-PCR | quantitative reverse transcription polymerase chain reaction |
| RNA-Seq | RNA sequencing |
| TNF | tumor necrosis factor |
| WT | wild-type |
References
- Chow, Y.Y.; Chin, K.-Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Rahmati, M.; Mobasheri, A.; Mozafari, M. Inflammatory mediators in osteoarthritis: A critical review of the state-of-the-art, current prospects, and future challenges. Bone 2016, 85, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef] [PubMed]
- Conaghan, P.G.; Cook, A.D.; Hamilton, J.A.; Tak, P.P. Therapeutic options for targeting inflammatory osteoarthritis pain. Nat. Rev. Rheumatol. 2019, 15, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef] [PubMed]
- Richter, D.L.; Schenck, R.C., Jr.; Wascher, D.C.; Treme, G. Knee Articular Cartilage Repair and Restoration Techniques: A Review of the Literature. Sports Health Multidiscip. Approach 2015, 8, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Szychlinska, M.A.; Calabrese, G.; Ravalli, S.; Dolcimascolo, A.; Castrogiovanni, P.; Fabbi, C.; Puglisi, C.; Lauretta, G.; Di Rosa, M.; Castorina, A.; et al. Evaluation of a Cell-Free Collagen Type I-Based Scaffold for Articular Cartilage Regeneration in an Orthotopic Rat Model. Materials 2020, 13, 2369. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lou, Z.; Zheng, S.; Wu, J.; Yao, Q.; Chen, R.; Kou, L.; Chen, D. Intra-articular drug delivery systems for osteoarthritis therapy: Shifting from sustained release to enhancing penetration into cartilage. Drug Deliv. 2022, 29, 767–791. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.-H.; Wu, C.-H.; Jou, I.-M.; Tu, Y.-K.; Hung, C.-H.; Hsieh, P.-L.; Tsai, K.-L. PKR activation causes inflammation and MMP-13 secretion in human degenerated articular chondrocytes. Redox Biol. 2018, 14, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zhang, H.-X.; Xing, M.-L.; Xu, Y.-B.; Li, P.; Huang, L.-K.; Bai, J.; Tian, Z.; Zhao, Z.-D. Knockdown of PRMT1 suppresses IL-1β-induced cartilage degradation and inflammatory responses in human chondrocytes through Gli1-mediated Hedgehog signaling pathway. Mol. Cell. Biochem. 2017, 438, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, L.; Lv, D.; Liu, H.; He, T.; Wang, X. Poly(ADP-ribose) polymerase 1 inhibition prevents interleukin-1β-induced inflammation in human osteoarthritic chondrocytes. Acta Biochim. Biophys. Sin. 2015, 47, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.-Z.; Yun, Z.; Wang, W.; Ma, B.-A. Dual specificity phosphatase 1 has a protective role in osteoarthritis fibroblast-like synoviocytes via inhibition of the MAPK signaling pathway. Mol. Med. Rep. 2017, 16, 8441–8447. [Google Scholar] [CrossRef]
- Kirk, S.G.; Samavati, L.; Liu, Y. MAP kinase phosphatase-1, a gatekeeper of the acute innate immune response. Life Sci. 2020, 241, 117157. [Google Scholar] [CrossRef] [PubMed]
