
PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
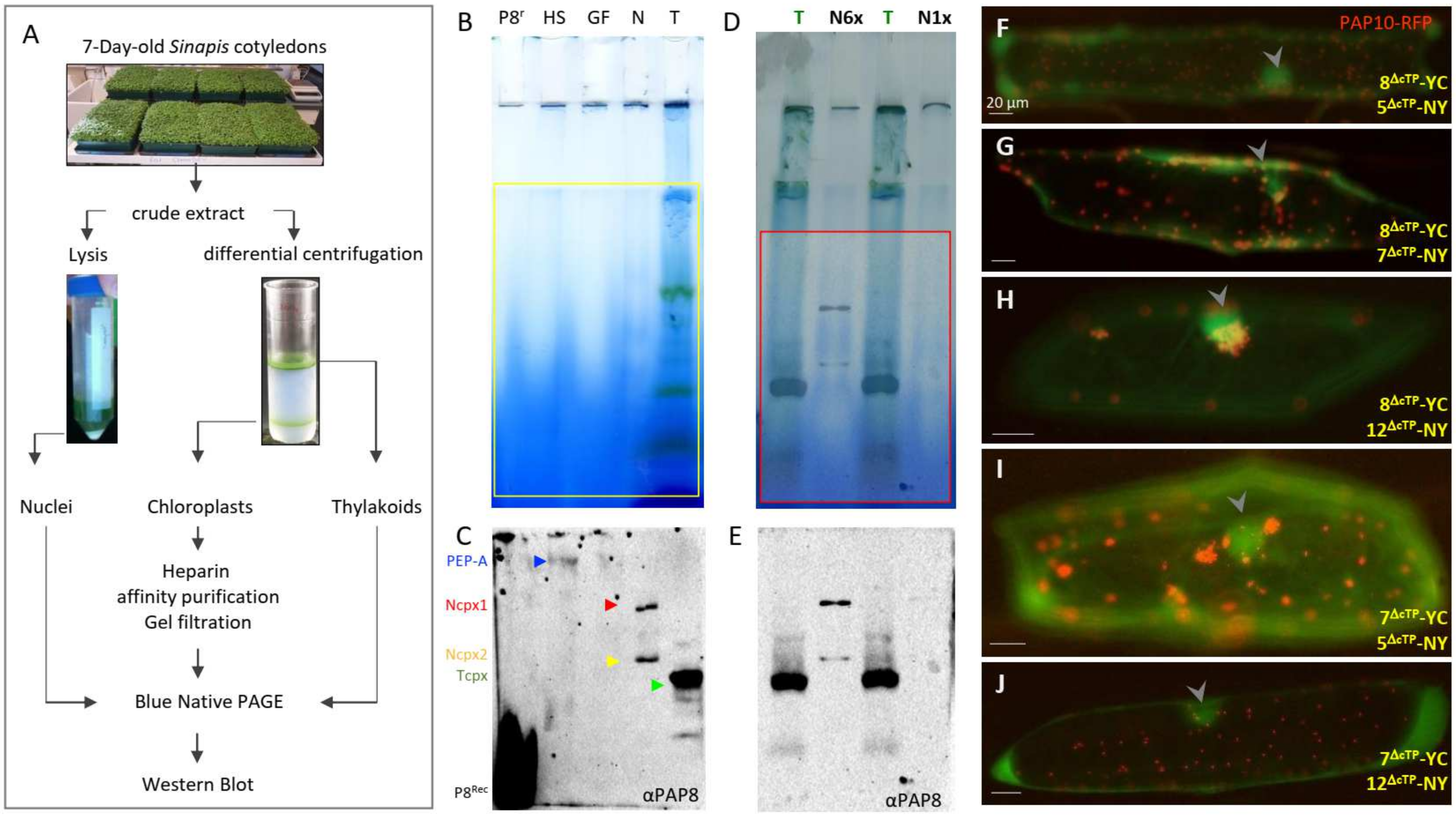
2.1. PAP8 Is Found in a Nuclear Complex during Photomorphogenesis
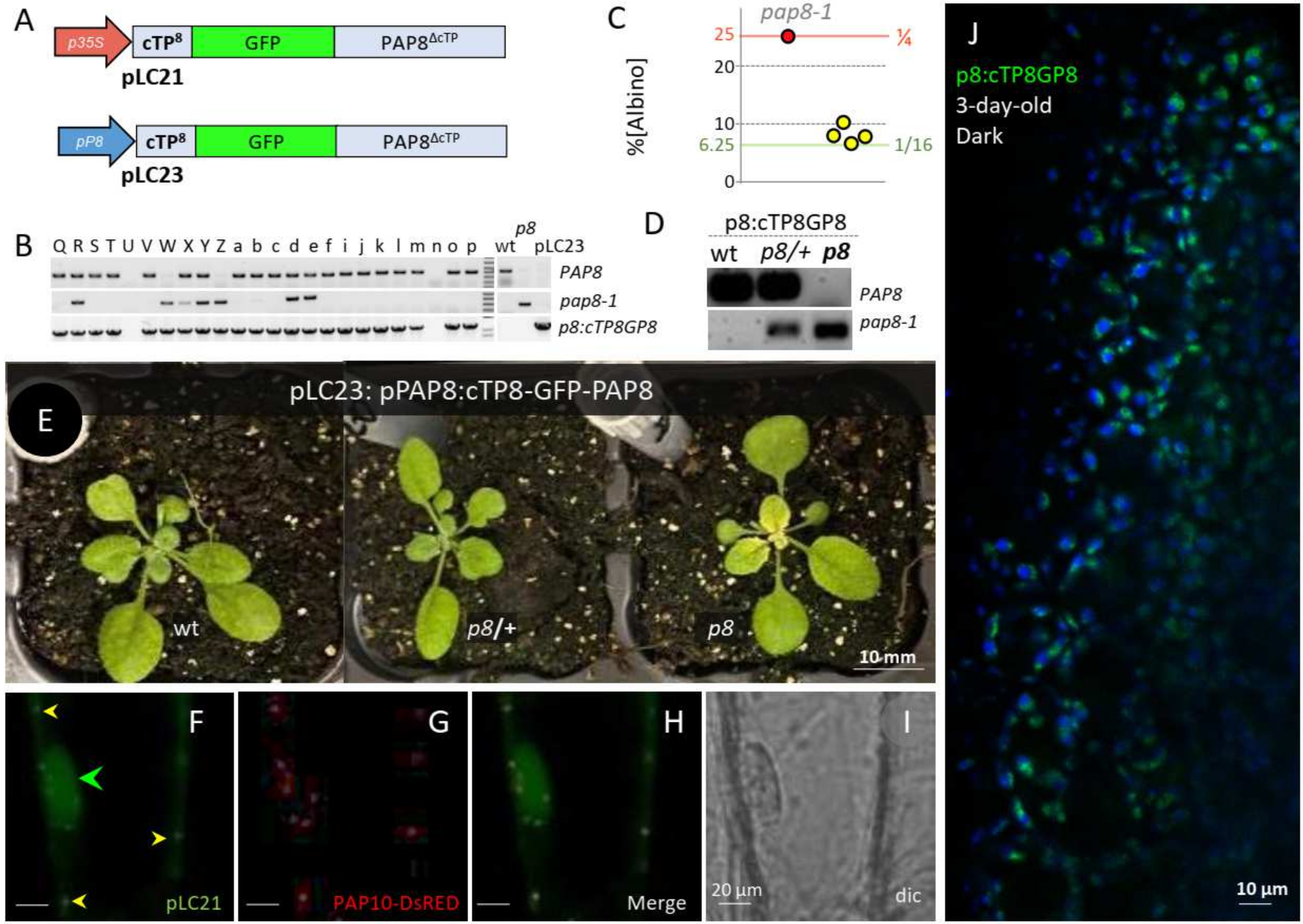
2.2. PAP8 Functional Fusions
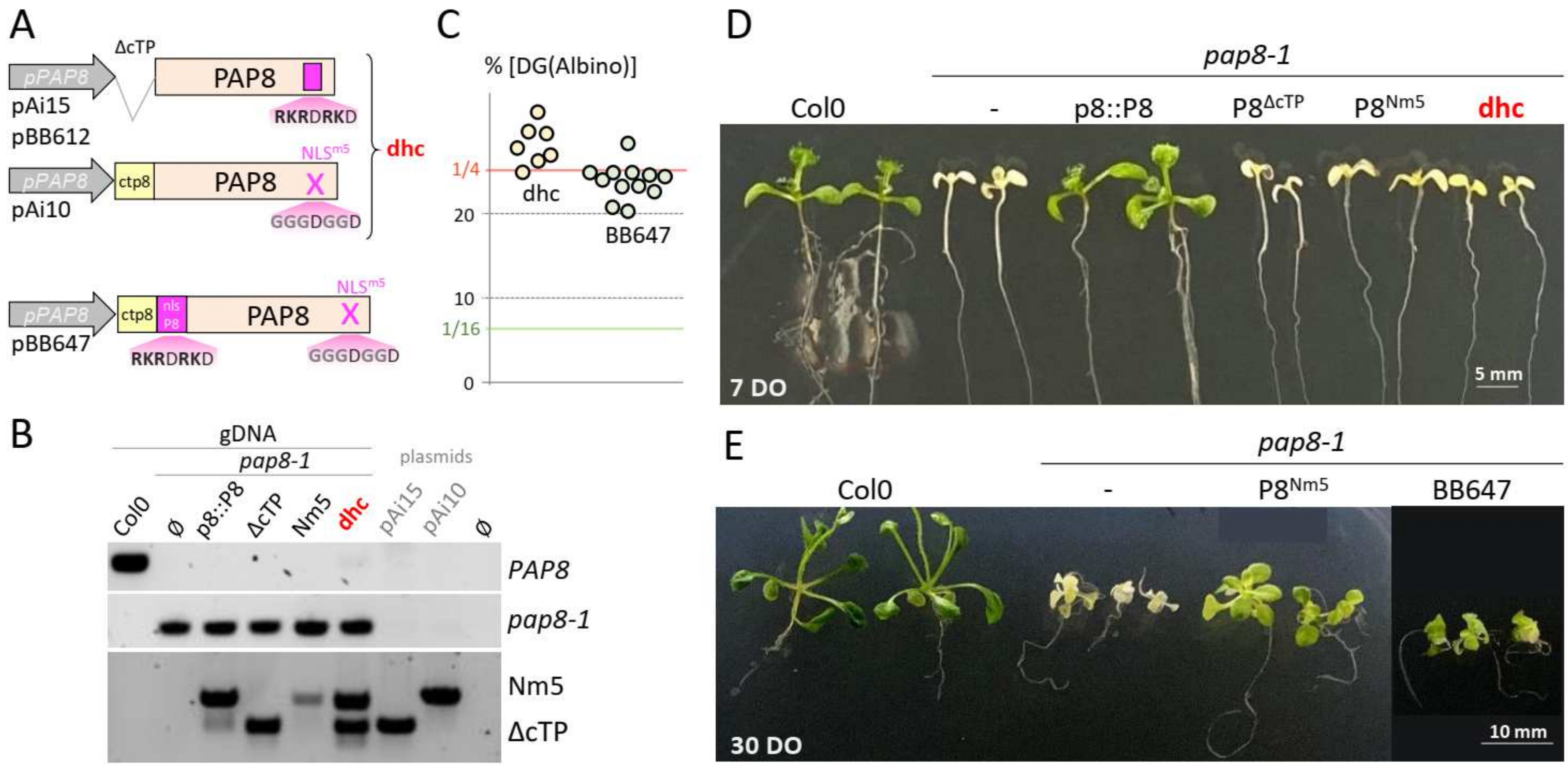
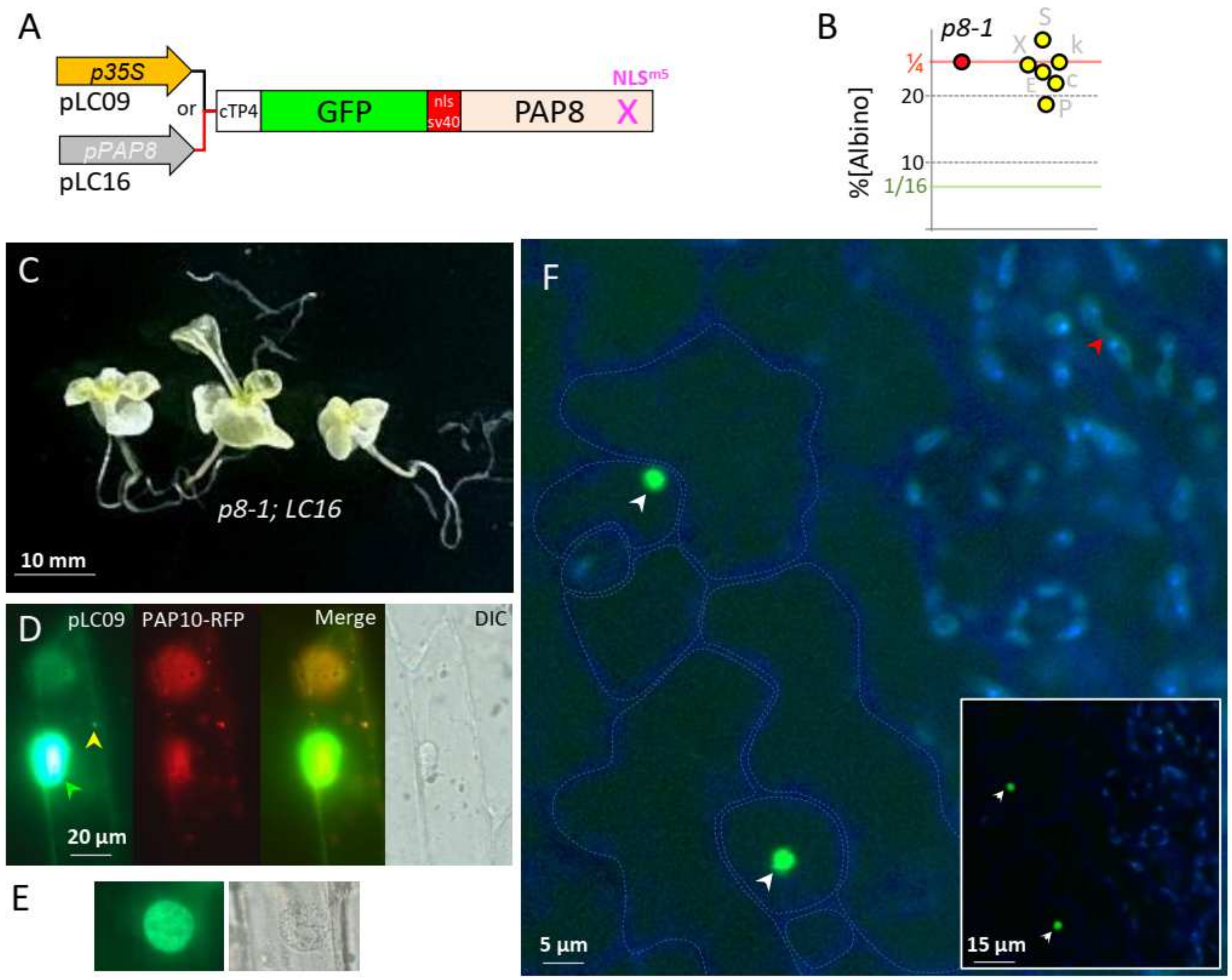
2.3. Uncoupling Localization and Function of PAP8
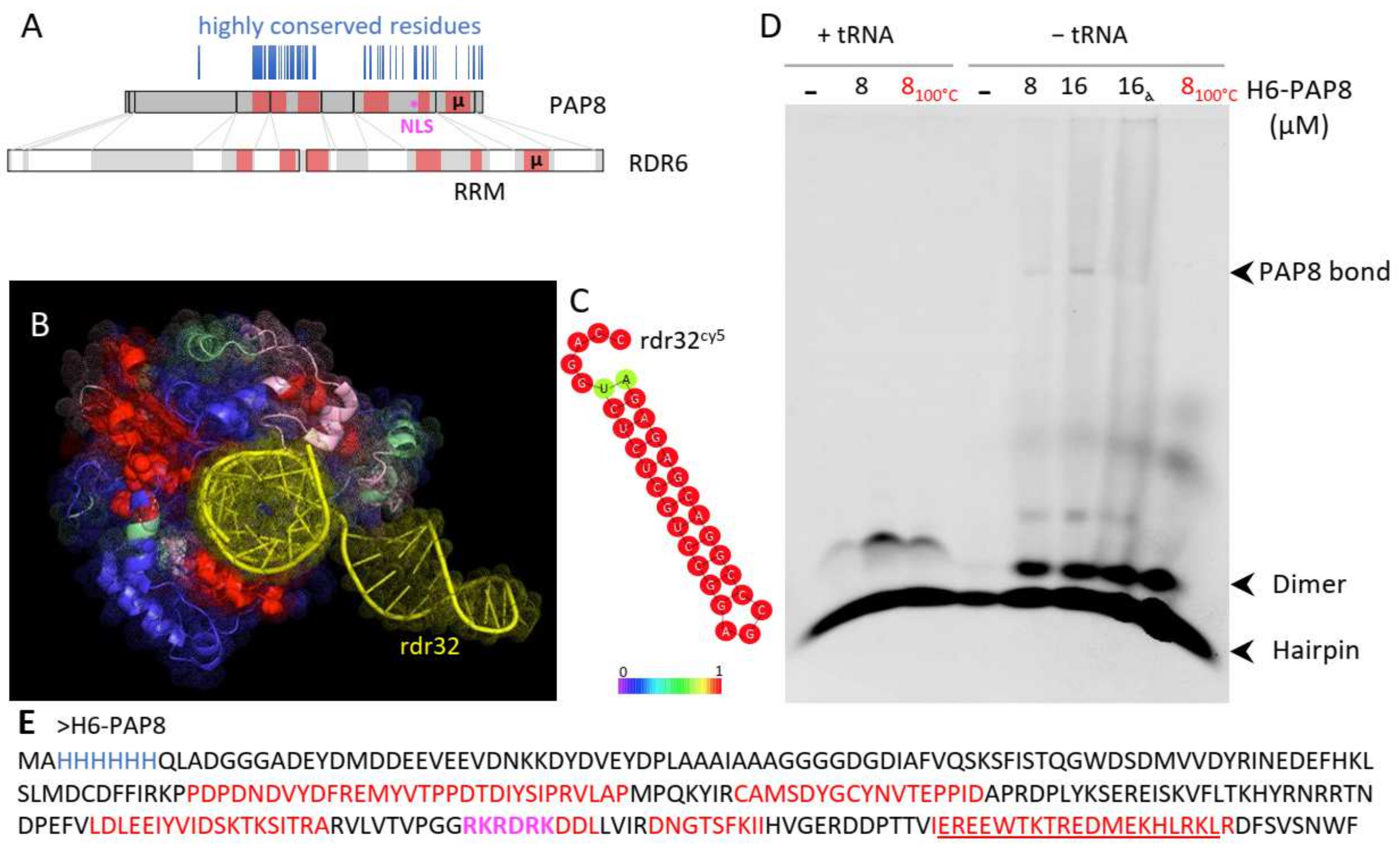
2.4. PAP8 Microhomology to RDR6 Reveals RNA Binding Motifs in PAP8
3. Discussion
4. Materials and Methods
4.1. Accessions
4.2. Live Material and Growth Conditions
4.3. Biochemical Purifications
4.4. Cloning
4.5. Transformation
4.6. Transgenics Characterization
4.7. Statistical Analysis
4.8. Protein Sub-Cellular Localization
4.9. rEMSA
4.10. Protein Production
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sato, S.; Nakamura, Y.; Kaneko, T.; Asamizu, E.; Tabata, S. Complete structure of the chloroplast genome of Arabidopsis thaliana. DNA Res. 1999, 6, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, P.; López-Juez, E. Biogenesis and homeostasis of chloroplasts and other plastids. Nat. Rev. Mol. Cell Biol. 2013, 14, 787–802, Erratum in Nat. Rev. Mol. Cell Biol. 2014, 15, 147. [Google Scholar] [CrossRef] [PubMed]
- Puthiyaveetil, S.; McKenzie, S.D.; Kayanja, G.E.; Ibrahim, I.M. Transcription initiation as a control point in plastid gene expression. Biochim. Biophys. Acta Gene Regul. Mech. 2021, 1864, 194689. [Google Scholar] [CrossRef] [PubMed]
