
Anti-Inflammatory Characteristics of Local Anesthetics: Inhibition of TNF-α Secretion of Lipopolysaccharide-Stimulated Leucocytes in Human Blood Samples

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Individuals
2.2. Individual Patterns of TNF-α Inhibition
2.3. Influence of Biometric Factors on TNF-α Inhibition by LAs
3. Discussion
3.1. LAs Reduce TNF-α Stimulation in White Blood Cells
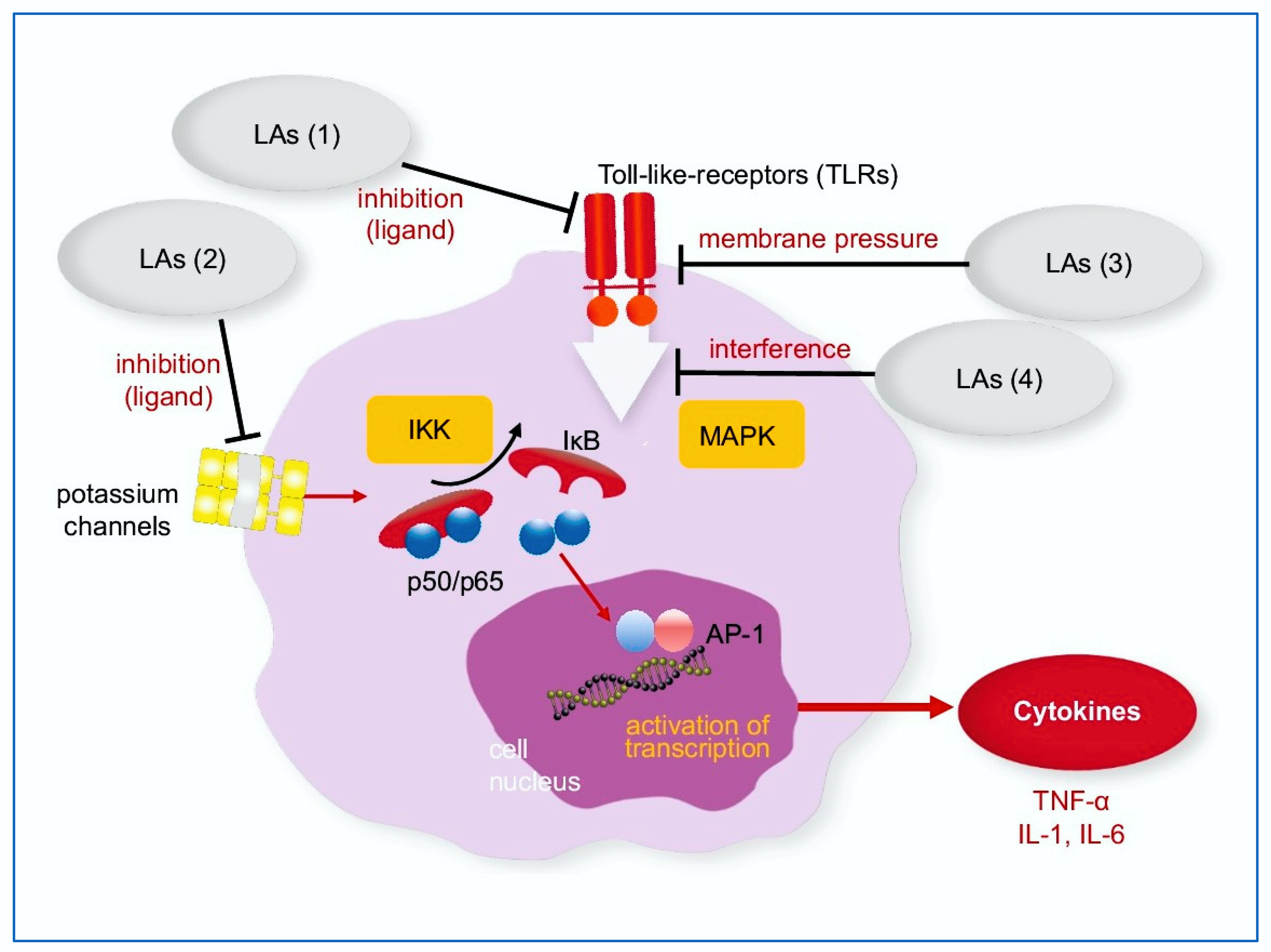
3.2. Molecular Pathway of the TNF-α Reduction by LA
3.3. Limitations and Strength of the Study
3.3.1. Applicability of the Test System
3.3.2. Why Such Marked Differences between Individuals?
3.3.3. Concentration of LAs in the Test System and In Vivo
3.3.4. Small Observational Study
3.4. Clinical Implications and Future Investigations
3.5. TNF-α Testing—A Prognosis of the Therapeutic Potential of LA?
4. Material and Methods
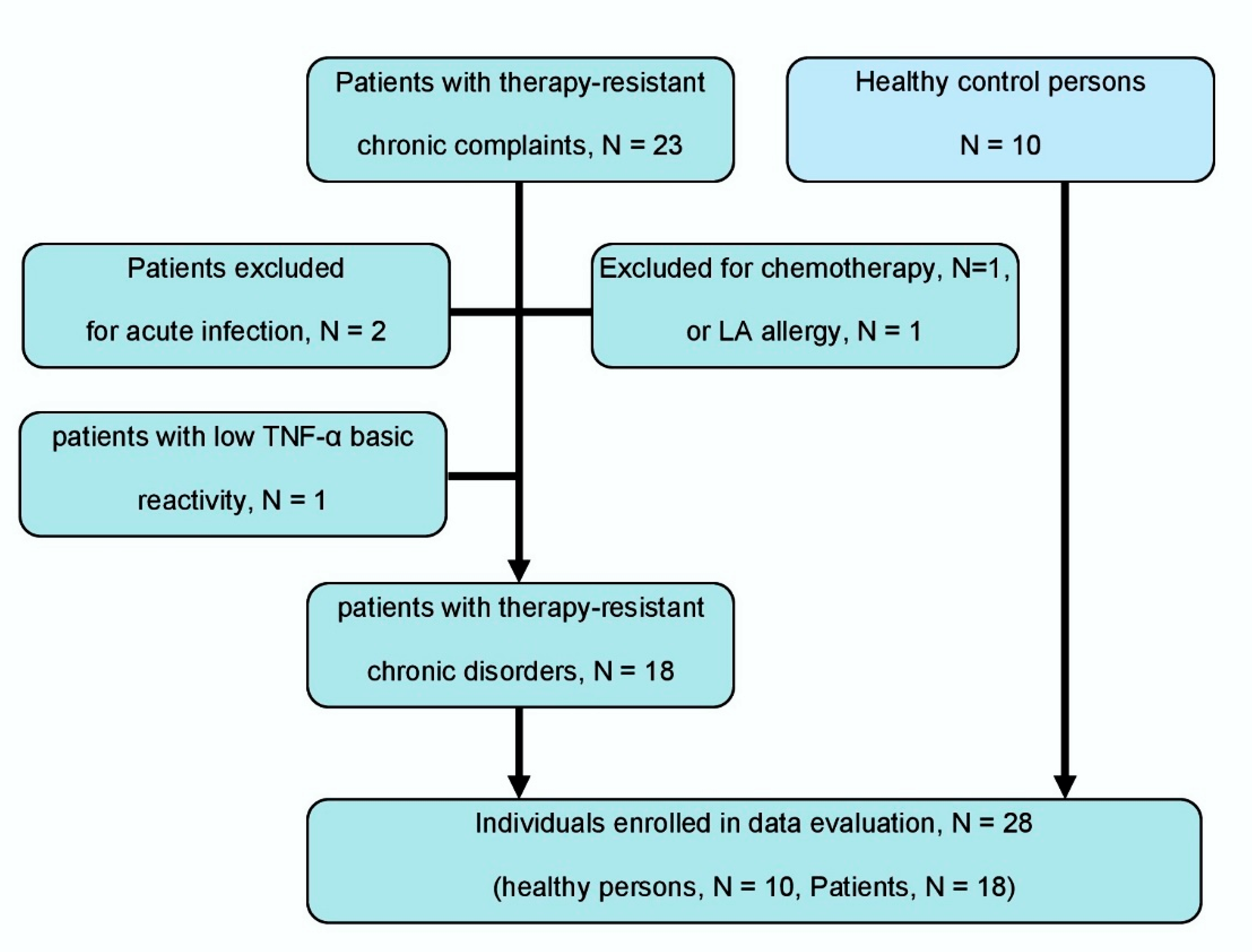
4.1. Dose-Finding Pre-Study and Individuals Tested
4.2. Whole Blood Stimulation Test
4.3. Acquisition of Covariates
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARDS | acute respiratory distress syndrome |
| BMI | body mass index |
| IKK | inhibitor of κB kinase |
| GPCR | G-protein coupled receptor |
| LAs | Local Anesthetics |
| LPS | lipopolysaccharide |
| MAPK | mitogen activated protein kinase |
| NAS | nominal analogue scale |
| SARS | severe acute respiratory syndrome |
| TLA | therapy with Local Anesthetics, Therapeutic Local Anesthesia |
| TLR | toll-like receptor |
| TLR4 | toll-like receptors 4 |
| TNF-α | tumor necrosis factor-alpha |
| VGSC | voltage-gated sodium channels |
References
- Cassuto, J.; Sinclair, R.; Bonderovic, M. Anti-inflammatory properties of local anesthetics and their present and potential clinical implications. Acta Anaesthesiol. Scand 2006, 50, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Weinschenk, S. Neural Therapy—A review of the therapeutic use of local anaesthetics. Acupunct Rel. 2012, 4, 25–29. [Google Scholar]
- Layer, P.; Bronisch, H.-J.; Henniges, U.M.; Koop, I.; Kahl, M.; Dignass, A.; Ell, C.; Freitag, M.; Keller, J. Effects of systemic administration of a local anesthetic on pain in acute pancreatitis: A randomized clinical trial. Pancreas 2011, 40, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Bjorck, S.; Dahlstrom, A.; Ahlman, H. Treatment of distal colitis with local anaesthetic agents. Pharm. Toxicol. 