
Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
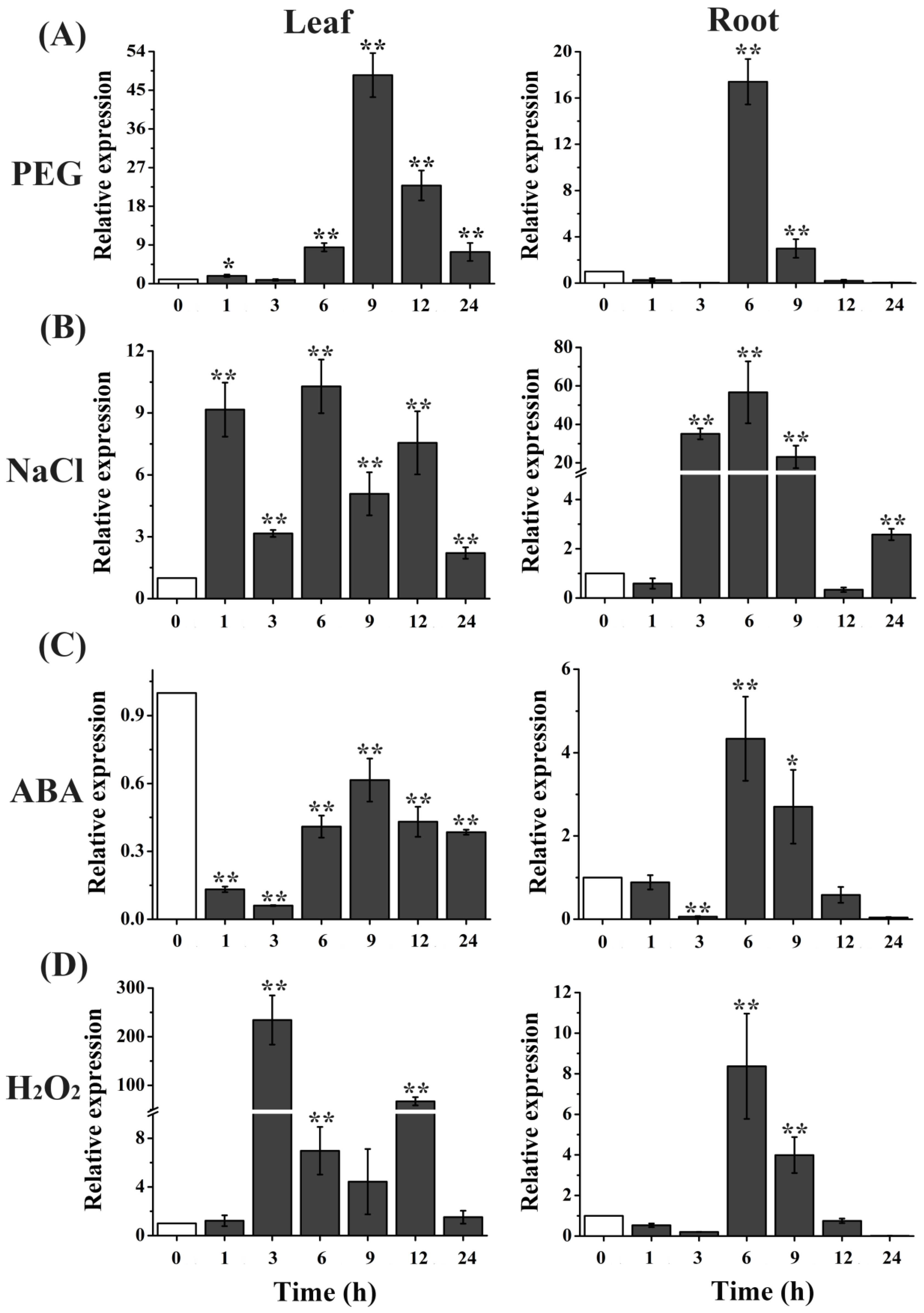
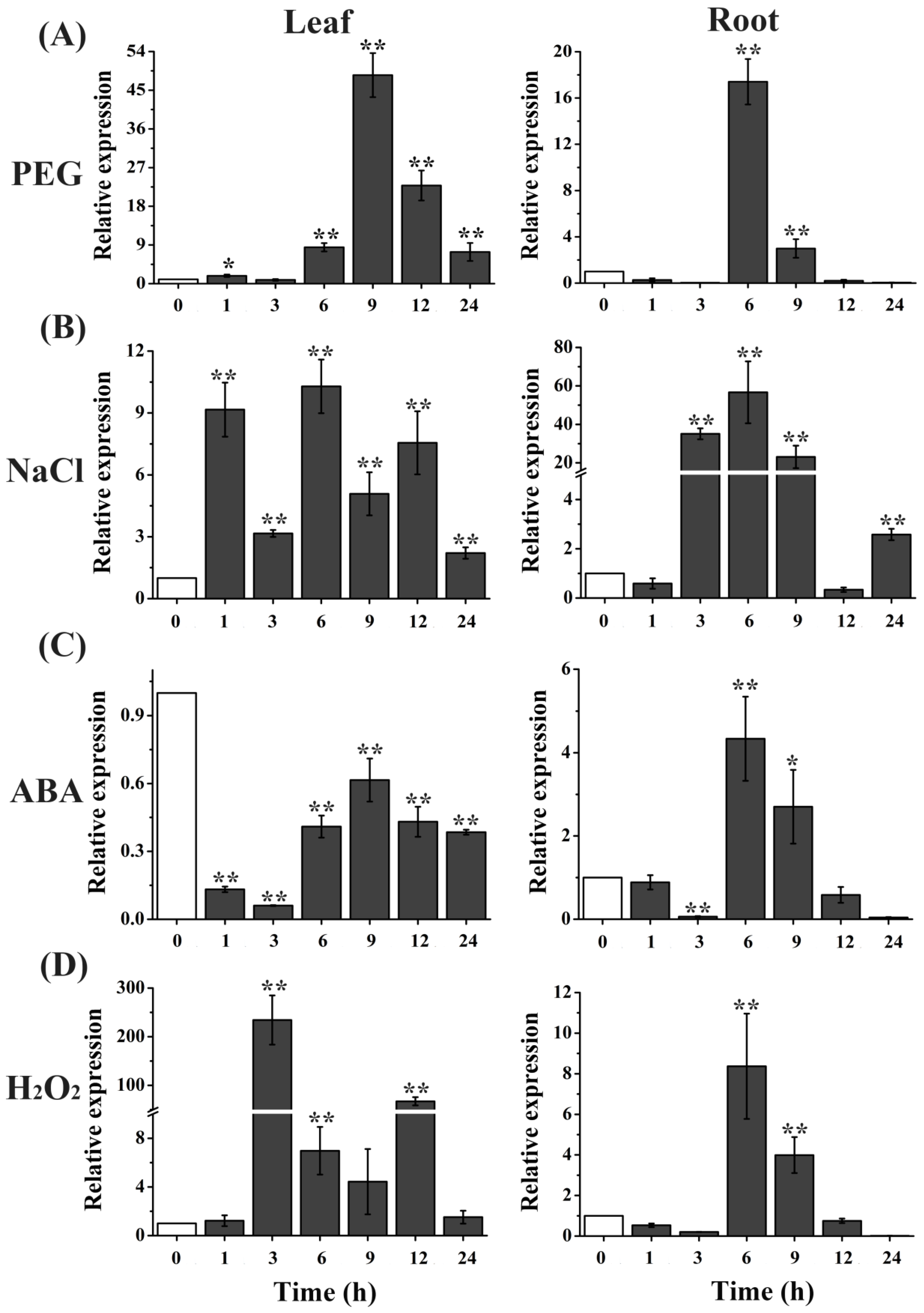
