Abstract
Non-alcoholic fatty liver disease (NAFLD) is an ‘umbrella’ term, comprising a spectrum ranging from benign, liver steatosis to non-alcoholic steatohepatitis, liver fibrosis and eventually cirrhosis and hepatocellular carcinoma. NAFLD has evolved as a major health problem in recent years. Discovering ways to prevent or delay the progression of NAFLD has become a global focus. Lifestyle modifications remain the cornerstone of NAFLD treatment, even though various pharmaceutical interventions are currently under clinical trial. Among them, sodium-glucose co-transporter type-2 inhibitors (SGLT-2i) are emerging as promising agents. Processes regulated by SGLT-2i, such as endoplasmic reticulum (ER) and oxidative stress, low-grade inflammation, autophagy and apoptosis are all implicated in NAFLD pathogenesis. In this review, we summarize the current understanding of the NAFLD pathophysiology, and specifically focus on the potential impact of SGLT-2i in NAFLD development and progression, providing current evidence from in vitro, animal and human studies. Given this evidence, further mechanistic studies would advance our understanding of the exact mechanisms underlying the pathogenesis of NAFLD and the potential beneficial actions of SGLT-2i in the context of NAFLD treatment.
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) has become a major health problem worldwide with an increasing prevalence ranging from 13% in Africa to 42% in South-East Asia [1,2]. The term NAFLD includes a variety of diseases, ranging from liver fat deposition in more than 5% of hepatocytes (steatosis—non-alcoholic fatty liver (NAFL)) to necroinflammation and fibrosis (non-alcoholic steatohepatitis (NASH)), which can progress into NASH-cirrhosis, and eventually to hepatocellular carcinoma [3,4]. Determining ways to delay the progression of NAFLD has become a global focus.
Lifestyle modifications—namely improved diet, weight management and increased physical activity—play a fundamental role in the treatment in NAFLD, since more than half of the patients with NAFLD have a high body mass index (BMI) [5,6]. Of note, drugs for specifically treating NAFLD are now being developed/approved, thus management of co-morbidities such as obesity, dyslipidemia or type 2 diabetes mellitus (T2DM) remains the cornerstone for the treatment of NAFLD patients [7,8].
Interestingly, since the cardiometabolic disorders associated with NAFLD account for the increased morbidity and mortality in these patients, the term metabolic (dysfunction) associated fatty liver disease (MAFLD) has been recently proposed by a panel of international experts as more appropriate, reflecting the underlying pathogenesis of the NAFLD spectrum [9,10,11]. Masarone et al., revealed that the prevalence of NAFLD was 94.82% among patients with metabolic syndromes (MS) and was presented in all T2DM patients with elevated transaminases when they performed biopsy [12]. Surprisingly, 58.52% of MS and 96.82% of T2DM patients in this study were diagnosed with NASH. As insulin resistance (IR) is crucially linked with both T2DM and NASH pathophysiology [13], they concluded that NASH may be one of the early clinical manifestations of T2DM [12]. Of note, NAFLD, particularly steatohepatitis, has also been associated with an increased risk of cardiovascular-related mortality regardless of age, sex, smoking habit, cholesterolemia and the remaining elements of MS [11]. Several prospective, observational and cross-sectional studies and meta-analyses demonstrated that NAFLD is associated with enhanced preclinical atherosclerotic damage as well as coronary, cerebral and peripheral vascular events with a negative impact on patients’ outcome [14]. Furthermore, the severity of liver biopsy-based fibrosis has been independently associated with the worsening of both systolic and diastolic cardiac dysfunction [15,16].
Sodium-glucose co-transporter type-2 (SGLT-2) inhibitors (SGLT-2i) are glucose-lowering agents that improve glucose control, whilst promoting weight loss and lowering serum uric acid levels. These agents have shown great advantages even in patients with no diabetes, gaining approval of use in non-diabetic patients with heart failure and chronic kidney diseases [17,18]. In addition to the anti-hyperglycemic effects and their ability to reduce body weight, SGLT-2i seem to exert potent antioxidant and anti-inflammatory effects making them promising candidates for NAFLD treatment.
Indeed, recent data from animal studies and clinical trials have demonstrated beneficial effects of SGLT-2i on fatty liver accumulation, as judged by improvement of biological markers of NAFLD, as well as by imaging techniques, albeit mainly in T2DM patients.
Herein, we provide current insights into the effects of SGLT-2i on the progression of NAFLD, focusing on the underlying mechanisms of action. Endoplasmic reticulum (ER) stress, oxidative stress, low-grade inflammation, autophagy and apoptosis are among the SGLT-2i-regulated processes that have been shown to mediate the beneficial effects of SGLT-2i on NAFLD. Accordingly, we present the available evidence from in vitro, animal and human studies regarding the potential impact of SGLT-2i on NAFLD development and progression.
2. Overview of NAFLD Pathogenesis
The main theory concerning the pathophysiology of NAFLD has changed over time, reflecting the advances in our understanding of this multi-factorial disease. For years, the “two-hit” theory was the prevailing one. According to this theory, the pathophysiology of NAFLD consisted of a first “hit” representing the stage of simple steatosis alone (NAFL), which involves hepatocytic lipid accumulation and hepatic insulin resistance [19], as well as of a second “hit” from other factors (e.g., oxidative stress, ER stress, other injury), which was required for the development and progression of hepatic inflammation (NASH) and fibrosis. Replacing this initial theory, the “multiple parallel-hit” model [20] has been more recently proposed to better explain the complex pathogenesis and progression of NAFLD. According to this theory, as its name suggests, different amalgamations of numerous (epi)genetic and environmental factors, representing “hits”, dynamically interplay with each other, and can drive the development and progression of NAFLD. These factors include specific genetic polymorphisms and epigenetic modifications [21], features of metabolic syndrome [22,23,24,25] (western diet, lack of physical activity, central obesity, dysregulation of adipoknes and IR), lipotoxicity [26,27], dysbiosis of the gut microbiota [26], dysregulation of autophagy and mitochondrial function [28,29,30], ER stress [31], hepatocyte homeostasis and death [32,33], as well as inflammatory and fibrotic responses [34,35]. Notably, when the hepatic capacity to handle the primary metabolic energy substrates is overwhelmed, toxic lipid species accumulate in the liver, leading to hepatocyte dysfunction and apoptosis, along with metabolically triggered inflammation and subsequent fibrosis [36].
The hallmark of NAFLD pathogenesis seems to be an increased adipocyte-like (dys)function of the hepatocytes, when the capacity of adipose tissue to store excess energy from the diet is diminished [37,38,39,40]. In conditions of energy surplus, increased metabolic substrates in tissues [27] drive hepatic de novo lipogenesis [41,42,43], while enhancing IR leading to a vicious cycle. The accumulation of intrahepatic lipid levels are governed by the balance between lipids synthesis, uptake and lysis [44]. Loss of equilibrium between lipogenesis and lipolysis leads to intracellular accumulation of free fatty acids (FFA) and subsequent hepatocellular damage, IR, worsening of liver function, formation of hepatic steatosis and progression to NASH, cirrhosis, and hepatocellular carcinoma [44].
In most individuals with NAFLD, dysregulation of adipokines (e.g., leptin and adiponectin) and metabolic-induced inflammation impair insulin signaling in adipocytes [45,46]. This impairment, in turn, contributes to reduced FA uptake and accelerated lipolysis in subcutaneous adipose tissue, resulting in exces sive delivery to the liver [45,46,47]. When hepatocytes are overloaded with lipids, physiologically minor pathways of β-oxidation in the peroxisomes and the ER are upregulated, thus increasing the hepatocyte reactive oxygen species (ROS) production [41,48] and generating highly reactive aldehyde by-products [49]. This phenomenon leads to nuclear and mitochondrial DNA damage, phospholipid membrane disruption and cell death. Moreover, due to the mitochondrial dysfunction and the consequent impaired β-oxidation found in patients with NAFLD, FA are alternatively esterified and collected in lipid droplets in the ER [50,51], generating toxic lipid metabolites, such as diacylglycerols [24], ceramides [52,53], and lysophosphatidyl choline species [53], which in turn lead to hepatocyte dysfunction (lipotoxicity) [54,55] and ER stress. The adaptive homeostatic mechanism of ER stress, called the unfolded protein response (UPR) [56], is impaired in NAFLD patients and, thus further cell stress-sensors are activated, whilst inflammatory and apoptotic pathways are upregulated [57,58]. In addition, the gut microbiota seem to also play a critical role in NAFL and NASH pathogenesis. Gut-derived pathogens and damage-associated molecular patterns activate an intrahepatic inflammatory process via Toll-like receptor signaling and NLR family pyrin domain containing-3 (NLRP3) inflammasome activation [35,59,60]. Hepatic innate immune cells, including Kupffer cells and dendritic cells, as well as hepatic stellate cells (HSCs) are then activated and the liver parenchyma is progressively infiltrated by recruited neutrophils, monocytes, T-lymphocytes, and macrophages [34,61]. Subsequently, the secreted cytokines and growth factors intensify the inflammatory process and further contribute to the fibrotic process as an ineffective attempt for tissue regeneration [35]. Overall, NAFLD is characterized by a complex pathogenetic mechanism, where various pathways are implicated, making the discovery of a “wonder drug” a difficult task (Figure 1).
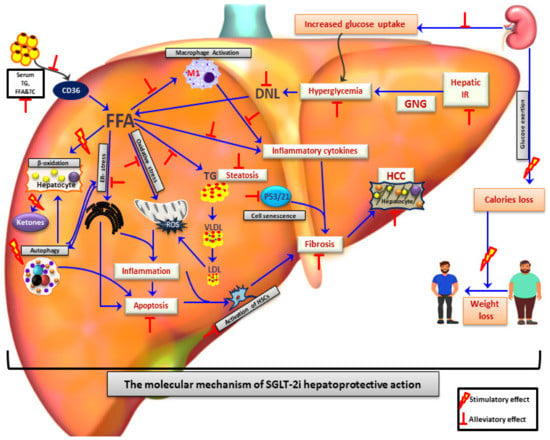
Figure 1.
SGLT-2 inhibitors as a promising therapeutic agents for treatment of NAFLD/NASH patients. SGLT-2i treatment contributes to alleviation of NAFLD by reduction of hyperglycaemia, improvement of systematic insulin resistance, elevation of caloric loss and reduction of body weight mostly due to glycosuria. Apart from that, SGLT-2i play a hepatoprotective effect through reduction of hepatic de novo lipogenesis, hepatic inflammation, apoptosis, ER-stress, oxidative stress, and increase of hepatic beta-oxidation. Reduced activation of hepatic satellite cells and p53/p21 pathways by SGLT-2i leads to amelioration of hepatic fibrosis and HCC development. FFA: Free fatty acids; DNL: De novo lipogenesis; HCC: Hepatocellular carcinoma; TC: Total cholesterol; TG: Triglycerides; LDL: Low density lipoprotein; VLDL: Very low density lipoprotein; GNG: Gluconeogenesis; HSC: Hepatic stellate cells; IR: Insulin resistance; ROS: Reactive oxygen species; ER-stress: Endoplasmic reticulum stress.
3. SGLT-2i Overview
Phlorizin, a b-Dglucoside, was the first natural non-selective SGLT-2i isolated from the bark of the apple tree in 1835 and was initially used for fever relief and treatment of infectious diseases [62,63]. Phlorizin contains a glucose moiety and an aglycone in which two aromatic carbocycles are joined by an alkyl spacer. Approximately fifty years later, scientists observed that phlorizin in high dose lowered plasma glucose levels through glucosuria independently of insulin secretion [62,63,64]. However, the mechanism of phlorizin action through the active glucose transport system of the proximal tubule was revealed in the early 1970s [62,65].
Thereafter, researchers began to discover the exact mechanism of phlorizin action and its potential application in the management of hyperglycaemia. Phlorizin, when delivered orally, undergoes hydrolysis of the O-glycosidic bond by intestinal glucosidases, leading to formation of phloretin, which is capable of GLUT transporter inhibition and uncoupling of oxidative phosphorylation. Due to Phlorizin’s low SGLT-2 selectivity, rapid degradation, inhibition of the ubiquitous glucose transporter 1 (GLUT1), poor intestinal absorption and its gastrointestinal side effects (e.g., diarrhea and dehydration), possibly due to its higher potency for SGLT-1, it failed to be developed as an antidiabetic drug [62,66,67,68].
Nevertheless, in order to overcome the aforementioned side effects of phlorizin, a researcher developed a devphlorizin-based analog with improved bioavailability/stability and more selectivity towards SGLT-2 than SGLT-1. Initially, they focused on the development of O-glucoside analogs such as T-1095, sergliflozin and remogliflozin [69,70]. However, due to their poor stability and incomplete SGLT-2 selectivity, pharmaceutical industries turned to another derivative of phlorizin known as C-glucosides [62].
Since then, there have been numerous attempts to synthesize phlorizin C-glucoside analogs with sufficient potency and selectivity for inhibition of SGLT-2. These attempts led to development of dapagliflozin by Meng et al., in 2008 [71]. Dapagliflozin demonstrated more than 1200-fold higher potency for human SGLT-2 versus SGLT-1. Apart from dapagliflozin, during the next years, several other C-glucoside inhibitors have been developed and approved by United States Food and Drug Administration (FDA). Canagliflozin, characterized by a thiophene derivative of C-glucoside, showed over 400-fold difference in inhibitory activities between human SGLT-2 and SGLT-1 [72]. Thereafter, empagliflozin, which has the highest selectivity for SGLT-2 over SGLT-1 (approximately 2700-fold), was the third agent approved by both the European Medicines Agency (EMA) and the FDA, while other SGLT-2i, namely luseogliflozin and topogliflozin, are approved so far for use only in Japan, and ipragliflozin only in Japan and Russia [73,74,75,76,77].
The structure of phlorizin and currently FDA-approved SGLT-2 inhibitors in the United States, including dapagliflozin, canagliflozin, empagliflozin, and ertugliflozin, along with their chemical formulae and brand names are presented in Figure 2 [78]. Diverse structures of other SGLT-2 inhibitors have been previously disclosed in various articles and in a number of patents [79,80].
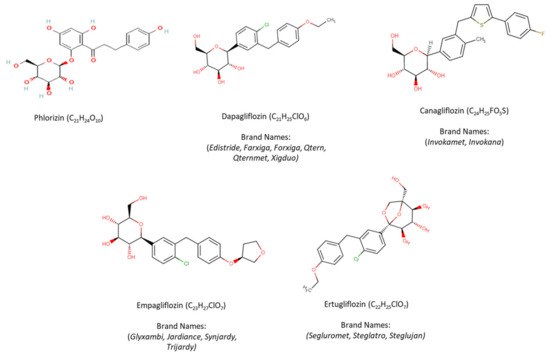
Figure 2.
Structure of phlorizin and FDA-approved SGLT-2 inhibitors.
4. SGLT-2 Inhibitors in NAFLD
4.1. Laboratory Experiments
4.1.1. In Vitro Data
The expression of both SGLT-1 and SGLT-2 co-transporters has been reported in human hepatocellular carcinoma cells (HepG2) [81,82,83], whilst SGLT-2 has also been identified in immortalized human primary hepatocyte cells (HuS-E/2), as well as immortalized normal human hepatocyte-derived liver cells (L02) [84,85].
