
Expression of SLC26A9 in Airways and Its Potential Role in Asthma


Abstract
:1. Introduction
2. Results
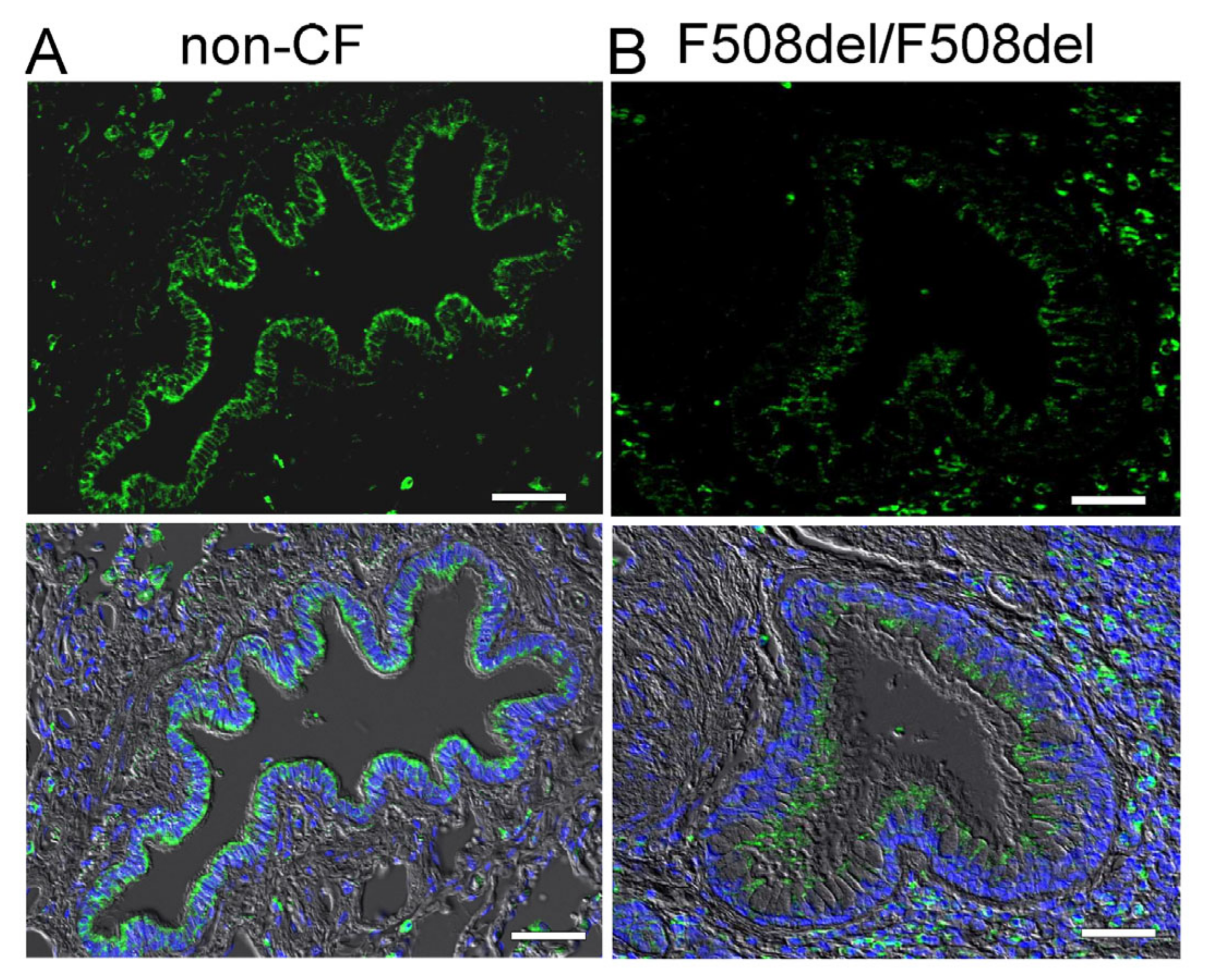
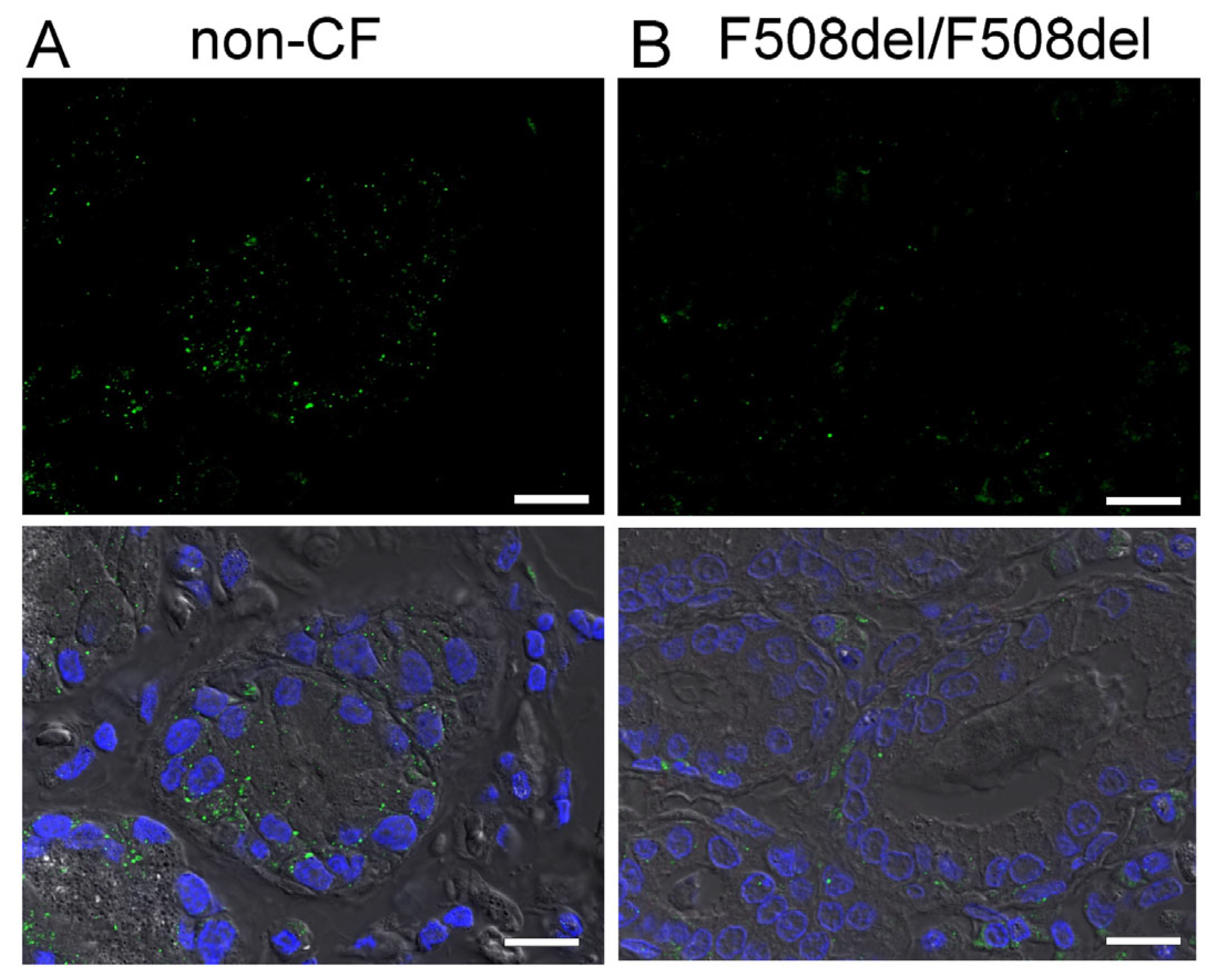
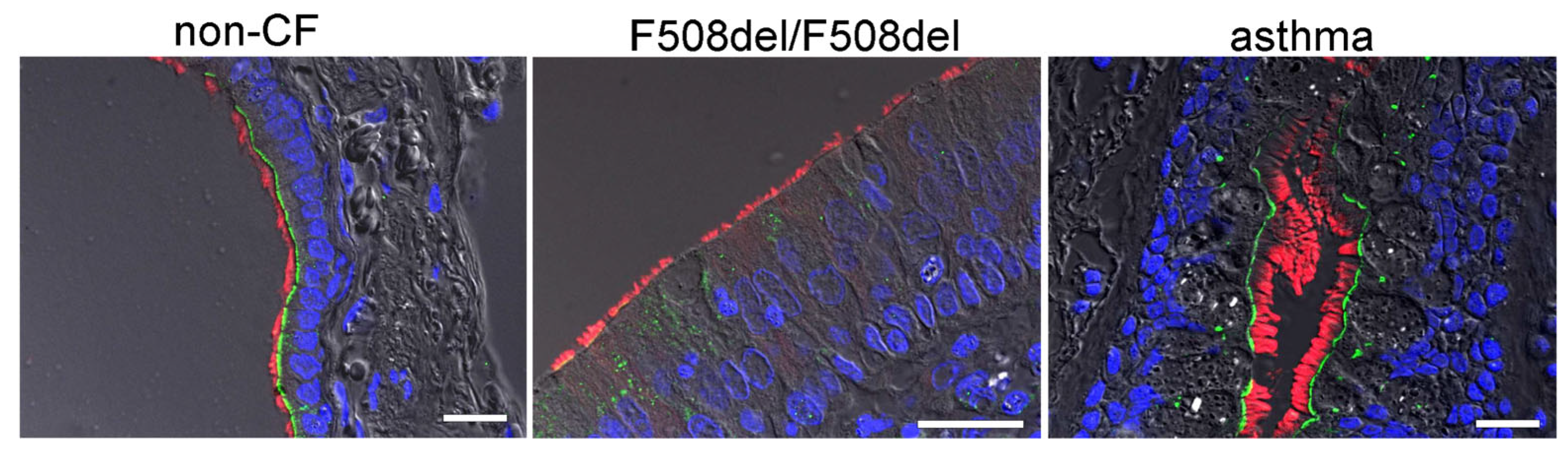
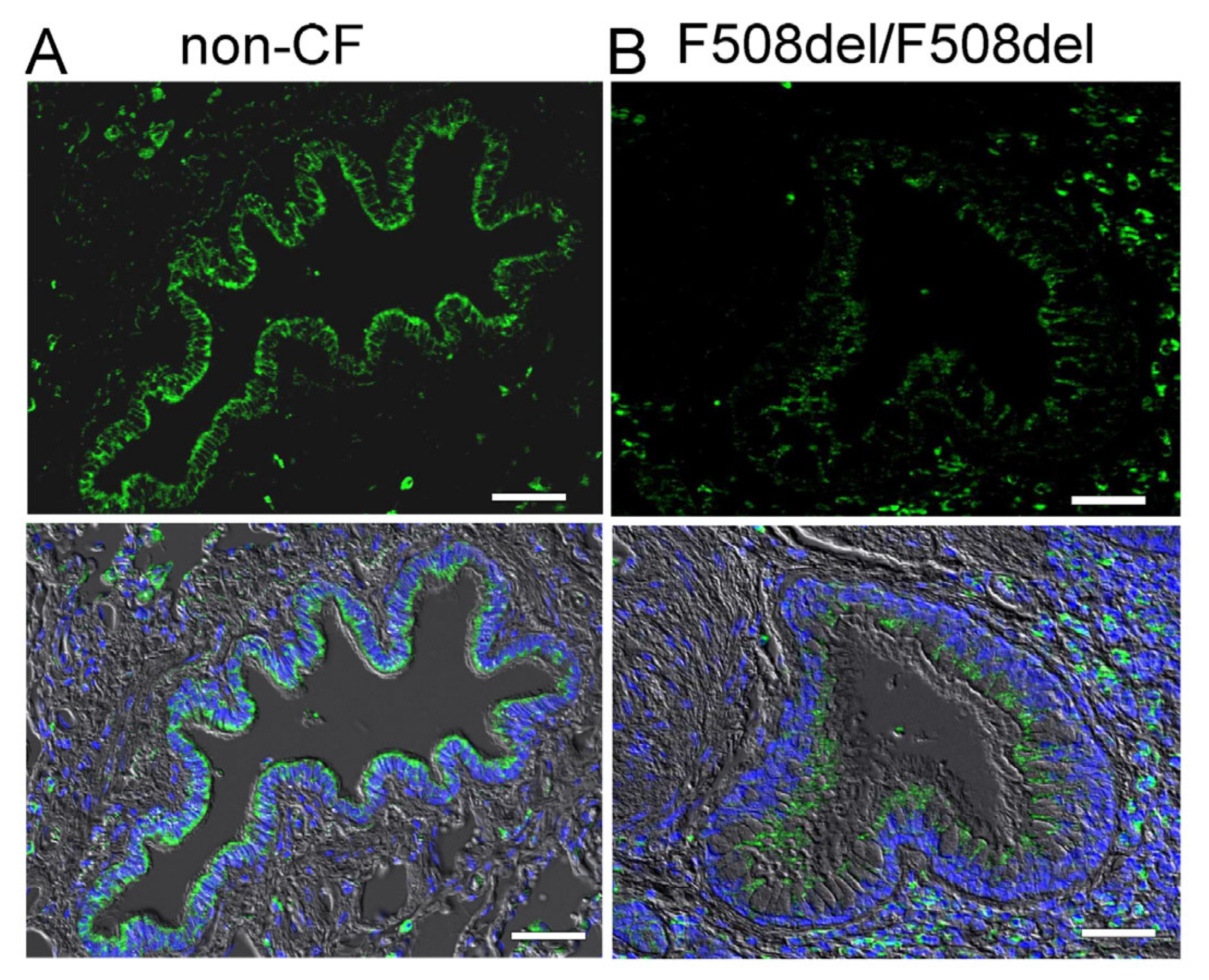
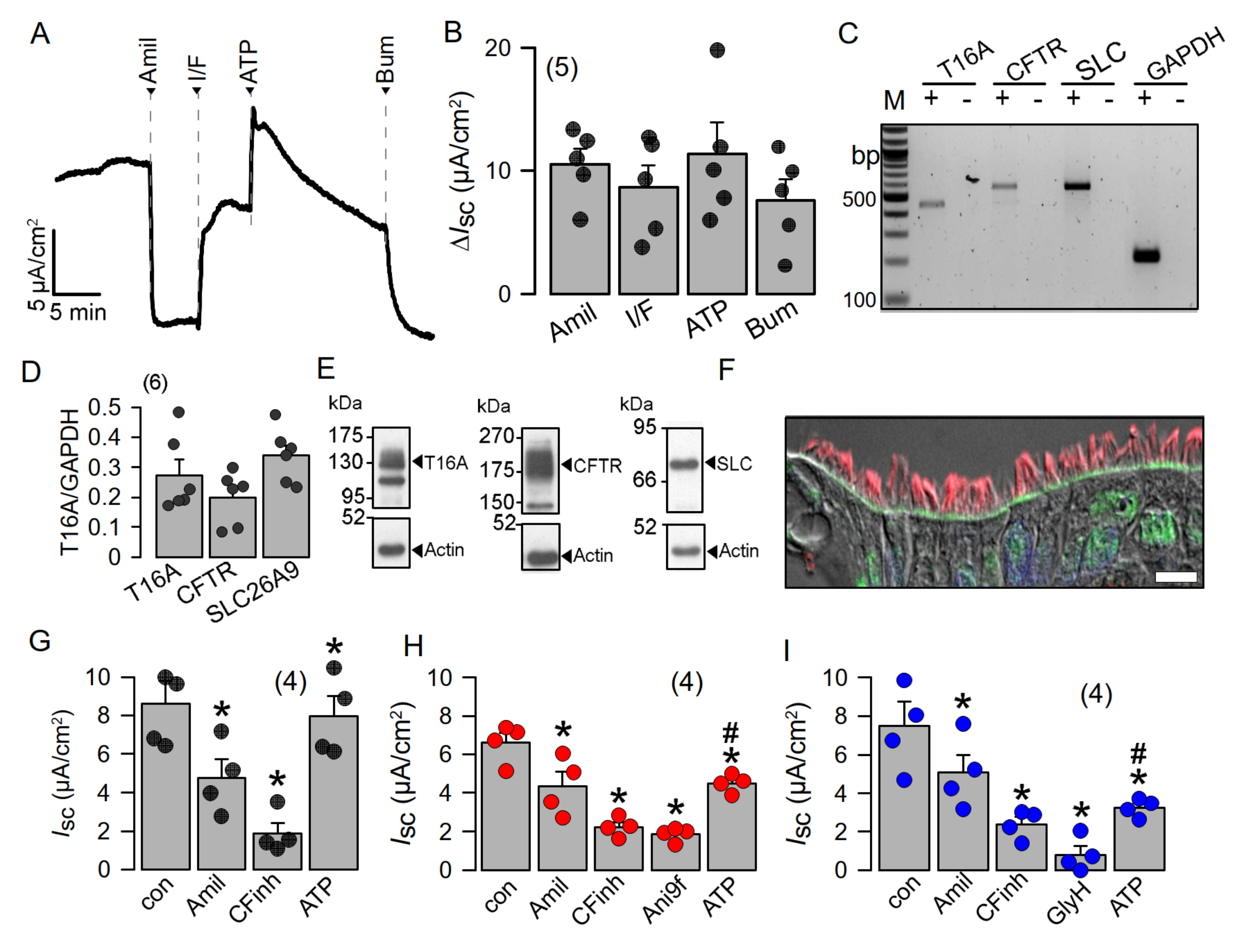
2.1. Apical Expression of SLC26A9 in Human Superficial Airway Epithelium Which Is Absent in CF
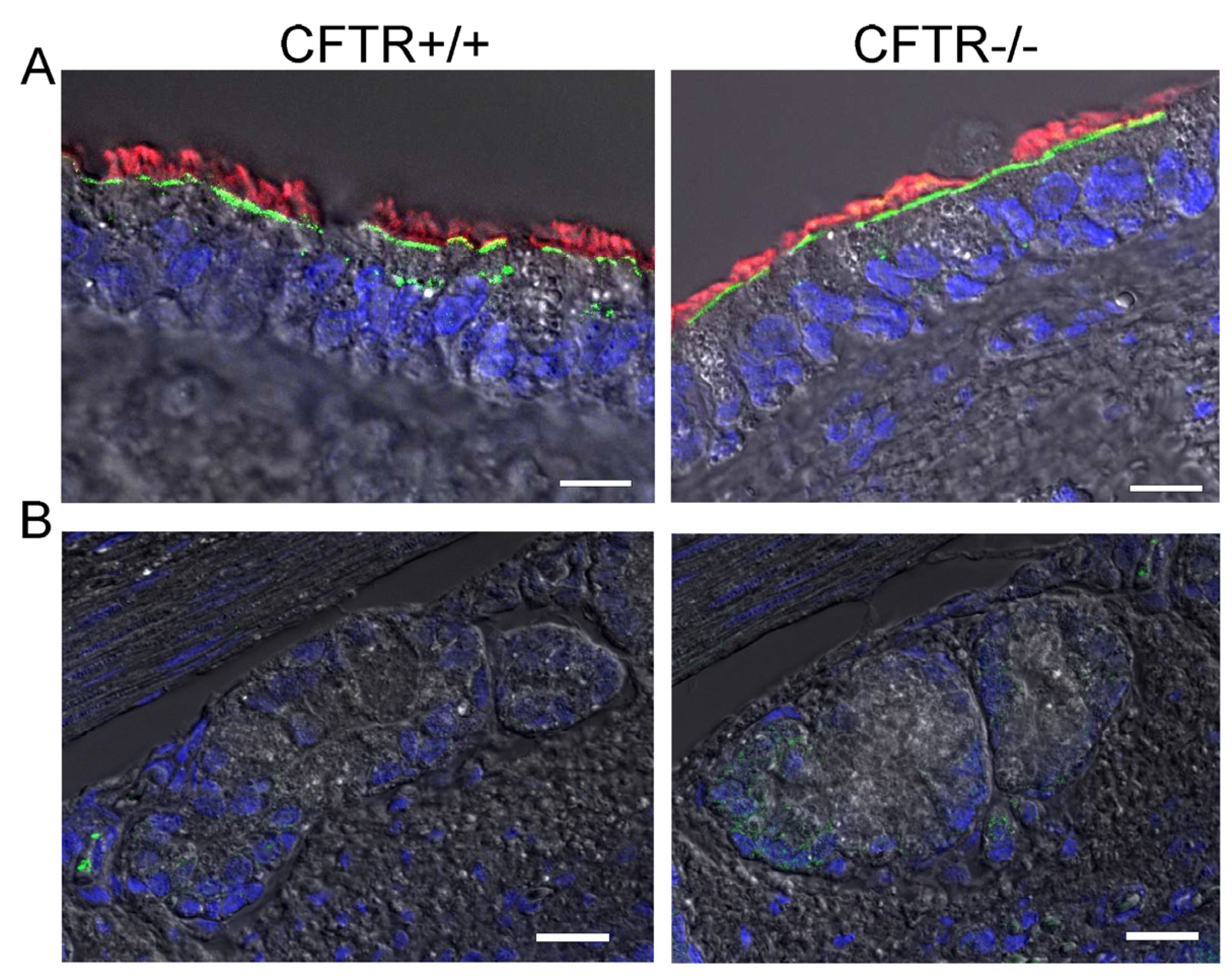
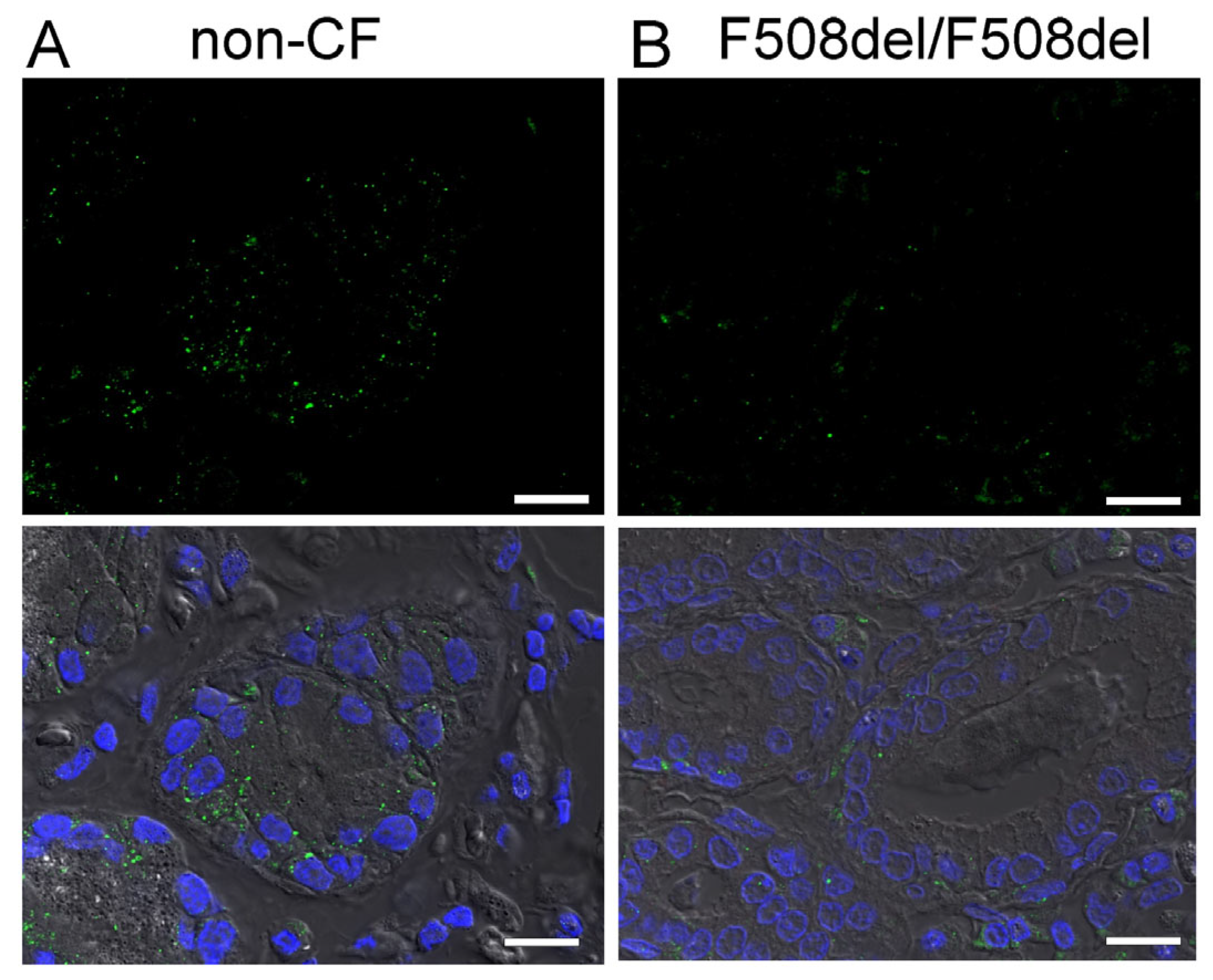
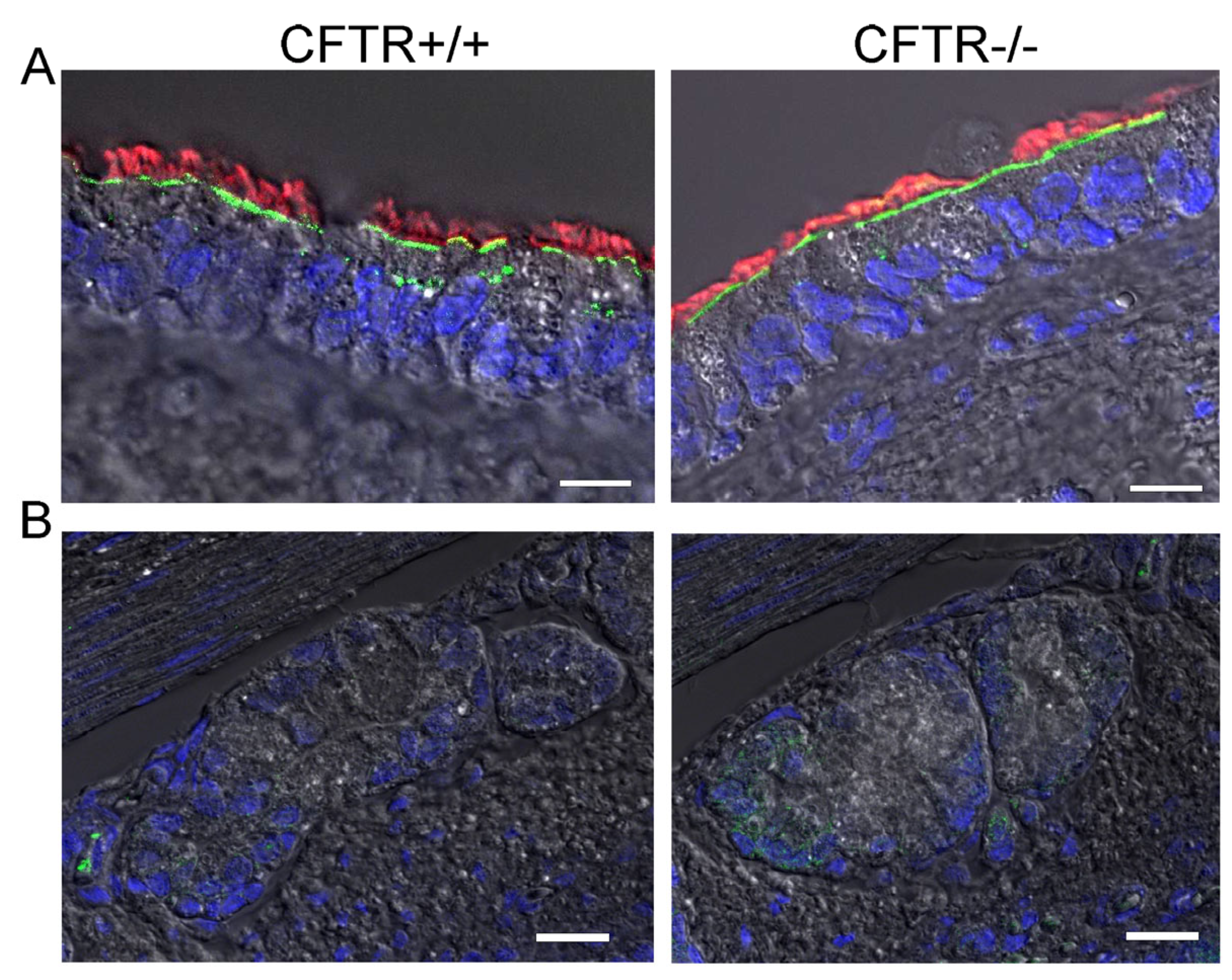
2.2. SLC26A9 Is Located in the Apical Membrane of Ciliated Airway Epithelial Cells of CFTR+/+ and CFTR−/− Piglet Lungs
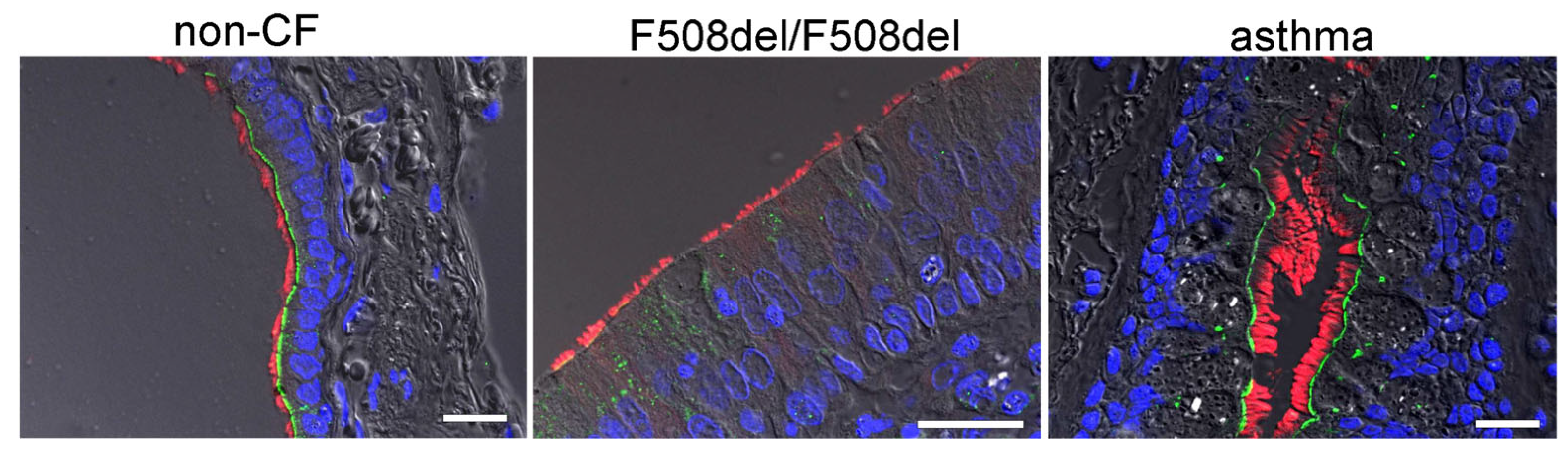
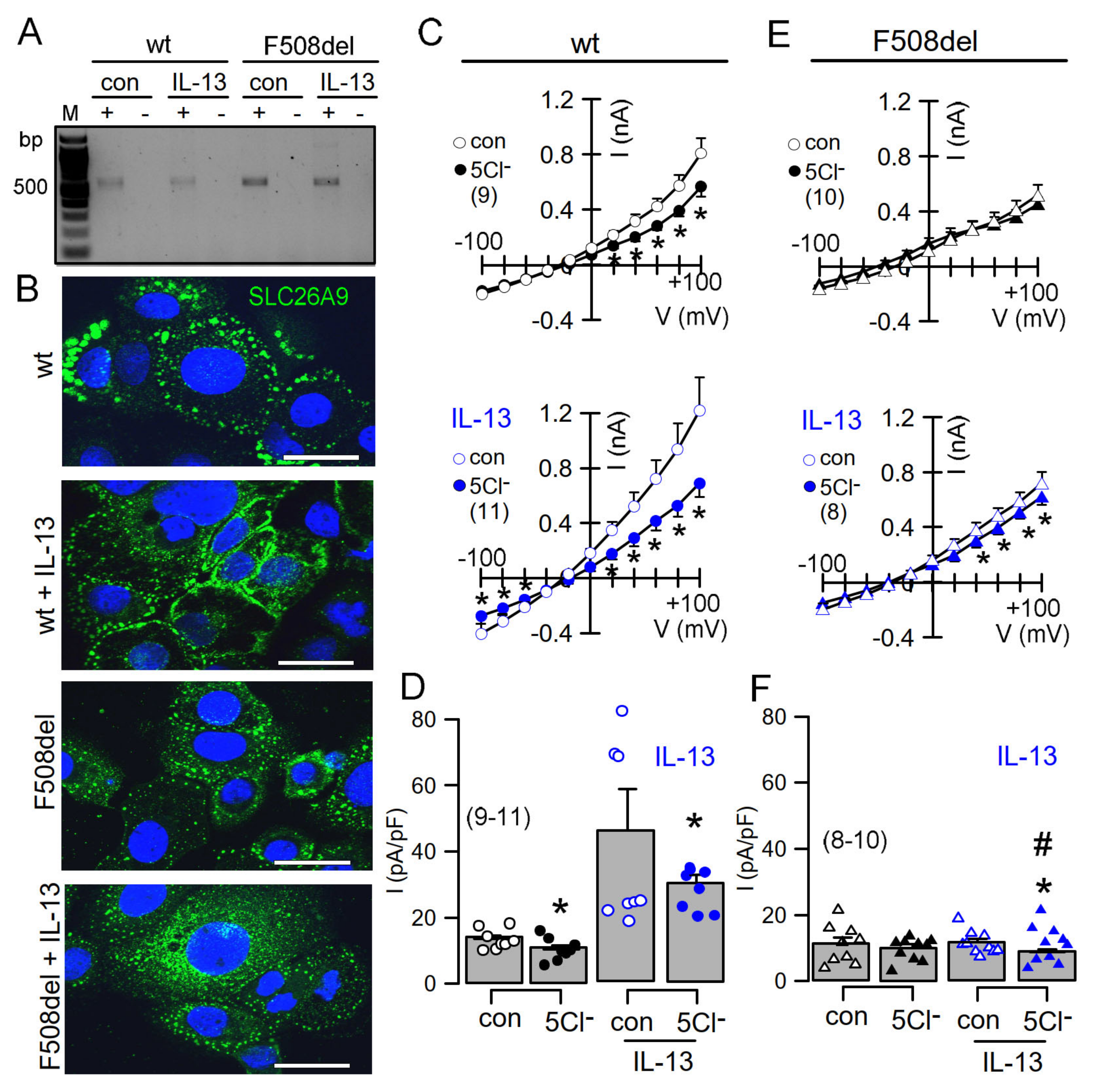
2.3. IL-13 Augments Membrane Expression of SLC26A9 and Induces Basal Cl− Currents in CFBE Airway Epithelial Cells Expressing wtCFTR, but Not in Cells Expressing F508del-CFTR
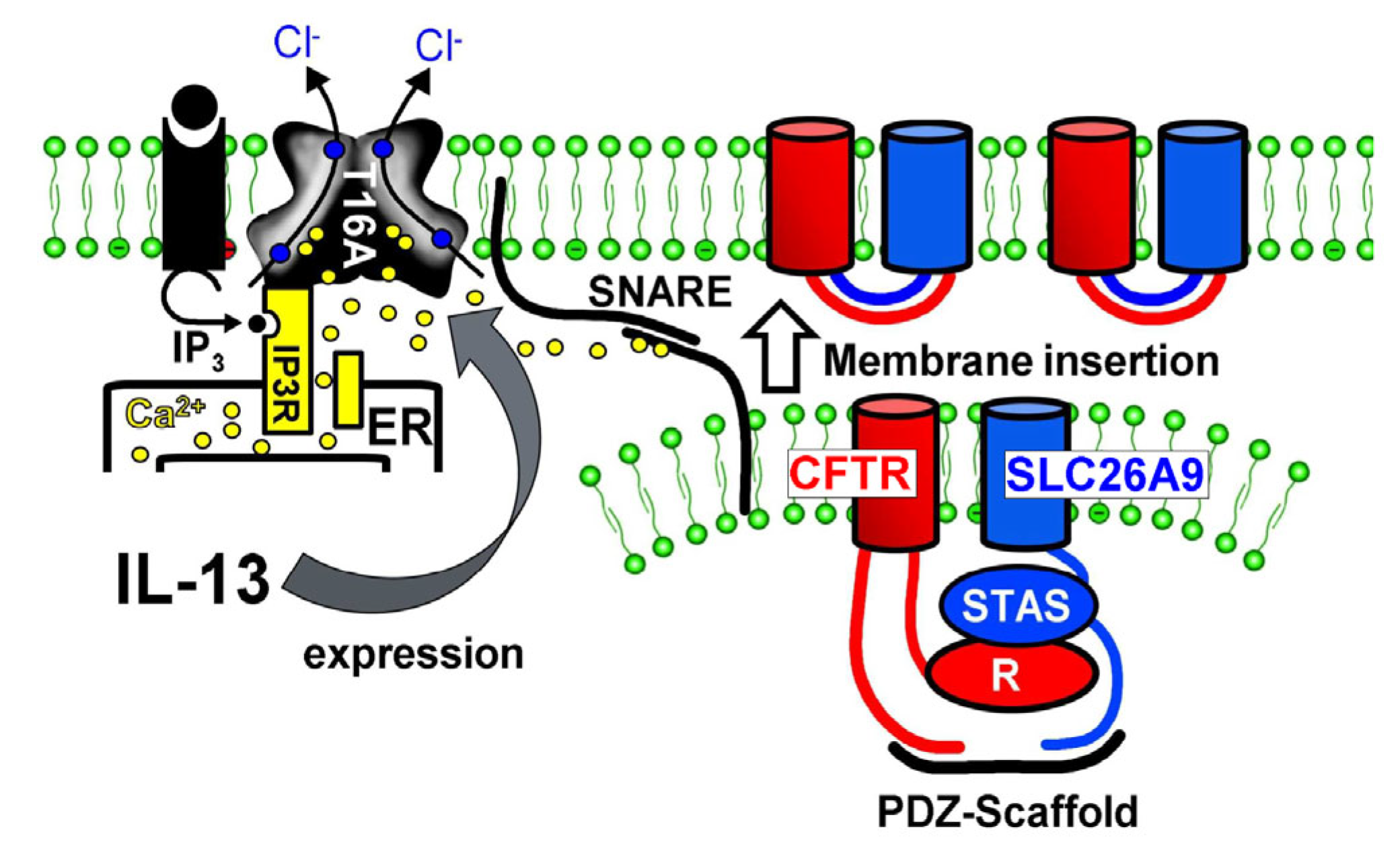
3. Discussion
3.1. SLC26A9 Is Expressed in the Apical Membrane of the Airway Epithelium
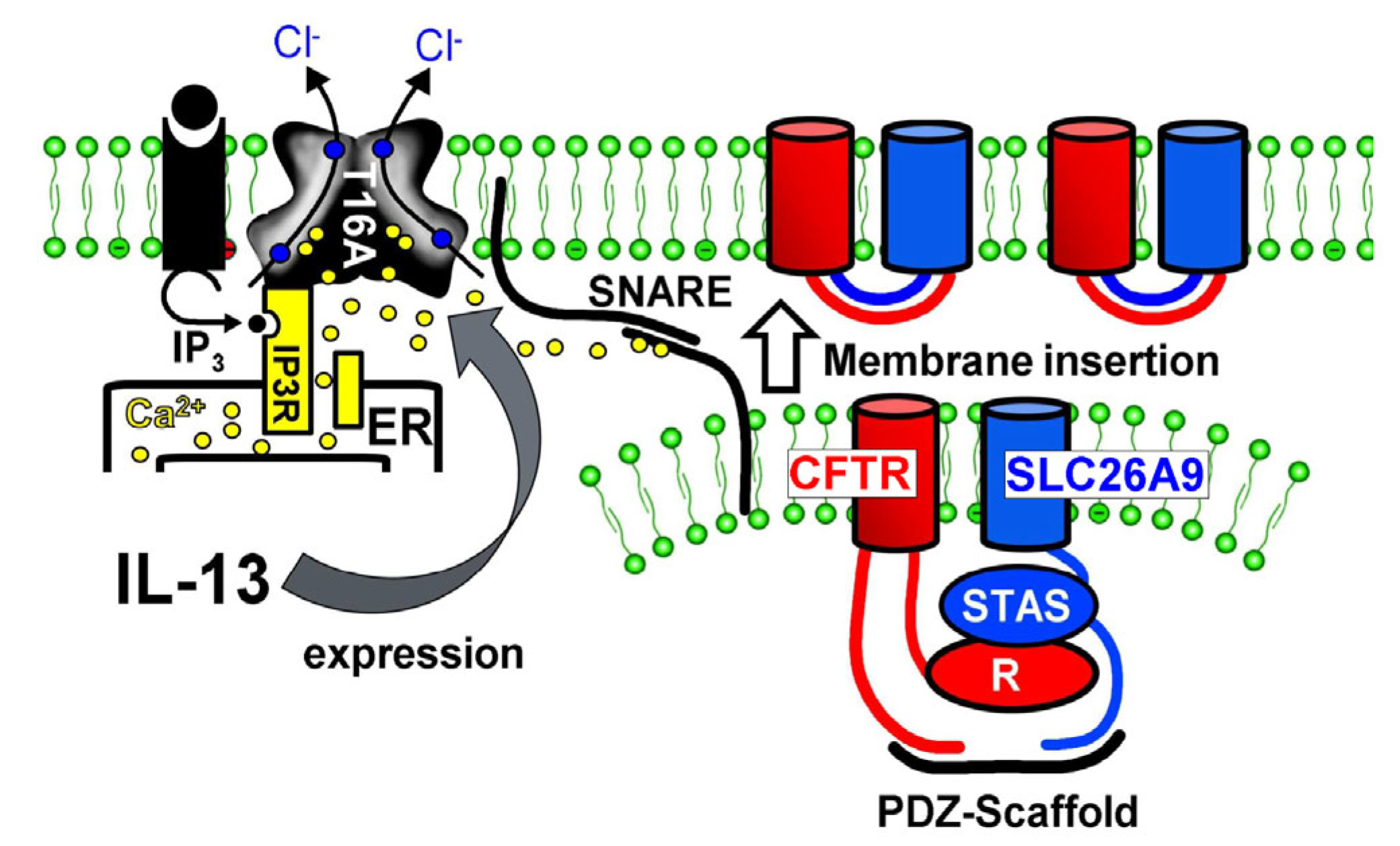
3.2. Plasma Membrane Expression of SLC26A9 Is CFTR-Dependent and Is Augmented by IL-13
4. Materials and Methods
4.1. Tissues, Cell Culture and Treatment
4.2. RT-PCR
4.3. Western Blot
4.4. Immunocytochemistry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alper, S.L.; Sharma, A.K. The SLC26 gene family of anion transporters and channels. Mol. Aspects Med. 2013, 34, 494–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amlal, H.; Xu, J.; Barone, S.; Zahedi, K.; Soleimani, M. The chloride channel/transporter Slc26a9 regulates the systemic arterial pressure and renal chloride