2.1. TaTIP4;1 Expression Was Strongly Induced by PEG, NaCl and H2O2 in Wheat
2.2. Generation of Transgenic Plants Overexpressing TaTIP4;1 in Arabidopsis and Rice
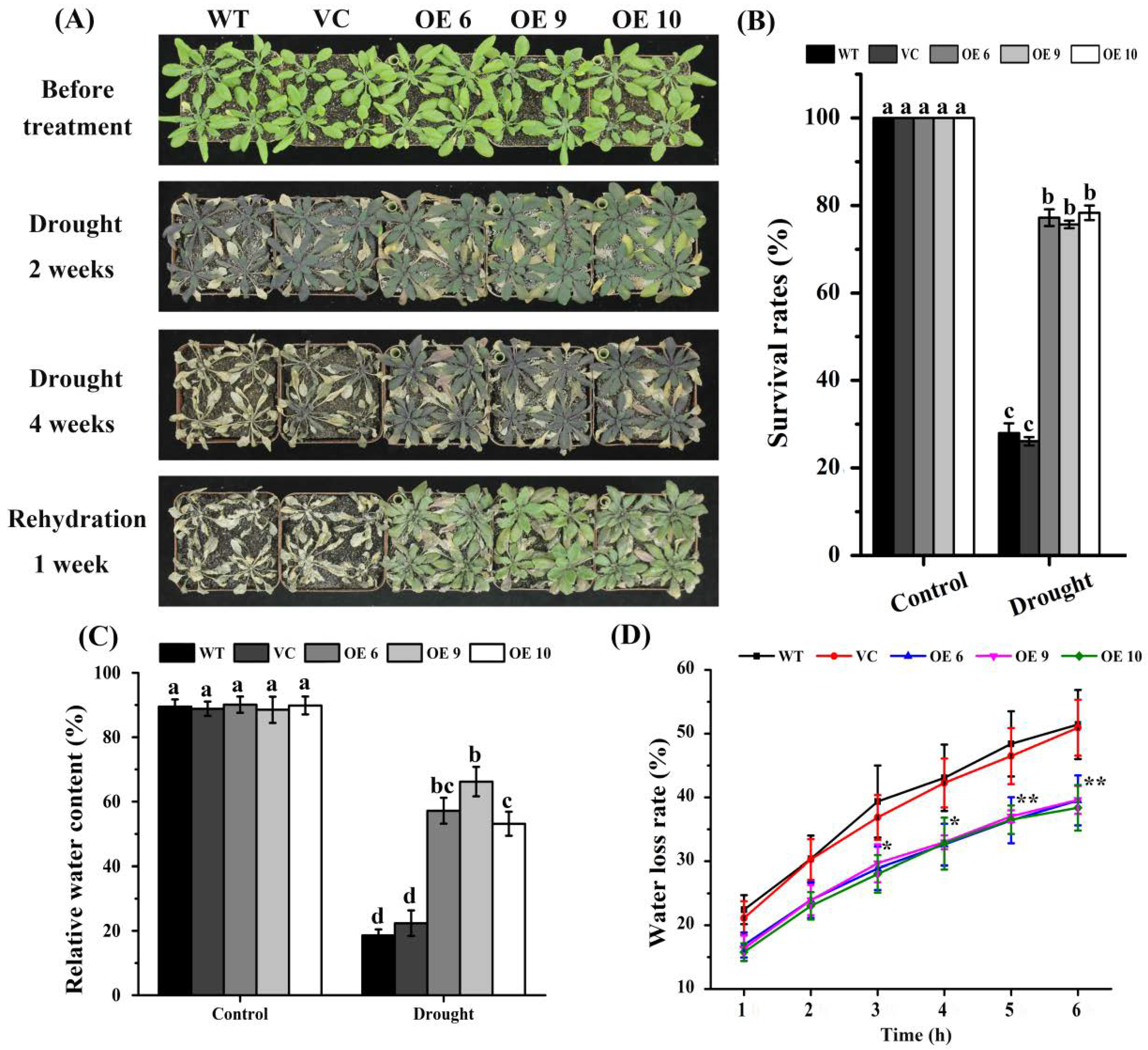
2.3. Overexpression of TaTIP4;1 Improved Drought Tolerance of Arabidopsis