Studies have indicated that most SGLT-2i exert anti-proliferative activity in several hepatocellular cell lines, through—among other mechanisms—attenuation of glucose uptake [86]. Incubation of both L02 and HepG2 cells with canagliflozin at various concentrations for 24 h resulted in a significant reduction of cell proliferation through an increase in G0/G1 and a decrease in G2/M phase cell population [86]. In addition, an apoptotic effect was exhibited through activation of caspase 3 in HepG2 cells [87]. From a mechanistic point of view, cell growth regulators, cyclins and cyclin-dependent kinases (CDKs) have been indicated as direct targets of SGLT-2i to control proliferation and survival processes. As such, canagliflozin-treated HepG2 cells demonstrated increased expression of the cell growth regulator hepatocyte nuclear factor 4α (HNF4α) [82]. Furthermore, reduced expression of cyclin D1, cyclin D2 and cdk4 leading to cell cycle arrest in a hepatocellular carcinoma (HCC) cell line have been reported after treatment with canagliflozin [82,87]. In another study, incubation of HepG2 cells with canagliflozin resulted in reduced clonogenic cell survival and elevation of anti-carcinogenic potential of γ-irradiation through modulation of ER stress-mediated autophagy and cell apoptosis [88]. On the contrary, trilobatin, a novel SGLT-1/2 inhibitor, increased the HepG2 cell proliferation rate at a dose of 10, 50 and 100 μΜ, while incubation of human HCC cells with tofogliflozin at various concentrations did not alter the HCC cell proliferation rate [89]. Incubation of HepG2 cells with dapagliflozin, canagliflozin and empagliflozin had no effect on cancer cell survival [81,86] and adhesion capacity [81], while the cell sensitivity to dapagliflozin was induced after UDP Glucuronosyltransferase Family 1 Member A9 (UGT1A9) silencing, indicated by an increased number of floating HepG2 cells; of note, UGT1A9 metabolizes and deactivates dapagliflozin [81].
Dapagliflozin treatment remarkably suppressed oleic acid (OA)-induced lipid accumulation and TG content in L02 cells through increased FA β-oxidation, as indicated by elevated proliferator-activated receptor-gamma coactivator-1 alpha (PGC-1α) levels and activation of the AMP-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) signaling pathway [84]. Several in vitro studies have shown that AMPK is a key regulator that mediates the various beneficial effects of SGLT-2i related to cholesterol and glucose metabolism in hepatic cells. AMPK activation by dapagliflozin prevented glucose absorption through reduced SGLT-2 expression in OA-stimulated HuS-E/2 cells [90]. This effect was eliminated after incubation of cells with compound C, a potent AMPK inhibitor [84]. Moreover, treatment of HepG2 cells with canagliflozin facilitated hepatic cholesterol efflux via activation of AMPK. In turn, AMPK activation led to increased expression of the liver X receptor (LXR) and its downstream proteins, resulting in subsequent stimulation of cholesterol reverse transport [91]. LXR activation also accelerates fecal cholesterol disposal through regulation of ATP-binding cassette (ABC) transporters ABCG5 and ABCG8 expression [92]. Canagliflozin treatment resulted in reduced expressions of ABCG5 and ABCG8 and LXR, while this effect was inhibited after treatment of cells with compound C [91]. An increased AMP/ADP ratio leads to AMPK activation. It is known that SGLT-2i reduces cellular ATP levels and indirectly activates the AMPK signaling pathway [83,93]. It has been shown that dapagliflozin alleviates the intracellular ATP levels via regulation of glucose metabolism [84].
Stimulation of hepatic cell lines with palmitic acid (PA) mimics hepatocyte situation in NASH [94,95]. Incubation of PA-stimulated L02 and HepG2 cell lines with dapagliflozin resulted in a significant reduction of intracellular lipid accumulation. This effect was attributed to down-regulation of proteins related to lipid synthesis, up-regulation of genes involved in fatty acid oxidation (e.g., peroxisome proliferator-activated receptor alpha (PPARα) and carnitine palmitoyltransferase 1 (CPT1a)), regulation of AMPK/mTOR pathway and autophagy. Interestingly, incubation of cells with compound C abolished the dapagliflozin beneficial effect on reduction of intracellular lipid accumulation [85]. Collectively, these data suggest that SGLT-2i induced improvement of NAFLD is directly dependent on AMPK signaling activation.
The divergent results of the various relevant studies suggest that the effects of SGLT-2i on cell proliferation, survival and apoptosis—which are important processes in hepatocellular carcinoma development and progression—appear to be drug-, dose- and duration-dependent.
4.1.2. Animal Studies
Overall, the animal models of NAFLD/NASH can be divided into two categories. Of these, one comprises genetically modified animal models, such as leptin deficient (ob/ob), leptin-resistant (db/db), Agouti mutation (KK-Ay), and apolipoprotein E knockout (ApoE−/−) mice, as well as the Prague hereditary hypertriglyceridemic (HHTg) rats, Zucker diabetic fatty (ZDF) rats and Otsuka Long-Evans Tokushima Fatty (OLETF) rat. The other category includes dietary/pharmacological manipulation-induced NAFLD/NASH animal models, such as those fed with either a methionine/choline-deficient (MCD) diet, the trans-fat containing AMLN (amylin liver non-alcoholic steatohepatitis, NASH) diet, and the methionine-reduced, choline-deficient and amino acid defined (CDAA) diet, as well as a high-fat diet (HFD), and a HFD + cholesterol diet or a high-fat, high-calorie (HFHC) diet [30,96,97,98,99,100]. Most of the recent data indicate that both SGLT-1 and SGLT-2 genes are expressed in hepatic tissues of mice and rats [30,85,87,101]. High levels of SGLT-2 protein have also been detected in hepatic macrophages and T cells [101]. While the exact underlying molecular mechanisms of SGLT-2i-induced beneficial effects on NAFLD have not yet been fully clarified, most of our knowledge regarding the potential hepato-protective mechanism(s) of SGLT-2i actions comes from animal studies.
Although the SGLT-2i-induced weight loss seems to play an important role in hepato-protective effects on NAFLD in humans [102,103,104], data from animal studies suggest that other mechanisms are more likely responsible for the noted SGLT-2i-mediated hepato-protection.
Administration of SGLT-2i leads to net loss of calories and consequent attenuation of body weight gain, while reducing the accumulated white adipose tissue [99,105,106,107]. In fact, it has been found that SGLT-2i promote weight loss through improvement of systemic IR, increased body temperature and basal metabolism via regulation of the AMP/ATP ratio [19]. Petito da Silva et al., reported that empagliflozin (Table 1) at a dose of 10 mg/kg/day given for five weeks significantly reduces body weight and body mass in HFD-fed male C57Bl/6 mice, despite a slight increase in their appetite [107]. Similar results were observed in HFD-fed C57BL/6J male mice after administration of canagliflozin (Table 2) at 30 mg/kg dose for four weeks in parallel with improvement of liver function, liver TG content and NAS score [108]. Additionally, short term and low dose administration of dapagliflozin (Table 3) reduced both ALT levels and body weight in both HFD and HFD+MCD diet fed C57BL/6J mice [109]. However, it should be noted that not all studies point towards a beneficial effect of SGLT-2i on body weight. Several studies have shown that administration of SGLT-2i does not exert any significant effect on body weight gain, epididymal fat weight or food intake [30,105,110,111]. In addition, although several studies reported strong effects of low dose ipragliflozin (Table 4) on weight loss [100,112,113], supplementation of an AMLN diet with 40 mg ipragliflozin/kg for 8 weeks did not significantly alter the body weight of AMLN diet fed mice, while it improved liver function, hepatic fibrosis and NA score [114]. Empagliflozin at the dose of 10 mg/kg/day improved liver function and NAFLD status, while it had no significant effect on the body weight of ZDF, HHTg and Wistar rats [115,116], as well as on the body weight and appetite of ApoE knockout mice [30].
Although there are studies showing that administration of SGLT-2i does not exert any significant effect on body weight gain [30,100,110,111,112,113,114], the vast majority of the literature points towards a beneficial effect on weight loss.
However, the protective role of SGLT-2i against NAFLD progression also seems to be mediated by effects beyond those on body weight. Indeed, it has been shown that SGLT-2i treatment improves IR and ameliorates the intracellular FFA, total cholesterol (TC) and TG accumulation through reducing the expression of genes involved in de novo lipogenesis, FA uptake and hepatic TG secretion, whilst it also promotes the expression of key regulatory genes of fatty acid β-oxidation [30,84,100,107,109,112,117]. Specifically, empagliflozin administration has been shown to reduce hepatic triacylglycerol (TAG) levels and improve IR through reduction of lipotoxic intermediates formation, such as diacylglycerols (DAG) [115], which at elevated levels contribute to IR by activating the protein kinase C (PKC)ε pathway [118]. Of note, a recent study by Hüttl et al., demonstrated that reduction of hepatic lipid accumulation following empagliflozin treatment is mediated via increased nuclear factor erythroid 2-related factor 2 (Nrf2), fibroblast growth factor 21 (FGF21) and altered gene expression of the P450 (CYP) enzyme superfamily of cytochrome [115]. Interestingly, Nrf2 and FGF21 regulate lipid metabolism through inhibiting lipogenesis and improving insulin sensitivity, respectively [119,120].
Sterol regulatory element-binding transcription factor 1 (SREBP1) is one of the key regulators of hepatic lipogenesis and plays a crucial role in the regulation of lipogenic genes, such as fatty acid synthase (Fasn), acetyl-CoAcarboxylase 1 (Acc1), and stearoyl-CoA desaturase1 (Scd-1). Increased SREBP1 expression has been linked to the aggravation of hepatic steatosis [121]. While reduced expression of SREBP1 and Scd-1 by both canagliflozin and empagliflozin has been observed in several studies [30,107,108,115], Kern et al., found that empagliflozin treatment for eight weeks does not markedly affect the Scd-1 levels in db/db mice [122]. Dapagliflozin (1 mg/kg/day) treatment for nine weeks resulted in down-regulation of Scd-1 gene expression, as well as reduced ACC1 phosphorylation in ZDF rats. Taking into account that SREBP1 activity is directly regulated by mTOR signaling [123], reduced expression of SREBP1 and its downstream lipogenic targets could be due to the reduced mTOR expression/activity observed after SGLT-2i administration [84,123].
Peroxisome proliferator-activated receptor gamma (PPAR-γ) upregulation triggers de novo lipogenesis and consequent deposition of lipid droplets into hepatocytes [124]. Eight weeks of dapagliflozin treatment reduced hepatic weight and prevented progression of hepatic steatosis compared to mice treated with insulin glargine [125]. This effect was found to be mediated through decreased expression of PPAR-γ targeted genes involved in fatty acid synthesis, such as Scd-1, monoacylglycerol O-acyltransferase 1 (Mogat1), Cell death inducing DFFA like effector A (Cidea) and cell death inducing DFFA like effector C (Cidec), without affecting insulin sensitivity, liver TC or oleate content [125].
SGLT-2i have been also shown to reverse the HFD-induced down regulation of genes involved in FA β-oxidation and lipolysis in the liver of various mouse models of NAFLD [30,84,107,108]. Indeed, it has been shown that expression of PPARα is induced by SGLT-2i [84,105,107,114]. PPARα activation is predominant for regulation of genes related to the FA β-oxidation process in mitochondria, such as peroxisomal acyl-CoA oxidase1 (ACOX1) and enoyl-CoA hydratase, CPT1, and cytochrome-mediated (CYP4A1 and CYP4A3) [44]. Ipragliflozin treatment has been shown to result in acceleration of β-oxidation and export of very-low-density lipoprotein (VLDL) through upregulation of PPARα, CPT1A, and Microsomal Triglyceride Transfer Protein (MTTP) gene expression in the liver of AMLN-fed C57BL/6J mice. MTTP plays a crucial role in export of hepatic LDL [114]. Similarly, dapagliflozin attenuated hepatic lipid accumulation in ZDF rats via up-regulation of the FA β-oxidation enzyme ACOX1 [85]. Tofogliflozin administration induced the expression of genes related to FA β-oxidation in non-tumorous liver lesions, while it had no effect on their expression in tumorous liver tissues [126]. However, not all studies show a promotive effect of SGLT-2i on the expression of enzymes and transcription factors involved in the regulation of FA β-oxidation [107,114,126]. Of note, the expression of PPARα remained unaffected in both Wistar and HHTg rats after eight weeks of empagliflozin administration [107,115]. Administration of canagliflozin at doses of 10 and 20 mg/kg/day ameliorated the HFD-induced down-regulation of the expression of the key regulatory molecule of hepatic lipolysis, known as zinc alpha 2 glycoprotein (ZAG). When the same SGLT-2i was given in KK-Ay mice, it reduced the accumulation of lipid droplets in the liver by increasing hepatic prostaglandin E2 (PGE2) protein levels [111]. Furthermore, the combined therapy with canagliflozin and exercise exerted an additive effect on hepatic PPARγ Coactivator 1 Beta (PGC1b) elevation, and reduced expression of hepatic lipogenic genes, such as Scd1 [108]. Notably, PGC1 plays a central role in the control of hepatic gluconeogenesis, while its overexpression has been associated with stimulation of hepatic FA oxidation [127].
The inhibition of de novo lipogenesis, and enhanced lipolysis and β-oxidation by SGLT-2-i lead to a consequent reduction in oxidative stress, hepatic inflammation and apoptosis [128]. Administration of canagliflozin to HFD-fed diabetic Wistar adult male rats resulted in reduced hepatic steatosis through amelioration of oxidative stress, inflammation and apoptosis, as indicated by reduced plasma malondialdehyde (MDA) levels, serum tumor necrosis factor (TNF) and caspase-3 levels, as well as hepatic expression of interleukin-6 (IL-6) [128,129]. Moreover, dapagliflozin treatment reduced the expression of hepatic inflammatory cytokine (TNF-α, IL-1β, IL-18) content and improved hepatic steatosis in HCHF fed male Wistar rats [130]. Tahara et al., have also shown anti-oxidative stress and anti-inflammatory effects of ipragliflozin on HFD-fed and streptozotocin–nicotinamide-induced type 2 diabetic mice [131]. Similarly, dapagliflozin treatment at a dose of 1 mg/kg/day decreased hepatic ROS production of myeloperoxidase (MPO) and F4/80, thus ameliorating serum ALT levels and hepatic fibrosis [132]. MPO is a chlorinating oxidant-generating enzyme that regulates the initiation of an acute inflammatory response and promotes the development of chronic inflammation through oxidant production [133]. Furthermore, canagliflozin treatment of F344 rats for 16 weeks resulted in reduced hepatic ROS production and MDA and 8-hydroxy-2′-deoxyguanosine (8-OHdG) levels [110]. MDA and 8-OHdG are known biomarkers of lipid peroxidation and oxidative DNA damage, respectively. Of note, canagliflozin and teneligliptin combination therapy showed a stronger effect on reduction of hepatic oxidative stress and amelioration of hepatic inflammation [110]. Treatment with both remogliflozin and ipragliflozin also reduced oxidative stress levels, as evaluated by decreased thiobarbituric acid reactive substances (TBARS) levels [100,134]. Instead, administration of dapagliflozin at 1 mg/kg/twice/day for four weeks had no significant effect either on hepatic TBARS and TG levels or on plasma ALT levels [135]. In addition, reduction of oxidative stress by empagliflozin treatment resulted in amelioration of hepatic inflammation [30,107,116] and steatosis, as judged by down-regulation of inflammatory markers [30,107,115]. Of interest, decreased macrophage infiltration expression [107,117,134] as well as elevation of autophagy markers [30,117] have also been linked to the anti-inflammatory actions of SGLT-2-i in the liver. Specifically, oxidative stress was reduced after empagliflozin treatment via up regulation of Nrf2, which led to increased antioxidant enzyme activity (SOD) and reduced hepatic inflammation [115]. Empagliflozin exerted anti-inflammatory actions in diet-induced obese mice and NASH mouse models through, at least in part, suppression of hepatic nuclear factor kappa-light-chain-enhancer of activated B cells (NFκΒ), monocyte chemoattractant protein-1 (MCP-1) and TNF-α expression, as well as inhibition of the IL17/IL23 axis [115,117,136]. Reduced hepatic inflammation has also been reported in mice and rats after treatment with low dose (1 mg/kg/day) dapagliflozin and ipragliflozin. Indeed, ipragliflozin administration for 12 weeks alleviated hepatic fibrosis through reduced expression of pro-inflammatory markers Emr1 and Itgax in non-tumoric lesions [126]. On the contrary, a recent study in mice by Hupa-Breier et al., demonstrated that four weeks treatment of diet-induced NASH with empagliflozin alone at a dose of 10 mg/kg/day did not exert any beneficial effect on NASH, while it significantly increased expression of pro-inflammatory and pro-fibrotic genes. In the same study, the combination of dulaglutide (a GLP-1 agonist) and empagliflozin exhibited a hepato-protective effect on diabetic background mice through modulation of the pro-inflammatory immune response and microbiome dysbiosis [137].
