Peptides Regulating Proliferative Activity and Inflammatory Pathways in the Monocyte/Macrophage THP-1 Cell Line

 ,
,  , , , ,
, , , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
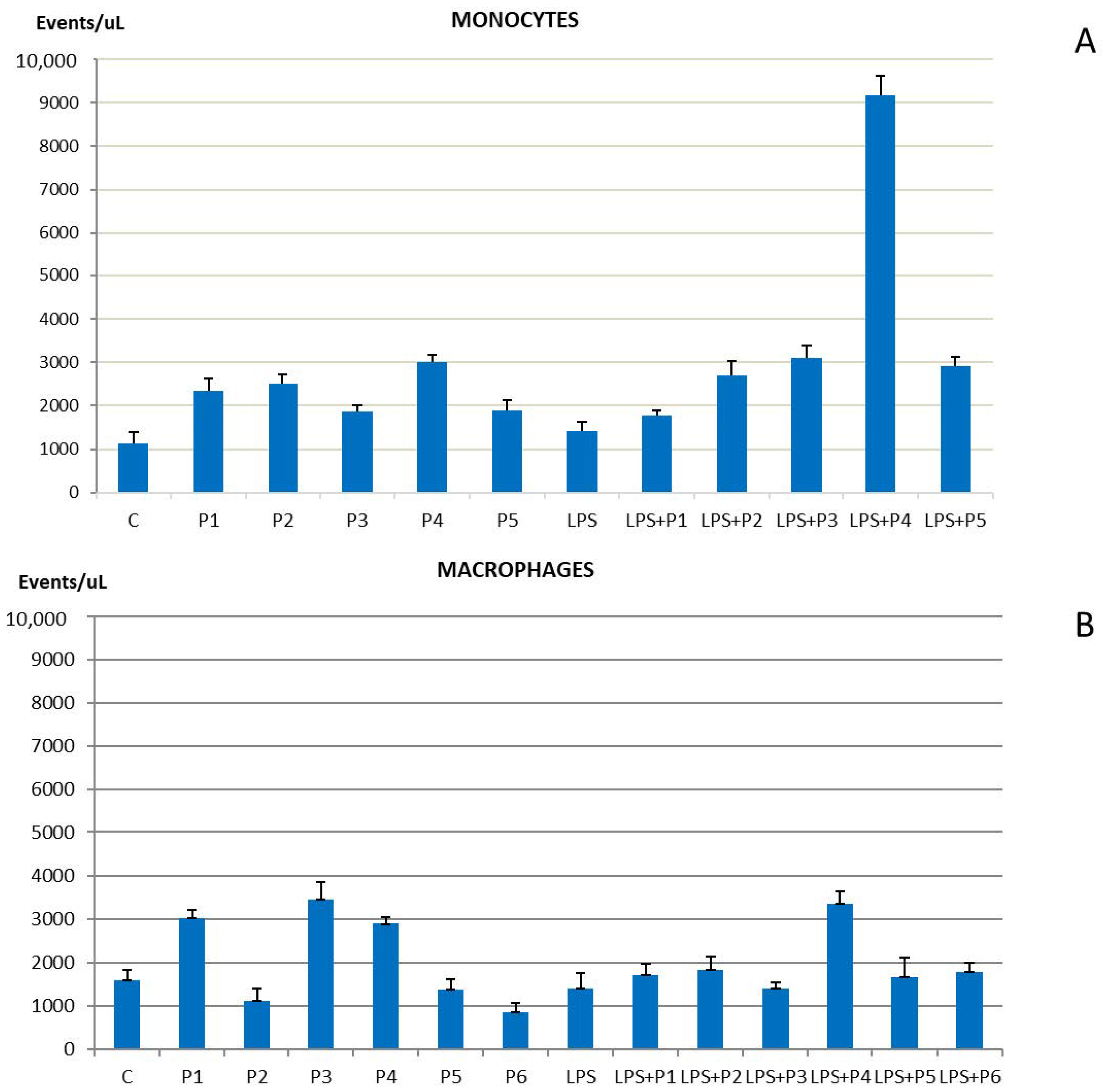
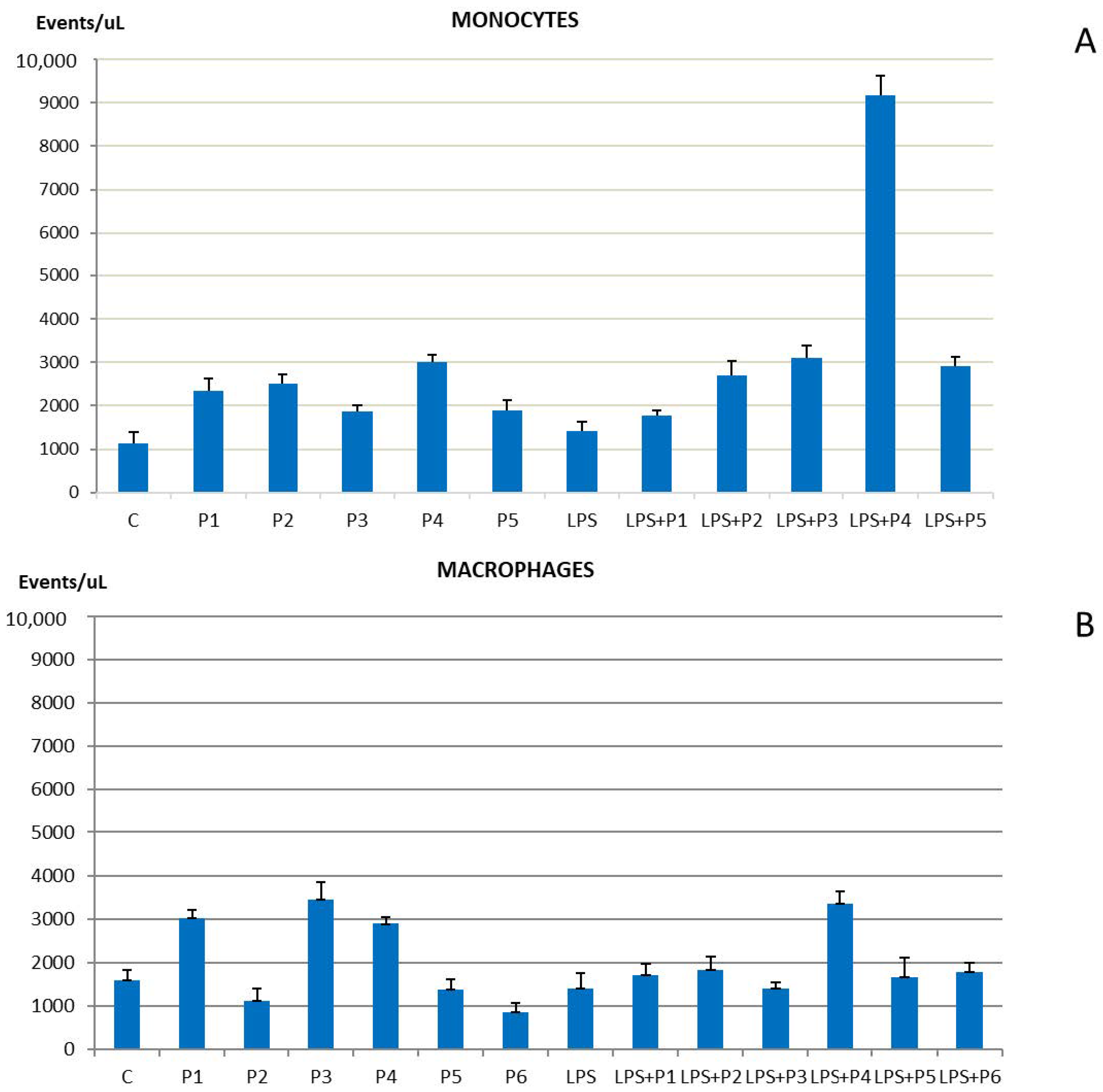
2.1. Phenotypic Markers of Cell Proliferation
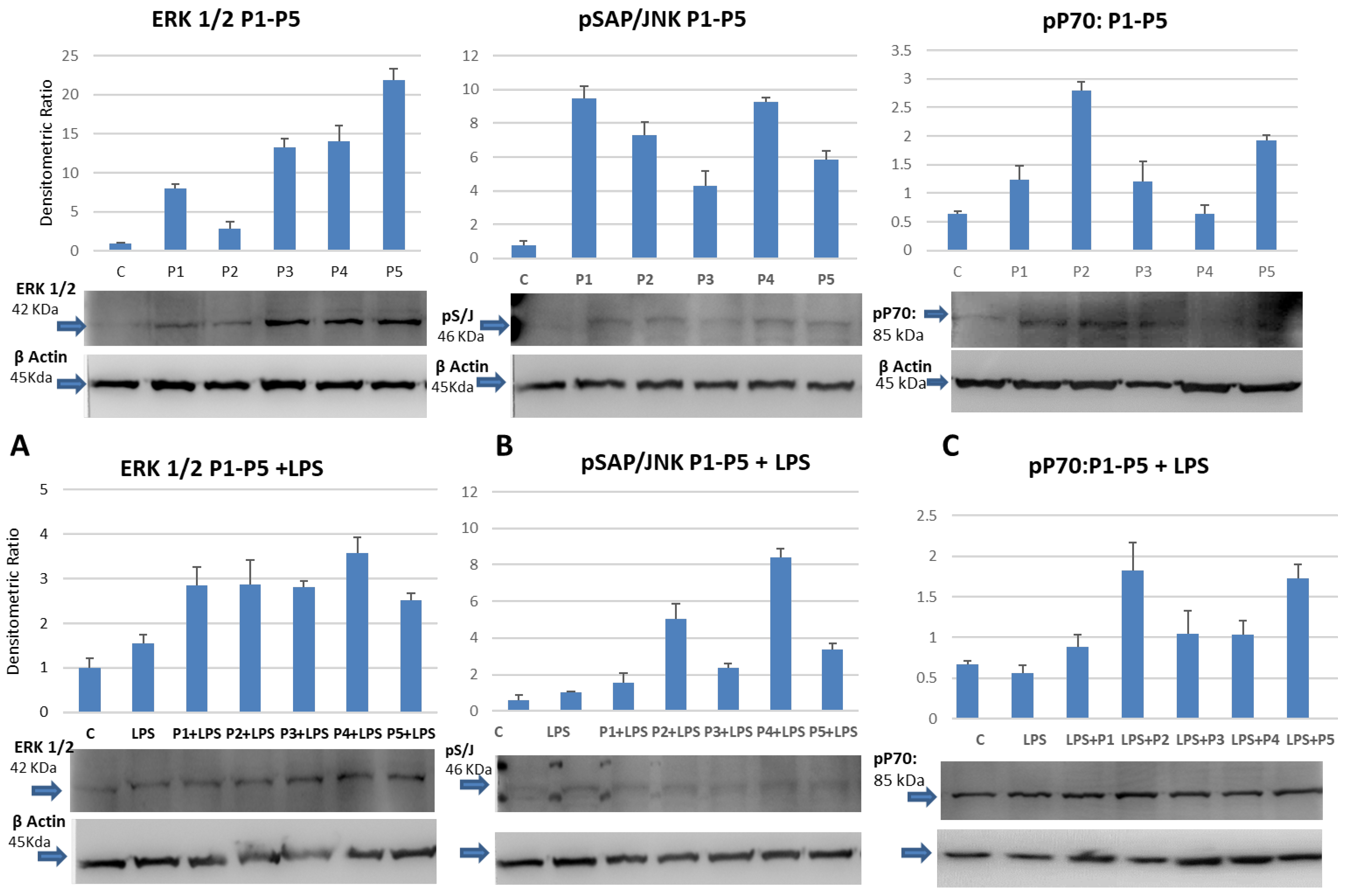
2.2. Induction of the Mitogen-Activated Phospho-ERK1/2 and Phospho-JNK in Cultured Monocytes
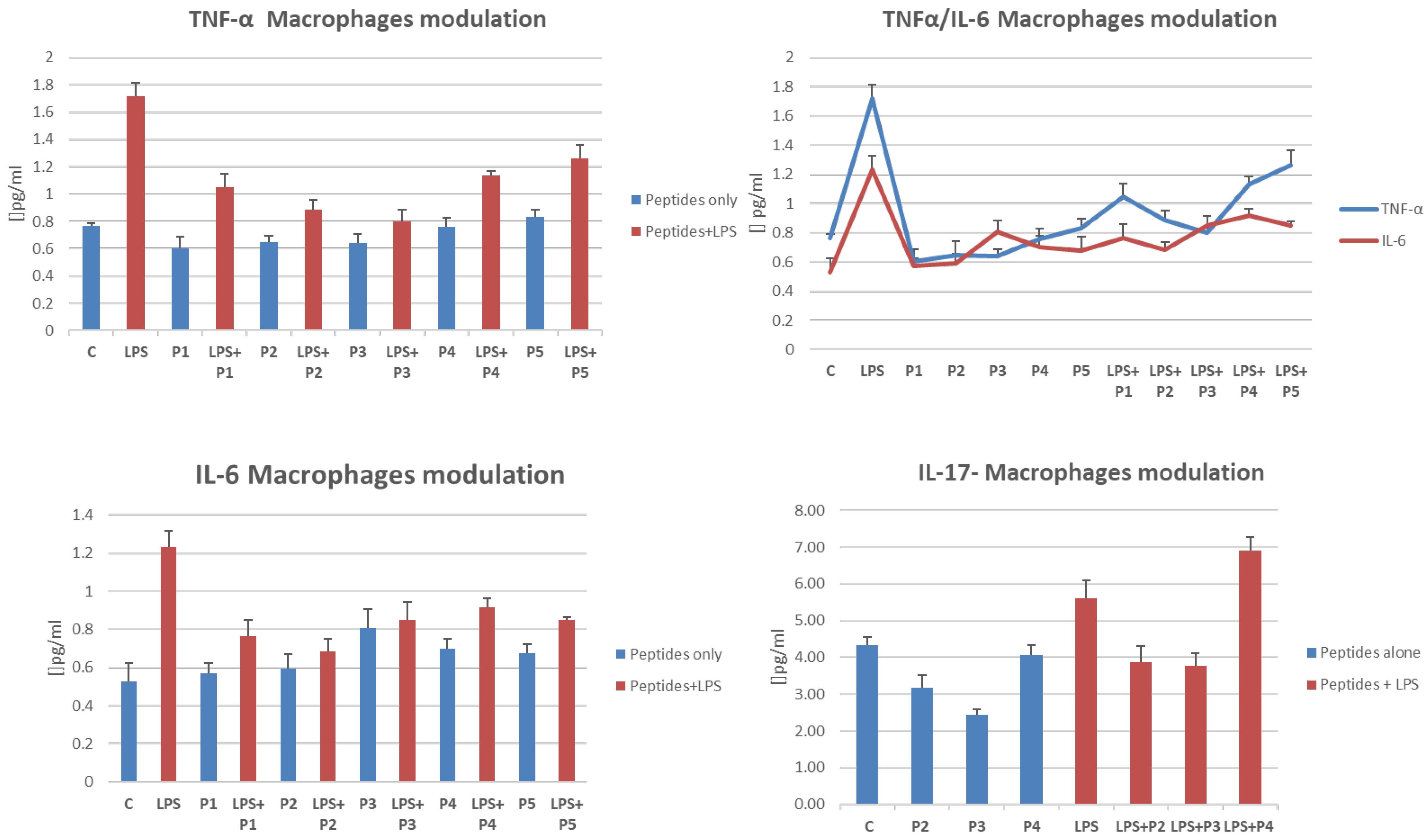
2.3. Chonluten (P5) Promotes Mild Level of TNF Secretion in Monocytes
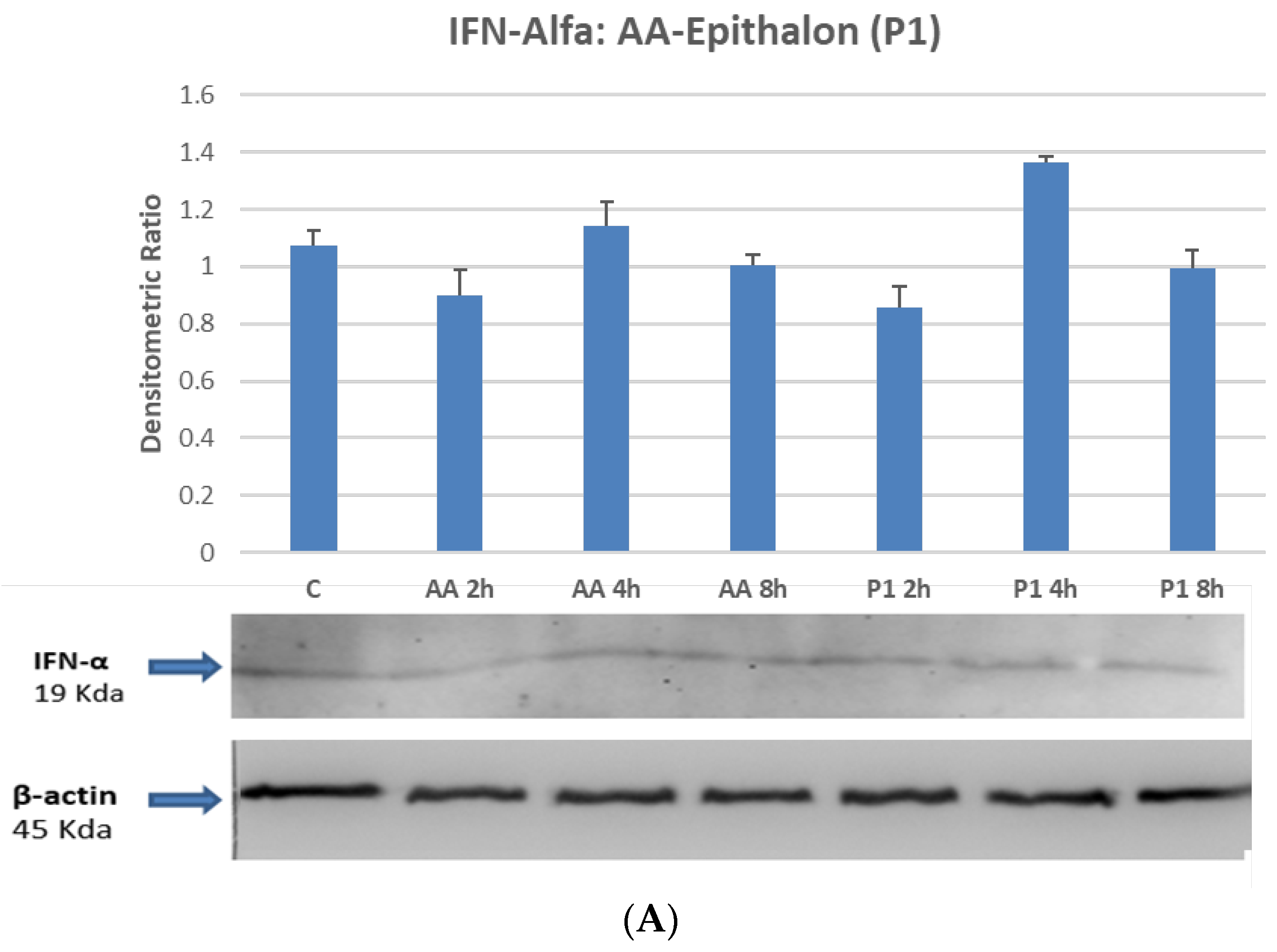
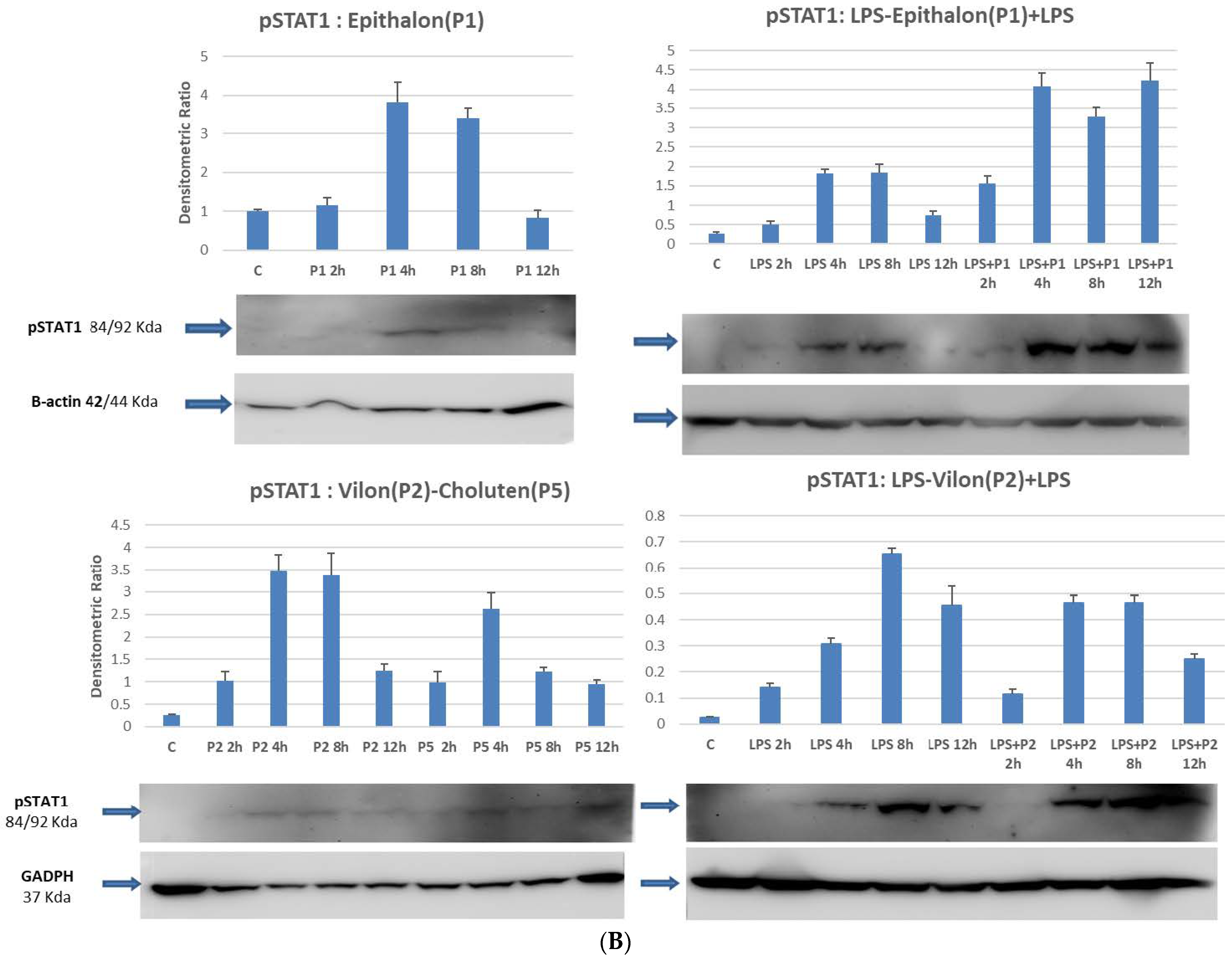
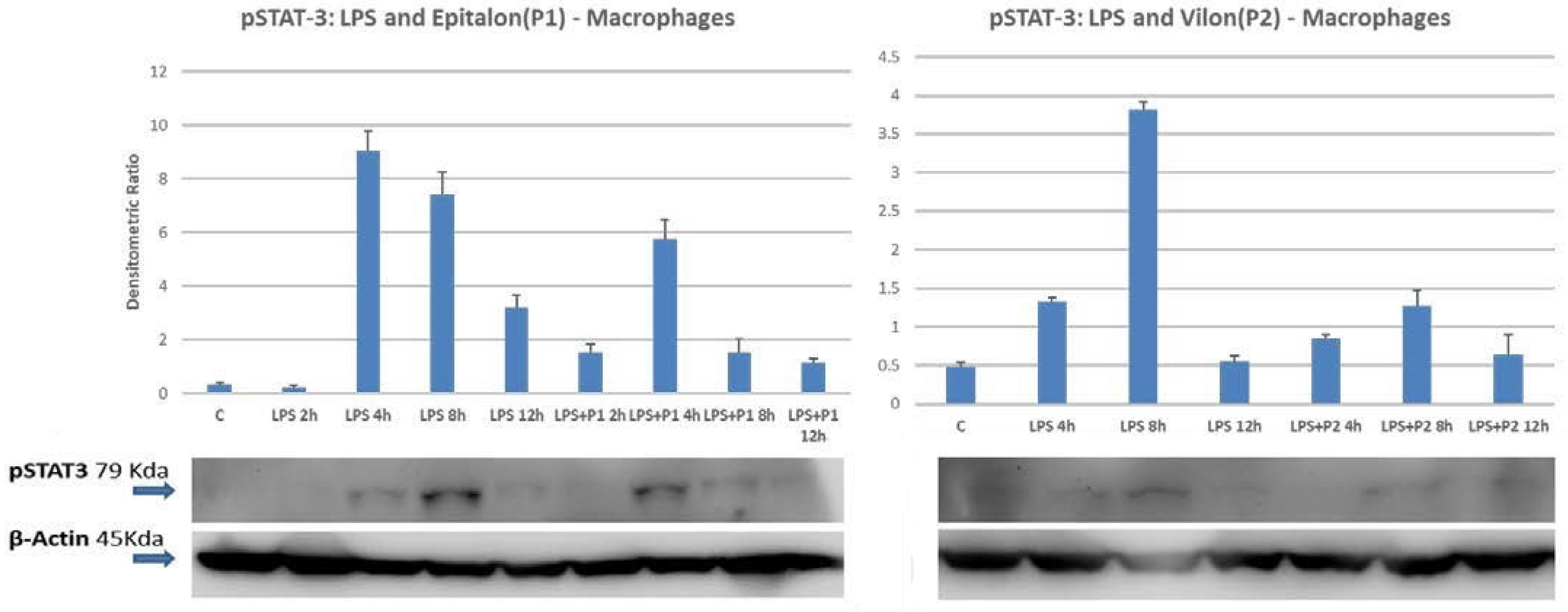
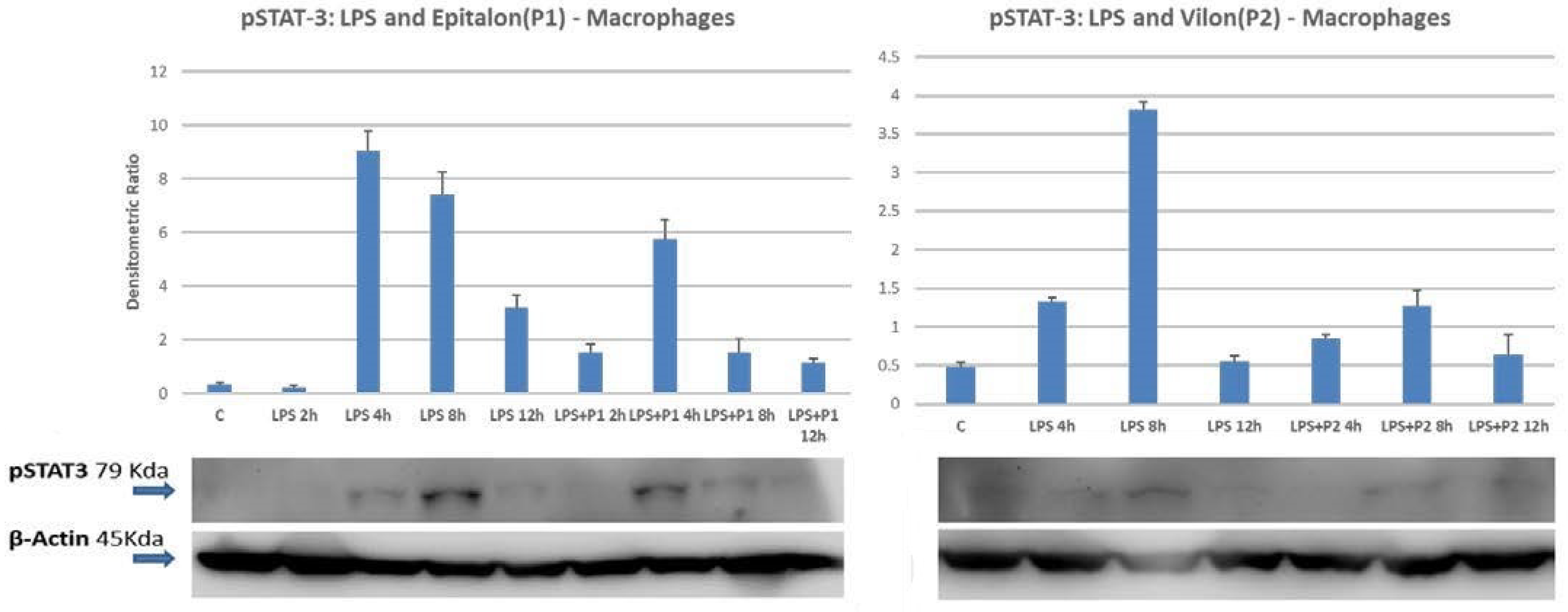
2.4. STAT1 and STAT3 Are Directly Phosphorylated by Peptides on Differentiated Macrophages
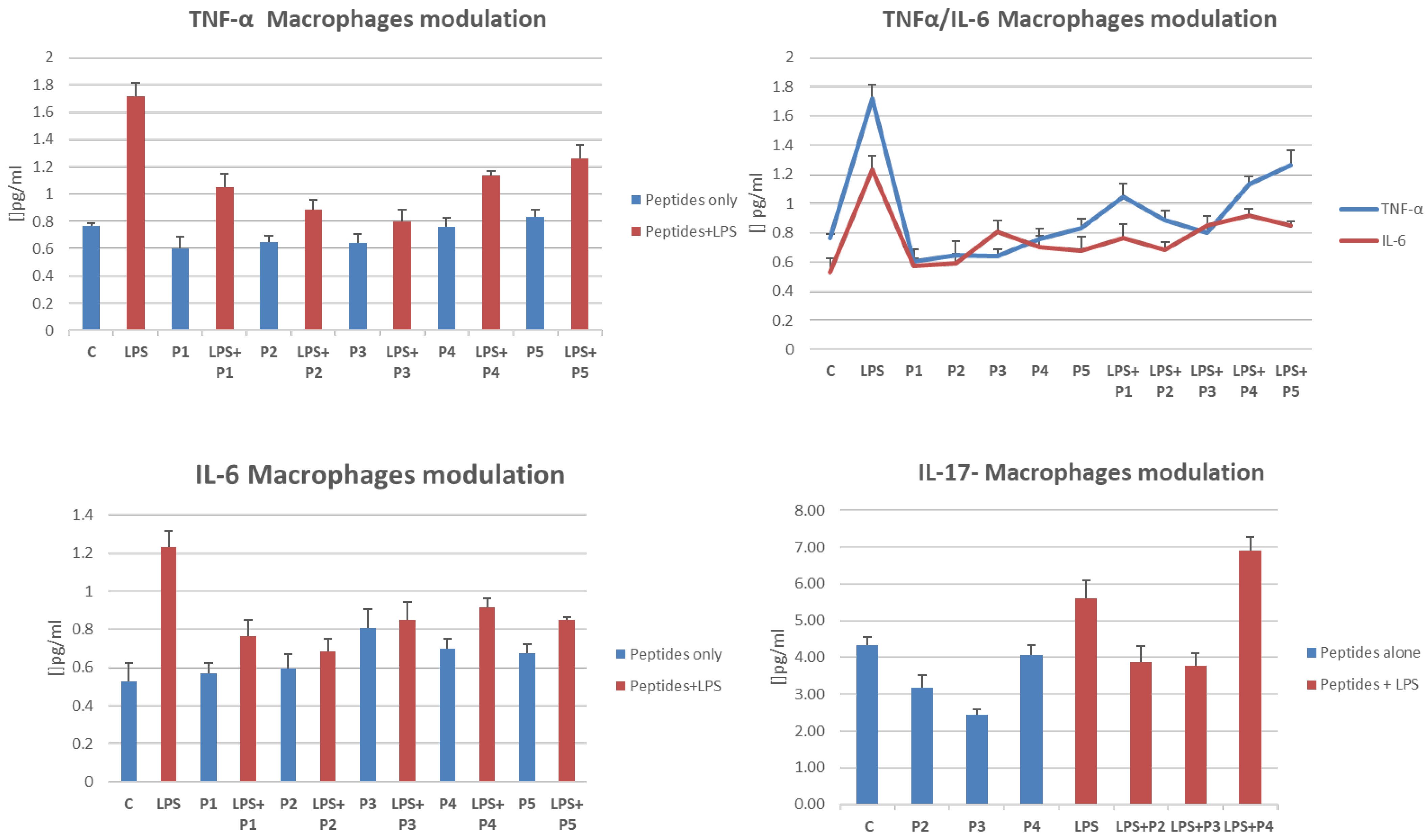
2.5. Cytokine Release Is Influenced by the Presence of Peptides in LPS-Treated THP-1 Cells
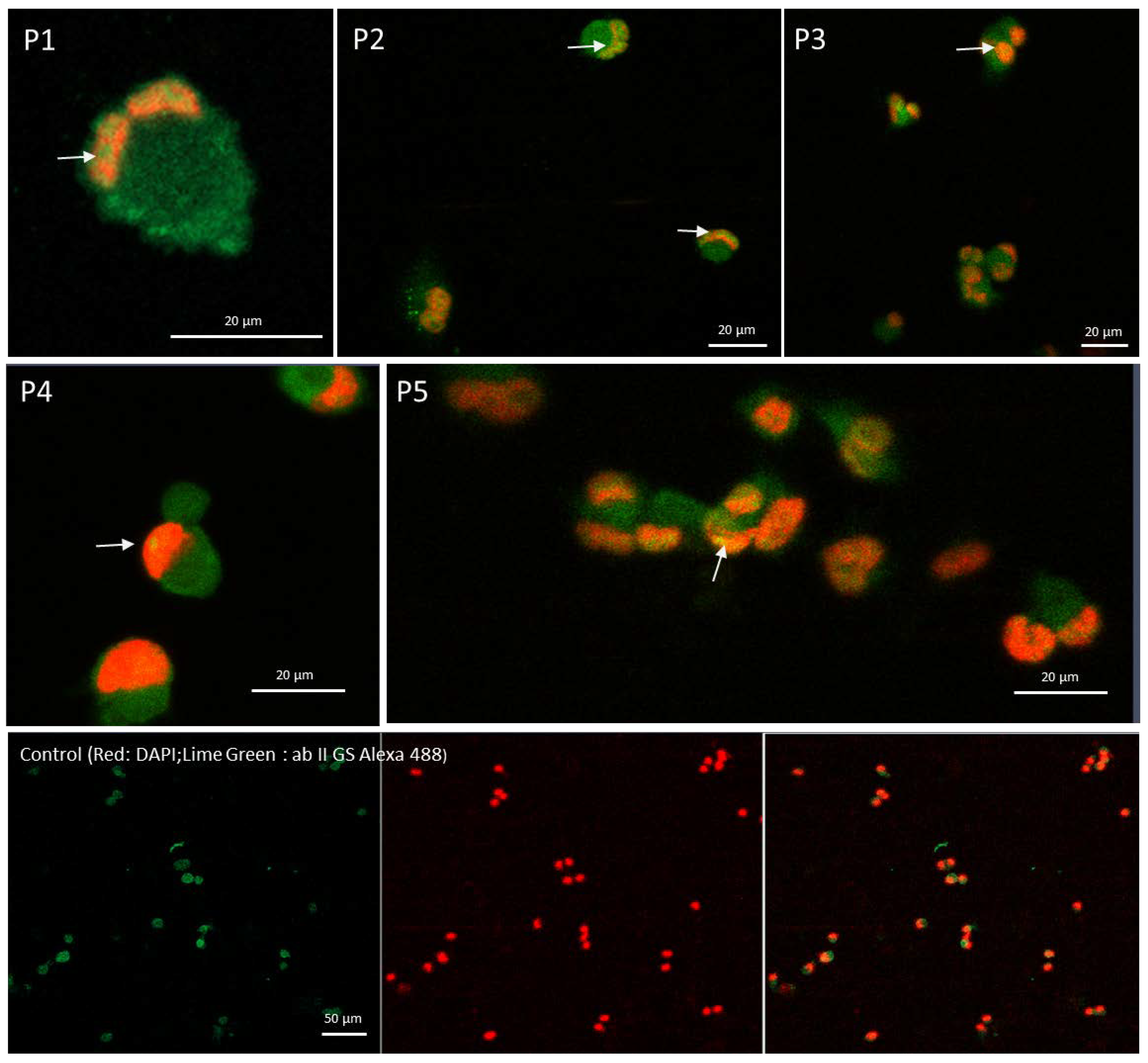
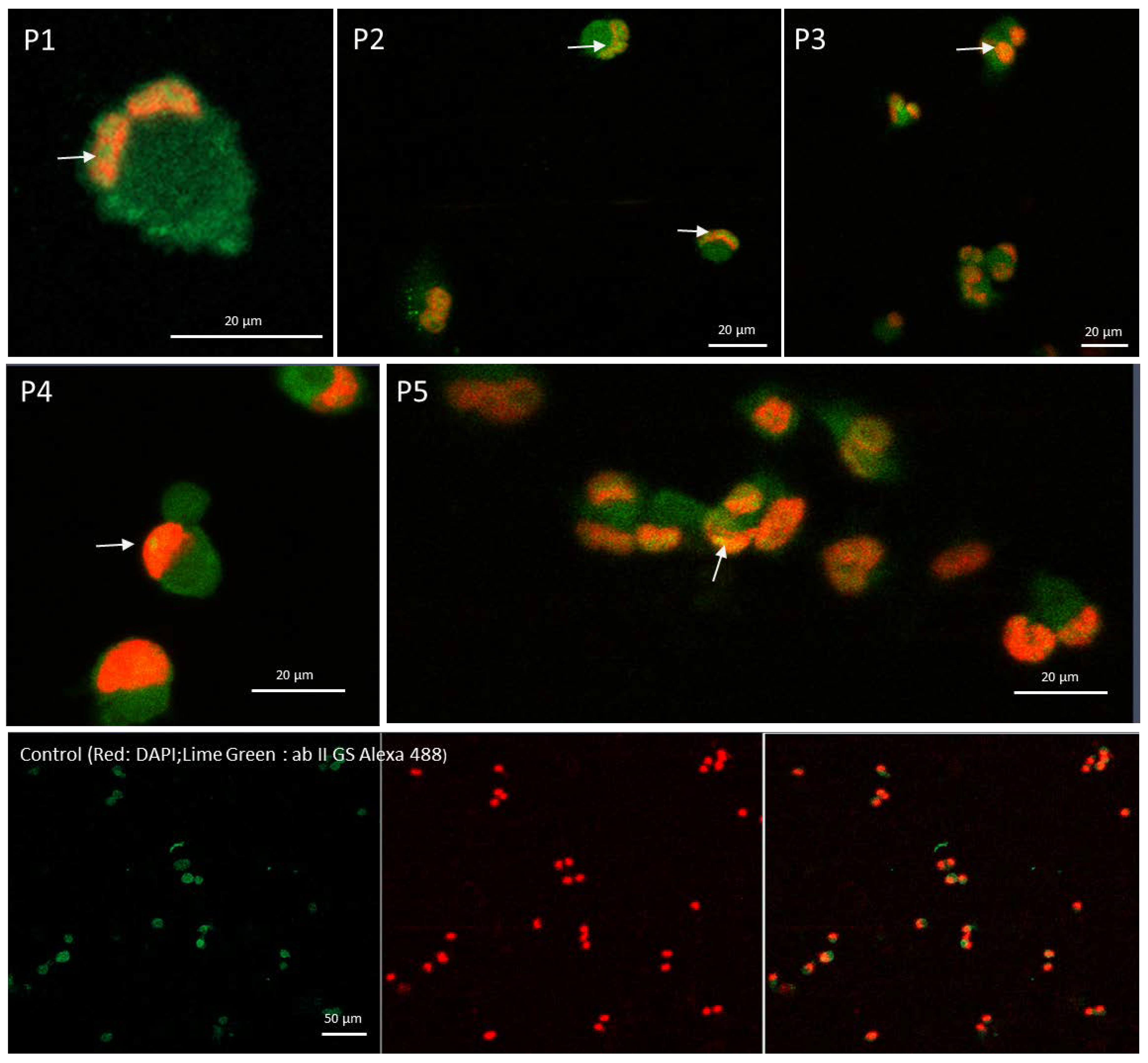
2.6. Confocal Microscopy Confirms That Peptide Mediates the Cytoplasmic STAT1 Transfer into Cells Nuclei
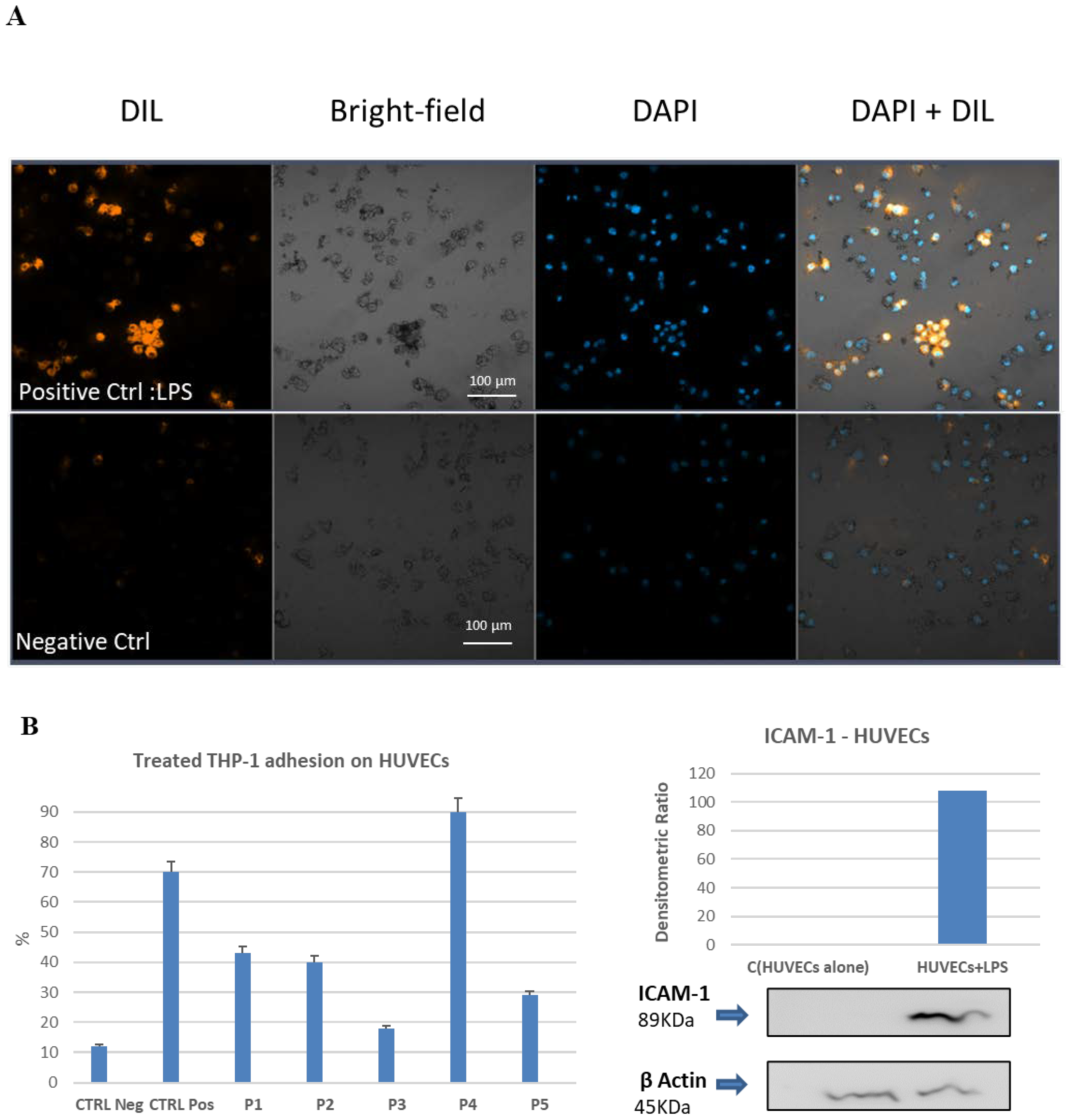
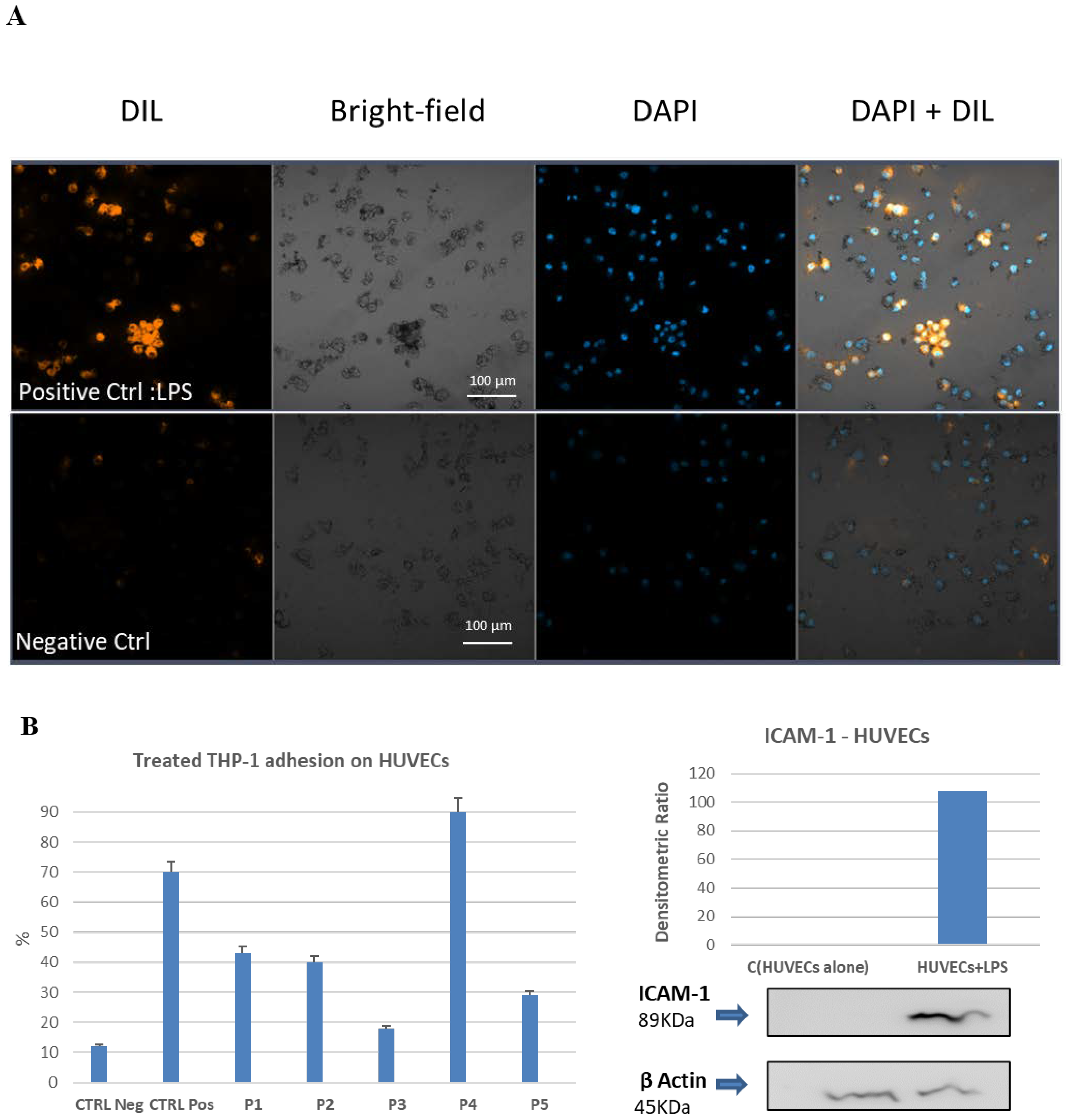
2.7. Cell–Cell Adhesion Assay between LPS-Activated Human Endothelial Cells and the THP-1 Monocytic Cells. Effect of Pretreatment with Bio-Peptides and Downregulation of Adhesion Activity
3. Discussion
4. Materials and Methods
4.1. Peptides
4.2. Cell Culture
4.3. Cell Viability
4.4. Flow Cytometry and Cell Cycle Analysis
4.5. Flow Cytometry Extracellular Vesicle Staining, Acquisition and Analysis
4.6. Protein Extraction and Western Blot
4.7. Confocal Microscopy
4.8. Cytokine Analysis
4.9. Monocytes Adhesion Assay
4.10. Validation Using Confocal Microscopy
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kantari, C.; Pederzoli-Ribeil, M.; Witko-Sarsat, V. The role of neutrophils and monocytes in innate immunity. Contrib. Microbiol. 2008, 15, 118–146. [Google Scholar] [PubMed]
- Kobayashi, S.D.; Voyich, J.M.; Burlak, C.; DeLeo, F.R. Neutrophils in the innate immune response. Arch. Immunol. Ther. Exp. 2005, 53, 505–517. [Google Scholar]
- Han, X.; Krempski, J.W.; Nadeau, K. Advances and novel developments in mechanisms of allergic inflammation. Allergy 2020, 75, 3100–3111. [Google Scholar] [CrossRef] [PubMed]