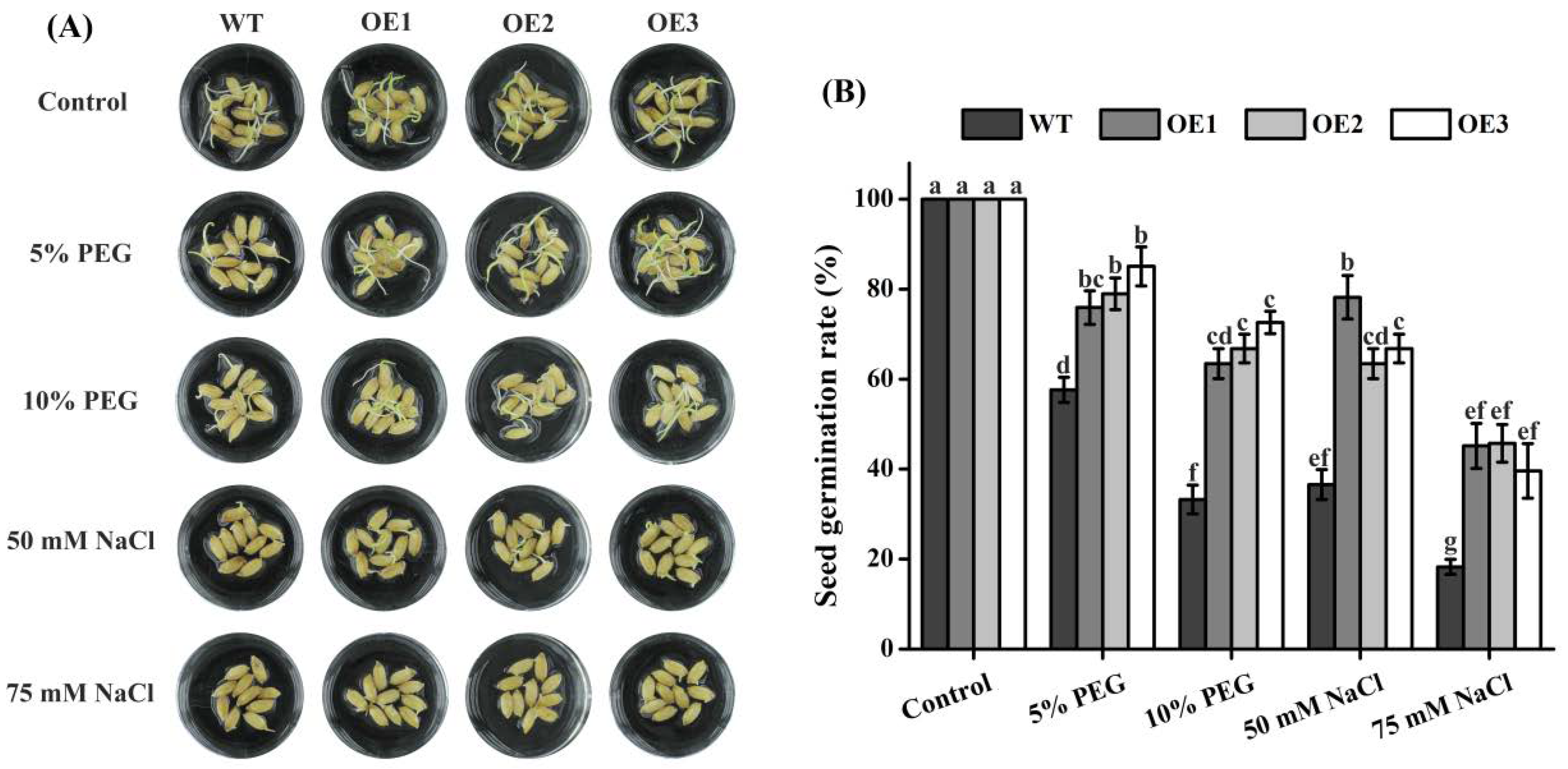
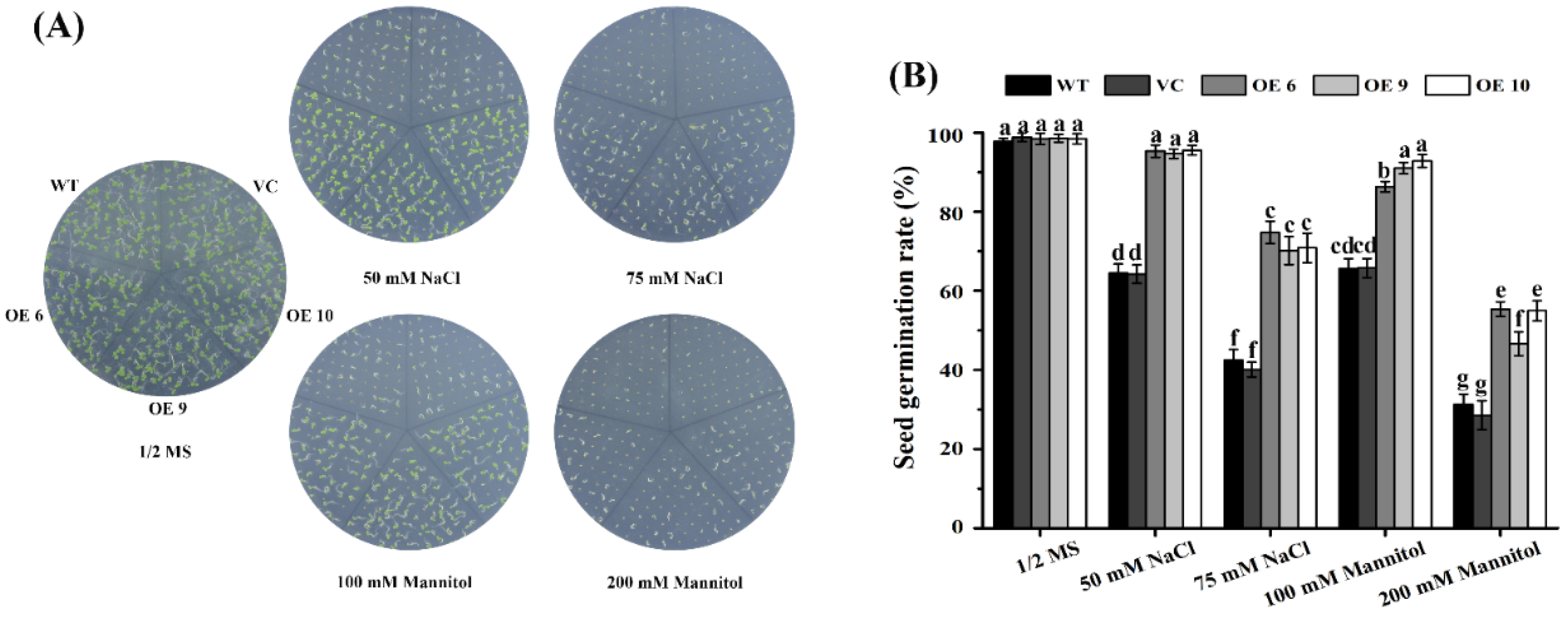
2.4. Arabidopsis TaTIP4;1 Overexpressors Had Increased Seed Germination Rates