2002, 90, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Herroeder, S.; Pecher, S.; Schönherr, M.E.; Kaulitz, G.; Hahnenkamp, K.; Friess, H.; Böttiger, B.W.; Bauer, H.; Dijkgraaf, M.G.W.; Durieux, M.E.; et al. Systemic lidocaine shortens length of hospital stay after colorectal surgery: A double-blinded, randomized, placebo-controlled trial. Ann. Surg. 2007, 246, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 95, 1033–1034. [Google Scholar] [CrossRef]
- Lahat, A.; Horin, S.B.; Lang, A.; Fudim, E.; Picard, O.; Chowers, Y. Lidocaine down-regulates nuclear factor-kappaB signalling and inhibits cytokine production and T cell proliferation. Clin. Exp. Immunol. 2008, 152, 320–327. [Google Scholar] [CrossRef]
- Yuan, T.; Li, Z.; Li, X.; Yu, G.; Wang, N.; Yang, X. Lidocaine attenuates lipopolysaccharide-induced inflammatory responses in microglia. J. Surg. Res. 2014, 192, 150–162. [Google Scholar] [CrossRef]
- Lang, A.; Horin, S.B.; Picard, O.; Fudim, E.; Amariglio, N.; Chowers, Y. Lidocaine inhibits epithelial chemokine secretion via inhibition of nuclear factor kappa B activation. Immunobiology 2010, 215, 304–313. [Google Scholar] [CrossRef]
- Herrlich, P.; Karin, M.; Weiss, C. Supreme EnLIGHTenment: Damage recognition and signaling in the mammalian UV response. Mol. Cell 2008, 29, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, Q.; Qi, Z.; Zheng, X. Lidocaine protects against renal and hepatic dysfunction in septic rats via downregulation of Tolllike receptor 4. Mol. Med. Rep. 2014, 9, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, M.; Li, S.; Wu, H.; Shen, Q.; Zhang, S.; Fang, L.; Liu, R. Nebulized lidocaine ameliorates allergic airway inflammation via downregulation of TLR2. Mol. Immunol. 2018, 97, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.; Marrero-Berrios, I.; Weinberg, J.; Manchikalapati, D.; SchianodiCola, J.; Schloss, R.; Yarmush, J. The effect of local anesthetic on pro-inflammatory macrophage modulation by mesenchymal stromal cells. Int. Immunopharmacol. 2016, 33, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Garutti, I.; Rancan, L.; Simón, C.; Cusati, G.; Sanchez-Pedrosa, G.; Moraga, F.; Olmedilla, L.; Lopez-Gil, M.T.; Vara, E. Intravenous lidocaine decreases tumor necrosis factor alpha expression both locally and systemically in pigs undergoing lung resection surgery. Anesth. Analg. 2014, 119, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Gaestel, M.; Kotlyarov, A.; Kracht, M. Targeting innate immunity protein kinase signalling in inflammation. Nat. Rev. Drug Discov. 2009, 8, 480–499. [Google Scholar] [CrossRef] [PubMed]