- Hoppstädter, J.; Ammit, A. Role of Dual-Specificity Phosphatase 1 in Glucocorticoid-Driven Anti-inflammatory Responses. Front. Immunol. 2019, 10, 1446. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.; Antoniades, W.; Okalova, J.; Roos, M.; Grimsey, N. Atypical p38 Signaling, Activation, and Implications for Disease. Int. J. Mol. Sci. 2021, 22, 4183. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.; Moilanen, E. Mitogen-Activated Protein Kinase Phosphatase 1 as an Inflammatory Factor and Drug Target. Basic Clin. Pharmacol. Toxicol. 2013, 114, 24–36. [Google Scholar] [CrossRef]
- Keränen, T.; Hömmö, T.; Hämäläinen, M.; Moilanen, E.; Korhonen, R. Anti-Inflammatory Effects of β2-Receptor Agonists Salbutamol and Terbutaline Are Mediated by MKP-1. PLoS ONE 2016, 11, e0148144. [Google Scholar] [CrossRef]
- Korhonen, R.; Hömmö, T.; Keränen, T.; Laavola, M.; Hämäläinen, M.; Vuolteenaho, K.; Lehtimäki, L.; Kankaanranta, H.; Moilanen, E. Attenuation of TNF production and experimentally induced inflammation by PDE4 inhibitor rolipram is mediated by MAPK phosphatase-1. Br. J. Pharmacol. 2013, 169, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, R.; Korhonen, R.; Moilanen, T.; Clark, A.; Moilanen, E. Aurothiomalate inhibits cyclooxygenase 2, matrix metalloproteinase 3, and interleukin-6 expression in chondrocytes by increasing MAPK phosphatase 1 expression and decreasing p38 phosphorylation: MAPK phosphatase 1 as a novel target for antirheumatic drugs. Arthritis Rheum. 2010, 62, 1650–1659. [Google Scholar] [CrossRef]
- Barnes, P.J. Glucocorticosteroids. Bact. Protein Toxins 2016, 237, 93–115. [Google Scholar]
- Hirsch, G.; Kitas, G.; Klocke, R. Intra-articular Corticosteroid Injection in Osteoarthritis of the Knee and Hip: Factors Predicting Pain Relief—A Systematic Review. Semin. Arthritis Rheum. 2013, 42, 451–473. [Google Scholar] [CrossRef] [PubMed]
- Pemmari, A.; Leppänen, T.; Hämäläinen, M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Widespread regulation of gene expression by glucocorticoids in chondrocytes from patients with osteoarthritis as determined by RNA-Seq. Arthritis Res. Ther. 2020, 22, 271. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.-M.; Lu, X.-H.; Lin, J.; Hu, J.; Rong, Z.-J.; Xu, W.-C.; Liu, Z.-W.; Zeng, W.-T. Knockdown of FOXM1 attenuates inflammatory response in human osteoarthritis chondrocytes. Int. Immunopharmacol. 2019, 68, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Capsoni, F.; Ongari, A.M.; Lonati, C.; Accetta, R.; Gatti, S.; Catania, A. α-Melanocyte-stimulating-hormone (α-MSH) modulates human chondrocyte activation induced by proinflammatory cytokines. BMC Musculoskelet. Disord. 2015, 16, 154. [Google Scholar] [CrossRef]
- Qu, R.; Chen, X.; Wang, W.; Qiu, C.; Ban, M.; Guo, L.; Vasilev, K.; Chen, J.; Li, W.; Zhao, Y. Ghrelin protects against osteoarthritis through interplay with Akt and NF-κB signaling pathways. FASEB J. 2018, 32, 1044–1058. [Google Scholar] [CrossRef]
- Little, C.B.; Barai, A.D.; Burkhardt, D.; Smith, S.M.; Fosang, A.J.; Werb, Z.; Shah, M.H.; Thompson, E.W. Matrix metalloproteinase 13-deficient mice are resistant to osteoarthritic cartilage erosion but not chondrocyte hypertrophy or osteophyte development. Arthritis Rheum. 2009, 60, 3723–3733. [Google Scholar] [CrossRef] [PubMed]