Meng et al., demonstrated that induction of AMPK/mTOR activity is dominant pathway in empagliflozin-induced beneficial effects on liver inflammation and ALT levels [117]. Dapagliflozin treatment also induced AMPK/mTOR pathway activation through phosphorylation of liver kinase B1 (LKB1; a ubiquitously expressed master serine/threonine kinase that activates downstream kinases of the AMPK pathway) [105,138]. Previous studies have demonstrated that AMPK activation is required for SGLT-2i induced autophagy-dependent lipid catabolism [30,117]. Both dapagliflozin and empagliflozin promote hepatic autophagy through increased AMPK phosphorylation and BECLIN gene expression, as well as reduced P62 levels and mTOR levels and activity [30,85].
Apart from impaired autophagy, ER stress is also implicated in the development of steatosis and the progression of NAFLD/NASH [30,107]. Consistent with the regulatory role of ER stress in the autophagy process, Nasiri-Ansari et al., demonstrated that stimulation of autophagy by empagliflozin leads to amelioration of ER stress and reduced hepatic cell apoptosis [30]. In particular, empaglifilozin administration protects against HFD-induced NAFLD through inhibiting all three branches of ER stress, namely inositol-requiring enzyme 1 (IRE1a), X-box binding protein 1 (Xbp1), activating transcription factor 4 (ATF4), C/EBP homologous protein (CHOP) and activating transcription factor 6 (ATF6) [30,107]. A recent study by Chen et al., also showed that down-regulation of the ATF6 signaling diminished ER stress-induced inflammation and apoptosis of hepatic cells [139]. The synergistic ER stress response and autophagy process regulated hepatic cell apoptosis in HFD-fed ApoE−/− mice with NASH, by reducing cleaved caspase 3 levels and elevation of B-cell lymphoma-2 (Bcl-2)/BCL2 Associated X (Bax) ratio [30]. Of note, canagliflozin was previously found to exert an anti-apoptotic effect on HFD-fed mice as revealed by robust Bcl-2 hepatic expression [129].
Chronic liver inflammation leads to transformation of hepatic stellate cells to myofibroblasts, thus contributing to liver fibrosis and progression of NAFLD to NASH [140]. Transforming growth factor beta (TGF-β) is known as the most potent inducer of liver fibrosis. Administration of canagliflozin for 16 and 20 weeks reduced the expression of hepatic TGF-β in both F344 rats and MC4R-KO mice, respectively [101,110]. Interestingly, TGF-β activation leads to increased fibronectin and collagen types I, II, and IV production [141]. Treatment of mice with both empagliflozin and canagliflozin resulted in reduced expression of different types of collagen, implying synergistic action(s) [87,137]. Luseogliflozin also exerts anti-fibrotic effects through reduction of collagen1a1, collagen1a2, TGF-β and smooth muscle actin (SMA) expression [142]. Extensive data exist regarding the crucial role of tissue inhibitors of metalloproteinases (TIMPs) in the progression of hepatic fibrosis. Reduced expression of TIMPs and amelioration of hepatic fibrosis have been observed after luseogliflozin and canagliflozin administration [101,126,142]. On the contrary, empagliflozin treatment did not affect the expression of stellate cell functionality markers, such as galectin-3, a-SMA, and collagen1a1 [99].
NASH has been associated with an increased risk of cirrhosis and HCC development. Hepatic tumorogenesis is the result of DNA instability caused by several factors, such as hepatic lipotoxicity, aberrant metabolism and inflammation [141,143]. As a protective effect of SGLT-2i on NAFLD/NASH progression has been shown in the vast majority of the studies, further research aimed to evaluate the effect of SGLT-2i on development and progression of HCC [87,101,142]. Activation of the hepatic P53/P21 signaling pathway is positively correlated with induced fibrosis and hepatocellular carcinogenesis. A 14-week treatment with tofogliflozin has been shown to effectively reduce the P21 expression, alleviating the progression of NASH through, at least in part, mitigating genes related to the hepatocyte cellular senescence-associated secretory phenotype [89]. Daily administration of canagliflozin prevented the occurrence of HCC in both STAM mice—a NASH model—(11 weeks) and HFD-fed MC4R-KO mice (52 weeks), as indicated by a significantly reduced number of hepatic tumorous lesions [87,101]. Moreover, 66 weeks administration of canagliflozin to CDAA-fed F344 rats exerted an anti-carcinogenic effect, as indicated by a reduced number of cells positively stained for placental glutathione S-transferase (a prominent marker of hepatocarcinogenesis) [110]. Finally, canagliflozin administration reduced the development of hepatic preneoplastic lesions in CDAA-fed rats through suppression of hepatic neovascularization markers, namely cluster of differentiation 31 (CD31) and vascular endothelial growth factor (VEGF) [110]. Overall, the key animal studies regarding the impact of SGLT-2i on NAFLD/NASH/HCC are summarized in Table 1, Table 2, Table 3 and Table 4.

Table 1.
Key animal studies regarding the impact of empagliflozin on NAFLD/NASH.
Table 1.
Key animal studies regarding the impact of empagliflozin on NAFLD/NASH.
| Study/Reference | Animal Model, Dose, & Duration | Effect on Body Weight and Liver Weight | Effect on Laboratory Values | Mechanism of Action | Effect on Insulin Sensitivity & Glucose Homeostasis | NAFLD Activity Score (NAS) & Fibrosis/Steatosis |
|---|---|---|---|---|---|---|
| Perakakis, N., et al., 2021 [99] | Male C57BL/6JRj on AMLN diet (HFD, fructose + cholesterol) 10 mg/kg/day 12 (w) | No effect | - | ⬇ Hepatic lactosylceramides | ⬇ Blood glucose levels No effect on Insulin sensitivity | ⬇ Lobular inflammation ⬇ NAS No effect on the hepatic steatosis and fibrosis |
| Meng, Z., et al., 2021 [117] | Male C57BL/6J on HFD + streptozotocin injection (T2DM with NAFLD) 10 mg/kg/day 8 (w) | ⬇ Body weight ⬇ Liver/Bw | ⬇ ALT ⬇ TG & TC ⬆ HDL | ⬇ Lipogenesis markers and lipid uptake genes (SREBP1, ChREBP, FASN, ACCα, SCDα, CD36) ⬆ Autophagy activation (AMPK/mTOR & BECLIN1, LC3BII) ⬆ IL-17/IL-23 axis inhibition (IL-23p19, IL-23, IL-1β, IL-17A, RORγt, p-STAT3/t-STAT3, IL-17) ⬇ M1 macrophage marker (CD11C, CD86, NOS2) ⬇ Th17-related chemokines and chemokine receptors (CCL20, CCR6, CCR4, CXCL1/CXCL2, CXCL1, CXCR2) | ⬇ Blood glucose levels | ⬇ Hepatic steatosis ⬇ NAS |
| Nasiri-Ansari, N., et al., 2021 [30] | Male ApoE knockout mice on HFD 10 mg/kg/day 10 (w) | No effects | ⬇ ALT, TG, TC levels ⬇ Serum TG/HDL levels | ⬇ Lipogenesis markers (SREBP1, Pck1, FASN) ⬇ Inflammatory markers (MCP-1, F4/80) ⬇ ER stress markers (GRP78, IRE1α, XBP1, ELF2α, ATF4, CHOP, GRP94) Autophagy markers (⬇ mTOR and P62 & ⬆ pAMPK/AMPK, BECLIN) ⬇ Apoptosis markers (Bax/Bcl-2 Ratio, cleaved Caspase-3) | ⬇ Blood glucose levels | ⬇ Lobular inflammation ⬇ Hepatic steatosis ⬇ NAS No effect on hepatic fibrosis |
| Petito-da-Silva, et al., 2019 [107] | Male C57Bl/6 mice on HFD 10 mg/kg/day 5 (w) | ⬇ BW ⬇ Liver/Bw | No effect on ALT | ⬇ Lipogenic genes (Fas, SREBP1c, PPARγ) ⬇ ΕR- stress markers (CHOP, ATF4, GADD45) Fatty acid β-oxidation (⬆ PPAR-α, ⬇ Acox1) ⬇ lipid droplet-associated protein (Fsp27/cidec) ⬇ Inflammatory markers (Nfκb, TNF-α) | Improved Glucose intelorence Improved Insulin sensitivity | ⬇ Hepatic TC ⬇ Hepatic steatosis |
| Jojima, T., et al., 2016 [136] | Male C57BL/6J on HFD + early STZ injection 10 mg/kg/day 3 (w) | ⬇ Liver/BW | ⬇ GA ⬇ ALT | ⬇ Inflammatory markers (IL6, TNF-α, MCP-1, SOCS3) ⬇ Plasma DPP-4 activity (CD26/DPP-4) | ⬇ Plasma glucose Levels | ⬇ Hepatic TG ⬇ NAS ⬇ Hepatic fibrosis |
| Hüttl, M., et al., 2021 [115] | HHTg & Wistar rats 10 mg/kg/day 8 (w) | No effect on BW | ⬇ TAG No effect on serum ALT | ⬇ Lipogenicgenes (Fas, Scd-1, SREBP1c, PPARγ) Improvement of hepatic lipid metabolism (⬆ Nrf, Cyp2e1, ⬇ FGF21, Cyp4a1, Cyp1a1, Cyp2b1) ⬇ Inflammatory markers (MCP-1) ⬆ Oxidative stress markers (⬇ Hepatic GSH/GSSG, SOD) ⬇ Hepatokines (Fetuin-A) | Improved Glucose intolerance Improved Insulin sensitivity | ⬇ Hepatic TG ⬇ lipotoxic diacylglycerols ⬇ Fibrosis |
Table 2.
Key animal studies regarding the impact of canagliflozin on NAFLD/NASH/HCC.
Table 2.
Key animal studies regarding the impact of canagliflozin on NAFLD/NASH/HCC.
| Study/Reference | Animal Model Dose & Duration | Effect on Body & Liver Weight | Effect on Laboratory Values | Mechanism of Action | Effect on Insulin Sensitivity & Glucose Homeostasis | NAFLD Activity Score (NAS) & Fibrosis/Steatosis |
|---|---|---|---|---|---|---|
| Yoshino, K., et al., 2021 [111] | obese diabetic KK-Ay mice putative dose of ~17 mg/kg/day 3 (w) | No effect on body weight No effect epididymal fat weigh ⬇ Liver/Bw | ⬇ TG No effect on serum ALT | ⬆ Prostaglandin E2 (PGE2) and resolvin E3 | ⬇ Plasma glucose levels | ⬇ Hepatic TG |
| Tanaka K., et al., 2020 [108] | Male C57BL/6J mice on HFD 30 mg/kg/day 4 (w) | ⬇ BW | ⬇ ALT ⬆ TG, ketone bodies | ⬆ lipid-dependent energy expenditure ⬇ Respiratory Qquotients ⬇ Lipogenesis markers (PPAR, FAS, Scd1) ⬆ Fatty acid β-oxidation markers (CPT1a, PGC1a, PGC1b) ⬇ Inflammatory markers (IL-1b) | Improved glucose intolerance Improved insulin sensitivity ⬇ lasma glucose & insulin levels | ⬇ Hepatic TG ⬇ NAS |
| Jojima, T., et al., 2019 [87] | STAM mice 30 mg/kg/day 4 & 11 (w) | ⬇ Liver/Bw (11 w) | ⬇ TG (11 w) ⬇ ALT (11 w) | ⬇ Inflammatory marker and fibrosis marker [SOCS-3, collagen 3 (4 w)] ⬇ Lipogenesis markers [FAS (11 w)] ⬇ Inhibits progression of NASH to Hepatocarcinogenesis GS & AFP | ⬇ Plasma glucose levels | ⬇ NAS (11 w) ⬇ Hepatic fibrosis (4 w) ⬇ Tumor number (11 w) |
| Shiba, K., et al., 2018 [101] | MC4R-KO mice on HFD 20–30 mg/kg/day 8, 20 & 52 (w) | ⬇ Liver weight (8 w) ⬆ Body weight (8 and 52 w) | ⬆ TG (8 and 20 w) ⬇ ALT (8 and 20 w) | ⬇ Lipogenic markers genes [Acc1 and Scd1 (8 and 20 w), Fasn (8 w)] ⬇ Gluconeogenic markers (G6pc, Pck1) ⬇ Inflammatory markers [F4/80 gene (20 w), TNFa (20 w), Cd11c (8 and 20 w)] ⬇ Fibrosis markers [Col1a1, TIMP-1 (8 and 20w), Acta2, Tgf1b (20 w)] | Improved insulin sensitivity and hyperglycemia (8 and 20 w) | ⬇ Hepatic steatosis (8 w) ⬇ Hepatic fibrosis (20 w) ⬇ NAS (20 w) ⬇ Tumor number (52 w) ⬇ Hepatic TG content (8 and 20 w) |
| Ozutsumi, T., et al., 2020 [110] | F344 rats on CDAA diet 10 mg/kg/day 16 (w) | No effect on body weight No effect on Liver/BW | ⬇ ALT | ⬇ Fibrosis markers (αSMA, TGF-β1, α1(I)-procollagen) ⬇ Inflammatory markers (CCL2, TNF-α, IL-6) ⬇ Hepatocarcinogenesis markers (GST-P, VEGF, CD31) ⬇ Oxidative stress markers (MDA, 8-OHdG) | No effect on insulin sensitivity No effect on plasma glucose levels | ⬇ Hepatic fibrosis & Steatosis ⬇ Hepatic Cirrhosis ⬇ Hepatic inflammation ⬇ Hepatic ballooning ⬇ NAS |
| Kabil, Sl, et al., 2018 [129] | Male Wister rats injected with STZ on HFD 10 and 20 mg/kg/day 8 (w) | ⬇ Liver weight ⬇ BW (20 mg) | ⬇ ALT ⬆ TC, TG & NEFA | ⬆ Hepatic lipolytic factor ZAG ⬇ Inflammatory markers (serum TNF-α, hepatic IL-6) Serum apoptotic markers (⬇ Caspase3, ⬆ Bcl-2) Hepatic oxidative stress (⬇ MDA, ⬆ SOD and GPx activity) Serum antioxidant enzyme activity (⬇ TOS and ⬆ TAS) | No effect on fasting insulin levels Fasting blood glucose | ⬇ Hepatic inflammation ⬇ Hepatic TC, TG, NEFA ⬇ Hepatic inflammation Hepatic Steatosis ⬇ NAS |
Table 3.
Key animal studies regarding the impact of dapagliflozin on NAFLD/NASH.
Table 3.