- Skuratovskaia, D.; Vulf, M.; Khaziakhmatova, O.; Malashchenko, V.; Komar, A.; Shunkin, E.; Shupletsova, V.; Goncharov, A.; Urazova, O.; Litvinova, L. Tissue-specific role of macrophages in noninfectious inflammatory disorders. Biomedicines 2020, 8, 400. [Google Scholar] [CrossRef]
- Ruder, B.; Becker, C. At the Forefront of the Mucosal Barrier: The Role of Macrophages in the Intestine. Cells 2020, 9, 2162. [Google Scholar] [CrossRef]
- Cortese, N.; Carriero, R.; Laghi, L.; Mantovani, A.; Marchesi, F. Prognostic significance of tumor-associated macrophages: Past, present and future. Semin. Immunol. 2020, 48, 101408. [Google Scholar] [CrossRef]
- Zhukova, O.V.; Kovaleva, T.F.; Arkhipova, E.V.; Ryabov, S.A.; Mukhina, I.V. Tumor-associated macrophages: Role in the pathological process of tumorigenesis and prospective therapeutic use. Biomed. Rep. 2020, 13, 47. [Google Scholar] [CrossRef]
- Larenas-Linnemann, D.; Rodríguez-Pérez, N.; Arias-Cruz, A.; Blandón-Vijil, M.V.; Del-Río-Navarro, B.E.; Estrada-Cardona, A.; Gereda, J.E.; Luna-Pech, J.A.; Navarrete-Rodríguez, E.M.; Onuma-Takane, E.; et al. Enhancing innate immunity against virus in times of COVID-19: Trying to untangle facts from fictions. World Allergy Organ. 2020, 13, 100476. [Google Scholar] [CrossRef]
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell. Mol. Immunol. 2020, 18, 45–56. [Google Scholar] [CrossRef]
- Li, M.; Chen, L.; Gao, Y.; Li, M.; Wang, X.; Qiang, L.; Wang, X. Recent Advances Targeting C-C chemokine receptor type 2 for Liver Diseases in Monocyte/Macrophage. Liver Int. 2020, 40, 2928–2936. [Google Scholar] [CrossRef]
- Mhaidly, R.; Mechta-Grigoriou, F. Fibroblast heterogeneity in tumor micro-environment: Role in immunosuppression and new therapies. Semin. Immunol. 2020, 48, 101417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kim, T.H.; Thauland, T.J.; Li, H.; Majedi, F.S.; Ly, C.; Gu, Z.; Butte, M.J.; Rowat, A.C.; Li, S. Unraveling the mechanobiology of immune cells. Curr. Opin. Biotechnol. 2020, 66, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.W. Studies on patients establish Crohn’s disease as a manifestation of impaired innate immunity. J. Intern. Med. 2019, 286, 373–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballerini, P.; Diomede, F.; Petragnani, N.; Cicchitti, S.; Merciaro, I.; Cavalcanti, M.F.; Trubiani, O. Conditioned medium from relapsing-remitting multiple sclerosis patients re- duces the expression and release of inflammatory cytokines induced by LPS-gingivalis in THP-1 and MO3.13 cell lines. Cytokine 2017, 96, 261–272. [Google Scholar] [CrossRef]