- Kamekura, S.; Hoshi, K.; Shimoaka, T.; Chung, U.; Chikuda, H.; Yamada, T.; Uchida, M.; Ogata, N.; Seichi, A.; Nakamura, K.; et al. Osteoarthritis development in novel experimental mouse models induced by knee joint instability. Osteoarthr. Cartil. 2005, 13, 632–641. [Google Scholar] [CrossRef]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of Osteoarthritis of the Hand, Hip, and Knee. Arthritis Care Res. 2020, 72, 149–162. [Google Scholar] [CrossRef]
- Zhao, Q.; Shepherd, E.G.; Manson, M.E.; Nelin, L.D.; Sorokin, A.; Liu, Y. The Role of Mitogen-activated Protein Kinase Phosphatase-1 in the Response of Alveolar Macrophages to Lipopolysaccharide. J. Biol. Chem. 2005, 280, 8101–8108. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, J.; Barnes, J.; Kokkonen, G.C.; Lee, J.C.; Liu, Y. Restraint of Proinflammatory Cytokine Biosynthesis by Mitogen-Activated Protein Kinase Phosphatase-1 in Lipopolysaccharide-Stimulated Macrophages. J. Immunol. 2002, 169, 6408–6416. [Google Scholar] [CrossRef] [PubMed]
- Issa, R.; Xie, S.; Khorasani, N.; Sukkar, M.; Adcock, I.; Lee, K.-Y.; Chung, K.F. Corticosteroid Inhibition of Growth-Related Oncogene Protein-α via Mitogen-Activated Kinase Phosphatase-1 in Airway Smooth Muscle Cells. J. Immunol. 2007, 178, 7366–7375. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Chivers, J.E.; Giembycz, M.A.; Newton, R. Long-Acting β2-Adrenoceptor Agonists Synergistically Enhance Glucocorticoid-Dependent Transcription in Human Airway Epithelial and Smooth Muscle Cells. Mol. Pharmacol. 2007, 73, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Tuure, L.; Hämäläinen, M.; Moilanen, E. PDE4 inhibitor rolipram inhibits the expression of microsomal prostaglandin E synthase-1 by a mechanism dependent on MAP kinase phosphatase-1. Pharmacol. Res. Perspect. 2017, 5, e00363. [Google Scholar] [CrossRef]
- Chi, H.; Barry, S.P.; Roth, R.J.; Wu, J.J.; Jones, E.A.; Bennett, A.M.; Flavell, R.A. Dynamic regulation of pro- and anti-inflammatory cytokines by MAPK phosphatase 1 (MKP-1) in innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Hammer, M.; Mages, J.; Dietrich, H.; Servatius, A.; Howells, N.; Cato, A.; Lang, R. Dual specificity phosphatase 1 (DUSP1) regulates a subset of LPS-induced genes and protects mice from lethal endotoxin shock. J. Exp. Med. 2006, 203, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, X.; Nelin, L.D.; Yao, Y.; Matta, R.; Manson, M.E.; Baliga, R.S.; Meng, X.; Smith, C.V.; Bauer, J.A.; et al. MAP kinase phosphatase 1 controls innate immune responses and suppresses endotoxic shock. J. Exp. Med. 2005, 203, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Salojin, K.V.; Owusu, I.B.; Millerchip, K.A.; Potter, M.; Platt, K.A.; Oravecz, T. Essential Role of MAPK Phosphatase-1 in the Negative Control of Innate Immune Responses. J. Immunol. 2006, 176, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.; Huotari, N.; Hömmö, T.; Leppänen, T.; Moilanen, E. The expression of interleukin-12 is increased by MAP kinase phosphatase-1 through a mechanism related to interferon regulatory factor 1. Mol. Immunol. 2012, 51, 219–226. [Google Scholar] [CrossRef]