under Salt and Osmotic Stress
2.5. Arabidopis Seedlings Overexpressing TaTIP4;1 Had Enhanced Tolerance to Salinity and Osmotic Stress
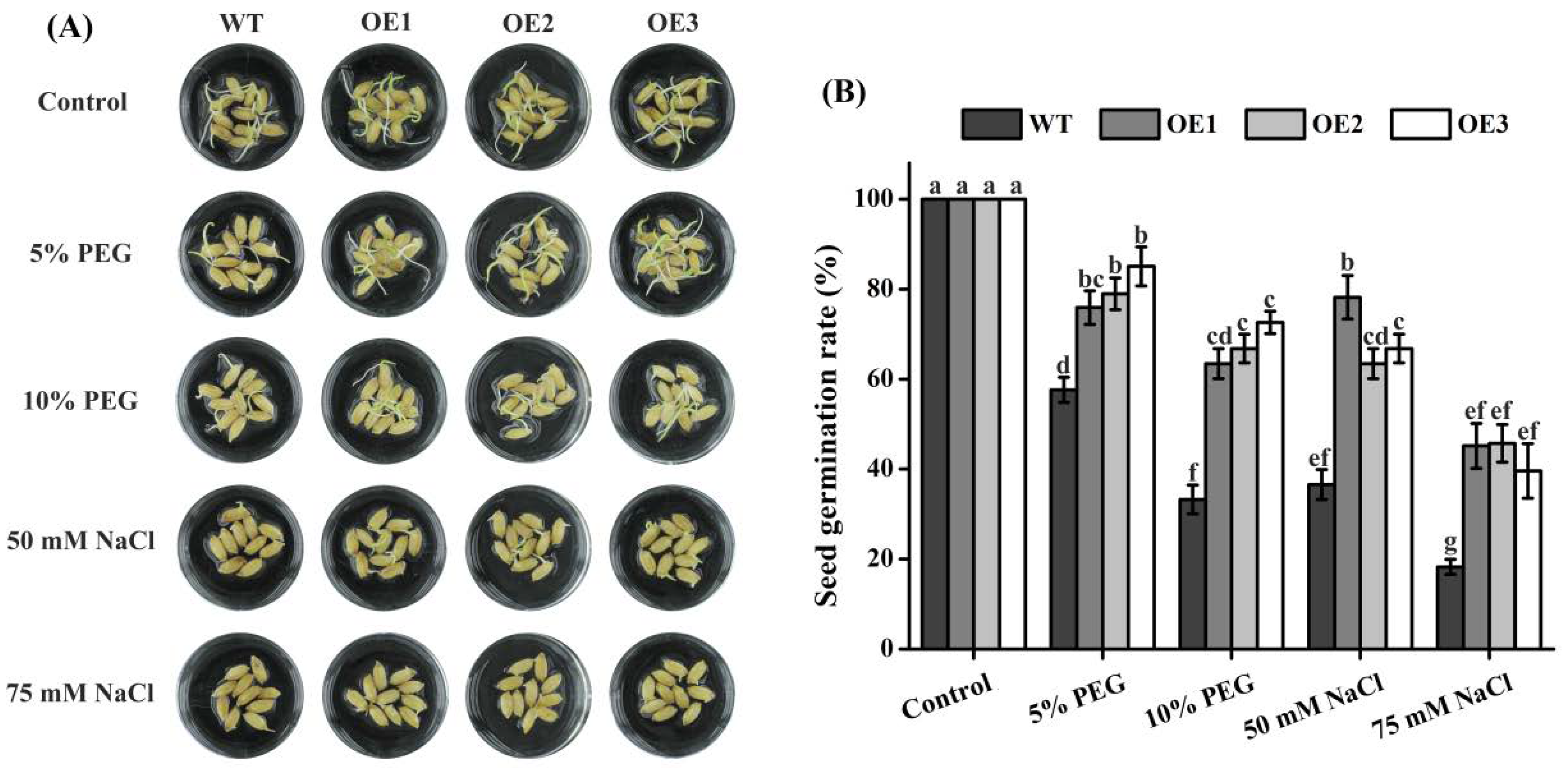
2.6. Seed Germination of Rice TaTIP4;1 Overexpressors Was Insensitive to Salt and Osmotic Stress