- Kessler, F.; Schnell, D. Chloroplast biogenesis: Diversity and regulation of the protein import apparatus. Curr. Opin. Cell Biol. 2009, 21, 494–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Verdeja, T.; Vuorijoki, L.; Strand, Å. Emerging from the darkness: Interplay between light and plastid signaling during chloroplast biogenesis. Physiol. Plant 2020, 169, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Liebers, M.; Grübler, B.; Chevalier, F.; Lerbs-Mache, S.; Merendino, L.; Blanvillain, R.; Pfannschmidt, T. Regulatory Shifts in Plastid Transcription Play a Key Role in Morphological Conversions of Plastids during Plant Development. Front. Plant Sci. 2017, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Steiner, S.; Schröter, Y.; Pfalz, J.; Pfannschmidt, T. Identification of essential subunits in the plastid-encoded RNA polymerase complex reveals building blocks for proper plastid development. Plant Physiol. 2011, 157, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Pfalz, J.; Liere, K.; Kandlbinder, A.; Dietz, K.J.; Oelmüller, R. pTAC2, -6, and -12 are components of the transcriptionally active plastid chromosome that are required for plastid gene expression. Plant Cell 2006, 18, 176–197. [Google Scholar] [CrossRef] [Green Version]
- Liebers, M.; Chevalier, F.; Blanvillain, R.; Pfannschmidt, T. PAP genes are tissue- and cell-specific markers of chloroplast development. Planta 2018, 248, 629–646. [Google Scholar] [CrossRef]
- Ji, Y.; Lehotai, N.; Zan, Y.; Dubreuil, C.; Díaz, M.G.; Strand, Å. A fully assembled plastid-encoded RNA polymerase complex detected in etioplasts and proplastids in Arabidopsis. Physiol. Plant. 2021, 171, 435–446. [Google Scholar] [CrossRef]