- Turpeinen, T.; Nieminen, R.; Moilanen, E.; Korhonen, R. Mitogen-Activated Protein Kinase Phosphatase-1 Negatively Regulates the Expression of Interleukin-6, Interleukin-8, and Cyclooxygenase-2 in A549 Human Lung Epithelial Cells. J. Pharmacol. Exp. Ther. 2010, 333, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Huotari, N.; Hömmö, T.; Taimi, V.; Nieminen, R.; Moilanen, E.; Korhonen, R. Regulation of tristetraprolin expression by mitogen-activated protein kinase phosphatase-1. APMIS 2012, 120, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Tuure, L.; Hämäläinen, M.; Nummenmaa, E.; Moilanen, T.; Moilanen, E. Downregulation of microsomal prostaglandin E synthase-1 (mPGES-1) expression in chondrocytes is regulated by MAP kinase phosphatase-1 (MKP-1). Int. Immunopharmacol. 2019, 71, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Keränen, T.; Moilanen, E.; Korhonen, R. Suppression of cytokine production by glucocorticoids is mediated by MKP-1 in human lung epithelial cells. Inflamm. Res. 2017, 66, 441–449. [Google Scholar] [CrossRef]
- Zhong, H.-M.; Ding, Q.-H.; Chen, W.-P.; Luo, R.-B. Vorinostat, a HDAC inhibitor, showed anti-osteoarthritic activities through inhibition of iNOS and MMP expression, p38 and ERK phosphorylation and blocking NF-κB nuclear translocation. Int. Immunopharmacol. 2013, 17, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Kim, H.P. Matrix metalloproteinase-13 expression in IL-1β-treated chondrocytes by activation of the p38 MAPK/c-Fos/AP-1 and JAK/STAT pathways. Arch. Pharmacal Res. 2011, 34, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Yang, H.; Bau, B.; Söder, S.; Aigner, T. Role of mitogen-activated protein kinases and NFκB on IL-1β-induced effects on collagen type II, MMP-1 and 13 mRNA expression in normal articular human chondrocytes. Rheumatol. Int. 2006, 26, 900–903. [Google Scholar] [CrossRef]
- Dorfman, K.; Carrasco, D.; Gruda, M.; Ryan, C.; Lira, S.A.; Bravo, R. Disruption of the erp/mkp-1 gene does not affect mouse development: Normal MAP kinase activity in ERP/MKP-1-deficient fibroblasts. Oncogene 1996, 13, 925–931. [Google Scholar]
- Nummenmaa, E.; Hämäläinen, M.; Pemmari, A.; Moilanen, L.J.; Tuure, L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models. Int. J. Mol. Sci. 2020, 22, 87. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, A.; Juslin, S.; Nieminen, R.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Adiponectin associates with markers of cartilage degradation in osteoarthritis and induces production of proinflammatory and catabolic factors through mitogen-activated protein kinase pathways. Arthritis Res. Ther. 2011, 13, R184. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 19 April 2021).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Tuure, L.; Hämäläinen, M.; Whittle, B.J.; Moilanen, E. Microsomal Prostaglandin E Synthase-1 Expression in Inflammatory Conditions Is Downregulated by Dexamethasone: Seminal Role of the Regulatory Phosphatase MKP-1. Front. Pharmacol. 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Mean WT (IL-1β) | Mean WT (IL-1β + dexa) | WT FC | WT FDR p | Mean KO (IL-1β) | Mean KO (IL-1β + dexa) | KO FC | KO FDR p | p between the Effect of Dexamethasone in WT vs. KO Cells |