excretion. J. Mol. Med. 2013, 91, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Anagnostopoulou, P.; Riederer, B.; Duerr, J.; Michel, S.; Binia, A.; Agrawal, R.; Liu, X.; Kalitzki, K.; Xiao, F.; Chen, M.; et al. SLC26A9-mediated chloride secretion prevents mucus obstruction in airway inflammation. J. Clin. Investig. 2012, 122, 3629–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebok, Z.; Collawn, J.F.; Wakefield, J.; Parker, W.; Li, Y.; Varga, K.; Sorscher, E.J.; Clancy, J.P. Failure of cAMP agonists to activate rescued deltaF508 CFTR in CFBE41o- airway epithelial monolayers. J. Physiol. 2005, 569, 601–615. [Google Scholar] [CrossRef]
- Benedetto, R.; Centeio, R.; Ousingsawat, J.; Schreiber, R.; Janda, M.; Kunzelmann, K. Transport properties in CFTR−/− knockout piglets suggest normal airway surface liquid pH and enhanced amiloride-sensitive Na(+) absorption. Pflugers Arch. 2020, 472, 1507–1519. [Google Scholar] [CrossRef]
- Benedetto, R.; Ousingsawat, J.; Cabrita, I.; Pinto, M.; Lerias, J.; Wanitchakool, P.; Schreiber, R.; Kunzelmann, K. Plasma membrane localized TMEM16 Proteins are Indispensable for expression of CFTR. J. Mol. Med. 2019, 97, 711–722. [Google Scholar] [CrossRef]
- Benedetto, R.; Ousingsawat, J.; Wanitchakool, P.; Zhang, Y.; Holtzman, M.J.; Amaral, M.; Rock, J.R.; Schreiber, R.; Kunzelmann, K. Epithelial Chloride Transport by CFTR Requires TMEM16A. Sci. Rep. 2017, 7, 12397. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, C.A.; Mitra, S.; Mishra, S.K.; Wang, X.; Zhao, Y.; Pilewski, J.M.; Madden, D.R.; Frizzell, R.A. The CFTR trafficking mutation F508del inhibits the constitutive activity of SLC26A9. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 312, L912–L925. [Google Scholar] [CrossRef]