- Muñoz, J.; Akhavan, N.S.; Mullins, A.P.; Arjmandi, B.H. Macrophage Polarization and Osteoporosis: A Review. Nutrients 2020, 12, 2999. [Google Scholar] [CrossRef]
- Wang, J.; Xia, J.; Huang, R.; Hu, Y.; Fan, J.; Shu, Q.; Xu, J. Mesenchymal stem cell-derived extracellular vesicles alter disease outcomes via endorsement of macrophage polarization. Stem Cell Res. Ther. 2020, 11, 424. [Google Scholar] [CrossRef]
- Viegas, C.; Caetano, L.A.; Cox, J.; Korkalainen, M.; Haines, S.R.; Dannemiller, K.C.; Viegas, S.; Reponen, T. The effects of waste sorting in environmental microbiome, THP-1 cell viability and inflammatory responses. Environ. Res. 2020, 185, 109450. [Google Scholar] [CrossRef]
- Morozov, V.G.; Khavinson, V.K. Natural and synthetic thymic peptides as therapeutics for immune dysfunction. Int. J. Immunopharmacol. 1997, 19, 501–505. [Google Scholar] [CrossRef]
- Khavinson, V.K. Peptides and Ageing. Neuroendocrinol. Lett. 2002, 23 (Suppl. 3), 144. [Google Scholar]
- Khavinson, V.K.; Kuznik, B.I.; Ryzhak, G.A. Peptide bioregulators: The new class of geroprotectors. Communication 1. Results of experimental studies. Adv. Gerontol. 2012, 25, 696–708. [Google Scholar]
- Kuznik, B.I.; Pateyuk, A.V.; Rusaeva, N.S. Effect of tetrapeptides Lys-Glu-Asp-Gly and Ala-Glu-Asp-Gly on the structure and function of the thyroid gland in neonatally hypophysectomized chickens. Bull. Exp. Biol. Med. 2008, 145, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Lin’kova, N.S.; Tarnovskaya, S.I. Short Peptides Regulate Gene Expression. Bull. Exp. Biol. Med. 2016, 162, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Ashapkin, V.; Khavinson, V.; Shilovsky, G.; Linkova, N.; Vanuyshin, B. Gene expression in human mesenchymal stem cell aging cultures: Modulation by short peptides. Mol. Biol. Rep. 2020, 47, 4323–4329. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.; Diomede, F.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S.; Sinjari, B. AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism. Molecules 2020, 25, 609. [Google Scholar] [CrossRef] [Green Version]
- Ashapkin, V.V.; Linkova, N.S.; Khavinson, V.K.; Vanyushin, B.F. Epigenetic mechanisms of peptidergic regulation of gene expression during aging of human cells. Biochemistry 2015, 80, 310–322. [Google Scholar] [CrossRef]
