
Transcription Profiles Reveal Age-Dependent Variations of Photosynthetic Properties and Sugar Metabolism in Grape Leaves (Vitis vinifera L.)



Abstract
1. Introduction
2. Results
2.1. Morphological Characteristics of Leaves with Different Ages
2.2. The Micro-Morphology of Stomata on Differently Aged Leaves
2.3. The Physiological Difference of Leaves at Different Ages
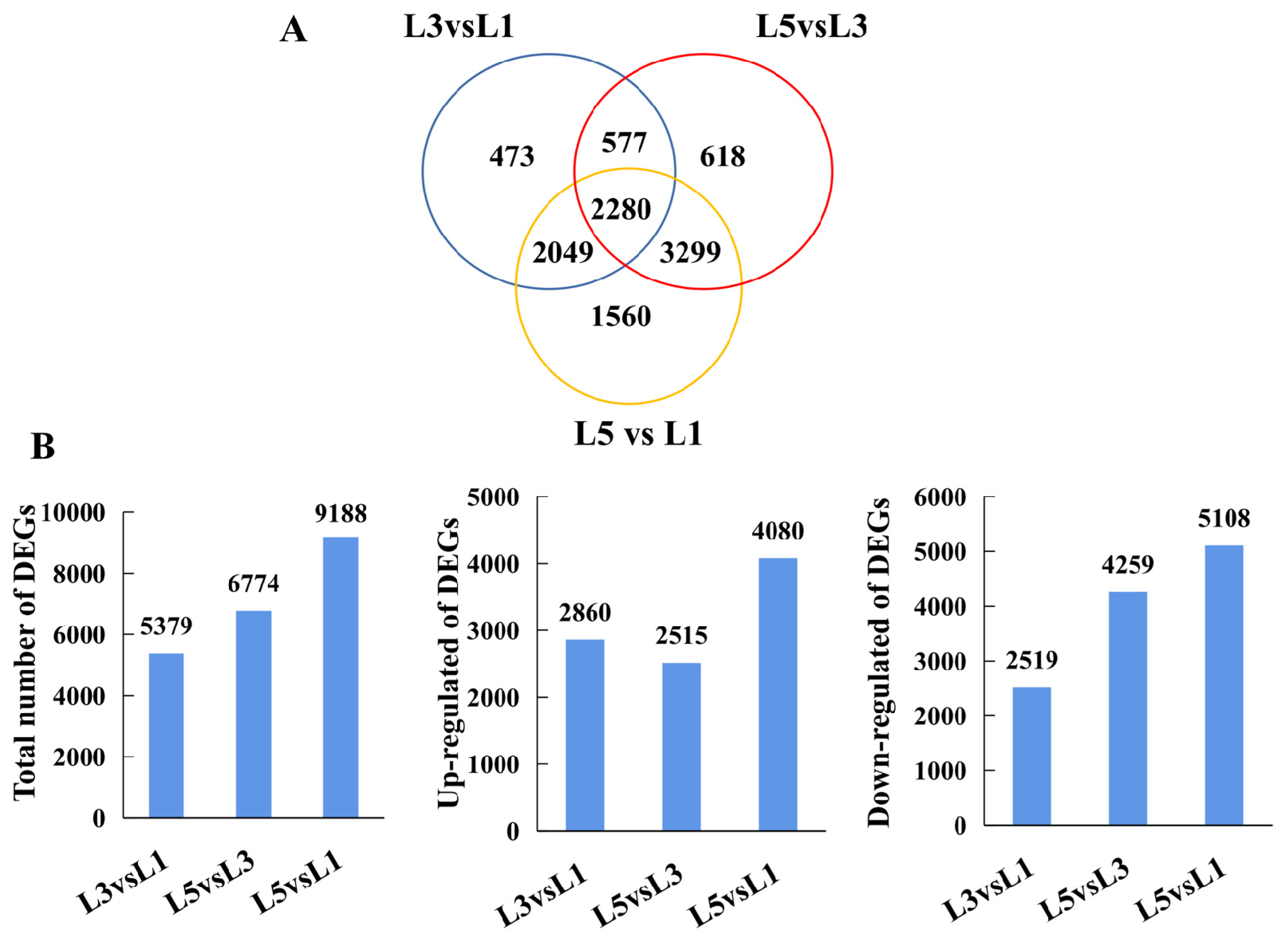
2.4. RNA-Seq Analysis and Functional Classification of DEGs
2.5. Expression Profile of the Genes Associated with Stomata Development and Movement in Differently Aged Leaves
2.6. Chlorophyll-Related Genes in Differently Aged Leaves
2.7. Expression Profiles of Photosynthesis-Related GENES in Differently Aged Leaves
2.8. Expression Profiles of Sucrose and Starch Genes in Differently Aged Leaves
3. Discussion
3.1. The Differences in Stomata Are Observed among Differently Aged Leaves at Morphology and Molecular Level
3.2. Dynamic Regulation of Chl Biosynthesis and Metabolism-Related Genes Resulted in the Changed Chla/b Contents
3.3. Genes Associated with Photosynthesis and Carbon Fixation Are Differentially Expressed among Differently Aged Leaves
3.4. Sucrose- and Starch-Biosynthesis-Related Genes Were Involved in Differences in Sugar Contents of Differently Aged Leaves
4. Materials and Methods
4.1. Plant Material and Sample Collection
4.2. Leaf Area Measurement
4.3. Photosynthesis Measurements
4.4. Scanning Electron Microscopy
4.5. Determination of Chl a/b and Sugar Content
4.6. Total RNA Extraction
4.7. RNA-Seq and Analysis
4.8. Identification and Analysis of Differentially Expressed Genes (DEGs)
4.9. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Braun, D.M.; Wang, L.; Ruan, Y.L. Understanding and manipulating sucrose phloem loading, unloading, metabolism, and signalling to enhance crop yield and food security. J. Exp. Bot. 2014, 65, 1713–1735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Wang, X.L.; Wang, X.F.; Xia, G.H.; Pan, Q.H.; Fan, R.C.; Wu, F.Q.; Yu, X.C.; Zhang, D.P. A shift of phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. Plant Physi. 2006, 142, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, N.; Vanhaeren, H.; Inze, D. Leaf size control: Complex coordination of cell division and expansion. Trends Plant Sci 2012, 17, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Albert, L.P.; Wu, J.; Prohaska, N.; de Camargo, P.B.; Huxman, T.E.; Tribuzy, E.S.; Ivanov, V.Y.; Oliveira, R.S.; Garcia, S.; Smith, M.N.; et al. Age-dependent leaf physiology and consequences for crown-scale carbon uptake during the dry season in an Amazon evergreen forest. New Phytol. 2018, 219, 870–884. [Google Scholar] [CrossRef]
- Turgeon, R. Phloem Loading: How Leaves Gain Their Independence. BioScience 2006, 56, 15–24. [Google Scholar] [CrossRef]