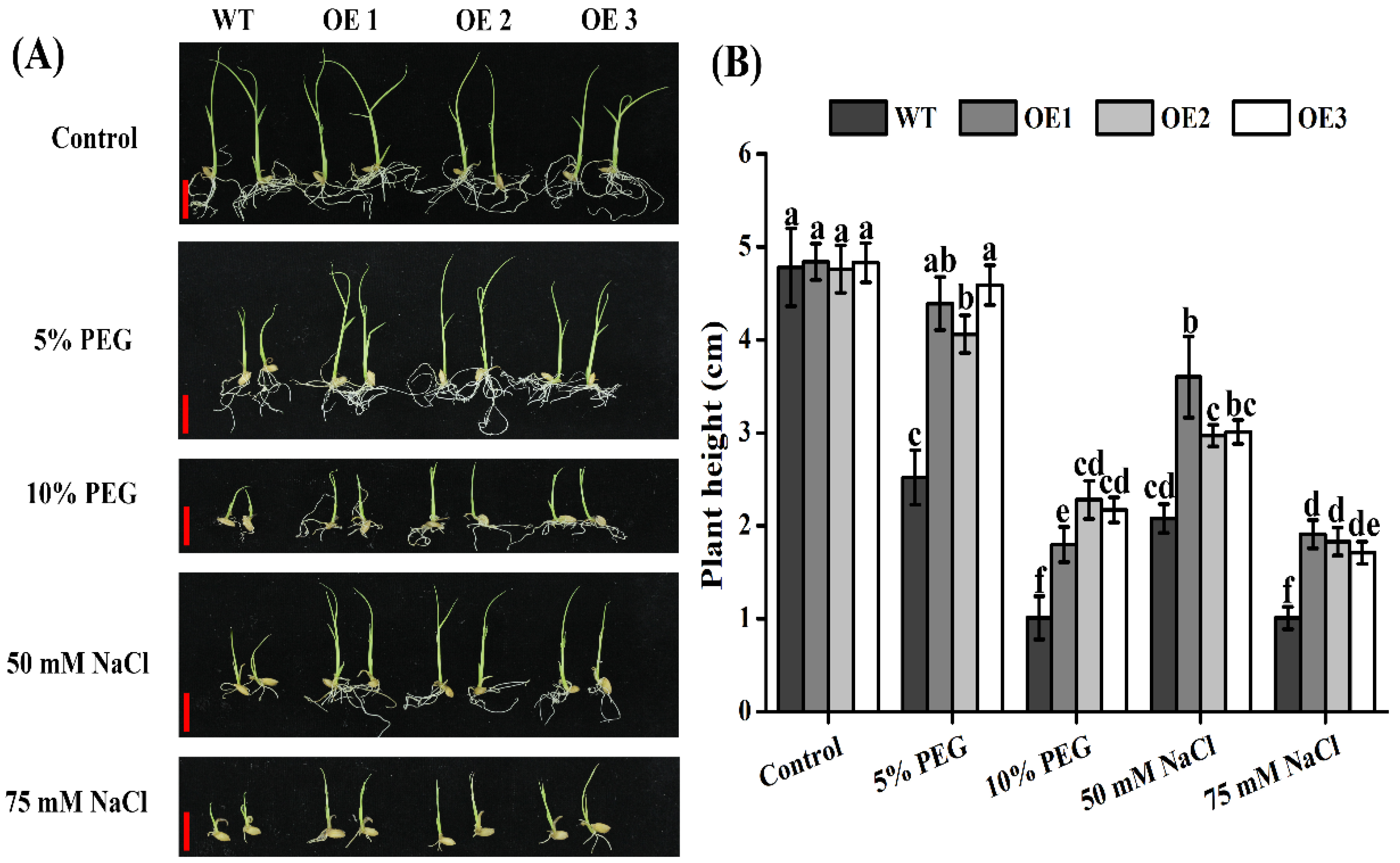
2.7. Rice TaTIP4;1 Overexpressors Grew Better Than WT after Challenged by PEG6000, Salt and Mannitol