- Bertrand, C.A.; Zhang, R.; Pilewski, J.M.; Frizzell, R.A. SLC26A9 is a constitutively active, CFTR-regulated anion conductance in human bronchial epithelia. J. Gen. Physiol. 2009, 133, 421–438. [Google Scholar] [CrossRef] [Green Version]
- Botelho, H.M.; Uliyakina, I.; Awatade, N.T.; Proenca, M.C.; Tischer, C.; Sirianant, L.; Kunzelmann, K.; Pepperkok, R.; Amaral, M.D. Protein traffic disorders: An effective high-throughput fluorescence microscopy pipeline for drug discovery. Sci. Rep. 2015, 5, 9038. [Google Scholar] [CrossRef] [Green Version]
- Cabrita, I.; Benedetto, R.; Wanitchakool, P.; Lerias, J.; Centeio, R.; Ousingsawat, J.; Schreiber, R.; Kunzelmann, K. TMEM16A Mediated Mucus Production in Human Airway Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2020, 64, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Centeio, R.; Ousingsawat, J.; Schreiber, R.; Kunzelmann, K. CLCA1 Regulates Airway Mucus Production and Ion Secretion Through TMEM16A. Int. J. Mol. Sci. 2021, 22, 5133. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Plata, C.; Zandi-Nejad, K.; Sindic, A.; Sussman, C.R.; Mercado, A.; Broumand, V.; Raghuram, V.; Mount, D.B.; Romero, M.F. Slc26a9-Anion Exchanger, Channel and Na(+) Transporter. J. Membr. Biol. 2009, 228, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Dallos, P.; Fakler, B. Prestin, a new type of motor protein. Nat. Rev. Mol. Cell Biol. 2002, 3, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Dorwart, M.R.; Shcheynikov, N.; Wang, Y.; Stippec, S.; Muallem, S. SLC26A9 is a Cl(-) channel regulated by the WNK kinases. J. Physiol. 2007, 584, 333–345. [Google Scholar] [CrossRef]