- Kindgren, P.; Strand, Å. Chloroplast transcription, untangling the Gordian Knot. New Phytol. 2015, 206, 889–891. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Blanvillain, R.; Merendino, L.; Courtois, F.; Chevalier, F.; Liebers, M.; Grübler, B.; Hommel, E.; Lerbs-Mache, S. Plastid RNA polymerases: Orchestration of enzymes with different evolutionary origins controls chloroplast biogenesis during the plant life cycle. J. Exp. Bot. 2015, 66, 6957–6973. [Google Scholar] [CrossRef]
- Yagi, Y.; Ishizaki, Y.; Nakahira, Y.; Tozawa, Y.; Shiina, T. Eukaryotic-type plastid nucleoid protein pTAC3 is essential for transcription by the bacterial-type plastid RNA polymerase. Proc. Natl. Acad. Sci. USA 2012, 109, 7541–7546. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.H.; Lee, S.; Um, T.Y.; Kim, J.K.; Do Choi, Y.; Jang, G. pTAC10, a key subunit of plastid-encoded RNA polymerase, promotes chloroplast development. Plant Physiol. 2017, 174, 435–449. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Galvão, R.M.; Li, M.; Burger, B.; Bugea, J.; Bolado, J.; Chory, J. Arabidopsis HEMERA/pTAC12 initiates photomorphogenesis by phytochromes. Cell 2010, 141, 1230–1240. [Google Scholar] [CrossRef] [Green Version]
- Galvão, R.M.; Li, M.; Kothadia, S.M.; Haskel, J.D.; Decker, P.V.; Van Buskirk, E.K.; Chen, M. Photoactivated phytochromes interact with HEMERA and promote its accumulation to establish photomorphogenesis in Arabidopsis. Genes Dev. 2012, 26, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.P.; Yu, Q.B.; Zhao, T.T.; Ma, Q.; Chen, G.X.; Yang, Z.N. A functional component of the transcriptionally active chromosome complex, Arabidopsis pTAC14, interacts with pTAC12/HEMERA and regulates plastid gene expression. Plant Physiol. 2011, 157, 1733–1745. [Google Scholar] [CrossRef] [Green Version]