Key animal studies regarding the impact of dapagliflozin on NAFLD/NASH.
| Study/Reference | Animal Model Dose & Duration | Effect on Body and Liver Weight | Effect on Laboratory Values | Mechanism of Action | Effect on Insulin Sensitivity & Glucose Homeostasis | NAFLD Activity Score (NAS) & Fibrosis/Steatosis |
|---|---|---|---|---|---|---|
| Han, T., et al., 2021 [105] | Male C57BL/6 J and ob/ob mice on HFD 1 mg/kg/day 4 (w) | No effect on BW | ⬇ TC | ⬆ β-oxidation (PPAR-α, CPT1, PGC1α) ⬇ Inflammatory markers (MCP1) | ⬇ Fasting blood glucose | ⬇ Hepatic oxidative stress ⬇ Hepatic lipid accumulation ⬇ Hepatic steatosis |
| Luo, J., et al., 2021 [84] | Male NIH mice on HFD 25 mg/kg/day 4 (w) | No effect on BW ⬆ Food intake | ⬇ ALT | ⬇ Lipogenic markers (SREBP1, ACC, FASN) ⬆ β-oxidation markers (PPARα, CPT1a) Regulation of lipid metabolism ⬆ pAMPK and ⬇ pmTOR | - | ⬇ Hepatic steatosis ⬇ Hepatic ballooning ⬇ HepaticTC, TG |
| Tang, L., et al., 2017 [132] | db/db mice 1.0 mg/kg/day via diet gel 4 (w) | No effect on BW | ⬇ ALT ⬇ TG | ⬇ Inflammatory markers (MPO, F4/80) ⬇ Oxidative stress markers (ROS) ⬇ Fibrosis markers (FN, Col I, Col III, LM) | ⬇ Plasma glucose levels | ⬇ Hepatic injury ⬇ Hepatic fibrosis ⬇ Hepatic inflammation |
| Yabiku, K., et al., 2020 [109] | Male C57BL/6J mice on HFD or HFD + MCDD 0.1 or 1.0 mg/kg/day 2 (w) | ⬇ BW (0.1 and 1 mg) in both diets | ⬇ ALT (0.1 and 1 mg) Mice on HFD | - | Improved glucose tolerance and insulin sensitivity | - |
| Omori, K., et al., 2019 [125] | db/db mice on ND 1.0 mg/kg/day 8 (w) | No effect on BW ⬇ Liver weight | ⬇ TG ⬇ Plasma C-peptide | No significant differences in the expression of fatty acid oxidation markers No significant differences in the expression of inflammatory markers ⬇ Fatty acid uptake and storage markers (PPARγ targeted genes as compared to Gla group) | Improved glucose tolerance | No significant changes in hepatic TG, Palmitate, Oleate, and Stearate content |
| Li, L., et al., 2021 [85] | ZDF rats 1 mg/kg/day 9 (w) | ⬇ BW ⬇ Liver weight ⬇ Liver weight/BW | ⬇ TG, TC, LDL, HDL | ⬇ Lipogenic markers (SREBP1, ACC1, p-ACC) ⬆ Fatty acid oxidation markers (ACOX1, CPT1, pACOX) Autophagy-related markers (⬆ LC3B, Beclin1, activation of AMPK/mTOR pathway and ⬇ P62) | ⬇ Plasma glucose and insulin levels | ⬇ Hepatic lipid accumulation ⬇ Hepatic steatosis |
| ElMahdy, M.K., et al., 2020 [130] | Male Wistar rats on HCHF diet 1 mg/kg/day 5 (w) | No significant effects on liver weight | ⬇ ALT, AST ⬇ TC, TG, LDL ⬆ HDL | ⬇ Inflammatory markers (TNF-α, IL-1β, IL-18) | - | ⬇ Hepatic steatosis |
Table 4.
Key animal studies regarding the impact of ipragliflozin, remogliflozin, tofogliflozin and luseogliflozin on NAFLD/NASH/HCC.
Table 4.
Key animal studies regarding the impact of ipragliflozin, remogliflozin, tofogliflozin and luseogliflozin on NAFLD/NASH/HCC.
| Study/Reference | Animal Model Dose & Duration | Effect on Body Weight &Body Composition | Effect on Laboratory Values | Mechanism of Action | Effect on Insulin Sensitivity & Glucose Homeostasis | NAFLD Activity Score (NAS) & Fibrosis/Steatosis |
|---|---|---|---|---|---|---|
| Ipragliflozin | ||||||
| Tahara, A. & Takasu, T., 2020 [100] | KK-Ay mice on HFD 0.1, 0.3, 1 and 3 mg/kg/day Alone or with Metformin 4 (w) | ⬇ BW weight (1 & 3 mg) ⬇ Liver weight (0.3, 1 & 3 mg) | ⬇ TG (0.3, 1 & 3 mg) ⬇ TC (1 & 3 mg) ⬇ AST (1 & 3 mg) ⬇ ALT (0.3, 1 & 3 mg) | ⬇ Inflammatory Markers [serum TNF-α, IL-6, MCP-1 and CRP (1 and 3mg); Liver TNF-α (3 mg) and IL-6, MCP-1 and CRP (1 & 3 mg)] ⬇ Serum and hepatic oxidative stress markers [TBARS and protein carbonyl (1 & 3 mg)] | Improve glucose intolerance Improved Insulin resistance Improved hyperlipidemia | ⬇ Hepatic TG, TC (1 & 3 mg) ⬇ Hepatic Hyperthrophy (1 & 3 mg) ⬇ Hepatic Inflammation (1 & 3 mg) ⬇ Hepatic fibrosis & steatosis (3 mg) |
| Tahara, A., et al., 2019 [113] | KK-Ay mice on HFD 0.1, 0.3, 1 and 3 mg/kg/day Alone or with Pioglitazone 4 (w) | ⬇ BW weight (1 & 3 mg) ⬇ Liver weight (0.3, 1 & 3 mg) | ⬇ TC (0.3, 1 & 3 mg) ⬇ TG (1 & 3 mg) ⬇ AST (1 & 3 mg) ⬇ ALT (1 & 3 mg) | ⬇ Genes involved in regulation of insulin sensitivity (Plasma adipocytokines, Leptin & FGF-21) ⬇ Inflammatory Markers [serum TNF-α, IL-6, MCP-1 and CRP (1 and 3 mg); Liver TNF-α (3 mg) and IL-6, MCP-1 and CRP (1 and 3 mg)] ⬇ Serum and hepatic oxidative stress markers [TBARS and protein carbonyl (1 & 3 mg)] | ⬇ Plasma glucose and insulin levels (0.3, 1 and 3 mg) | ⬇ Hepatic TG (0.3, 1 & 3 mg) ⬇ Hepatic TC (1 & 3 mg) ⬇ Hepatic Hyperthrophy (1 & 3 mg) ⬇ Hepatic Inflammation (3 mg) ⬇ Hepatic fibrosis (3 mg) |
| Komiya, Ch, et al., 2016 [112] | ob/ob and WT mice on HFD 11 mg/kg/day 4 (w) | ⬇ Hyperphagia ⬇ BW weight ⬇ Liver weight | ⬇ ALT ⬇ TG ⬇ Plasma glucagon | ⬇ Lipogenic markers genes (SREBP1, Fasn, Acc1, Scd1) ⬇ Gluconeogenic markers (Pck1) ⬇ Inflammatory markers (F4/80, Cd11c) | Improved Insulin resistance Improved fasting glucose levels | ⬇ Hepatic TG ⬇ Hepatic lipid ⬇ Hepatic steatosis |
| Honda, Y., et al., 2016 [114] | C57BL/6J male mice on AMLN diet 40 mg/kg/day 8 (w) | No effect on BW ⬇ Liver weight | ⬇ ALT, AST ⬇ FFA | ⬆ β-oxidation (PPAR-α, CPT1, MTTP) ⬇ Hepatocytes apoptosis (TUNEL) ⬇ Lipogenic markers genes (SREBP1 & Acc1) | Improved Insulin resistance | ⬇ Hepatic TG & FFA ⬇ Hepatic fibrosis ⬇ Hepatocyte ballooning ⬇ Lobular inflammation ⬇ NAS |
| Hayashizaki-Someya, Y., et al., 2015 [144] | Male Wistar rats on CDAA diet 0.3 and 3 mg/kg/day 5 (w) | ⬇ BW weight (3 mg) | No effect on ALT, AST | - | No effect on fasting blood glucose levels | ⬇ Hepatic TG (3 mg) ⬇ Hepatic lipid droplet size ⬇ Hepatic fibrosis (0.3 & 3 mg) ⬇ Hepatic HP |
| Yoshioka, N., et al., 2021 [126] | Mc4r KO mice on HFD and injected with single dose of diethylnitrosamine 5 mg/kg/day 12 (w) | ⬇ BW weight ⬇ Liver weight | ⬇ ALT, AST ⬇ LDH | ⬇ Lipogenic markers genes (Fasn in non-tumor) ⬇ Fibrosis markers (Emr1, Itgax in non-tumor) ⬇ Cell senescence markers (Cxcl1 in tumor lesion; p21, Cxcl1, MMp12, mmp13 in non-tumor) ⬆ β-oxidation (PPAR-α in tumor lesion; PPAR-α, CPT1, PGC1 in non-tumor) ⬇ Cell apoptosis (Bax and Pcna) | ⬇ Plasma glucose & insulin levels | ⬇ Hepatic TG ⬇ Lobular inflammation ⬇ Hepatocyte ballooning ⬇ NAS ⬇ Hepatic steatosis & fibrosis ⬇ Hepatic tumor number & size |
| (Remogliflozin) | ||||||
| Nakano, S., et al., 2015 [134] | C57BL/6J mice on HFD32 13.2 ± 2.2 and 33.9 ± 2.0 mg/kg/day 4 (w) | ⬇ Liver weight ⬇ Liver/BW | ⬇ ALT & AST | ⬇ Inflammatory markers [Hepatic TNF-α (13.2 mg), hepatic MCP-1 (13.2 and 33.9 mg)] ⬇ Oxidative stress (serum and hepatic TBARS) | Improved non fasting glucose levels | ⬇ Hepatic TG ⬇ Hepatic fibrosis |
| Tofogliflozin | ||||||
| Obara, K., et al., 2017 [89] | db/db mice on HFD and injected with single dose of diethylnitrosamine 1 and 10 mg/kg/day 14 (w) | ⬇ Liver weight (10 mg) | ⬇ ALT (10 mg) ⬇ FFA (1 & 10 mg) | ⬇ Inflammatory markers (10 mg) (F4/80) | ⬇ Plasma glucose levels Improved insulin insensitivity | ⬇ Foci of cellular alteration (10 mg) ⬇ Hepatic pre-neoplastic lesions (10 mg) ⬇ Hepatocyte balooning (10 mg) ⬇ Hepatic steatosis (10 mg) ⬇ NAS (1 & 10 mg) |
| Luseogliflozin | ||||||
| Qiang, Sh, et al., 2015 [142] | C57BL/6 mice injected with STZ on HFDT Mixing in food at 0.1%. w/w food 8 (w) | ⬇ Liver weight | ⬇ ALT ⬆ TG, NEFA | ⬇ Hepatic fibrosis markers (collagen1a1, collagen1a2, TGF, SMA, TIMP1) ⬇ Inflammatory markers (MCP-1, IL1, IL-12, IL-6, f4/80) | ⬇ Plasma glucose levels | ⬇ Hepatic TC, TG &NEFA |
Abbreviations: W: week, BW: Body Weight, STZ: streptozotocin, WT: Wild Type, Gla:insulin glargine, ALT: Alanine aminotransferase, AST: Aspartate aminotransferase, TG: Triglycerides, TC: Total cholesterol, FFA: Free fatty acids, GA: glycated albumin, STAT3: Signal Transducer And Activator Of Transcription 3, ROR: RAR Related Orphan Receptor, Bax: BCL2 Associated X, IRE: Inositol-requiring enzyme 1, -Xbp1: X-box binding protein 1, ATF4: Activating transcription factor 4, CHOP: C/EBP homologous protein, ATF-6:Activating transcription factor 6, ChREBP: Carbohydrate response element binding protein, SREBP1: Sterol regulatory element-binding transcription factor 1, Scd1: Stearoyl-CoA desaturase1, ACC1: Acetyl-CoAcarboxylase 1, ACOX: Peroxisomal acyl-CoA oxidase1 Fasn: Fatty acid synthase, IL: Interleukin, TNF-α: Tumor necrosis factor, NFκΒ: Nuclear factor kappa-light-chain-enhancer of activated B cells, MCP-1:Monocyte chemoattractant protein-1, CRP:C-reactive protein, PPAR-α: Peroxisome proliferator activated receptor alpha, CPT1: Carnitine palmitoyltransferase 1, PGC1: PPARγ coactivator 1, TOS: Total Oxidant Status, TAS: Total antioxidant status, p21: cyclin-dependent kinase inhibitor, Cxcl: Chemokine (C-X-C motif) ligand, MMP: Matrix metalloproteinases, TIMP: Tissue inhibitors of matrix metalloproteinases, CYP: Cytochromes P450, MDA: Malondialdehyde.
4.2. Human Trials
A number of clinical studies have highlighted the benefit of SGLT-2i in patients with T2DM and NAFLD (Table 5) [145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169]. In the majority of them, the administration of SGLT-2i has resulted in improvement of serum levels of liver enzymes and hepatic steatosis, as evaluated by magnetic resonance imaging (MRI), ultrasound (U/S), non-invasive biomarkers, such as AST to platelet ratio (APRI) index, NAFLD fibrosis score (NFS) and Fibrosis-4 (FIB-4) score, or even by liver biopsy (LB). In some of these studies, improvement in hepatic fibrosis was found, using transient elastography (TE) or LB, even though this finding was not univocal [146,153,158,159,164,165,168].
Of note, the overall improvement in hepatic steatosis is also found in patients treated with other classes of anti-diabetic drugs, like thiazolidinediones and dipeptidyl peptidase-4 (DPP-4) inhibitors, raising the question of steatosis improvement due to glycemic control [148,160,162,170]. Moreover, the vast majority of the aforementioned studies are limited by the small sample size, heterogenous inclusion criteria, especially regarding the presence of NAFLD, as well as the duration of follow up, thus meta-analyses have been conducted to better assess the true benefit of SGLT-2i in patients with DM and NAFLD [171,172,173]. In the largest one, comprising 9 randomized trials, with 7281 and 4088 patients in the SGLT-2i and control arm, (standard of care (SOC) or placebo), respectively, the use of SGLT-2i resulted in improvement of serum transaminases, body weight, and liver fat as measured by proton density fat fraction. Accordingly, the authors discuss that this improvement derives mainly from the achievement of glycemic control and weight loss. However, in a 2020 study by Kahl et al., including 84 patients with DM and excellent glycemic control, randomly assigned to empagliflozin or placebo, patients on empagliflozin had improved liver fat content, as assessed by MRI [161], strongly suggesting that the good glycemic control and weight loss are not the only mechanisms associated with the beneficial effects of SGLT-2i treatment on hepatic steatosis.
In non-DM patients, only a small single center study exists which studied 12 patients under dapagliflozin and 10 patients under teneligliptin, a DPP4 inhibitor, for a total of 12 weeks, showing that after this intervention period, serum transaminases were decreased in both groups, while in the dapagliflozin group, total body water and body fat decreased, leading to decreased total body weight [174].
Regarding the pathophysiology of NAFLD improvement under SGLT-2i treatment, various mechanisms have been suggested. Treatment with SGLT-2i results in decreases in both glucose and insulin levels (especially in patients with DM), which lead to a large reduction of hepatic de novo lipid synthesis [175]. Moreover, glucagon-secreting alpha pancreatic cells also express SGLT-2, thus the administration of SGLT-2i stimulates glucagon secretion [175,176,177]. In turn, the subsequently elevated plasma glucagon levels stimulate β-oxidation, and this shift from carbohydrate to fatty acid metabolism leads to reduced liver triglyceride content and consequently hepatic steatosis [175,178,179]. Another potential mechanism is mediated by the antioxidant effects of SGLT-2i. Apart from their ability to reduce high glucose-induced oxidative stress, SGLT-2i reduce free radical generation, suppress pro-oxidants, and upregulate antioxidant systems, such as superoxide dismutases (SODs) and glutathione (GSH) peroxidases (Figure 1) [116,180,181,182,183,184].
Table 5.
Studies on the impact of SGLT-2i on NAFLD in patients with T2DM.
Table 5.