- Coussens, M.L.; Zitvogel, L.; Palucka, A.K. Neutralizing tumor-promoting chronic inflammation: A magic bullet? Science 2013, 339, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.H.; Chiang, B.L.; Chang, S.C.; Ho, H.N.; Wang, J.T.; Chen, Y.C.; Hsiao, C.H.; Hseuh, P.R.; Chie, W.C.; Yang, P.C. Clinical manifestations and inflammatory cytokine responses in patients with severe acute respiratory syndrome. J. Formos. Med. Assoc. 2005, 104, 715–723. [Google Scholar]
- Piegeler, T.; Votta-Velis, E.G.; Liu, G.; Place, A.T.; Schwartz, D.E.; Beck-Schimmer, B.; Minshall, R.D.; Borgeat, A. Antimetastatic potential of amide-linked local anesthetics: Inhibition of lung adenocarcinoma cell migration and inflammatory Src signaling independent of sodium channel blockade. Anesthesiology 2012, 117, 548–559. [Google Scholar] [CrossRef] [Green Version]
- Grandhi, K.R.; Perona, B. Mechanisms of Action by Which Local Anesthetics Reduce Cancer Recurrence: A Systematic Review. Pain Med. 2019, 21, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Grage, S.L.; Culetto, A.; Culetto, A.S.; Weinschenk, S. Membrane-Mediated Activity of Local Anesthetics. Mol. Pharmacol. 2021, 100, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Picardi, S.; Stevens, M.F.; Hahnenkamp, K.; Durieux, M.E.; Lirk, P.; Hollmann, M.W. Time-dependent modulation of muscarinic m1/m3 receptor signalling by local anaesthetics. Br.J. Anaesth. 2014, 112, 370–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahnenkamp, K.; Durieux, M.E.; Hahnenkamp, A.; Schauerte, S.K.; Hoenemann, C.W.; Vegh, V.; Theilmeier, G.; Hollmann, M.W. Local anaesthetics inhibit signalling of human NMDA receptors recombinantly expressed in Xenopus laevis oocytes: Role of protein kinase C. Br. J. Anaesth. 2006, 96, 77–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, M.W.; McIntire, W.E.; Garrison, J.C.; Durieux, M.E. Inhibition of mammalian Gq protein function by local anesthetics. Anesthesiology 2002, 97, 1451–1457. [Google Scholar] [CrossRef] [PubMed]