- El Khouri, E.; Toure, A. Functional interaction of the cystic fibrosis transmembrane conductance regulator with members of the SLC26 family of anion transporters (SLC26A8 and SLC26A9): Physiological and pathophysiological relevance. Int. J. Biochem. Cell Biol. 2014, 52, 58–67. [Google Scholar] [CrossRef]
- Klymiuk, N.; Mundhenk, L.; Kraehe, K.; Wuensch, A.; Plog, S.; Emrich, D.; Langenmayer, M.C.; Stehr, M.; Holzinger, A.; Kroner, C.; et al. Sequential targeting of CFTR by BAC vectors generates a novel pig model of cystic fibrosis. J. Mol. Med. 2012, 90, 597–608. [Google Scholar] [CrossRef]
- Ko, S.B.; Shcheynikov, N.; Choi, J.Y.; Luo, X.; Ishibashi, K.; Thomas, P.J.; Kim, J.Y.; Kim, K.H.; Lee, M.G.; Naruse, S.; et al. A molecular mechanism for aberrant CFTR-dependent HCO(3)(-) transport in cystic fibrosis. EMBO J. 2002, 21, 5662–5672. [Google Scholar] [CrossRef]
- Ko, S.B.; Zeng, W.; Dorwart, M.R.; Luo, X.; Kim, K.H.; Millen, L.; Goto, H.; Naruse, S.; Soyombo, A.; Thomas, P.J.; et al. Gating of CFTR by the STAS domain of SLC26 transporters. Nat. Cell Biol. 2004, 6, 343–350. [Google Scholar] [CrossRef]
- Lerias, J.; Pinto, M.; Benedetto, R.; Schreiber, R.; Amaral, M.; Aureli, M.; Kunzelmann, K. Compartmentalized crosstalk of CFTR and TMEM16A (ANO1) through EPAC1 and ADCY1. Cell Signal. 2018, 44, 10–19. [Google Scholar] [CrossRef]