- Miyazawa, S.I.; Makino, A.; Terashima, I. Changes in mesophyll anatomy and sink–Source relationships during leaf development in quercus glauca, an evergreen tree showing delayed leaf greening. Plant Cell Environ. 2010, 26, 745–755. [Google Scholar] [CrossRef]
- Yoo, S.D.; Greer, D.H.; Laing, W.A.; McManus, M.T. Changes in photosynthetic efficiency and carotenoid composition in leaves of white clover at different developmental stages. Plant Physiol. Biochem. 2003, 41, 887–893. [Google Scholar] [CrossRef]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Johnson, M.P. Photosynthesis. Essays Biochem. 2016, 60, 255–273. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef]
- Vitlin Gruber, A.; Feiz, L. Rubisco Assembly in the Chloroplast. Front. Mol. Biosci. 2018, 5, 24. [Google Scholar] [CrossRef]
- Yu, A.; Xie, Y.; Pan, X.; Zhang, H.; Cao, P.; Su, X.; Chang, W.; Li, M. Photosynthetic Phosphoribulokinase Structures: Enzymatic Mechanisms and the Redox Regulation of the Calvin-Benson-Bassham Cycle. Plant Cell 2020, 32, 1556–1573. [Google Scholar] [CrossRef] [PubMed]
- Gurrieri, L.; Fermani, S.; Zaffagnini, M.; Sparla, F.; Trost, P. Calvin-Benson cycle regulation is getting complex. Trends Plant Sci. 2021, 26, 898–912. [Google Scholar] [CrossRef]
- Heyneke, E.; Fernia, A.R. Metabolic regulation of photosynthesis. Biochem. Soc. Trans. 2018, 46, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, M.; Lopezcalcagno, P.E.; Raines, C.A.; Furbank, R.T.; Von Caemmerer, S. Overexpression of the Rieske FeS protein of the Cytochrome b6f complex increases C4 photosynthesis in Setaria viridis. Commun. Biol. 2019, 2, 314. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Xu, J.; Teper, D.; Lee, D.; Wang, N. The transcriptome landscapes of citrus leaf in different developmental stages. Plant Mol. Biol. 2021, 106, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Chikov, V.I.; Abdrakhimov, F.A.; Bakirova, G.G.; Batasheva, S.N. The role of sink-source relationships between different organs in regulation of photosynthesis and productivity. Acta. Hortic. 2009, 835, 87–98. [Google Scholar] [CrossRef]
- Hedrich, R.; Shabala, S. Stomata in a saline world. Curr. Opin. Plant Biol. 2018, 46, 87–95. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef]
- Sakoda, K.; Yamori, W.; Shimada, T.; Sugano, S.S.; Hara-Nishimura, I.; Tanaka, Y. Higher Stomatal Density Improves Photosynthetic Induction and Biomass Production in Arabidopsis under Fluctuating Light. Frontiers in plant science. Front. Plant Sci. 2020, 11, 589603. [Google Scholar] [CrossRef]
- Nadeau, J.A.; Sack, F.D. Control of stomatal distribution on the Arabidopsis leaf surface. Science 2002, 296, 1697–1700. [Google Scholar] [CrossRef] [PubMed]