- Puopolo, M.; Binshtok, A.M.; Yao, G.-L.; Oh, S.B.; Woolf, C.J.; Bean, B.P. Permeation and block of TRPV1 channels by the cationic lidocaine derivative QX-314. J. Neurophysiol. 2013, 109, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Picardi, S.; Cartellieri, S.; Groves, D.; Hahnenekamp, K.; Gerner, P.; Durieux, M.E.; Stevens, M.F.; Lirk, P.; Hollmann, M.W. Local anesthetic-induced inhibition of human neutrophil priming: The influence of structure, lipophilicity, and charge. Reg. Anesth. Pain Med. 2013, 38, 9–15. [Google Scholar] [CrossRef]
- Hollmann, M.W.; Durieux, M.E. Local anesthetics and the inflammatory response: A new therapeutic indication? Anesthesiology 2000, 93, 858–875. [Google Scholar] [CrossRef]
- Hollmann, M.W.; Fischer, L.G.; Byford, A.M.; Durieux, M.D. Local anesthetic inhibition of m1 muscarinic acetylcholine signaling. Anesthesiology 2000, 93, 497–509. [Google Scholar] [CrossRef]
- Sinclair, R.; Eriksson, A.S.; Gretzer, C.; Thomsen, M.D. Inhibitory effects of amide local anaesthetics on stimulus-induced human leukocyte metabolic activation, LTB4 release and IL-1 secretion in vitro. Acta Anaesthesiol. Scand 1993, 37, 159–165. [Google Scholar] [CrossRef]
- Hollmann, M.W.; Wieczorek, K.S.; Berger, A.; Durieux, M.E. Local anesthetic inhibition of G protein-coupled receptor signaling by interference with Galpha(q) protein function. Mol. Pharmacol. 2001, 59, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.H.; Kennedy, J.L.; Ou, D.W. Inhibition of cytokine release by cocaine. Int. J. Immunopharmacol. 1994, 16, 295–300. [Google Scholar] [CrossRef]
- Kindler, C.H.; Paul, M.; Zou, H.; Liu, C.; Winegar, B.D.; Gray, A.T.; Yost, C.S. Amide local anesthetics potently inhibit the human tandem pore domain background K+ channel TASK-2 (KCNK5). J. Pharmacol. Exp. Ther. 2003, 306, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papavlassopoulos, M.; Stamme, C.; Thon, L.; Adam, D.; Hillemann, D.; Seydel, U.; Schromm, A.B. MaxiK blockade selectively inhibits the lipopolysaccharide-induced I kappa B-alpha /NF-kappa B signaling pathway in macrophages. J. Immunol. 2006, 177, 4086–4093. [Google Scholar] [CrossRef] [Green Version]
- Hollmann, M.W.; Herroeder, S.; Kurz, K.S.; Hoenemann, C.W.; Struemper, D.; Hahnenkamp, K.; Durieux, M.E. Time-dependent inhibition of G protein-coupled receptor signaling by local anesthetics. Anesthesiology 2004, 100, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Boneberg, M.E.; Hartung, T. Mistletoe lectin-1 increases tumor necrosis factor-alpha release in lipopolysaccharide-stimulated whole blood via inhibition of interleukin-10 production. J. Pharmacol. Exp. Ther. 2001, 298, 996–1000. [Google Scholar] [PubMed]