Studies on the impact of SGLT-2i on NAFLD in patients with T2DM.
| Study | Study Design | No of Pts | SGLT-2i/Drug Used (No of Pts) | Control Group | Treatment Duration (Weeks) | NAFLD Diagnosis ** | Key Results |
|---|---|---|---|---|---|---|---|
| Eriksson, J., et al., 2018 [150] | Randomised, double-blind, prospective | 84 | Dapagliflozin (42) | OM-3CA or placebo | 12 | MRI | Reduction of serum transaminases, CK-18, FGF-21 in Dapagliflozin group and liver fat in Dapagliflozin + OM-3CA group |
| Kahl, S., et al., 2020, [161] | Randomised, double-blind, prospective | 84 * | Empagliflozin (42) | Placebo | 24 | MRI | LFC improvement only in empagliflozin |
| Chehrehgosha, H., et al., 2021 [165] | Randomised, double-blind, prospective | 78 | Empagliflozin (21) | Pioglitzone or placebo | 24 | TE | Better CAP, LS, no difference vs. pioglitzone for serum transaminases or FIB-4 |
| Gaborit, B., et al., 2021 [167] | Randomised, double-blind, prospective | 34 | Empagliflozin (18) | Placebo | 12 | MRI | Reduction in liver fat vs. placebo |
| Bando, Y., et al., 2017 [145] | Randomised, open label, prospective | 62 | Ipragliflozin (40) | SOC | 12 | C/T | Improvement in serum transaminases. VFA, L/S ratio compared to SOC |
| Ito, D., et al., 2017 [147] | Randomized, open label, prospective | 66 | Ipragliflozin (32) | Pioglitazone | 24 | C/T or U/S | Improvement of L/S ratio, ALT, ferritin not statistically significant between 2 groups; ipragliflozin more weight and VFA reduction |
| Kuchay, M.S., et al., 2018 [152] | Randomized, open label, prospective | 42 | Empagliflozin (22) | SOC | 20 | MRI | Reduction of liver fat and ALT |
| Shibuya, T., et al., 2018 [154] | Randomized, open label, prospective | 32 | Luseogliflozin (16) | Metformin | 26 (6 months) | C/T or U/S | Improvement in L/S ratio compared to baseline |
| Shimizu, M., et al., 2019 [155] | Randomized, open label, prospective | 57 | Dapagliflozin (33) | SOC | 24 | U/S | Improvement of CAP and LS, especially for high LS at the trial beginning |
| Han, E., et al., 2020 [160] | Randomized, open label, prospective | 44 | Ipragliflozin (+metformin +pioglitazone) (29) | Metformin + pioglitazone | 24 | U/S | Better FLI, CAP, NAFLD liver fat score |
| Kinoshita, T., et al., 2020 [162] | Randomized, open label, prospective | 98 | Dapagliflozin (32) | Pioglitazone (33) Glimepiride (33) | 28 | C/T | Improvement of L/S ratio and ALT with pioglitazone and dapagliflozin |
| Takahashi, H., et al., 2021 [168] | Randomized, open label, prospective | 55 | Ipragliflozin (27) | SOC, except pioglitazone, GLP1 | 72 | LB | Statistically significant improvement in NASH resolution and fibrosis improvement in SGLT-2i vs. SOC |
| Yoneda, M., et al., 2021 [169] | Randomized, open label, prospective | 40 | Topogliflozin (21) | Pioglitzone | 24 | MRI | Decrease of liver steatosis in both groups, body weight decrease in topogliflozin |
| Arai, T., et al., 2021 [164] | Open label, Prospective | 100 | Canagliflozin (29) Ipragliflozin (12) Tofogliflozin (6) Dapagliflozin (4) Luseogliflozin (4) Empagliflozin (1) | SOC | 48 | U/S | Decrease in LS and CAP in SGLT-2i during treatment, statistically significant decrease in SGLT-2i vs SOC in ALT, FIB-4 |
| Akuta, N., et al., 2017 [146] | Single-arm, Prospective | 5 | Canagliflozin (5) | N/A | 24 | LB | Improvement of NAS score, liver steatosis; fibrosis improvement in 2 pts |
| Itani, T., et al., 2018 [151] | Single arm, Prospective | 35 | Canagliflozin (35) | N/A | 26 (6 months) | U/S | Improvement in ALT, ferritin, FIB-4 at 3 and 6 months |
| Miyake, T., et al., 2018 [153] | Single arm, Prospective | 43 | Ipragliflozin (43) | N/A | 24 | 12 LB, 41 U/S | Reduction in serum transaminases, CAP, not statistically significant reduction in fibrosis |
| Sumida, Y., et al., 2019 [156] | Single-arm, Prospective | 40 | Luseogliflozin (40) | N/A | 24 | U/S | Reduction in transaminases, serum ferritin and liver fat in MRI |
| Akuta, N., et al., 2019 [157] | Single arm, Prospective | 9 | Canagliflozin (9) | N/A | 24 | LB | Histological improvement in all patients |
| Akuta, N., et al., 2020 [159] | Single arm, Prospective | 7 | Canagliflozin (7) | N/A | 24 | LB | Histopathological improvement at 24 weeks sustained to >1 year, transaminases and ferritin better at 24 weeks |
| Seko, Y., et al., 2017 [148] | Retrospective | 45 | Canagliflozin (18) Ipragliflozin (6) | Sitagliptin | 24 | LB | Significant decrease in serum transaminases with both drugs, not statistically significant between SGLT-2i and sitagliptin |
| Choi, D.H., et al., 2018 [149] | Retrospective | 102 (all abnormal ALT) | Dapagliflozin + Metformin (50) | DPP4 + Metformin | 44.4 ± 18.4 for dapagliflozin and 50.4 ± 21.6 for DPP4 | U/S | Statistically significant decrease in dapagliflozin vs. DPP4 |
| Yamashima, M., et al., 2019 [158] | Retrospective | 22 | Ipragliflozin (18) Dapagliflozin (2) Tofogliflozin (1) Empagliflozin (1) | N/A | 52 (22 pts) and 104 (15 pts) | 12 LB, 10 U/S | Lower serum transaminases levels at 12 and 24 months, better CAR and shear wave velocity at 12 months |
| Yano, K., et al., 2020 [163] | Retrospective | 69 | Dapagliflozin (10) Canagliflozin (7) Ipragliflozin (3) Empagliflozin (2) | SOC | 162 | LB | Improvement of serum transaminases in both groups (No head to head comparison) |
| Euh, W., et al., 2021 [166] | Retrospective | 283 | Dapagliflozin (58) Empagliflozin (34) Ipragliflozin (3) | SOC, except GLP-1 and Insulin | 39 | U/S | Statistically significant reduction in ALT and body weight in SLT2i vs. SOC |
* All patients with excellent glycemic control. ** Test used to diagnose/assess NAFLD.Abbreviations: RCT: Randomised controlled trial, L/S ratio: Liver to spleen ratio, VFA: Visceral fat area, C/T: Computed tomography, MRI: Magnetic Resonance Imaging, OM-3CA: omega-3 carboxylic acids, LFC: Liver fat content, FIB-4: Fibrosis-4 index, ALT: Alanine aminotransferase, FLI: Fatty liver index, CAP: Controlled attenuation parameter, SGLT-2i: Sodium-glucose co-transporter type-2 inhibitors, NAS score: NAFLD Activity Score, LB: Liver biopsy, GLP-1: Glucagon-like peptide-1, pts: patients.
5. Conclusions
There is increasing interest regarding the promising effect(s) of SGLT-2i for the treatment of NAFLD, regardless of the co-existence of T2DM. In addition to weight loss, the beneficial effect(s) of SGLT-2i on NAFLD development and progression appear to be mediated directly through regulation of multiple processes, including ER stress, oxidative stress, low-grade inflammation, autophagy and apoptosis, as revealed by in vitro, animal and clinical studies. Moreover, the observed different effects between members of the SGLT-2i class suggest that there are features specific to individual drugs of this class regarding the underlying mechanism(s) of action and their corresponding effects on NAFLD.
Funding
This review article received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, incidence, and outcome of non-alcoholic fatty liver disease in Asia, 1999–2019: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2019, 4, 389–398. [Google Scholar] [CrossRef]
- Hui, J.M.; Kench, J.G.; Chitturi, S.; Sud, A.; Farrell, G.C.; Byth, K.; Hall, P.; Khan, M.; George, J. Long-term outcomes of cirrhosis in nonalcoholic steatohepatitis compared with hepatitis C. Hepatology 2003, 38, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Bugianesi, E.; Leone, N.; Vanni, E.; Marchesini, G.; Brunello, F.; Carucci, P.; Musso, A.; De Paolis, P.; Capussotti, L.; Salizzoni, M.; et al. Expanding the natural history of nonalcoholic steatohepatitis: From cryptogenic cirrhosis to hepatocellular carcinoma. Gastroenterology 2002, 123, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Yki-Jarvinen, H. Non-alcoholic fatty liver disease as a cause and a consequence of metabolic syndrome. Lancet Diabetes Endocrinol. 2014, 2, 901–910. [Google Scholar] [CrossRef]
- Petroni, M.L.; Brodosi, L.; Bugianesi, E.; Marchesini, G. Management of non-alcoholic fatty liver disease. BMJ 2021, 372, m4747. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Fouad, Y.; Waked, I.; Bollipo, S.; Gomaa, A.; Ajlouni, Y.; Attia, D. What’s in a name? Renaming ‘NAFLD’ to ‘MAFLD’. Liver Int. 2020, 40, 1254–1261. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, L.; Pafundi, P.C.; Galiero, R.; Caturano, A.; Morone, M.V.; Silvestri, C.; Giordano, M.; Salvatore, T.; Sasso, F.C. Mechanisms of Non-Alcoholic Fatty Liver Disease in the Metabolic Syndrome. A Narrative Review. Antioxidants 2021, 10, 270. [Google Scholar] [CrossRef]
- Masarone, M.; Rosato, V.; Aglitti, A.; Bucci, T.; Caruso, R.; Salvatore, T.; Sasso, F.C.; Tripodi, M.F.; Persico, M. Liver biopsy in type 2 diabetes mellitus: Steatohepatitis represents the sole feature of liver damage. PLoS ONE 2017, 12, e0178473. [Google Scholar] [CrossRef]
- Tanase, D.M.; Gosav, E.M.; Costea, C.F.; Ciocoiu, M.; Lacatusu, C.M.; Maranduca, M.A.; Ouatu, A.; Floria, M. The Intricate Relationship between Type 2 Diabetes Mellitus (T2DM), Insulin Resistance (IR), and Nonalcoholic Fatty Liver Disease (NAFLD). J. Diabetes Res. 2020, 2020, 3920196. [Google Scholar] [CrossRef]
- Fargion, S.; Porzio, M.; Fracanzani, A.L. Nonalcoholic fatty liver disease and vascular disease: State-of-the-art. World J. Gastroenterol. 2014, 20, 13306–13324. [Google Scholar] [CrossRef]
- Galiero, R.; Caturano, A.; Vetrano, E.; Cesaro, A.; Rinaldi, L.; Salvatore, T.; Marfella, R.; Sardu, C.; Moscarella, E.; Gragnano, F.; et al. Pathophysiological mechanisms and clinical evidence of relationship between Nonalcoholic fatty liver disease (NAFLD) and cardiovascular disease. Rev. Cardiovasc. Med. 2021, 22, 755–768. [Google Scholar] [CrossRef]
- Petta, S.; Argano, C.; Colomba, D.; Camma, C.; Di Marco, V.; Cabibi, D.; Tuttolomondo, A.; Marchesini, G.; Pinto, A.; Licata, G.; et al. Epicardial fat, cardiac geometry and cardiac function in patients with non-alcoholic fatty liver disease: Association with the severity of liver disease. J. Hepatol. 2015, 62, 928–933. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (FDA). FDA Approves New Treatment for a Type of Heart Failure. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-new-treatment-type-heart-failure (accessed on 10 December 2021).
- U.S. Food and Drug Administration (FDA). FDA Approves Treatment for Chronic Kidney Disease. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-treatment-chronic-kidney-disease (accessed on 15 December 2021).
- Kim, J.W.; Lee, Y.J.; You, Y.H.; Moon, M.K.; Yoon, K.H.; Ahn, Y.B.; Ko, S.H. Effect of sodium-glucose cotransporter 2 inhibitor, empagliflozin, and alpha-glucosidase inhibitor, voglibose, on hepatic steatosis in an animal model of type 2 diabetes. J. Cell Biochem. 2018, 20, 8534–8546. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Eslam, M.; Valenti, L.; Romeo, S. Genetics and epigenetics of NAFLD and NASH: Clinical impact. J. Hepatol. 2018, 68, 268–279. [Google Scholar] [CrossRef]
- Mahady, S.E.; George, J. Exercise and diet in the management of nonalcoholic fatty liver disease. Metabolism 2016, 65, 1172–1182. [Google Scholar] [CrossRef] [Green Version]
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef]
- Marra, F.; Bertolani, C. Adipokines in liver diseases. Hepatology 2009, 50, 957–969. [Google Scholar] [CrossRef]
- Marra, F.; Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. J. Hepatol. 2018, 68, 280–295. [Google Scholar] [CrossRef]
- Gluchowski, N.L.; Becuwe, M.; Walther, T.C.; Farese, R.V., Jr. Lipid droplets and liver disease: From basic biology to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 343–355. [Google Scholar] [CrossRef]
- Mansouri, A.; Gattolliat, C.H.; Asselah, T. Mitochondrial Dysfunction and Signaling in Chronic Liver Diseases. Gastroenterology 2018, 155, 629–647. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, K.; Kong, A.; Zhou, Y.; Chen, D.; Gu, J.; Shi, H. Dysregulation of autophagy acts as a pathogenic mechanism of non-alcoholic fatty liver disease (NAFLD) induced by common environmental pollutants. Ecotoxicol. Environ. Saf. 2021, 217, 112256. [Google Scholar] [CrossRef]
- Nasiri-Ansari, N.; Nikolopoulou, C.; Papoutsi, K.; Kyrou, I.; Mantzoros, C.S.; Kyriakopoulos, G.; Chatzigeorgiou, A.; Kalotychou, V.; Randeva, M.S.; Chatha, K.; et al. Empagliflozin Attenuates Non-Alcoholic Fatty Liver Disease (NAFLD) in High Fat Diet Fed ApoE−/− Mice by Activating Autophagy and Reducing ER Stress and Apoptosis. Int. J. Mol. Sci. 2021, 22, 818. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, X.; Lu, Y.; Wang, E.; Zhang, Z.; Yang, J.; Zhang, H.; Li, X. Hepatic steatosis exacerbated by endoplasmic reticulum stress-mediated downregulation of FXR in aging mice. J. Hepatol. 2014, 60, 847–854. [Google Scholar] [CrossRef]
- Zhang, X.; Han, J.; Man, K.; Li, X.; Du, J.; Chu, E.S.; Go, M.Y.; Sung, J.J.; Yu, J. CXC chemokine receptor 3 promotes steatohepatitis in mice through mediating inflammatory cytokines, macrophages and autophagy. J. Hepatol. 2016, 64, 160–170. [Google Scholar] [CrossRef] [Green Version]