- Hunt, L.; Gray, J.E. The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr. Biol. 2009, 19, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhao, Y.; Lu, Y.; Su, X.; Chen, Y.; Shen, Y.; Lin, J.; Li, X. In vivo single-particle tracking of the aquaporin AtPIP2;1 in stomata reveals cell type-specific dynamics. Plant Physi. 2021, 185, 1666–1681. [Google Scholar] [CrossRef]
- Yamori, W.; Kusumi, K.; Iba, K.; Terashima, I. Increased stomatal conductance induces rapid changes to photosynthetic rate in response to naturally fluctuating light conditions in rice. Plant Cell Environ. 2020, 43, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Grimm, B. Organization of chlorophyll biosynthesis and insertion of chlorophyll into the chlorophyll-binding proteins in chloroplasts. Photosynth. Res. 2015, 126, 189–202. [Google Scholar] [CrossRef]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole Metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef]
- Lecourieux, F.; Kappel, C.; Lecourieux, D.; Serrano, A.; Torres, E.; Arce-Johnson, P.; Delrot, S. An update on sugar transport and signalling in grapevine. J. Exp. Bot. 2014, 65, 821–832. [Google Scholar] [CrossRef]
- Pervaiz, T.; Haifeng, J.; Salman Haider, M.; Cheng, Z.; Cui, M.; Wang, M.; Cui, L.; Wang, X.; Fang, J. Transcriptomic Analysis of Grapevine (cv. Summer Black) Leaf, Using the Illumina Platform. PLoS ONE 2016, 11, e0147369. [Google Scholar] [CrossRef]
- Lin, G.; Zhang, L.; Han, Z.; Yang, X.; Liu, W.; Li, E.; Chang, J.; Qi, Y.; Shpak, E.D.; Chai, J. A receptor-like protein acts as a specificity switch for the regulation of stomatal development. Genes Dev. 2017, 31, 927–938. [Google Scholar] [CrossRef]
- Hunt, L.; Bailey, K.J.; Gray, J.E. The signalling peptide EPFL9 is a positive regulator of stomatal development. New Phytol. 2010, 186, 609–614. [Google Scholar] [CrossRef]
- Franks, P.J.; Doheny-Adams, T.W.; Britton-Harper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Hu, H.; Qin, X.; Zeise, B.; Xu, D.; Rappel, W.-J.; Boron, W.F.; Schroeder, J.I. Reconstitution of CO2 Regulation of SLAC1 Anion Channel and Function of CO2-Permeable PIP2;1 Aquaporin as CARBONIC ANHYDRASE4 Interactor. Plant Cell 2016, 28, 568–582. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Wang, X.H.; Huang, J.Y.; Zhang, C.G.; Chen, Y.L. Phosphorylation of plasma membrane aquaporin PIP2;1 in C-terminal affects light-induced stomatal opening in Arabidopsis. Plant Signal. Behav. 2020, 15, 1795394. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, M.; Matus, J.T.; Francia, P.; Rusconi, F.; Cañón, P.; Medina, C.; Conti, L.; Cominelli, E.; Tonelli, C.; Arce-Johnson, P. The grapevine guard cell-related VvMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant Biol. 2011, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Pons, T.L.; Westbeek, M.H. Analysis of differences in photosynthetic nitrogen-use efficiency between four contrasting species. Physiol. Plant. 2004, 122, 68–78. [Google Scholar] [CrossRef]