- Keiss, H.P.; Dirsch, V.M.; Hartung, T.; Haffner, T.; Trueman, L.; Auger, J.; Kahane, R.; Vollmar, A.M. Garlic (Allium sativum L.) modulates cytokine expression in lipopolysaccharide-activated human blood thereby inhibiting NF-kappaB activity. J. Nutr. 2003, 133, 2171–2175. [Google Scholar] [CrossRef] [PubMed]
- Kiemer, A.K.; Hartung, T.; Huber, C.; Vollmar, A.M. Phyllanthus amarus has anti-inflammatory potential by inhibition of iNOS, COX-2, and cytokines via the NF-kappaB pathway. J. Hepatol. 2003, 38, 289–297. [Google Scholar] [CrossRef]
- Finnerty, T.D.; Buggy, D.J. A novel role for lidocaine in COVID-19 patients? Br. J. Anaesth. 2020, 125, e391–e394. [Google Scholar] [CrossRef]
- Takao, Y.; Mikawa, K.; Nishina, K.; Maekawa, N.; Obara, N. Lidocaine attenuates hyperoxic lung injury in rabbits. Acta Anaesthesiol. Scand 1996, 40, 318–325. [Google Scholar] [CrossRef]
- Weinschenk, S.; Benrath, J. Wie gefährlich ist Procain? Eine Stellungnahme unter Bezug auf die aktuelle wissenschaftliche Evidenz. Dtsch Z Akupunkt. 2019, 43, 101–104. [Google Scholar]
- Bossuyt, P.M.; Reitsma, J.B.; Bruns, D.E.; Gatsonis, C.A.; Glasziou, P.P.; Irwig, L.M.; Lijmer, J.G.; Moher, D.; Rennie, D.; de Vet, H.C.M. Towards complete and accurate reporting of studies of diagnostic accuracy: The STARD initiative. BMJ 2003, 326, 41–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlins, P.; Mander, T.; Sadeghi, R.; Hill, S.; Gammon, G.; Foxwell, B.; Wrigley, S.; Moore, M. Inhibition of endotoxin-induced TNF-alpha production in macrophages by 5Z-7-oxo-zeaenol and other fungal resorcylic acid lactones. Int. J. Immunopharmacol. 1999, 21, 799–814. [Google Scholar] [CrossRef]
- Hartel, C.; Puttkamer, J.V.; Gallner, F.; Strunk, T.; Schultz, C. Dose-dependent immunomodulatory effects of acetylsalicylic acid and indomethacin in human whole blood: Potential role of cyclooxygenase-2 inhibition. Scand J. Immunol. 2004, 60, 412–420. [Google Scholar] [CrossRef]