- Grübler, B.; Merendino, L.; Twardziok, S.O.; Mininno, M.; Allorent, G.; Chevalier, F.; Liebers, M.; Blanvillain, R.; Mayer, K.F.X.; Lerbs-Mache, S.; et al. Light and Plastid Signals Regulate Different Sets of Genes in the Albino Mutant Pap7-1. Plant Physiol. 2017, 175, 1203–1219. [Google Scholar] [CrossRef] [Green Version]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Ustün, S.; Melzer, M.; Petersen, K.; Lein, W.; Börnke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: Evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Yu, Q.B.; Lv, R.H.; Yin, Q.Q.; Chen, G.Y.; Xu, L.; Yang, Z.N. The reduced plastid-encoded polymerase-dependent plastid gene expression leads to the delayed greening of the Arabidopsis fln2 mutant. PLoS ONE 2013, 8, e73092. [Google Scholar] [CrossRef] [Green Version]
- Myouga, F.; Hosoda, C.; Umezawa, T.; Iizumi, H.; Kuromori, T.; Motohashi, R.; Shono, Y.; Nagata, N.; Ikeuchi, M.; Shinozaki, K. A heterocomplex of iron superoxide dismutases defends chloroplast nucleoids against oxidative stress and is essential for chloroplast development in Arabidopsis. Plant Cell 2008, 20, 3148–3162. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Myouga, F.; Takechi, K.; Sato, H.; Nabeshima, K.; Nagata, N.; Takio, S.; Shinozaki, K.; Takano, H. An Arabidopsis homolog of the bacterial peptidoglycan synthesis enzyme MurE has an essential role in chloroplast development. Plant J. 2008, 53, 924–934. [Google Scholar] [CrossRef]