- Wu, T.; Plett, P.A.; Chua, H.L.; Jacobsen, M.; Sandusky, G.E.; MacVittie, T.J.; Orschell, C.M. Immune Reconstitution and Thymic Involution in the Acute and Delayed Hematopoietic Radiation Syndromes. Health Phys. 2020, 119, 647–658. [Google Scholar] [CrossRef]
- Kinsella, S.; Dudakov, J.A. When the Damage Is Done: Injury and Repair in Thymus Function. Front. Immunol. 2020, 11, 1745. [Google Scholar] [CrossRef]
- Chalisova, N.I.; Khavinson, V.; Nozdrachev, A.D. Modulating and protective effects of thymic peptides in lymphoid tissue culture. Dokl. Biol. Sci. 2001, 379, 316–318. [Google Scholar] [CrossRef]
- Anisimov, V.N.; Khavinson, V.K.; Morozov, V.G. Immunomodulatory synthetic dipeptide L-Glu-L-Trp slows down aging and inhibits spontaneous carcinogenesis in rats. Biogerontology 2000, 1, 55–59. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Kuznik, B.I.; Tarnovskaya, S.I.; Lin’kova, N.S. Short Peptides and Telomere Length Regulator Hormone Irisin. Bull. Exp. Biol. Med. 2016, 160, 347–349. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Kopylov, A.T.; Vaskovsky, B.V.; Ryzhak, G.A.; Lin’kova, N.S. Identification of Peptide AEDG in the Polypeptide Complex of the Pineal Gland. Bull. Exp. Biol. Med. 2017, 164, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Caputi, S.; Trubiani, O.; Sinjari, B.; Trofimova, S.; Diomede, F.; Linkova, N.; Diatlova, A.; Khavinson, V. Effect of short peptides on neuronal differentiation of stem cells. Int. J. Immunopathol. Pharmacol. 2019, 33, 2058738419828613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozov, A.V.; Khizhkin, E.A.; Svechkina, E.B.; Vinogradova, I.A.; Ilyukha, V.A.; Anisimov, V.N.; Khavinson, V.K. Effects of Geroprotectors on Age-Related Changes in Proteolytic Digestive Enzyme Activities at Different Lighting Conditions. Bull. Exp. Biol. Med. 2015, 159, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.; Linkova, N.; Dyatlova, A.; Kuznik, B.; Umnov, R. Peptides: Prospects for Use in the Treatment of COVID-19. Molecules 2020, 25, 4389. [Google Scholar] [CrossRef]
- Kraskovskaya, N.A.; Kukanova, E.O.; Lin’kova, N.S.; Popugaeva, E.A.; Khavinson, V.K. Tripeptides Restore the Number of Neuronal Spines under Conditions of In Vitro Modeled Alzheimer’s Disease. Bull. Exp. Biol. Med. 2017, 163, 550–553. [Google Scholar] [CrossRef]