- Ouagued, M.; Martin-Chouly, C.A.E.; Brinchault, G.; Leportier-Comoy, C.; Depincé, A.; Bertrand, C.; Lagente, V.; Belleguic, C.; Pruniaux, M.P. The novel phosphodiesterase 4 inhibitor, CI-1044, inhibits LPS-induced TNF-alpha production in whole blood from COPD patients. Pulm Pharmacol. Ther. 2005, 18, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Zhan, J. Effects of remifentanyl and fentanyl on LPS-induced cytokine release in human whole blood in vitro. Mol. Biol. Rep. 2009, 36, 1113–1117. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 21. [Google Scholar] [CrossRef]
- R-Core-Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]

{kind=link}
{kind=link}
| Covariate | All Individuals | Patients, N = 18 | Healthy, N = 10 | p Value |
|---|---|---|---|---|
| Number N = | 28 | 18 | 10 | --- |
| Age (years) | 49.9 (18.8–79.3) | 62.5 (42.4–79.3) | 34.8 (18.8–49.8) | <0.001 |
| Sex = female | 20 (71.4%) | 12 (66.7%) | 8 (80%) | 0.454 |
| Body mass index | 24.2 (19.4–30.1) | 24.2 (19.4–30.1) | 24.1 (20.4–25.4) | 0.632 |
| Chronic pain disorder | 13 | 13 | 0 | --- |
| Recurrent inflammation | 8 | 8 | 0 | --- |
| Serum TNF-α > 8.5 pg/mL | 2 | 2 | n. a. | --- |
| Person | Patient/Control | Bupivacaine (%) | Lidocaine (%) | Mepivacaine (%) | Procaine (%) |
|---|---|---|---|---|---|
| 1 | Patient | 75 | 61 | 66 | 82 |
| 2 | Patient | 126 | 108 | 87 | 94 |
| 3 | Patient | 121 | 136 | 106 | 93 |
| 4 | Patient | 63 | 65 | 100 | 101 |
| 5 | Patient | 61 | 67 | 65 | 77 |
| 6 | Patient | 90 | 90 | 109 | 117 |
| 7 | Patient | 81 | 96 | ||
| 8 | Patient | 85 | 48 | 43 | 49 |
| 9 | Patient | 72 | 83 | 74 | 103 |
| 10 | Patient | 93 | 96 | 82 | 104 |
| 11 | Patient | 144 | 92 | 87 | 82 |
| 12 | Patient | 73 | 105 | 54 | |
| 13 | Patient | 121 | 103 | 97 | 109 |
| 14 | Patient | 49 | 85 | 80 | |
| 15 | Patient | 79 | 81 | 90 | |
| 16 | Patient | 96 | 88 | 137 | |
| 17 | Patient | 95 | 59 | 82 | 80 |
| 18 | Patient | 134 | 20 | 52 | 68 |
| 19 | Test person | 92 | 51 | 98 | 54 |
| 20 | Test person | 200 | 69 | 166 | 74 |
| 21 | Test person | 106 | 77 | 100 | 60 |
| 22 | Test person | 57 | 155 | 60 | 81 |
| 23 | Test person | 92 | 98 | 117 | 38 |
| 24 | Test person | 54 | 43 | 96 | 61 |
| 25 | Test person | 77 | 76 | 102 | 122 |
| 26 | Test person | 69 | 83 | 56 | 88 |
| 27 | Test person | 86 | 50 | 85 | 133 |
| 28 | Test person | 96 | 110 | 93 | 68 |
| Bupivacaine | Lidocaine | Mepivacaine | Procaine | p-Value | |
|---|---|---|---|---|---|
| All persons | 44.4% | 61.5% | 44.4% | 50.0% | 0.562 |
| Patients | 47.1% | 56.3% | 58.8% | 37.5% | 0.607 |
| Healthy individuals | 40.0% | 70.0% | 20.0% | 70.0% | 0.066 |