- Yu, Q.B.; Lu, Y.; Ma, Q.; Zhao, T.T.; Huang, C.; Zhao, H.F.; Zhang, X.L.; Lv, R.H.; Yang, Z.N. TAC7, an essential component of the plastid transcriptionally active chromosome complex, interacts with FLN1, TAC10, TAC12 and TAC14 to regulate chloroplast gene expression in Arabidopsis thaliana. Physiol. Plant. 2013, 148, 408–421. [Google Scholar] [CrossRef]
- Ding, S.; Zhang, Y.; Hu, Z.; Huang, X.; Zhang, B.; Lu, Q.; Wen, X.; Wang, Y.; Lu, C. mTERF5 Acts as a Transcriptional Pausing Factor to Positively Regulate Transcription of Chloroplast psbEFL. J. Mol. Plant 2019, 12, 1259–1277. [Google Scholar] [CrossRef]
- Tadini, L.; Pesaresi, P.; Kleine, T.; Rossi, F.; Guljamow, A.; Sommer, F.; Mühlhaus, T.; Schroda, M.; Masiero, S.; Pribil, M.; et al. GUN1 Controls Accumulation of the Plastid Ribosomal Protein S1 at the Protein Level and Interacts with Proteins Involved in Plastid Protein Homeostasis. Plant Physiol. 2016, 170, 1817–1830. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Tang, R.; Shi, Y.; Ke, X.; Wang, Y.; Che, Y.; Luan, S.; Hou, X. Arabidopsis Seedling Lethal 1 Interacting with Plastid-Encoded RNA Polymerase Complex Proteins Is Essential for Chloroplast Development. Front. Plant Sci. 2020, 11, 602782. [Google Scholar] [CrossRef]
- Liebers, M.; Gillet, F.X.; Israel, A.; Pounot, K.; Chambon, L.; Chieb, M.; Chevalier, F.; Ruedas, R.; Favier, A.; Gans, P.; et al. Nucleo-plastidic PAP8/pTAC6 couples chloroplast formation with photomorphogenesis. EMBO J. 2020, 39, e104941. [Google Scholar] [CrossRef]