- Kumar, A.M.; Soll, D. Antisense HEMA1 RNA expression inhibits heme and chlorophyll biosynthesis in Arabidopsis. Plant Physiol. 2000, 122, 49–56. [Google Scholar] [CrossRef]
- Matsumoto, F.; Obayashi, T.; Sasaki-Sekimoto, Y.; Ohta, H.; Takamiya, K.; Masuda, T. Gene expression profiling of the tetrapyrrole metabolic pathway in Arabidopsis with a mini-array system. Plant Physiol. 2004, 135, 2379–2391. [Google Scholar] [CrossRef]
- Oster, U.; Tanaka, R.; Tanaka, A.; Rüdiger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J. 2000, 21, 305–310. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Kim, Y.S.; Yoo, S.C.; Hörtensteiner, S.; Paek, N.C. 7-Hydroxymethyl chlorophyll a reductase functions in metabolic channeling of chlorophyll breakdown intermediates during leaf senescence. Biochem. Biophys. Res. Commun. 2013, 430, 32–37. [Google Scholar] [CrossRef]
- Takashima, T.; Hikosaka, K.; Hirose, T. Photosynthesis or persistence: Nitrogen allocation in leaves of evergreen and deciduous Quercus species. Plant Cell Environ. 2004, 27, 1047–1054. [Google Scholar] [CrossRef]
- Salesse-Smith, C.E.; Sharwood, R.E.; Busch, F.A.; Kromdijk, J.; Bardal, V.; Stern, D.B. Overexpression of Rubisco subunits with RAF1 increases Rubisco content in maize. Nat. Plants 2018, 4, 802–810. [Google Scholar] [CrossRef]
- Simkin, A.J.; Mcausland, L.; Lawson, T.; Raines, C.A. Overexpression of the RieskeFeS protein increases electron transport rates and biomass yield. Plant Physiol. 2017, 175, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, S.; Lawson, T.; Zakhleniuk, O.V.; Lloyd, J.C.; Raines, C.A.; Fryer, M. Increased sedoheptulose-1,7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 2005, 138, 451–460. [Google Scholar] [CrossRef]
- Avron, M.; Gibbs, M. Properties of phosphoribulokinase of whole chloroplasts. Plant Physiol 1974, 53, 136–139. [Google Scholar] [CrossRef]
- Melandri, B.A.; Baccarini, A.; Pupillo, P. Glyceraldehyde-3-phosphate dehydrogenase in photosynthetic tissues: Kinetic evidence for competitivity between NADP and NAD. Biochem. Biophys. Res. Commun. 1968, 33, 160–164. [Google Scholar] [CrossRef]
- Baroli, I.; Price, G.D.; Caemmerer, B.S.V. The contribution of photosynthesis to the red light response of stomatal conductance. Plant Physiol. 2008, 146, 737–747. [Google Scholar] [CrossRef]
- Uematsu, K.; Suzuki, N.; Iwamae, T.; Inui, M.; Yukawa, H. Increased fructose 1,6-bisphosphate aldolase in plastids enhances growth and photosynthesis of tobacco plants. J. Exp. Bot. 2012, 63, 3001–3009. [Google Scholar] [CrossRef]