- Liu, X.; Li, T.; Riederer, B.; Lenzen, H.; Ludolph, L.; Yeruva, S.; Tuo, B.; Soleimani, M.; Seidler, U. Loss of Slc26a9 anion transporter alters intestinal electrolyte and HCO3(-) transport and reduces survival in CFTR-deficient mice. Pflugers Arch. 2015, 467, 1261–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loriol, C.; Dulong, S.; Avella, M.; Gabillat, N.; Boulukos, K.; Borgese, F.; Ehrenfeld, J. Characterization of SLC26A9, facilitation of Cl(-) transport by bicarbonate. Cell Physiol. Biochem. 2008, 22, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Needham, P.G.; Goeckeler-Fried, J.L.; Zhang, C.; Sun, Z.; Wetzel, A.R.; Bertrand, C.A.; Brodsky, J.L. SLC26A9 is selected for endoplasmic reticulum associated degradation (ERAD) via Hsp70-dependent targeting of the soluble STAS domain. Biochem. J. 2021, 47, 4203–4220. [Google Scholar] [CrossRef]
- Ohana, E.; Yang, D.; Shcheynikov, N.; Muallem, S. Diverse transport modes by the Solute Carrier 26 family of anion transporters. J. Physiol. 2008, 24, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Ousingsawat, J.; Schreiber, R.; Kunzelmann, K. Differential contribution of SLC26A9 to Cl(-) conductance in polarized and non-polarized epithelial cells. J. Cell Physiol. 2011, 227, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Ousingsawat, J.; Cabrita, I.; Bettels, R.E.; Große-Onnebrink, J.; Schmalstieg, C.; Biskup, S.; Reunert, J.; Rust, S.; Schreiber, R.; et al. TMEM16A deficiency: A potentially fatal neonatal disease resulting from impaired chloride currents. J. Med. Genet 2020, 58, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.V.N.; Ribeiro, J.D.; Bertuzzo, C.S.; Marson, F.A.L. Interaction among variants in the SLC gene family (SLC6A14, SLC26A9, SLC11A1, and SLC9A3) and CFTR mutations with clinical markers of cystic fibrosis. Pediatric Pulmonol. 2018, 53, 888–900. [Google Scholar] [CrossRef]