- Sevostianova, N.N.; Linkova, N.S.; Polyakova, V.O.; Chervyakova, N.A.; Kostylev, A.V.; Durnova, A.O.; Kvetnoy, I.M.; Abdulragimov, R.I.; Khavinson, V.H. Immunomodulating effects of Vilon and its analogue in the culture of human and animal thymus cells. Bull. Exp. Biol. Med. 2013, 154, 562–565. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Linkova, N.S.; Kvetnoy, I.M.; Kvetnaia, T.V.; Polyakova, V.O.; Korf, H.W. Molecular cellular mechanisms of peptide regulation of melatonin synthesis in pinealocyte culture. Bull. Exp. Biol. Med. 2012, 153, 255–258. [Google Scholar] [CrossRef]
- Kolchina, N.; Khavinson, V.; Linkova, N.; Yakimov, A.; Baitin, D.; Afanasyeva, A.; Petukhov, M. Systematic search for structural motifs of peptide binding to double-stranded DNA. Nucleic Acids Res. 2019, 47, 10553–10563. [Google Scholar] [CrossRef]
- Chaudhry, M.S.; Velardi, E.; Dudakov, J.A.; van den Brink, M.R. Thymus: The next (re)generation. Immunol. Rev. 2016, 271, 56–71. [Google Scholar] [CrossRef] [Green Version]
- Geenen, V.; Trussart, C.; Michaux, H.; Halouani, A.; Jaïdane, H.; Collée, C.; Renard, C.; Daukandt, M.; Ledent, P.; Martens, H. The presentation of neuroendocrine self-peptides in the thymus: An essential event for individual life and vertebrate survival. Ann. N. Y. Acad. Sci. 2019, 1455, 113–125. [Google Scholar] [CrossRef]
- Pan, L.; De Smet, I. Expanding the Mitogen-Activated Protein Kinase (MAPK) Universe: An Update on MAP4Ks. Front. Plant Sci. 2020, 11, 1220. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, J.; Silke, J. An overview of mammalian p38 mitogen-activated protein kinases, central regulators of cell stress and receptor signaling. F1000Research 2020, 9, 653. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Toniato, E.; Colagrande, A.; Alesse, E.; Alleva, C.; Screpanti, I.; Morrone, S.; Scarpa, S.; Frati, L.; Hayday, A.C.; et al. Heavy-metal modulation of the human intercellular adhesion molecule (ICAM-1) gene expression. Biochim. Biophys. Acta 1995, 1261, 107–114. [Google Scholar] [CrossRef]
- Rayees, S.; Rochford, I.; Joshi, J.C.; Joshi, B.; Banerjee, S.; Mehta, D. Macrophage TLR4 and PAR2 Signaling: Role in Regulating Vascular Inflammatory Injury and Repair. Front. Immunol. 2020, 11, 2091. [Google Scholar] [CrossRef]
- Sinjari, B.; Diomede, F.; Khavinson, V.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S. Short Peptides Protect Oral Stem Cells from Ageing. Stem Cell Rev. Rep. 2020, 16, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Bikker, R.; Welz, B.; Christmann, M.; Brand, K. TNF Tolerance in Monocytes and Macrophages: Characteristics and Molecular Mechanisms. J. Immunol. Res. 2017, 2017, 9570129. [Google Scholar] [CrossRef] [Green Version]