- Alkhouri, N.; Carter-Kent, C.; Feldstein, A.E. Apoptosis in nonalcoholic fatty liver disease: Diagnostic and therapeutic implications. Expert Rev. Gastroenterol. Hepatol. 2011, 5, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Zhang, X.J.; Li, H. The Role of Innate Immune Cells in Nonalcoholic Steatohepatitis. Hepatology 2019, 70, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.A.; Friedman, S.L. Inflammatory and fibrotic mechanisms in NAFLD-Implications for new treatment strategies. J. Intern. Med. 2021, 291, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Ertunc, M.E.; Hotamisligil, G.S. Lipid signaling and lipotoxicity in metaflammation: Indications for metabolic disease pathogenesis and treatment. J. Lipid Res. 2016, 57, 2099–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, D.; Kotronen, A.; Westerbacka, J.; Puig, O.; Arkkila, P.; Kiviluoto, T.; Laitinen, S.; Kolak, M.; Fisher, R.M.; Hamsten, A.; et al. Gene expression in human NAFLD. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1281–G1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magkos, F.; Fraterrigo, G.; Yoshino, J.; Luecking, C.; Kirbach, K.; Kelly, S.C.; de Las Fuentes, L.; He, S.; Okunade, A.L.; Patterson, B.W.; et al. Effects of Moderate and Subsequent Progressive Weight Loss on Metabolic Function and Adipose Tissue Biology in Humans with Obesity. Cell Metab. 2016, 23, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Vilar-Gomez, E.; Martinez-Perez, Y.; Calzadilla-Bertot, L.; Torres-Gonzalez, A.; Gra-Oramas, B.; Gonzalez-Fabian, L.; Friedman, S.L.; Diago, M.; Romero-Gomez, M. Weight Loss through Lifestyle Modification Significantly Reduces Features of Nonalcoholic Steatohepatitis. Gastroenterology 2015, 149, 367–378.e5; quiz e14–e15. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Noworolski, S.M.; Erkin-Cakmak, A.; Korn, N.J.; Wen, M.J.; Tai, V.W.; Jones, G.M.; Palii, S.P.; Velasco-Alin, M.; Pan, K.; et al. Effects of Dietary Fructose Restriction on Liver Fat, De Novo Lipogenesis, and Insulin Kinetics in Children with Obesity. Gastroenterology 2017, 153, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Invest. 2005, 115, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Viscarra, J.; Kim, S.J.; Sul, H.S. Transcriptional regulation of hepatic lipogenesis. Nat. Rev. Mol. Cell Biol. 2015, 16, 678–689. [Google Scholar] [CrossRef] [Green Version]
- Abdelmalek, M.F.; Lazo, M.; Horska, A.; Bonekamp, S.; Lipkin, E.W.; Balasubramanyam, A.; Bantle, J.P.; Johnson, R.J.; Diehl, A.M.; Clark, J.M. Higher dietary fructose is associated with impaired hepatic adenosine triphosphate homeostasis in obese individuals with type 2 diabetes. Hepatology 2012, 56, 952–960. [Google Scholar] [CrossRef] [Green Version]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [Green Version]
- Xu, A.; Wang, Y.; Keshaw, H.; Xu, L.Y.; Lam, K.S.; Cooper, G.J. The fat-derived hormone adiponectin alleviates alcoholic and nonalcoholic fatty liver diseases in mice. J. Clin. Invest. 2003, 112, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Myers, M.G.; Cowley, M.A.; Münzberg, H. Mechanisms of leptin action and leptin resistance. Annu. Rev. Physiol. 2008, 70, 537–556. [Google Scholar] [CrossRef] [Green Version]
- Sabio, G.; Das, M.; Mora, A.; Zhang, Z.; Jun, J.Y.; Ko, H.J.; Barrett, T.; Kim, J.K.; Davis, R.J. A stress signaling pathway in adipose tissue regulates hepatic insulin resistance. Science 2008, 322, 1539–1543. [Google Scholar] [CrossRef] [Green Version]
- Aubert, J.; Begriche, K.; Knockaert, L.; Robin, M.A.; Fromenty, B. Increased expression of cytochrome P450 2E1 in nonalcoholic fatty liver disease: Mechanisms and pathophysiological role. Clin. Res. Hepatol. Gastroenterol. 2011, 35, 630–637. [Google Scholar] [CrossRef]
- Delli Bovi, A.P.; Marciano, F.; Mandato, C.; Siano, M.A.; Savoia, M.; Vajro, P. Oxidative Stress in Non-alcoholic Fatty Liver Disease. An Updated Mini Review. Front. Med. 2021, 8, 595371. [Google Scholar] [CrossRef]
- Cases, S.; Smith, S.J.; Zheng, Y.W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welch, C.B.; Lusis, A.J.; et al. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. USA 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [Green Version]
- Cases, S.; Stone, S.J.; Zhou, P.; Yen, E.; Tow, B.; Lardizabal, K.D.; Voelker, T.; Farese, R.V., Jr. Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase, and related family members. J. Biol. Chem. 2001, 276, 38870–38876. [Google Scholar] [CrossRef] [Green Version]
- Luukkonen, P.K.; Zhou, Y.; Sädevirta, S.; Leivonen, M.; Arola, J.; Orešič, M.; Hyötyläinen, T.; Yki-Järvinen, H. Hepatic ceramides dissociate steatosis and insulin resistance in patients with non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Han, M.S.; Park, S.Y.; Shinzawa, K.; Kim, S.; Chung, K.W.; Lee, J.H.; Kwon, C.H.; Lee, K.W.; Lee, J.H.; Park, C.K.; et al. Lysophosphatidylcholine as a death effector in the lipoapoptosis of hepatocytes. J. Lipid Res. 2008, 49, 84–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassnacht, M.; Kreissl, M.C.; Weismann, D.; Allolio, B. New targets and therapeutic approaches for endocrine malignancies. Pharmacol. Ther. 2009, 123, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Yang, L.; McCall, S.; Huang, J.; Yu, X.X.; Pandey, S.K.; Bhanot, S.; Monia, B.P.; Li, Y.X.; Diehl, A.M. Inhibiting triglyceride synthesis improves hepatic steatosis but exacerbates liver damage and fibrosis in obese mice with nonalcoholic steatohepatitis. Hepatology 2007, 45, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Mirshahi, F.; Cheung, O.; Natarajan, R.; Maher, J.W.; Kellum, J.M.; Sanyal, A.J. Activation and dysregulation of the unfolded protein response in nonalcoholic fatty liver disease. Gastroenterology 2008, 134, 568–576. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Pagliassotti, M.J. Saturated fatty acid-mediated endoplasmic reticulum stress and apoptosis are augmented by trans-10, cis-12-conjugated linoleic acid in liver cells. Mol. Cell Biochem. 2007, 303, 105–113. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Topczewski, F.; Pagliassotti, M.J. Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E275–E281. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.M.; Wang, K.; Adler, A.J.; Vella, A.T.; Zhou, B. Macrophage polarization and meta-inflammation. Transl. Res. 2018, 191, 29–44. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R.; Szabo, G. Interleukin-1 and inflammasomes in alcoholic liver disease/acute alcoholic hepatitis and nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. Hepatology 2016, 64, 955–965. [Google Scholar] [CrossRef]
- Gomes, A.L.; Teijeiro, A.; Burén, S.; Tummala, K.S.; Yilmaz, M.; Waisman, A.; Theurillat, J.P.; Perna, C.; Djouder, N. Metabolic Inflammation-Associated IL-17A Causes Non-alcoholic Steatohepatitis and Hepatocellular Carcinoma. Cancer Cell 2016, 30, 161–175. [Google Scholar] [CrossRef]
- Choi, C.I. Sodium-Glucose Cotransporter 2 (SGLT2) Inhibitors from Natural Products: Discovery of Next-Generation Antihyperglycemic Agents. Molecules 2016, 21, 1136. [Google Scholar] [CrossRef] [Green Version]
- Hardman, T.C.; Rutherford, P.; Duprey, S.W.; Wierzbicki, A.S. Sodium-glucose co-transporter 2 inhibitors: From apple tree to ‘Sweet Pee’. Curr. Pharm. Des. 2010, 16, 3830–3838. [Google Scholar] [CrossRef]
- Blaschek, W. Natural Products as Lead Compounds for Sodium Glucose Cotransporter (SGLT) Inhibitors. Planta Med. 2017, 83, 985–993. [Google Scholar] [CrossRef]
- Vick, H.; Diedrich, D.F.; Baumann, K. Reevaluation of renal tubular glucose transport inhibition by phlorizin analogs. Am. J. Physiol. 1973, 224, 552–557. [Google Scholar] [CrossRef]
- Ehrenkranz, J.R.; Lewis, N.G.; Kahn, C.R.; Roth, J. Phlorizin: A review. Diabetes Metab. Res. Rev. 2005, 21, 31–38. [Google Scholar] [CrossRef]
- Tian, L.; Cao, J.; Zhao, T.; Liu, Y.; Khan, A.; Cheng, G. The Bioavailability, Extraction, Biosynthesis and Distribution of Natural Dihydrochalcone: Phloridzin. Int. J. Mol. Sci. 2021, 22, 962. [Google Scholar] [CrossRef]
- Lehmann, A.; Hornby, P.J. Intestinal SGLT1 in metabolic health and disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G887–G898. [Google Scholar] [CrossRef] [Green Version]
- Katsuno, K.; Fujimori, Y.; Takemura, Y.; Hiratochi, M.; Itoh, F.; Komatsu, Y.; Fujikura, H.; Isaji, M. Sergliflozin, a novel selective inhibitor of low-affinity sodium glucose cotransporter (SGLT2), validates the critical role of SGLT2 in renal glucose reabsorption and modulates plasma glucose level. J. Pharmacol. Exp. Ther. 2007, 320, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Oku, A.; Ueta, K.; Arakawa, K.; Ishihara, T.; Nawano, M.; Kuronuma, Y.; Matsumoto, M.; Saito, A.; Tsujihara, K.; Anai, M.; et al. T-1095, an inhibitor of renal Na+-glucose cotransporters, may provide a novel approach to treating diabetes. Diabetes 1999, 48, 1794–1800. [Google Scholar] [CrossRef]
- Meng, W.; Ellsworth, B.A.; Nirschl, A.A.; McCann, P.J.; Patel, M.; Girotra, R.N.; Wu, G.; Sher, P.M.; Morrison, E.P.; Biller, S.A.; et al. Discovery of dapagliflozin: A potent, selective renal sodium-dependent glucose cotransporter 2 (SGLT2) inhibitor for the treatment of type 2 diabetes. J. Med. Chem. 2008, 51, 1145–1149. [Google Scholar] [CrossRef]
- Nomura, S.; Sakamaki, S.; Hongu, M.; Kawanishi, E.; Koga, Y.; Sakamoto, T.; Yamamoto, Y.; Ueta, K.; Kimata, H.; Nakayama, K.; et al. Discovery of canagliflozin, a novel C-glucoside with thiophene ring, as sodium-dependent glucose cotransporter 2 inhibitor for the treatment of type 2 diabetes mellitus. J. Med. Chem. 2010, 53, 6355–6360. [Google Scholar] [CrossRef]
- Hsia, D.S.; Grove, O.; Cefalu, W.T. An update on sodium-glucose co-transporter-2 inhibitors for the treatment of diabetes mellitus. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 73–79. [Google Scholar] [CrossRef]
- Markham, A.; Elkinson, S. Luseogliflozin: First global approval. Drugs 2014, 74, 945–950. [Google Scholar] [CrossRef]
- Poole, R.M.; Dungo, R.T. Ipragliflozin: First global approval. Drugs 2014, 74, 611–617. [Google Scholar] [CrossRef]
- Poole, R.M.; Prossler, J.E. Tofogliflozin: First global approval. Drugs 2014, 74, 939–944. [Google Scholar] [CrossRef]
- Xu, J.; Hirai, T.; Koya, D.; Kitada, M. Effects of SGLT2 Inhibitors on Atherosclerosis: Lessons from Cardiovascular Clinical Outcomes in Type 2 Diabetic Patients and Basic Researches. J. Clin. Med. 2021, 11, 137. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Isaji, M. Sodium-glucose cotransporter inhibitors for diabetes. Curr. Opin. Investig. Drugs 2007, 8, 285–292. [Google Scholar]
- Isaji, M. SGLT2 inhibitors: Molecular design and potential differences in effect. Kidney Int. Suppl. 2011, 79 (Suppl. S20), S14–S19. [Google Scholar] [CrossRef] [Green Version]
- Okada, J.; Yamada, E.; Saito, T.; Yokoo, H.; Osaki, A.; Shimoda, Y.; Ozawa, A.; Nakajima, Y.; Pessin, J.E.; Okada, S.; et al. Dapagliflozin Inhibits Cell Adhesion to Collagen I and IV and Increases Ectodomain Proteolytic Cleavage of DDR1 by Increasing ADAM10 Activity. Molecules 2020, 25, 495. [Google Scholar] [CrossRef] [Green Version]
- Kaji, K.; Nishimura, N.; Seki, K.; Sato, S.; Saikawa, S.; Nakanishi, K.; Furukawa, M.; Kawaratani, H.; Kitade, M.; Moriya, K.; et al. Sodium glucose cotransporter 2 inhibitor canagliflozin attenuates liver cancer cell growth and angiogenic activity by inhibiting glucose uptake. Int. J. Cancer 2018, 142, 1712–1722. [Google Scholar] [CrossRef] [Green Version]
- Hawley, S.A.; Ford, R.J.; Smith, B.K.; Gowans, G.J.; Mancini, S.J.; Pitt, R.D.; Day, E.A.; Salt, I.P.; Steinberg, G.R.; Hardie, D.G. The Na+/Glucose Cotransporter Inhibitor Canagliflozin Activates AMPK by Inhibiting Mitochondrial Function and Increasing Cellular AMP Levels. Diabetes 2016, 65, 2784–2794. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Sun, P.; Wang, Y.; Chen, Y.; Niu, Y.; Ding, Y.; Xu, N.; Zhang, Y.; Xie, W. Dapagliflozin attenuates steatosis in livers of high-fat diet-induced mice and oleic acid-treated L02 cells via regulating AMPK/mTOR pathway. Eur. J. Pharmacol. 2021, 907, 174304. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Q.; Huang, W.; Han, Y.; Tan, H.; An, M.; Xiang, Q.; Zhou, R.; Yang, L.; Cheng, Y. Dapagliflozin Alleviates Hepatic Steatosis by Restoring Autophagy via the AMPK-mTOR Pathway. Front. Pharmacol. 2021, 12, 589273. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, M.; Yin, F.; Wang, Y.; Li, X.; Wu, Y.; Ye, C.; Liu, J. Trilobatin, a Novel SGLT1/2 Inhibitor, Selectively Induces the Proliferation of Human Hepatoblastoma Cells. Molecules 2019, 24, 3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jojima, T.; Wakamatsu, S.; Kase, M.; Iijima, T.; Maejima, Y.; Shimomura, K.; Kogai, T.; Tomaru, T.; Usui, I.; Aso, Y. The SGLT2 Inhibitor Canagliflozin Prevents Carcinogenesis in a Mouse Model of Diabetes and Non-Alcoholic Steatohepatitis-Related Hepatocarcinogenesis: Association with SGLT2 Expression in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2019, 20, 5237. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rafei, M.K.; Thabet, N.M.; Rashed, L.A.; Moustafa, E.M. Canagliflozin, a SGLT-2 inhibitor, relieves ER stress, modulates autophagy and induces apoptosis in irradiated HepG2 cells: Signal transduction between PI3K/AKT/GSK-3beta/mTOR and Wnt/beta-catenin pathways; in vitro. J. Cancer Res. Ther. 2021, 17, 1404–1418. [Google Scholar] [CrossRef] [PubMed]
- Obara, K.; Shirakami, Y.; Maruta, A.; Ideta, T.; Miyazaki, T.; Kochi, T.; Sakai, H.; Tanaka, T.; Seishima, M.; Shimizu, M. Preventive effects of the sodium glucose cotransporter 2 inhibitor tofogliflozin on diethylnitrosamine-induced liver tumorigenesis in obese and diabetic mice. Oncotarget 2017, 8, 58353–58363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, H.; Lee, J.C.; Huang, H.C.; Huang, H.; Liu, H.K.; Huang, C. Delayed intervention with a novel SGLT2 inhibitor NGI001 suppresses diet-induced metabolic dysfunction and non-alcoholic fatty liver disease in mice. Br. J. Pharmacol. 2020, 177, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, Y.; Liu, Q.; Tang, Q.; Zhang, Z.; Zhang, J.; Huang, C.; Huang, H.; Zhang, G.; Zhou, J.; et al. Canagliflozin Facilitates Reverse Cholesterol Transport through Activation of AMPK/ABC Transporter Pathway. Drug Des. Devel. Ther. 2021, 15, 2117–2128. [Google Scholar] [CrossRef]