- Pinto, M.C.; Quaresma, M.C.; Silva, I.A.L.; Railean, V.; Ramalho, S.S.; Amaral, M.D. Synergy in Cystic Fibrosis Therapies: Targeting SLC26A9. Int. J. Mol. Sci. 2021, 22, 13064. [Google Scholar] [CrossRef]
- Quesada, R.; Dutzler, R. Alternative chloride transport pathways as pharmacological targets for the treatment of cystic fibrosis. J. Cyst. Fibrosis 2019, 19, S37–S41. [Google Scholar] [CrossRef] [Green Version]
- Rakonczay, Z., Jr.; Hegyi, P.; Hasegawa, M.; Inoue, M.; You, J.; Iida, A.; Ignáth, I.; Alton, E.W.; Griesenbach, U.; Ovári, G.; et al. CFTR gene transfer to human cystic fibrosis pancreatic duct cells using a Sendai virus vector. J. Cell Physiol. 2008, 214, 442–455. [Google Scholar] [CrossRef]
- Rode, B.; Dirami, T.; Bakouh, N.; Rizk-Rabin, M.; Norez, C.; Lhuillier, P.; Lorès, P.; Jollivet, M.; Melin, P.; Zvetkova, I.; et al. The testis anion transporter TAT1 (SLC26A8) physically and functionally interacts with the cystic fibrosis transmembrane conductance regulator channel: A potential role during sperm capacitation. Hum. Mol. Genet. 2012, 21, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Salomon, J.J.; Spahn, S.; Wang, X.; Fullekrug, J.; Bertrand, C.A.; Mall, M.A. Generation and functional characterization of epithelial cells with stable expression of SLC26A9 Cl- channels. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L593–L602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Thomas, D.Y.; Hanrahan, J.W. The anion transporter SLC26A9 localizes to tight junctions and is degraded by the proteasome when co-expressed with F508del-CFTR. J. Biol. Chem. 2019, 294, 18269–18284. [Google Scholar] [CrossRef] [PubMed]