- Krause, K.; Oetke, S.; Krupinska, K. Dual targeting and retrograde translocation: Regulators of plant nuclear gene expression can be sequestered by plastids. Int. J. Mol. Sci. 2012, 13, 11085–11101. [Google Scholar] [CrossRef] [Green Version]
- Krupinska, K.; Blanco, N.E.; Oetke, S.; Zottini, M. Genome communication in plants mediated by organelle-nucleus-located proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190397. [Google Scholar] [CrossRef]
- Powikrowska, M.; Oetke, S.; Jensen, P.E.; Krupinska, K. Dynamic composition, shaping and organization of plastid nucleoids. Front. Plant Sci. 2014, 5, 424. [Google Scholar] [CrossRef]
- Ingelsson, B.; Vener, A.V. Phosphoproteomics of Arabidopsis chloroplasts reveals involvement of the STN7 kinase in phosphorylation of nucleoid protein pTAC16. FEBS Lett. 2012, 586, 1265–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, P.; Kortus, M.G.; Nix, J.C.; Davis, R.E.; Peersen, O.B. Structures of coxsackievirus, rhinovirus, and poliovirus polymerase elongation complexes solved by engineering RNA mediated crystal contacts. PLoS ONE 2013, 8, e60272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevarez, P.A.; Qiu, Y.; Inoue, H.; Yoo, C.Y.; Benfey, P.N.; Schnell, D.J.; Chen, M. Mechanism of Dual Targeting of the Phytochrome Signaling Component HEMERA/pTAC12 to Plastids and the Nucleus. Plant Physiol. 2017, 173, 1953–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.J.; Yoo, C.Y.; Liu, J.; Wang, H.; Cao, J.; Li, F.W.; Pryer, K.M.; Sun, T.P.; Weigel, D.; Zhou, P.; et al. NCP activates chloroplast transcription by controlling phytochrome-dependent dual nuclear and plastidial switches. Nat. Commun. 2019, 10, 2630. [Google Scholar] [CrossRef] [Green Version]