- Wang, B.; Tontonoz, P. Liver X receptors in lipid signalling and membrane homeostasis. Nat. Rev. Endocrinol. 2018, 14, 452–463. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, J.; Yu, S.J.; Ma, H.L.; Chen, J.; Ding, X.F.; Chen, G.; Liang, Y.; Zhang, Q. Sodium-glucose co-transporter-2 (SGLT-2) inhibition reduces glucose uptake to induce breast cancer cell growth arrest through AMPK/mTOR pathway. Biomed. Pharmacother. 2020, 132, 110821. [Google Scholar] [CrossRef]
- Xiao, Q.; Zhang, S.; Yang, C.; Du, R.; Zhao, J.; Li, J.; Xu, Y.; Qin, Y.; Gao, Y.; Huang, W. Ginsenoside Rg1 Ameliorates Palmitic Acid-Induced Hepatic Steatosis and Inflammation in HepG2 Cells via the AMPK/NF-kappaB Pathway. Int. J. Endocrinol. 2019, 2019, 7514802. [Google Scholar] [CrossRef] [Green Version]
- Soret, P.A.; Magusto, J.; Housset, C.; Gautheron, J. In Vitro and In Vivo Models of Non-Alcoholic Fatty Liver Disease: A Critical Appraisal. J. Clin. Med. 2020, 10, 36. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Goldin, R.D. Mouse models in non-alcoholic fatty liver disease and steatohepatitis research. Int. J. Exp. Pathol. 2006, 87, 1–16. [Google Scholar] [CrossRef]
- Ibrahim, S.H.; Hirsova, P.; Malhi, H.; Gores, G.J. Animal Models of Nonalcoholic Steatohepatitis: Eat, Delete, and Inflame. Dig. Dis. Sci. 2016, 61, 1325–1336. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Terauchi, Y. Lessons from mouse models of high-fat diet-induced NAFLD. Int. J. Mol. Sci. 2013, 14, 21240–21257. [Google Scholar] [CrossRef] [Green Version]
- Perakakis, N.; Chrysafi, P.; Feigh, M.; Veidal, S.S.; Mantzoros, C.S. Empagliflozin Improves Metabolic and Hepatic Outcomes in a Non-Diabetic Obese Biopsy-Proven Mouse Model of Advanced NASH. Int. J. Mol. Sci. 2021, 22, 6332. [Google Scholar] [CrossRef]
- Tahara, A.; Takasu, T. Therapeutic Effects of SGLT2 Inhibitor Ipragliflozin and Metformin on NASH in Type 2 Diabetic Mice. Endocr. Res. 2020, 45, 147–161. [Google Scholar] [CrossRef]
- Shiba, K.; Tsuchiya, K.; Komiya, C.; Miyachi, Y.; Mori, K.; Shimazu, N.; Yamaguchi, S.; Ogasawara, N.; Katoh, M.; Itoh, M.; et al. Canagliflozin, an SGLT2 inhibitor, attenuates the development of hepatocellular carcinoma in a mouse model of human NASH. Sci. Rep. 2018, 8, 2362. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.; Wilding, J.P.H.; Barber, T.M.; Alam, U.; Cuthbertson, D.J. Weight loss variability with SGLT2 inhibitors and GLP-1 receptor agonists in type 2 diabetes mellitus and obesity: Mechanistic possibilities. Obes. Rev. 2019, 20, 816–828. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Yang, W.; Gao, X.; Chen, Y.; Zhou, L.; Zhang, S.; Han, X.; Ji, L. The Association between the Dosage of SGLT2 Inhibitor and Weight Reduction in Type 2 Diabetes Patients: A Meta-Analysis. Obesity 2018, 26, 70–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherney, D.Z.I.; Dekkers, C.C.J.; Barbour, S.J.; Cattran, D.; Abdul Gafor, A.H.; Greasley, P.J.; Laverman, G.D.; Lim, S.K.; Di Tanna, G.L.; Reich, H.N.; et al. Effects of the SGLT2 inhibitor dapagliflozin on proteinuria in non-diabetic patients with chronic kidney disease (DIAMOND): A randomised, double-blind, crossover trial. Lancet Diabetes Endocrinol. 2020, 8, 582–593. [Google Scholar] [CrossRef]
- Han, T.; Fan, Y.J.; Gao, J.; Fatima, M.R.; Zhang, Y.L.; Ding, Y.M.; Bai, L.A.; Wang, C.X. Sodium glucose cotransporter 2 inhibitor dapagliflozin depressed adiposity and ameliorated hepatic steatosis in high-fat diet induced obese mice. Adipocyte 2021, 10, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Obata, A.; Kubota, N.; Kubota, T.; Iwamoto, M.; Sato, H.; Sakurai, Y.; Takamoto, I.; Katsuyama, H.; Suzuki, Y.; Fukazawa, M.; et al. Tofogliflozin Improves Insulin Resistance in Skeletal Muscle and Accelerates Lipolysis in Adipose Tissue in Male Mice. Endocrinology 2016, 157, 1029–1042. [Google Scholar] [CrossRef]
- Petito-da-Silva, T.I.; Souza-Mello, V.; Barbosa-da-Silva, S. Empaglifozin mitigates NAFLD in high-fat-fed mice by alleviating insulin resistance, lipogenesis and ER stress. Mol. Cell Endocrinol. 2019, 498, 110539. [Google Scholar] [CrossRef]
- Tanaka, K.; Takahashi, H.; Katagiri, S.; Sasaki, K.; Ohsugi, Y.; Watanabe, K.; Rasadul, I.M.D.; Mine, K.; Nagafuchi, S.; Iwata, T.; et al. Combined effect of canagliflozin and exercise training on high-fat diet-fed mice. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E492–E503. [Google Scholar] [CrossRef]
- Yabiku, K.; Nakamoto, K.; Tsubakimoto, M. Effects of Sodium-Glucose Cotransporter 2 Inhibition on Glucose Metabolism, Liver Function, Ascites, and Hemodynamics in a Mouse Model of Nonalcoholic Steatohepatitis and Type 2 Diabetes. J. Diabetes Res. 2020, 2020, 1682904. [Google Scholar] [CrossRef]
- Ozutsumi, T.; Namisaki, T.; Shimozato, N.; Kaji, K.; Tsuji, Y.; Kaya, D.; Fujinaga, Y.; Furukawa, M.; Nakanishi, K.; Sato, S.; et al. Combined Treatment with Sodium-Glucose Cotransporter-2 Inhibitor (Canagliflozin) and Dipeptidyl Peptidase-4 Inhibitor (Teneligliptin) Alleviates NASH Progression in A Non-Diabetic Rat Model of Steatohepatitis. Int. J. Mol. Sci. 2020, 21, 2164. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, K.; Hosooka, T.; Shinohara, M.; Aoki, C.; Hosokawa, Y.; Imamori, M.; Ogawa, W. Canagliflozin ameliorates hepatic fat deposition in obese diabetic mice: Role of prostaglandin E2. Biochem. Biophys. Res. Commun. 2021, 557, 62–68. [Google Scholar] [CrossRef]
- Komiya, C.; Tsuchiya, K.; Shiba, K.; Miyachi, Y.; Furuke, S.; Shimazu, N.; Yamaguchi, S.; Kanno, K.; Ogawa, Y. Ipragliflozin Improves Hepatic Steatosis in Obese Mice and Liver Dysfunction in Type 2 Diabetic Patients Irrespective of Body Weight Reduction. PLoS ONE 2016, 11, e0151511. [Google Scholar] [CrossRef] [Green Version]
- Tahara, A.; Takasu, T. SGLT2 inhibitor ipragliflozin alone and combined with pioglitazone prevents progression of nonalcoholic steatohepatitis in a type 2 diabetes rodent model. Physiol. Rep. 2019, 7, e14286. [Google Scholar] [CrossRef] [Green Version]
- Honda, Y.; Imajo, K.; Kato, T.; Kessoku, T.; Ogawa, Y.; Tomeno, W.; Kato, S.; Mawatari, H.; Fujita, K.; Yoneda, M.; et al. The Selective SGLT2 Inhibitor Ipragliflozin Has a Therapeutic Effect on Nonalcoholic Steatohepatitis in Mice. PLoS ONE 2016, 11, e0146337. [Google Scholar] [CrossRef] [Green Version]
- Huttl, M.; Markova, I.; Miklankova, D.; Zapletalova, I.; Poruba, M.; Haluzik, M.; Vaneckova, I.; Malinska, H. In a Prediabetic Model, Empagliflozin Improves Hepatic Lipid Metabolism Independently of Obesity and before Onset of Hyperglycemia. Int. J. Mol. Sci. 2021, 22, 11513. [Google Scholar] [CrossRef]
- Steven, S.; Oelze, M.; Hanf, A.; Kroller-Schon, S.; Kashani, F.; Roohani, S.; Welschof, P.; Kopp, M.; Godtel-Armbrust, U.; Xia, N.; et al. The SGLT2 inhibitor empagliflozin improves the primary diabetic complications in ZDF rats. Redox Biol. 2017, 13, 370–385. [Google Scholar] [CrossRef]
- Meng, Z.; Liu, X.; Li, T.; Fang, T.; Cheng, Y.; Han, L.; Sun, B.; Chen, L. The SGLT2 inhibitor empagliflozin negatively regulates IL-17/IL-23 axis-mediated inflammatory responses in T2DM with NAFLD via the AMPK/mTOR/autophagy pathway. Int. Immunopharmacol. 2021, 94, 107492. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Roles of Diacylglycerols and Ceramides in Hepatic Insulin Resistance. Trends Pharmacol. Sci. 2017, 38, 649–665. [Google Scholar] [CrossRef]
- Chambel, S.S.; Santos-Goncalves, A.; Duarte, T.L. The Dual Role of Nrf2 in Nonalcoholic Fatty Liver Disease: Regulation of Antioxidant Defenses and Hepatic Lipid Metabolism. Biomed. Res. Int. 2015, 2015, 597134. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Kong, Y.; Peng, D. Fibroblast growth factor 21 in lipid metabolism and non-alcoholic fatty liver disease. Clin. Chim. Acta 2019, 498, 30–37. [Google Scholar] [CrossRef]
- Eberle, D.; Hegarty, B.; Bossard, P.; Ferre, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Kern, M.; Kloting, N.; Mark, M.; Mayoux, E.; Klein, T.; Bluher, M. The SGLT2 inhibitor empagliflozin improves insulin sensitivity in db/db mice both as monotherapy and in combination with linagliptin. Metabolism 2016, 65, 114–123. [Google Scholar] [CrossRef]
- Bakan, I.; Laplante, M. Connecting mTORC1 signaling to SREBP-1 activation. Curr. Opin. Lipidol. 2012, 23, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Souza-Mello, V. Peroxisome proliferator-activated receptors as targets to treat non-alcoholic fatty liver disease. World J. Hepatol. 2015, 7, 1012–1019. [Google Scholar] [CrossRef]
- Omori, K.; Nakamura, A.; Miyoshi, H.; Takahashi, K.; Kitao, N.; Nomoto, H.; Kameda, H.; Cho, K.Y.; Takagi, R.; Hatanaka, K.C.; et al. Effects of dapagliflozin and/or insulin glargine on beta cell mass and hepatic steatosis in db/db mice. Metabolism 2019, 98, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, N.; Tanaka, M.; Ochi, K.; Watanabe, A.; Ono, K.; Sawada, M.; Ogi, T.; Itoh, M.; Ito, A.; Shiraki, Y.; et al. The sodium-glucose cotransporter-2 inhibitor Tofogliflozin prevents the progression of nonalcoholic steatohepatitis-associated liver tumors in a novel murine model. Biomed. Pharmacother. 2021, 140, 111738. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.M.; Meers, G.M.; Booth, F.W.; Fritsche, K.L.; Hardin, C.D.; Thyfault, J.P.; Ibdah, J.A. PGC-1alpha overexpression results in increased hepatic fatty acid oxidation with reduced triacylglycerol accumulation and secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G979–G992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrysavgis, L.; Papatheodoridi, A.M.; Chatzigeorgiou, A.; Cholongitas, E. The impact of sodium glucose co-transporter 2 inhibitors on non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2021, 36, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Kabil, S.L.; Mahmoud, N.M. Canagliflozin protects against non-alcoholic steatohepatitis in type-2 diabetic rats through zinc alpha-2 glycoprotein up-regulation. Eur. J. Pharmacol. 2018, 828, 135–145. [Google Scholar] [CrossRef] [PubMed]
- ElMahdy, M.K.; Helal, M.G.; Ebrahim, T.M. Potential anti-inflammatory effect of dapagliflozin in HCHF diet- induced fatty liver degeneration through inhibition of TNF-alpha, IL-1beta, and IL-18 in rat liver. Int. Immunopharmacol. 2020, 86, 106730. [Google Scholar] [CrossRef] [PubMed]
- Tahara, A.; Kurosaki, E.; Yokono, M.; Yamajuku, D.; Kihara, R.; Hayashizaki, Y.; Takasu, T.; Imamura, M.; Li, Q.; Tomiyama, H.; et al. Effects of SGLT2 selective inhibitor ipragliflozin on hyperglycemia, hyperlipidemia, hepatic steatosis, oxidative stress, inflammation, and obesity in type 2 diabetic mice. Eur. J. Pharmacol. 2013, 715, 246–255. [Google Scholar] [CrossRef]
- Tang, L.; Wu, Y.; Tian, M.; Sjostrom, C.D.; Johansson, U.; Peng, X.R.; Smith, D.M.; Huang, Y. Dapagliflozin slows the progression of the renal and liver fibrosis associated with type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E563–E576. [Google Scholar] [CrossRef]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef]
- Nakano, S.; Katsuno, K.; Isaji, M.; Nagasawa, T.; Buehrer, B.; Walker, S.; Wilkison, W.O.; Cheatham, B. Remogliflozin Etabonate Improves Fatty Liver Disease in Diet-Induced Obese Male Mice. J. Clin. Exp. Hepatol. 2015, 5, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Ookawara, M.; Matsuda, K.; Watanabe, M.; Moritoh, Y. The GPR40 Full Agonist SCO-267 Improves Liver Parameters in a Mouse Model of Nonalcoholic Fatty Liver Disease without Affecting Glucose or Body Weight. J. Pharmacol. Exp. Ther. 2020, 375, 21–27. [Google Scholar] [CrossRef]
- Jojima, T.; Tomotsune, T.; Iijima, T.; Akimoto, K.; Suzuki, K.; Aso, Y. Empagliflozin (an SGLT2 inhibitor), alone or in combination with linagliptin (a DPP-4 inhibitor), prevents steatohepatitis in a novel mouse model of non-alcoholic steatohepatitis and diabetes. Diabetol. Metab. Syndr. 2016, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Hupa-Breier, K.L.; Dywicki, J.; Hartleben, B.; Wellhoner, F.; Heidrich, B.; Taubert, R.; Mederacke, Y.E.; Lieber, M.; Iordanidis, K.; Manns, M.P.; et al. Dulaglutide Alone and in Combination with Empagliflozin Attenuate Inflammatory Pathways and Microbiome Dysbiosis in a Non-Diabetic Mouse Model of NASH. Biomedicines 2021, 9, 353. [Google Scholar] [CrossRef]
- Kuwako, K.I.; Okano, H. Versatile Roles of LKB1 Kinase Signaling in Neural Development and Homeostasis. Front. Mol. Neurosci. 2018, 11, 354. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Y.; Yang, L.; Liu, P.; Zhang, Y.; Wang, X. MiR-149 attenuates endoplasmic reticulum stress-induced inflammation and apoptosis in nonalcoholic fatty liver disease by negatively targeting ATF6 pathway. Immunol. Lett. 2020, 222, 40–48. [Google Scholar] [CrossRef]
- Heyens, L.J.M.; Busschots, D.; Koek, G.H.; Robaeys, G.; Francque, S. Liver Fibrosis in Non-alcoholic Fatty Liver Disease: From Liver Biopsy to Non-invasive Biomarkers in Diagnosis and Treatment. Front. Med. 2021, 8, 615978. [Google Scholar] [CrossRef]
- Zisser, A.; Ipsen, D.H.; Tveden-Nyborg, P. Hepatic Stellate Cell Activation and Inactivation in NASH-Fibrosis-Roles as Putative Treatment Targets? Biomedicines 2021, 9, 365. [Google Scholar] [CrossRef]
- Qiang, S.; Nakatsu, Y.; Seno, Y.; Fujishiro, M.; Sakoda, H.; Kushiyama, A.; Mori, K.; Matsunaga, Y.; Yamamotoya, T.; Kamata, H.; et al. Treatment with the SGLT2 inhibitor luseogliflozin improves nonalcoholic steatohepatitis in a rodent model with diabetes mellitus. Diabetol. Metab. Syndr. 2015, 7, 104. [Google Scholar] [CrossRef] [Green Version]