- Bosshart, H.; Heinzelmann, H. THP-1 cells as a model for human monocytes. Ann. Transl. Med. 2016, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, S.; De Majo, F.; Kim, J.; Trenti, A.; Trevisi, L.; Fadini, G.P.; Bolego, C.; Zandstra, P.W.; Cignarella, A.; Vitiello, L. Convenience versus Biological Significance: Are PMA-Differentiated THP-1 Cells a Reliable Substitute for Blood-Derived Macrophages When Studying In vitro Polarization? Front. Pharmacol. 2018, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Swirski, F.K.; Hilgendorf, I.; Robbins, C. From proliferation to proliferation: Monocyte lineage comes full circle. Semin. Immunopathol. 2014, 36, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.M.; Hadadi, E.; Dang, T.M.; Yeap, W.H.; Tan, C.T.Y.; Ng, T.P.; Larbi, A.; Wong, S.C. The pro-inflammatory phenotype of the human non-classical monocyte subset is attributed to senescence. Cell Death Dis. 2018, 9, 266. [Google Scholar] [CrossRef]
- Lloberas, J.; Valverde-Estrella, L.; Tur, J.; Vico, T.; Celada, A. Mitogen-Activated Protein Kinases and Mitogen Kinase Phosphatase 1: A Critical Interplay in Macrophage Biology. Front. Mol. Biosci. 2016, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Mebratu, Y.; Tesfaigzi, Y. How ERK1/2 activation controls cell proliferation and cell death is subcellular localization the answer? Cell Cycle 2009, 8, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Muniyappa, H.; Das Kumuda, C. Activation of c-Jun N-terminal kinase (JNK) by widely used specific p38 MAPK inhibitors SB202190 and SB203580: A MLK-3-MKK7-dependent mechanism. Cell. Signal. 2008, 20, 675–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromberg, J.F. Activation of STAT proteins and growth control. BioEssays 2001, 23, 161–169. [Google Scholar] [CrossRef]
- Wanga, Y.B.; van Boxel-Dezaireb, A.H.H.; Cheonb, H.; Yanga, J.; Stark, G.R. STAT3 activation in response to IL-6 is prolonged by the binding of IL-6 receptor to EGF receptor. Proc. Natl. Acad. Sci. USA 2013, 110, 16975–16980. [Google Scholar] [CrossRef] [Green Version]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.; Dockrell, D.H. The identification of markers of macrophage differentiation in PMA-stimulated THP-1 cells and monocyte-derived macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef]