- Simkin, A.J.; McAusland, L.; Headland, L.R.; Lawson, T.; Raines, C.A. Multigene manipulation of photosynthetic carbon assimilation increases CO2 fixation and biomass yield in tobacco. J. Exp. Bot. 2015, 66, 4075–4090. [Google Scholar] [CrossRef]
- Chen, C.; Yuan, Y.; Zhang, C.; Li, H.; Ma, F.; Li, M. Sucrose phloem unloading follows an apoplastic pathway with high sucrose synthase in Actinidia fruit. Plant Sci. 2017, 255, 40–50. [Google Scholar] [CrossRef]
- Albi, T.; Ruiz, M.T.; de Los Reyes, P.; Valverde, F.; Romero, J.M. Characterization of the Sucrose Phosphate Phosphatase (SPP) Isoforms from Arabidopsis thaliana and Role of the S6PPc Domain in Dimerization. PLoS ONE 2016, 11, e0166308. [Google Scholar] [CrossRef]
- Lunn, J.E.; ap Rees, T. Apparent equilibrium constant and mass-action ratio for sucrose-phosphate synthase in seeds of Pisum sativum. Biochem. J. 1990, 267, 739–743. [Google Scholar] [CrossRef]
- Wan, H.; Wu, L.; Yang, Y.; Zhou, G.; Ruan, Y.L. Evolution of Sucrose Metabolism: The Dichotomy of Invertases and Beyond. Trends Plant Sci. 2018, 23, 163–177. [Google Scholar] [CrossRef]
- Fettke, J.; Chia, T.; Eckermann, N.; Smith, A.; Steup, M. A transglucosidase necessary for starch degradation and maltose metabolism in leaves at night acts on cytosolic heteroglycans (SHG). Plant J. 2006, 46, 668–684. [Google Scholar] [CrossRef]
- Riggs, J.W.; Cavales, P.C.; Chapiro, S.M.; Callis, J. Identification and biochemical characterization of the fructokinase gene family in Arabidopsis thaliana. BMC Plant Biol. 2017, 17, 83. [Google Scholar] [CrossRef]
- Clifton, P.; Keogh, J. Starch. In Encyclopedia of Food and Health; Academic Press: New York, NY, USA, 2015; pp. 146–151. [Google Scholar]
- Gandin, A.; Lapointe, L.; Dizengremel, P. The alternative respiratory pathway allows sink to cope with changes in carbon availability in the sink-limited plant Erythronium americanum. J. Exp. Bot. 2009, 60, 4235–4248. [Google Scholar] [CrossRef]
- Burnison, B.K. Modified dimethyl sulfoxide (dmso) extraction for chlorophyll analysis of phytoplankton. Can. J. Fish. Aquat. 2011, 37, 729–733. [Google Scholar] [CrossRef]
- Barros, L.B.; Larissa, M.; Biasi, L.A.; Profio, F.D.; Reynolds, A.G. Use of hplc for characterization of sugar and phenolic compounds in vitis labrusca juice. Idesia 2014, 32, 89–94. [Google Scholar] [CrossRef][Green Version]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463. [Google Scholar]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenfuhrer, J.; Depends, R.; Db, G. Type Package Title topGO: Enrichment Analysis for Gene Ontology. Version 2.10.0. 1 October 2013. Available online: https://bioconductor.org/packages/devel/bioc/manuals/topGO/man/topGO.pdf (accessed on 11 January 2022).