- Yoo, C.Y.; Pasoreck, E.K.; Wang, H.; Cao, J.; Blaha, G.M.; Weigel, D.; Chen, M. Phytochrome activates the plastid-encoded RNA polymerase for chloroplast biogenesis via nucleus-to-plastid signaling. Nat. Commun. 2019, 10, 2629. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Li, M.; Pasoreck, E.K.; Long, L.; Shi, Y.; Galvao, R.M.; Chou, C.L.; Wang, H.; Sun, A.Y.; Zhang, Y.C.; et al. HEMERA Couples the Proteolysis and Transcriptional Activity of Phytochrome Interacting Factors in Arabidopsis Photomorphogenesis. Plant Cell 2015, 27, 1409–1427. [Google Scholar] [CrossRef] [Green Version]
- Favier, A.; Gans, P.; Boeri Erba, E.; Signor, L.; Muthukumar, S.S.; Pfannschmidt, T.; Blanvillain, R.; Cobessi, D. The Plastid-Encoded RNA Polymerase-Associated Protein PAP9 Is a Superoxide Dismutase with Unusual Structural Features. Front. Plant Sci. 2021, 12, 668897. [Google Scholar] [CrossRef]
- de Vries, J.; Stanton, A.; Archibald, J.M.; Gould, S.B. Streptophyte Terrestrialization in Light of Plastid Evolution. Trends Plant Sci. 2016, 21, 467–476. [Google Scholar] [CrossRef]
- Bock, R.; Timmis, J.N. Reconstructing evolution: Gene transfer from plastids to the nucleus. Bioessays 2008, 30, 556–566. [Google Scholar] [CrossRef] [Green Version]
- Chieb, M.; Liebers, M.; Chevalier, F.; Lerbs-Mache, S.; Blanvillain, R.; Pfannschmidt, T. Determination of the DNA/RNA-Associated Subproteome from Chloroplasts and Other Plastid Types. Methods Mol. Biol. 2018, 1829, 253–271. [Google Scholar] [CrossRef]