- Prontera, C.; Martelli, N.; Evangelista, V.; D’Urbano, E.; Manarini, S.; Recchiuti, A.; Dragani, A.; Passeri, C.; Davì, G.; Romano, M. Homocysteine Modulates the CD40/CD40L System. J. Am. Coll. Cardiol. 2007, 49, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Lanuti, P.; Bertagnolo, V.; Gaspari, A.R.; Ciccocioppo, F.; Pierdomenico, L.; Bascelli, A.; Sabatino, G.; Miscia, S.; Marchisio, M. Parallel regulation of PKC-alpha and PKC-delta characterizes the occurrence of erythroid differentiation from human primary hematopoietic progenitors. Exp. Hematol. 2006, 34, 1624–1634. [Google Scholar]
- Lanuti, P.; Marchisio, M.; Cantilena, S.; Paludi, M.; Bascelli, A.; Gaspari, A.R.; Grifone, G.; Centurione, M.A.; Papa, S.; Di Pietro, R.; et al. A flow cytometry procedure for simultaneous characterization of cell DNA content and expression of intracellular protein kinase C-zeta. J. Immunol. Methods 2006, 315, 37–48. [Google Scholar]
- Simeone, P.; Celia, C.; Bologna, G.; Ercolino, E.; Pierdomenico, L.; Cilurzo, F.; Grande, R.; Diomede, F.; Vespa, S.; Canonico, B.; et al. Diameters and Fluorescence Calibration for Extracellular Vesicle Analyses by Flow Cytometry. Int. J. Mol. Sci. 2020, 21, 7885. [Google Scholar] [CrossRef]
- Marchisio, M.; Simeone, P.; Bologna, G.; Ercolino, E.; Pierdomenico, L.; Pieragostino, D.; Ventrella, A.; Antonini, F.; Del Zotto, G.; Vergara, D.; et al. Flow Cytometry Analysis of Circulating Extracellular Vesicle Subtypes from Fresh Peripheral Blood Samples. Int. J. Mol. Sci. 2020, 22, 48. [Google Scholar]
- Lanuti, P.; Simeone, P.; Rotta, G.; Almici, C.; Avvisati, G.; Azzaro, R.; Bologna, G.; Budillon, A.; Di Cerbo, M.; Di Gennaro, E.; et al. A standardized flow cytometry network study for the assessment of circulating endothelial cell physiological ranges. Sci. Rep. 2018, 8, 5823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieragostino, D.; Lanuti, P.; Cicalini, I.; Cufaro, M.C.; Ciccocioppo, F.; Ronci, M.; Simeone, P.; Onofrj, M.; van der Pol, E.; Fontana, A.; et al. Proteomics characterization of extracellular vesicles sorted by flow cytometry reveals a disease-specific molecular cross-talk from cerebrospinal fluid and tears in multiple sclerosis. J. Proteom. 2019, 204, 103403. [Google Scholar] [CrossRef]
- Brocco, D.; Lanuti, P.; Simeone, P.; Bologna, G.; Pieragostino, D.; Cufaro, M.C.; Graziano, V.; Peri, M.; Di Marino, P.; De Tursi, M.; et al. Circulating Cancer Stem Cell-Derived Extracellular Vesicles as a Novel Biomarker for Clinical Outcome Evaluation. J. Oncol. 2019, 2019, 5879616. [Google Scholar] [CrossRef] [PubMed]