- Yu, G.; Wang, L.; Han, Y.; He, Q. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Gene ID | Gene Name | log2FC(L3vsL1) | log2FC(L5vsL1) | log2FC(L5vsL3) | padjL3vsL1 | padjL5vsL1 | padjL5vsL3 |
|---|---|---|---|---|---|---|---|---|
| Stomata pattern | VIT_05s0062g01380 | EPIDERMAL PATTERNING FACTOR LIKE9 (EPFL9) | −1.32 | −4.17 | −2.90 | 4.35 × 10−1 | 3.63 × 10−1 | 6.29 × 10−2 |
| VIT_18s0164g00010 | EPIDERMAL PATTERNING FACTOR2 (EPF2) | −2.92 | −9.76 | −6.88 | 2.32 × 10−23 | 2.41 × 10−15 | 7.37 × 10−7 | |
| VIT_18s0001g08350 | EPIDERMAL PATTERNING FACTOR LIKE2 (EPFL2) | −5.79 | −6.66 | −0.95 | 5.64 × 10−7 | 4.33 × 10−7 | 7.47 × 10−1 | |
| VIT_06s0004g07880 | EPIDERMAL PATTERNING FACTOR LIKE6 (EPFL6) | −2.68 | −1.26 | 1.38 | 9.31 × 10−36 | 1.19 × 10−10 | 3.18 × 10−7 | |
| VIT_05s0020g04050 | EPIDERMAL PATTERNING FACTOR LIKE4 (EPFL4) | −6.17 | −8.99 | −2.95 | 2.17 × 10−15 | 1.35 × 10−13 | 3.38 × 10−1 | |
| VIT_00s0391g00030 | EPIDERMAL PATTERNING FACTOR LIKE1 (EPFL1) | −2.07 | −3.64 | −1.61 | 1.07 × 10−11 | 2.90 × 10−18 | 2.12 × 10−3 | |
| VIT_00s0386g00040 | EPIDERMAL PATTERNING FACTOR LIKE2 (EPFL2) | −1.52 | −1.49 | −0.02 | 1.09 × 10−6 | 1.37 × 10−6 | 9.54 × 10−1 | |
| VIT_09s0002g07030 | TOO MANY MOUTHS (TMM) | −5.67 | −11.02 | −5.40 | 7.77 × 10−55 | 3.53 × 10−20 | 4.66 × 10−4 | |
| VIT_16s0022g02030 | ERECTA-like kinase 1 (ERL1) | −2.91 | −5.93 | −3.06 | 1.96 × 10−58 | 4.28 × 10−117 | 1.95 × 10−17 | |
| Stomata movement | VIT_02s0025g04930 | STOMATAL ANION CHANNEL PROTEIN 1 (SLAC1) | 3.07 | 3.77 | 0.65 | 4.33 × 10−28 | 3.22 × 10−41 | 2.47 × 10−4 |
| VIT_01s0010g02940 | CALMODULIN-LIKE31 (CML31) | −4.29 | −2.67 | 1.58 | 2.13 × 10−3 | 3.12 × 10−2 | 2.68 × 10−1 | |
| VIT_18s0122g00180 | CALMODULIN-LIKE37 (CML37) | −3.29 | −1.97 | 1.27 | 4.78 × 10−4 | 5.35 × 10−2 | 2.39 × 10−3 | |
| VIT_18s0001g11830 | CALMODULIN-LIKE41 (CML41) | −0.69 | 1.59 | 2.24 | 3.64 × 10−3 | 5.45 × 10−8 | 1.02 × 10−17 | |
| VIT_18s0001g03880 | CALMODULIN-LIKE29 (CML29) | 6.49 | 8.58 | 2.04 | 4.50 × 10−76 | 1.24 × 10−226 | 5.25 × 10−23 | |
| VIT_18s0001g01630 | CALMODULIN-LIKE44 (CML44) | 5.63 | 9.25 | 3.58 | 3.88 × 10−195 | 7.97 × 10−293 | 1.02 × 10−91 | |
| VIT_17s0000g01630 | CALMODULIN-LIKE19 (CML19) | 0.06 | −1.70 | −1.80 | 8.32 × 10−1 | 2.01 × 10−8 | 5.19 × 10−13 | |
| VIT_16s0039g01880 | CALMODULIN-LIKE18 (CML18) | 0.20 | 1.28 | 1.04 | 6.22 × 10−1 | 2.80 × 10−3 | 1.45 × 10−2 | |
| VIT_14s0030g02150 | CALMODULIN-LIKE1 (CML11) | −0.71 | 3.39 | 4.09 | 6.45 × 10−1 | 6.46 × 10−6 | 2.41 × 10−6 | |
| VIT_11s0016g05740 | CALMODULIN-LIKE5 (CML5) | 1.23 | 1.25 | −0.03 | 1.02 × 10−12 | 6.04 × 10−13 | 8.69 × 10−1 | |
| VIT_03s0063g00530 | CALMODULIN-LIKE30 (CML30) | 1.50 | 3.11 | 1.56 | 8.51 × 10−8 | 3.62 × 10−26 | 1.27 × 10−10 | |