- Seidler, U.; Nikolovska, K. Slc26 Family of Anion Transporters in the Gastrointestinal Tract: Expression, Function, Regulation, and Role in Disease. Comp. Physiol. 2019, 9, 839–872. [Google Scholar] [CrossRef]
- Seo, Y.; Kim, J.; Chang, J.; Kim, S.S.; Namkung, W.; Kim, I. Synthesis and biological evaluation of novel Ani9 derivatives as potent and selective ANO1 inhibitors. Eur. J. Med. Chem. 2018, 160, 245–255. [Google Scholar] [CrossRef]
- Strug, L.J.; Gonska, T.; He, G.; Keenan, K.; Ip, W.; Boelle, P.Y.; Lin, F.; Panjwani, N.; Gong, J.; Li, W.; et al. Cystic fibrosis gene modifier SLC26A9 modulates airway response to CFTR-directed therapeutics. Hum. Mol. Genet. 2016, 25, 4590–4600. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.D.; Sawicka, M.; Dutzler, R. Cryo-EM structures and functional characterization of murine Slc26a9 reveal mechanism of uncoupled chloride transport. eLife 2019, 8, e46986. [Google Scholar] [CrossRef]
- Walters, M.S.; Gomi, K.; Ashbridge, B.; Moore, M.A.; Arbelaez, V.; Heldrich, J.; Ding, B.S.; Rafii, S.; Staudt, M.R.; Crystal, R.G. Generation of a human airway epithelium derived basal cell line with multipotent differentiation capacity. Respir. Res. 2013, 14, 135. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Henriksnäs, J.; Barone, S.; Witte, D.; Shull, G.E.; Forte, J.G.; Holm, L.; Soleimani, M. SLC26A9 is expressed in gastric surface epithelial cells, mediates Cl-/HCO3- exchange, and is inhibited by NH4+. Am. J. Physiol. Cell Physiol. 2005, 289, C493–C505. [Google Scholar] [CrossRef]