| p-value (between groups) | 0.722 | 0.483 | 0.049 | 0.107 |
| Variable | Bupivacaine | Lidocaine | Mepivacaine | Procaine | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NR | Resp | p Value | NR | Resp | p Value | NR | Resp | p Value | NR | Resp | p Value | |
| Age (mean) | 48.49 | 47.14 | 0.981 | 52.10 | 43.94 | 0.384 | 43.17 | 53.79 | 0.113 | 59.22 | 41.97 | 0.013 * |
| Sex (% female) | 80.00 | 66.67 | 0.432 | 70.00 | 68.75 | 0.946 | 66.67 | 83.33 | 0.326 | 69.23 | 69.23 | 1.000 |
| BMI (mean) | 23.97 | 23.86 | 0.884 | 24.62 | 23.86 | 0.493 | 24.00 | 23.82 | 0.393 | 24.59 | 23.58 | 0.342 |
| Patients vs. Controls (% Pat) | 60.00 | 66.67 | 0.722 | 70.00 | 56.25 | 0.483 | 46.67 | 83.33 | 0.049 * | 76.92 | 46.15 | 0.107 |
| Type of Complaints (% with inflammation) | 12.50 | 50.00 | 0.124 | 33.33 | 42.86 | 0.725 | 16.67 | 37.5 | 0.393 | 25.00 | 20.00 | 0.835 |
| Duration of Complaints (mean) | 9.04 | 14.15 | 0.401 | 9.10 | 12.69 | 0.617 | 10.62 | 11.69 | 0.948 | 11.54 | 11.32 | 0.826 |
| Serum TNF-α (mean) | 5.63 | 7.83 | 0.155 | 6.12 | 6.30 | 1.000 | 6.20 | 6.94 | 1.000 | 7.086 | 6.18 | 0.607 |
| Bupivacaine | Basal | 1:2 | 1:5 | 1:10 | 1:20 | 1:40 | 1:80 | 1:160 |
|---|---|---|---|---|---|---|---|---|
| P1 | 184 | 23.5 * | 94.3 | -- | 99.4 | -- | -- | 177 |
| P2 | 402 | 394 | 294 | -- | 308 | -- | -- | 506 |
| P3 | 1844 | 102 * | 99.4 * | -- | 1276 | -- | -- | 1642 |
| P4 | 1216 | 19.3 * | 943 | -- | 1044 | -- | -- | 1038 |
| P5 | 833 | 66.2 * | 41.3 * | -- | 622 | -- | -- | 772 |
| P6 | 1034 | 955 | 983 | -- | 1018 | -- | -- | 965 |
| Lidocaine | Basal | 1:2 | 1:5 | 1:10 | 1:20 | 1:40 | 1:80 | 1:160 |
| P1 | 263 | -- | -- | 183 * | 244 | 387 | 304 | 299 |
| P2 | 293 | -- | -- | 169 * | 293 | 244 | 214 | 292 |
| P3 | 1183 | -- | -- | 926 | 1296 | 990 | 1001 | 1077 |
| P4 | 943 | -- | -- | 656 | 701 | 632 | 692 | 891 |
| P5 | 891 | -- | -- | 93 * | 782 | 621 | 606 | 837 |
| P6 | 772 | -- | -- | 523 | 479 | 417 | 407 | 622 |
| P7 | 875 | -- | 6,7 * | 662 | 669 | -- | 933 | -- |
| P8 | 1844 | -- | 1021 | 1737 | 1528 | -- | 1793 | -- |
| P9 | 1216 | -- | 481 | 617 | 613 | -- | 1027 | -- |
| P10 | 832 | -- | 11.8 * | 902 | 915 | -- | 892 | -- |
| Mepivacaine | Basal | 1:2 | 1:5 | 1:10 | 1:20 | 1:40 | 1:80 | 1:160 |
| P1 | 191 | -- | -- | 79.1 * | 102 | 155 | 142 | 177 |
| P2 | 281 | -- | -- | 170 | 214 | 231 | 219 | 226 |
| P3 | 286 | -- | -- | 256 | 214 | 291 | 367 | 341 |