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastro Hepat. 2019, 16, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Hayashizaki-Someya, Y.; Kurosaki, E.; Takasu, T.; Mitori, H.; Yamazaki, S.; Koide, K.; Takakura, S. Ipragliflozin, an SGLT2 inhibitor, exhibits a prophylactic effect on hepatic steatosis and fibrosis induced by choline-deficient l-amino acid-defined diet in rats. Eur. J. Pharmacol. 2015, 754, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Bando, Y.; Ogawa, A.; Ishikura, K.; Kanehara, H.; Hisada, A.; Notumata, K.; Okafuji, K.; Toya, D. The effects of ipragliflozin on the liver-to-spleen attenuation ratio as assessed by computed tomography and on alanine transaminase levels in Japanese patients with type 2 diabetes mellitus. Diabetol. Int. 2017, 8, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Akuta, N.; Watanabe, C.; Kawamura, Y.; Arase, Y.; Saitoh, S.; Fujiyama, S.; Sezaki, H.; Hosaka, T.; Kobayashi, M.; Kobayashi, M.; et al. Effects of a sodium-glucose cotransporter 2 inhibitor in nonalcoholic fatty liver disease complicated by diabetes mellitus: Preliminary prospective study based on serial liver biopsies. Hepatol. Commun. 2017, 1, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Shimizu, S.; Inoue, K.; Saito, D.; Yanagisawa, M.; Inukai, K.; Akiyama, Y.; Morimoto, Y.; Noda, M.; Shimada, A. Comparison of Ipragliflozin and Pioglitazone Effects on Nonalcoholic Fatty Liver Disease in Patients with Type 2 Diabetes: A Randomized, 24-Week, Open-Label, Active-Controlled Trial. Diabetes Care 2017, 40, 1364–1372. [Google Scholar] [CrossRef] [Green Version]
- Seko, Y.; Sumida, Y.; Tanaka, S.; Mori, K.; Taketani, H.; Ishiba, H.; Hara, T.; Okajima, A.; Umemura, A.; Nishikawa, T.; et al. Effect of sodium glucose cotransporter 2 inhibitor on liver function tests in Japanese patients with non-alcoholic fatty liver disease and type 2 diabetes mellitus. Hepatol. Res. 2017, 47, 1072–1078. [Google Scholar] [CrossRef]
- Choi, D.H.; Jung, C.H.; Mok, J.O.; Kim, C.H.; Kang, S.K.; Kim, B.Y. Effect of Dapagliflozin on Alanine Aminotransferase Improvement in Type 2 Diabetes Mellitus with Non-alcoholic Fatty Liver Disease. Endocrinol. Metab. 2018, 33, 387–394. [Google Scholar] [CrossRef]
- Eriksson, J.W.; Lundkvist, P.; Jansson, P.A.; Johansson, L.; Kvarnström, M.; Moris, L.; Miliotis, T.; Forsberg, G.B.; Risérus, U.; Lind, L.; et al. Effects of dapagliflozin and n-3 carboxylic acids on non-alcoholic fatty liver disease in people with type 2 diabetes: A double-blind randomised placebo-controlled study. Diabetologia 2018, 61, 1923–1934. [Google Scholar] [CrossRef] [Green Version]
- Itani, T.; Ishihara, T. Efficacy of canagliflozin against nonalcoholic fatty liver disease: A prospective cohort study. Obes. Sci. Pract. 2018, 4, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Kuchay, M.S.; Krishan, S.; Mishra, S.K.; Farooqui, K.J.; Singh, M.K.; Wasir, J.S.; Bansal, B.; Kaur, P.; Jevalikar, G.; Gill, H.K.; et al. Effect of Empagliflozin on Liver Fat in Patients with Type 2 Diabetes and Nonalcoholic Fatty Liver Disease: A Randomized Controlled Trial (E-LIFT Trial). Diabetes Care 2018, 41, 1801–1808. [Google Scholar] [CrossRef] [Green Version]
- Miyake, T.; Yoshida, S.; Furukawa, S.; Sakai, T.; Tada, F.; Senba, H.; Yamamoto, S.; Koizumi, Y.; Yoshida, O.; Hirooka, M.; et al. Ipragliflozin Ameliorates Liver Damage in Non-alcoholic Fatty Liver Disease. Open Med. 2018, 13, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, T.; Fushimi, N.; Kawai, M.; Yoshida, Y.; Hachiya, H.; Ito, S.; Kawai, H.; Ohashi, N.; Mori, A. Luseogliflozin improves liver fat deposition compared to metformin in type 2 diabetes patients with non-alcoholic fatty liver disease: A prospective randomized controlled pilot study. Diabetes Obes. Metab. 2018, 20, 438–442. [Google Scholar] [CrossRef]
- Shimizu, M.; Suzuki, K.; Kato, K.; Jojima, T.; Iijima, T.; Murohisa, T.; Iijima, M.; Takekawa, H.; Usui, I.; Hiraishi, H.; et al. Evaluation of the effects of dapagliflozin, a sodium-glucose co-transporter-2 inhibitor, on hepatic steatosis and fibrosis using transient elastography in patients with type 2 diabetes and non-alcoholic fatty liver disease. Diabetes Obes. Metab. 2019, 21, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Murotani, K.; Saito, M.; Tamasawa, A.; Osonoi, Y.; Yoneda, M.; Osonoi, T. Effect of luseogliflozin on hepatic fat content in type 2 diabetes patients with non-alcoholic fatty liver disease: A prospective, single-arm trial (LEAD trial). Hepatol. Res. 2019, 49, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akuta, N.; Kawamura, Y.; Watanabe, C.; Nishimura, A.; Okubo, M.; Mori, Y.; Fujiyama, S.; Sezaki, H.; Hosaka, T.; Kobayashi, M.; et al. Impact of sodium glucose cotransporter 2 inhibitor on histological features and glucose metabolism of non-alcoholic fatty liver disease complicated by diabetes mellitus. Hepatol. Res. 2019, 49, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Yamashima, M.; Miyaaki, H.; Miuma, S.; Shibata, H.; Sasaki, R.; Haraguchi, M.; Fukushima, M.; Nakao, K. The Long-term Efficacy of Sodium Glucose Co-transporter 2 Inhibitor in Patients with Non-alcoholic Fatty Liver Disease. Intern. Med. 2019, 58, 1987–1992. [Google Scholar] [CrossRef] [Green Version]
- Akuta, N.; Kawamura, Y.; Fujiyama, S.; Sezaki, H.; Hosaka, T.; Kobayashi, M.; Kobayashi, M.; Saitoh, S.; Suzuki, F.; Suzuki, Y.; et al. SGLT2 Inhibitor Treatment Outcome in Nonalcoholic Fatty Liver Disease Complicated with Diabetes Mellitus: The Long-term Effects on Clinical Features and Liver Histopathology. Intern. Med. 2020, 59, 1931–1937. [Google Scholar] [CrossRef]
- Han, E.; Lee, Y.H.; Lee, B.W.; Kang, E.S.; Cha, B.S. Ipragliflozin Additively Ameliorates Non-Alcoholic Fatty Liver Disease in Patients with Type 2 Diabetes Controlled with Metformin and Pioglitazone: A 24-Week Randomized Controlled Trial. J. Clin. Med. 2020, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Kahl, S.; Gancheva, S.; Straßburger, K.; Herder, C.; Machann, J.; Katsuyama, H.; Kabisch, S.; Henkel, E.; Kopf, S.; Lagerpusch, M.; et al. Empagliflozin Effectively Lowers Liver Fat Content in Well-Controlled Type 2 Diabetes: A Randomized, Double-Blind, Phase 4, Placebo-Controlled Trial. Diabetes Care 2020, 43, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Shimoda, M.; Nakashima, K.; Fushimi, Y.; Hirata, Y.; Tanabe, A.; Tatsumi, F.; Hirukawa, H.; Sanada, J.; Kohara, K.; et al. Comparison of the effects of three kinds of glucose-lowering drugs on non-alcoholic fatty liver disease in patients with type 2 diabetes: A randomized, open-label, three-arm, active control study. J. Diabetes Investig. 2020, 11, 1612–1622. [Google Scholar] [CrossRef]
- Yano, K.; Seko, Y.; Takahashi, A.; Okishio, S.; Kataoka, S.; Takemura, M.; Okuda, K.; Mizuno, N.; Taketani, H.; Umemura, A.; et al. Effect of Sodium Glucose Cotransporter 2 Inhibitors on Renal Function in Patients with Nonalcoholic Fatty Liver Disease and Type 2 Diabetes in Japan. Diagnostics 2020, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Atsukawa, M.; Tsubota, A.; Mikami, S.; Ono, H.; Kawano, T.; Yoshida, Y.; Tanabe, T.; Okubo, T.; Hayama, K.; et al. Effect of sodium-glucose cotransporter 2 inhibitor in patients with non-alcoholic fatty liver disease and type 2 diabetes mellitus: A propensity score-matched analysis of real-world data. Ther. Adv. Endocrinol. Metab. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Chehrehgosha, H.; Sohrabi, M.R.; Ismail-Beigi, F.; Malek, M.; Reza Babaei, M.; Zamani, F.; Ajdarkosh, H.; Khoonsari, M.; Fallah, A.E.; Khamseh, M.E. Empagliflozin Improves Liver Steatosis and Fibrosis in Patients with Non-Alcoholic Fatty Liver Disease and Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Diabetes Ther. 2021, 12, 843–861. [Google Scholar] [CrossRef]
- Euh, W.; Lim, S.; Kim, J.W. Sodium-Glucose Cotransporter-2 Inhibitors Ameliorate Liver Enzyme Abnormalities in Korean Patients with Type 2 Diabetes Mellitus and Nonalcoholic Fatty Liver Disease. Front. Endocrinol. 2021, 12, 613389. [Google Scholar] [CrossRef]
- Gaborit, B.; Ancel, P.; Abdullah, A.E.; Maurice, F.; Abdesselam, I.; Calen, A.; Soghomonian, A.; Houssays, M.; Varlet, I.; Eisinger, M.; et al. Effect of empagliflozin on ectopic fat stores and myocardial energetics in type 2 diabetes: The EMPACEF study. Cardiovasc. Diabetol. 2021, 20, 57. [Google Scholar] [CrossRef]
- Takahashi, H.; Kessoku, T.; Kawanaka, M.; Nonaka, M.; Hyogo, H.; Fujii, H.; Nakajima, T.; Imajo, K.; Tanaka, K.; Kubotsu, Y.; et al. Ipragliflozin Improves the Hepatic Outcomes of Patients with Diabetes with NAFLD. Hepatol. Commun. 2021, 6, 120–132. [Google Scholar] [CrossRef]
- Yoneda, M.; Honda, Y.; Ogawa, Y.; Kessoku, T.; Kobayashi, T.; Imajo, K.; Ozaki, A.; Nogami, A.; Taguri, M.; Yamanaka, T.; et al. Comparing the effects of tofogliflozin and pioglitazone in non-alcoholic fatty liver disease patients with type 2 diabetes mellitus (ToPiND study): A randomized prospective open-label controlled trial. BMJ Open Diabetes Res. Care 2021, 9, e001990. [Google Scholar] [CrossRef]
- Pradhan, R.; Yin, H.; Yu, O.; Azoulay, L. Glucagon-Like Peptide 1 Receptor Agonists and Sodium-Glucose Cotransporter 2 Inhibitors and Risk of Nonalcoholic Fatty Liver Disease Among Patients with Type 2 Diabetes. Diabetes Care 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Sinha, B.; Datta, D.; Ghosal, S. Meta-analysis of the effects of sodium glucose cotransporter 2 inhibitors in non-alcoholic fatty liver disease patients with type 2 diabetes. JGH Open 2021, 5, 219–227. [Google Scholar] [CrossRef]
- Mo, M.; Huang, Z.; Liang, Y.; Liao, Y.; Xia, N. The safety and efficacy evaluation of Sodium-Glucose co-transporter 2 inhibitors for patients with non-alcoholic fatty Liver disease: An updated meta-analysis. Dig. Liver Dis. 2021, in press. [Google Scholar] [CrossRef]
- Wong, C.; Yaow, C.Y.L.; Ng, C.H.; Chin, Y.H.; Low, Y.F.; Lim, A.Y.L.; Muthiah, M.D.; Khoo, C.M. Sodium-Glucose Co-Transporter 2 Inhibitors for Non-Alcoholic Fatty Liver Disease in Asian Patients with Type 2 Diabetes: A Meta-Analysis. Front. Endocrinol. 2020, 11, 609135. [Google Scholar] [CrossRef] [PubMed]
- Tobita, H.; Yazaki, T.; Kataoka, M.; Kotani, S.; Oka, A.; Mishiro, T.; Oshima, N.; Kawashima, K.; Ishimura, N.; Naora, K.; et al. Comparison of dapagliflozin and teneligliptin in nonalcoholic fatty liver disease patients without type 2 diabetes mellitus: A prospective randomized study. J. Clin. Biochem. Nutr. 2021, 68, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Daniele, G.; Xiong, J.; Solis-Herrera, C.; Merovci, A.; Eldor, R.; Tripathy, D.; DeFronzo, R.A.; Norton, L.; Abdul-Ghani, M. Dapagliflozin Enhances Fat Oxidation and Ketone Production in Patients with Type 2 Diabetes. Diabetes Care 2016, 39, 2036–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Luo, Y.; Wang, X.; Orlicky, D.J.; Myakala, K.; Yang, P.; Levi, M. The Sodium-Glucose Cotransporter 2 Inhibitor Dapagliflozin Prevents Renal and Liver Disease in Western Diet Induced Obesity Mice. Int. J. Mol. Sci. 2018, 19, 137. [Google Scholar] [CrossRef] [Green Version]
- Bonner, C.; Kerr-Conte, J.; Gmyr, V.; Queniat, G.; Moerman, E.; Thévenet, J.; Beaucamps, C.; Delalleau, N.; Popescu, I.; Malaisse, W.J.; et al. Inhibition of the glucose transporter SGLT2 with dapagliflozin in pancreatic alpha cells triggers glucagon secretion. Nat. Med. 2015, 21, 512–517. [Google Scholar] [CrossRef]
- Ferrannini, E.; Baldi, S.; Frascerra, S.; Astiarraga, B.; Heise, T.; Bizzotto, R.; Mari, A.; Pieber, T.R.; Muscelli, E. Shift to Fatty Substrate Utilization in Response to Sodium-Glucose Cotransporter 2 Inhibition in Subjects without Diabetes and Patients with Type 2 Diabetes. Diabetes 2016, 65, 1190–1195. [Google Scholar] [CrossRef] [Green Version]
- Yokono, M.; Takasu, T.; Hayashizaki, Y.; Mitsuoka, K.; Kihara, R.; Muramatsu, Y.; Miyoshi, S.; Tahara, A.; Kurosaki, E.; Li, Q.; et al. SGLT2 selective inhibitor ipragliflozin reduces body fat mass by increasing fatty acid oxidation in high-fat diet-induced obese rats. Eur. J. Pharmacol. 2014, 727, 66–74. [Google Scholar] [CrossRef]
- Terami, N.; Ogawa, D.; Tachibana, H.; Hatanaka, T.; Wada, J.; Nakatsuka, A.; Eguchi, J.; Horiguchi, C.S.; Nishii, N.; Yamada, H.; et al. Long-term treatment with the sodium glucose cotransporter 2 inhibitor, dapagliflozin, ameliorates glucose homeostasis and diabetic nephropathy in db/db mice. PLoS ONE 2014, 9, e100777. [Google Scholar] [CrossRef]
- Tahara, A.; Kurosaki, E.; Yokono, M.; Yamajuku, D.; Kihara, R.; Hayashizaki, Y.; Takasu, T.; Imamura, M.; Li, Q.; Tomiyama, H.; et al. Effects of sodium-glucose cotransporter 2 selective inhibitor ipragliflozin on hyperglycaemia, oxidative stress, inflammation and liver injury in streptozotocin-induced type 1 diabetic rats. J. Pharm. Pharmacol. 2014, 66, 975–987. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Atkin, S.L.; Butler, A.E.; Sahebkar, A. Sodium-glucose cotransporter inhibitors and oxidative stress: An update. J. Cell Physiol. 2019, 234, 3231–3237. [Google Scholar] [CrossRef]
- Oshima, H.; Miki, T.; Kuno, A.; Mizuno, M.; Sato, T.; Tanno, M.; Yano, T.; Nakata, K.; Kimura, Y.; Abe, K.; et al. Empagliflozin, an SGLT2 Inhibitor, Reduced the Mortality Rate after Acute Myocardial Infarction with Modification of Cardiac Metabolomes and Antioxidants in Diabetic Rats. J. Pharmacol. Exp. Ther. 2019, 368, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Osorio, H.; Coronel, I.; Arellano, A.; Pacheco, U.; Bautista, R.; Franco, M.; Escalante, B. Sodium-glucose cotransporter inhibition prevents oxidative stress in the kidney of diabetic rats. Oxid Med. Cell Longev. 2012, 2012, 542042. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).