|---|---|---|---|---|---|---|---|---|---|
| MMP-3 | 173,754.8 | 35,166.6 | –25.8 | <0.0001 | 451,987.4 | 81,385.2 | –5.5 | <0.0001 | <0.0001 |
| MMP-9 | 126.2 | 28.1 | –6.6 | <0.0001 | 164.8 | 22.0 | –6.1 | <0.0001 | 0.72 |
| MMP-10 | 84.7 | 19.1 | –11.5 | <0.0001 | 224.3 | 45.8 | –4.7 | <0.0001 | 0.0069 |
| MMP-12 | 1242.1 | 446.7 | –14.6 | <0.0001 | 5572.8 | 516.9 | –11.0 | <0.0001 | 0.033 |
| MMP-13 | 67,643.1 | 38,510.2 | –2.3 | <0.0001 | 62,517.4 | 59,714.4 | –1.0 | 0.51 | <0.0001 |
| ADAMTS5 | 4783.2 | 2340.8 | –3.1 | <0.0001 | 8471.5 | 7194.1 | –1.2 | <0.0001 | <0.0001 |
| ADAM22 | 93.7 | 55.0 | –2.0 | <0.0001 | 56.5 | 26.8 | –2.0 | <0.0001 | 0.94 |
| Primer/Probe | Sequence | |
|---|---|---|
| mGAPDH | forward | 5′-GCATGGCCTTCCGTGTTC-3′ |
| reverse | 5′-GATGTCATCATACTTGGCAGGTTT-3′ | |
| probe | 5′-TCGTGGATCTGACGTGCCGCC-3′ | |
| mMMP-13 | forward | 5′-TTGTGTTTGCAGAGCACTACTTGA-3′ |
| reverse | 5′-AACTGTGGAGGTCACTGTAGACTTCTT-3′ | |
| probe | 5′-CATCCTGCGACTCTTGCGGGAATC-3′ | |
| mMKP-1 | forward | 5′-CTCCTGGTTCAACGAGGCTATT-3′ |
| reverse | 5′-TGCCGGCCTGGCAAT-3′ | |
| probe | 5′-CCATCAAGGATGCTGGAGGGAGAGTGTT-3′ | |
| hGAPDH | forward | 5′-AAGGTCGGAGTCAACGGATTT-3′ |
| reverse | 5′-GCAACAATATCCACTTTACCAGAGTTAA-3′ | |
| probe | 5′-CGCCTGGTCACCAGGGCTGC-3′ | |
| hMMP-13 | forward | 5′-TGATCTCTTTTGGAATTAAGGAGCAT-3′ |
| reverse | 5′-GGAACTACTTGTCCAGGTTTCATCAT-3′ | |
| probe | 5′-CCCTCTGGCCTGCTGGCTCATG-3′ | |
| hMKP-1 | forward | 5′-ACGAGGCCATTGACTTCATAGAC-3′ |
| reverse | 5′-TCGATTAGTCCTCATAAGGTAAGCAA-3′ | |
| probe | 5′-CCACTGCCAGGCAGGCATTTCC-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehtola, T.; Nummenmaa, E.; Tuure, L.; Hämäläinen, M.; Nieminen, R.M.; Moilanen, T.; Pemmari, A.; Moilanen, E. Dexamethasone Attenuates the Expression of MMP-13 in Chondrocytes through MKP-1. Int. J. Mol. Sci. 2022, 23, 3880. https://doi.org/10.3390/ijms23073880
Lehtola T, Nummenmaa E, Tuure L, Hämäläinen M, Nieminen RM, Moilanen T, Pemmari A, Moilanen E. Dexamethasone Attenuates the Expression of MMP-13 in Chondrocytes through MKP-1. International Journal of Molecular Sciences. 2022; 23(7):3880. https://doi.org/10.3390/ijms23073880
Chicago/Turabian StyleLehtola, Tiina, Elina Nummenmaa, Lauri Tuure, Mari Hämäläinen, Riina M. Nieminen, Teemu Moilanen, Antti Pemmari, and Eeva Moilanen. 2022. "Dexamethasone Attenuates the Expression of MMP-13 in Chondrocytes through MKP-1" International Journal of Molecular Sciences 23, no. 7: 3880. https://doi.org/10.3390/ijms23073880
APA StyleLehtola, T., Nummenmaa, E., Tuure, L., Hämäläinen, M., Nieminen, R. M., Moilanen, T., Pemmari, A., & Moilanen, E. (2022). Dexamethasone Attenuates the Expression of MMP-13 in Chondrocytes through MKP-1. International Journal of Molecular Sciences, 23(7), 3880. https://doi.org/10.3390/ijms23073880