- Xu, J.; Song, P.; Miller, M.L.; Borgese, F.; Barone, S.; Riederer, B.; Wang, Z.; Alper, S.L.; Forte, J.G.; Shull, G.E.; et al. Deletion of the chloride transporter Slc26a9 causes loss of tubulovesicles in parietal cells and impairs acid secretion in the stomach. Proc. Natl. Acad. Sci. USA 2008, 105, 17955–17960. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Accession Number | Primer | Size (bp) |
|---|---|---|
| CFTR NM_000492.4 | s: 5′- CTCATTAGAAGGAGATGCTCCTG as: 5’- GCTCTTGTGGACAGTAATATATCG | 568 |
| SLC26A9 NM_052934.4 | s: 5´- CATTTGCTGTGCGCTTTCTG as: 5´- CCGCTTCTCCTGCTTCTTG | 568 |
| TMEM16A NM_018043.7 | s: 5´- CGACTACGTGTACATTTTCCG as: 5´- GATTCCGATGTCTTTGGCTC | 445 |
| GAPDH NM_001289726 | s: 5´- GTATTGGGCGCCTGGTCAC as: 5´- CTCCTGGAAGATGGTGATGG | 200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ousingsawat, J.; Centeio, R.; Schreiber, R.; Kunzelmann, K. Expression of SLC26A9 in Airways and Its Potential Role in Asthma. Int. J. Mol. Sci. 2022, 23, 2998. https://doi.org/10.3390/ijms23062998
Ousingsawat J, Centeio R, Schreiber R, Kunzelmann K. Expression of SLC26A9 in Airways and Its Potential Role in Asthma. International Journal of Molecular Sciences. 2022; 23(6):2998. https://doi.org/10.3390/ijms23062998
Chicago/Turabian StyleOusingsawat, Jiraporn, Raquel Centeio, Rainer Schreiber, and Karl Kunzelmann. 2022. "Expression of SLC26A9 in Airways and Its Potential Role in Asthma" International Journal of Molecular Sciences 23, no. 6: 2998. https://doi.org/10.3390/ijms23062998
APA StyleOusingsawat, J., Centeio, R., Schreiber, R., & Kunzelmann, K. (2022). Expression of SLC26A9 in Airways and Its Potential Role in Asthma. International Journal of Molecular Sciences, 23(6), 2998. https://doi.org/10.3390/ijms23062998