- De Iuliis, V.; Marino, A.; Caruso, M.; Capodifoglio, S.; Flati, V.; Marynuk, A.; Marricareda, V.; Ursi, S.; Lanuti, P.; Talora, C.; et al. Autophagy processes are dependent on EGF receptor signaling. Oncotarget 2018, 9, 30289–30303. [Google Scholar] [CrossRef]
- Holmes, K.L.; Otten, G.; Yokoyama, W.M. Flow cytometry analysis using the Becton Dickinson FACS Calibur. Curr. Protoc. Immunol. 2002, 49, 5.4.1–5.4.22. [Google Scholar] [CrossRef]
- Vincent, V.; Thakkar, H.; Verma, A.; Sen, A.; Chandran, N.; Singh, A. A novel flow cytometry-based quantitative monocyte adhesion assay to estimate endothelial cell activation in vitro. Biotechniques 2020, 68, 325–333. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avolio, F.; Martinotti, S.; Khavinson, V.K.; Esposito, J.E.; Giambuzzi, G.; Marino, A.; Mironova, E.; Pulcini, R.; Robuffo, I.; Bologna, G.; et al. Peptides Regulating Proliferative Activity and Inflammatory Pathways in the Monocyte/Macrophage THP-1 Cell Line. Int. J. Mol. Sci. 2022, 23, 3607. https://doi.org/10.3390/ijms23073607
Avolio F, Martinotti S, Khavinson VK, Esposito JE, Giambuzzi G, Marino A, Mironova E, Pulcini R, Robuffo I, Bologna G, et al. Peptides Regulating Proliferative Activity and Inflammatory Pathways in the Monocyte/Macrophage THP-1 Cell Line. International Journal of Molecular Sciences. 2022; 23(7):3607. https://doi.org/10.3390/ijms23073607
Chicago/Turabian StyleAvolio, Francesco, Stefano Martinotti, Vladimir Kh. Khavinson, Jessica Elisabetta Esposito, Giulia Giambuzzi, Antonio Marino, Ekaterina Mironova, Riccardo Pulcini, Iole Robuffo, Giuseppina Bologna, and et al. 2022. "Peptides Regulating Proliferative Activity and Inflammatory Pathways in the Monocyte/Macrophage THP-1 Cell Line" International Journal of Molecular Sciences 23, no. 7: 3607. https://doi.org/10.3390/ijms23073607
APA StyleAvolio, F., Martinotti, S., Khavinson, V. K., Esposito, J. E., Giambuzzi, G., Marino, A., Mironova, E., Pulcini, R., Robuffo, I., Bologna, G., Simeone, P., Lanuti, P., Guarnieri, S., Trofimova, S., Procopio, A. D., & Toniato, E. (2022). Peptides Regulating Proliferative Activity and Inflammatory Pathways in the Monocyte/Macrophage THP-1 Cell Line. International Journal of Molecular Sciences, 23(7), 3607. https://doi.org/10.3390/ijms23073607