- Blanvillain, R.; Wei, S.; Wei, P.; Kim, J.H.; Ow, D.W. Stress tolerance to stress escape in plants: Role of the OXS2 zinc-finger transcription factor family. EMBO J. 2011, 30, 3812–3822. [Google Scholar] [CrossRef] [Green Version]
- Bensmihen, S.; To, A.; Lambert, G.; Kroj, T.; Giraudat, J.; Parcy, F. Analysis of an activated ABI5 allele using a new selection method for transgenic Arabidopsis seeds. FEBS Lett. 2004, 561, 127–131. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chambon, L.; Gillet, F.-X.; Chieb, M.; Cobessi, D.; Pfannschmidt, T.; Blanvillain, R. PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin. Int. J. Mol. Sci. 2022, 23, 3059. https://doi.org/10.3390/ijms23063059
Chambon L, Gillet F-X, Chieb M, Cobessi D, Pfannschmidt T, Blanvillain R. PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin. International Journal of Molecular Sciences. 2022; 23(6):3059. https://doi.org/10.3390/ijms23063059
Chicago/Turabian StyleChambon, Louise, François-Xavier Gillet, Maha Chieb, David Cobessi, Thomas Pfannschmidt, and Robert Blanvillain. 2022. "PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin" International Journal of Molecular Sciences 23, no. 6: 3059. https://doi.org/10.3390/ijms23063059
APA StyleChambon, L., Gillet, F.-X., Chieb, M., Cobessi, D., Pfannschmidt, T., & Blanvillain, R. (2022). PAP8/pTAC6 Is Part of a Nuclear Protein Complex and Displays RNA Recognition Motifs of Viral Origin. International Journal of Molecular Sciences, 23(6), 3059. https://doi.org/10.3390/ijms23063059