| P4 | 723 | -- | -- | 154 * | 326 | 344 | 329 | 487 |
| P5 | 1735 | -- | -- | 943 | 952 | 893 | 1236 | 1105 |
| P6 | 1022 | -- | -- | 734 | 693 | 702 | 811 | 1001 |
| P7 | 1230 | 5,7 * | 6.1 * | -- | 742 | -- | 1312 | -- |
| P8 | 522 | 94,2 * | 602 | -- | 612 | -- | 570 | -- |
| P9 | 184 | 18,1 * | 194 | -- | 177 | -- | 191 | -- |
| P10 | 402 | 30,5 * | 402 | -- | 409 | -- | 422 | -- |
| Procaine | Basal | 1:2 | 1:5 | 1:10 | 1:20 | 1:40 | 1:80 | 1:160 |
| P1 | 385 | 325 | 317 | 410 | 364 | 388 | ||
| P2 | 282 | 187 * | 237 | 292 | 299 | 287 | ||
| P3 | 150 | 84 * | 176 | 170 | 173 | 159 | ||
| P4 | 832 | 122 * | 456 | 495 | 422 | 820 | ||
| P5 | 1253 | 788 * | 622 | 612 | 988 | 1014 | ||
| P6 | 928 | 688 | 692 | 701 | 821 | 1011 | ||
| P7 | 801 | -- | 73.2 * | -- | 433 | -- | -- | 501 |
| P8 | 473 | -- | 94.4 * | -- | 348 | -- | -- | 384 |
| P9 | 875 | -- | 422 | -- | 522 | -- | -- | 839 |
| P10 | 1230 | -- | 91.2 * | -- | 992 | -- | -- | 1021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinschenk, S.; Weiss, C.; Benrath, J.; von Baehr, V.; Strowitzki, T.; Feißt, M. Anti-Inflammatory Characteristics of Local Anesthetics: Inhibition of TNF-α Secretion of Lipopolysaccharide-Stimulated Leucocytes in Human Blood Samples. Int. J. Mol. Sci. 2022, 23, 3283. https://doi.org/10.3390/ijms23063283
Weinschenk S, Weiss C, Benrath J, von Baehr V, Strowitzki T, Feißt M. Anti-Inflammatory Characteristics of Local Anesthetics: Inhibition of TNF-α Secretion of Lipopolysaccharide-Stimulated Leucocytes in Human Blood Samples. International Journal of Molecular Sciences. 2022; 23(6):3283. https://doi.org/10.3390/ijms23063283
Chicago/Turabian StyleWeinschenk, Stefan, Carsten Weiss, Justus Benrath, Volker von Baehr, Thomas Strowitzki, and Manuel Feißt. 2022. "Anti-Inflammatory Characteristics of Local Anesthetics: Inhibition of TNF-α Secretion of Lipopolysaccharide-Stimulated Leucocytes in Human Blood Samples" International Journal of Molecular Sciences 23, no. 6: 3283. https://doi.org/10.3390/ijms23063283
APA StyleWeinschenk, S., Weiss, C., Benrath, J., von Baehr, V., Strowitzki, T., & Feißt, M. (2022). Anti-Inflammatory Characteristics of Local Anesthetics: Inhibition of TNF-α Secretion of Lipopolysaccharide-Stimulated Leucocytes in Human Blood Samples. International Journal of Molecular Sciences, 23(6), 3283. https://doi.org/10.3390/ijms23063283