| VIT_14s0068g02190 | CHLORIDE CHANNEL B (CLC-B) | −0.38 | −2.37 | −2.04 | 8.09 × 10−2 | 1.14 × 10−11 | 6.02 × 10−10 | |
| VIT_07s0130g00400 | CHLORIDE CHANNEL C (CLC-C) | 1.41 | 1.44 | −0.02 | 2.63 × 10−16 | 2.03 × 10−18 | 8.68 × 10−1 | |
| VIT_03s0038g04260 | CHLORIDE CHANNEL B (CLC-E) | 0.98 | 1.76 | 0.73 | 1.56 × 10−14 | 5.44 × 10−33 | 2.67 × 10−8 | |
| VIT_08s0040g01890 | PLASMA MEMBRANE INTRINSIC PROTEIN2-1 (PIP2-1) | −0.63 | −5.08 | −4.50 | 3.75 × 10−4 | 1.32 × 10−11 | 1.79 × 10−9 | |
| VIT_06s0004g02850 | PLASMA MEMBRANE INTRINSIC PROTEINS2-7 (PIP2-7) | −0.18 | −2.45 | −2.32 | 3.92 × 10−1 | 4.52 × 10−17 | 1.34 × 10−24 | |
| VIT_03s0038g02520 | PLASMA MEMBRANE INTRINSIC PROTEINS2-7 (PIP2-7) | −0.75 | −2.00 | −1.30 | 1.25 × 10−9 | 1.49 × 10−26 | 1.81 × 10−11 | |
| VIT_03s0038g01410 | PLASMA MEMBRANE INTRINSIC PROTEINS1-3 (PIP1-3) | 4.09 | 1.46 | −2.67 | 1.40 × 10−110 | 7.08 × 10−11 | 9.78 × 10−48 | |
| VIT_15s0046g02420 | PLASMA MEMBRANE INTRINSIC PROTEINS1-1 (PIP1-1) | −0.43 | −1.71 | −1.33 | 1.47 × 10−2 | 3.29 × 10−19 | 2.09 × 10−18 | |
| VIT_15s0046g02410 | PLASMA MEMBRANE INTRINSIC PROTEINS1-2 (PIP1-2) | −1.06 | −4.40 | −3.38 | 3.61 × 10−9 | 2.70 × 10−94 | 5.84 × 10−46 | |
| VIT_13s0067g00220 | PLASMA MEMBRANE INTRINSIC PROTEINS1-2 (PIP1-2) | −0.07 | −2.88 | −2.86 | 6.54 × 10−1 | 6.33 × 10−4 | 6.21 × 10−4 | |
| VIT_08s0056g00800 | MYB transcription factor 60 (MYB60) | 2.87 | 2.91 | 0.00 | 1.39 × 10−38 | 3.20 × 10−39 | 9.99 × 10−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-M.; You, J.-L.; Nie, W.-F.; Sun, M.-H.; Xie, Z.-S. Transcription Profiles Reveal Age-Dependent Variations of Photosynthetic Properties and Sugar Metabolism in Grape Leaves (Vitis vinifera L.). Int. J. Mol. Sci. 2022, 23, 2243. https://doi.org/10.3390/ijms23042243
Li Y-M, You J-L, Nie W-F, Sun M-H, Xie Z-S. Transcription Profiles Reveal Age-Dependent Variations of Photosynthetic Properties and Sugar Metabolism in Grape Leaves (Vitis vinifera L.). International Journal of Molecular Sciences. 2022; 23(4):2243. https://doi.org/10.3390/ijms23042243
Chicago/Turabian StyleLi, You-Mei, Jia-Ling You, Wen-Feng Nie, Meng-Hao Sun, and Zhao-Sen Xie. 2022. "Transcription Profiles Reveal Age-Dependent Variations of Photosynthetic Properties and Sugar Metabolism in Grape Leaves (Vitis vinifera L.)" International Journal of Molecular Sciences 23, no. 4: 2243. https://doi.org/10.3390/ijms23042243
APA StyleLi, Y.-M., You, J.-L., Nie, W.-F., Sun, M.-H., & Xie, Z.-S. (2022). Transcription Profiles Reveal Age-Dependent Variations of Photosynthetic Properties and Sugar Metabolism in Grape Leaves (Vitis vinifera L.). International Journal of Molecular Sciences, 23(4), 2243. https://doi.org/10.3390/ijms23042243