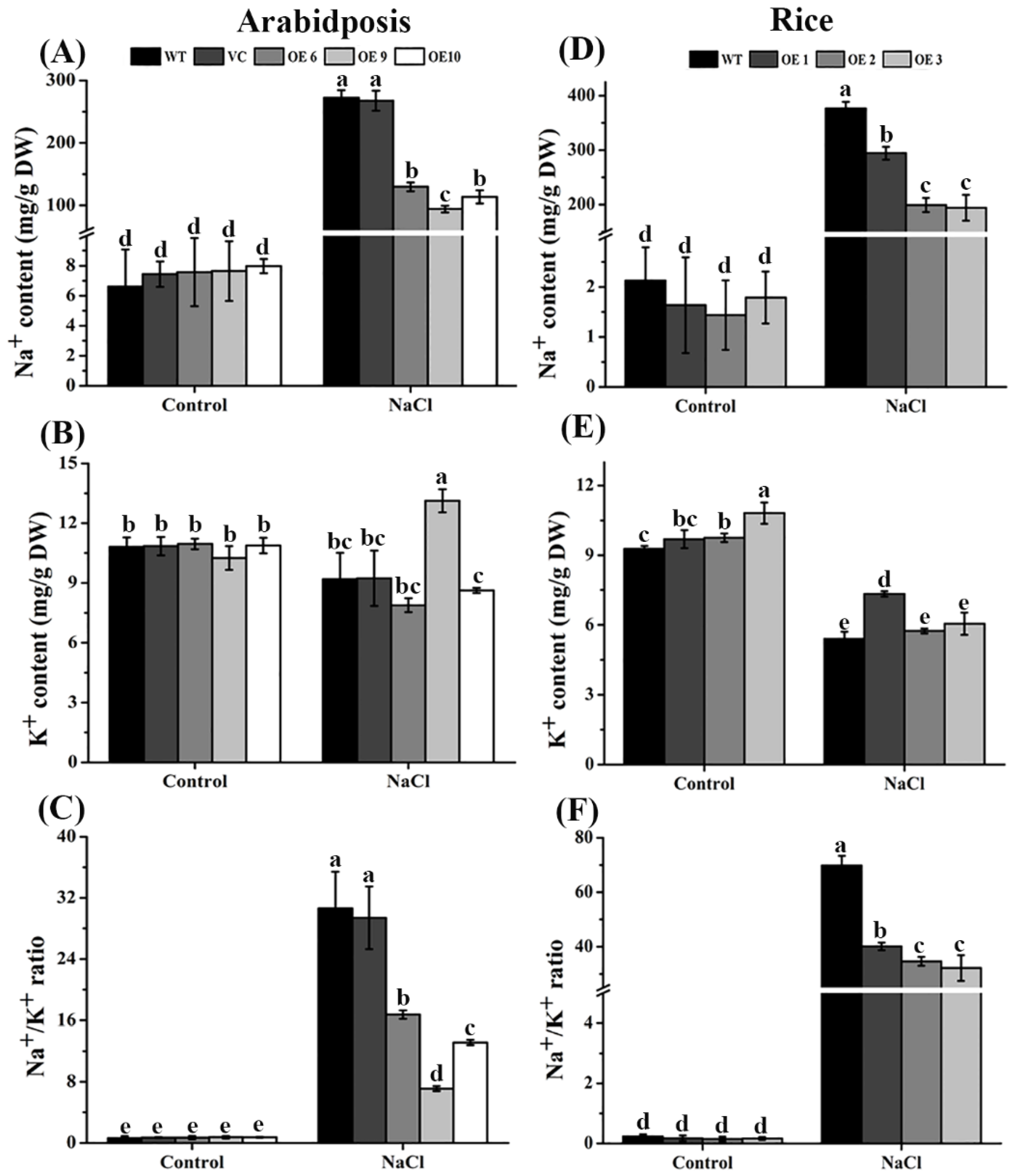
2.8. Salt-Stimulated Increases of Na+ Contents Were Markedly Reduced in TaTIP4;1 OE Lines of Arabidopsis and Rice
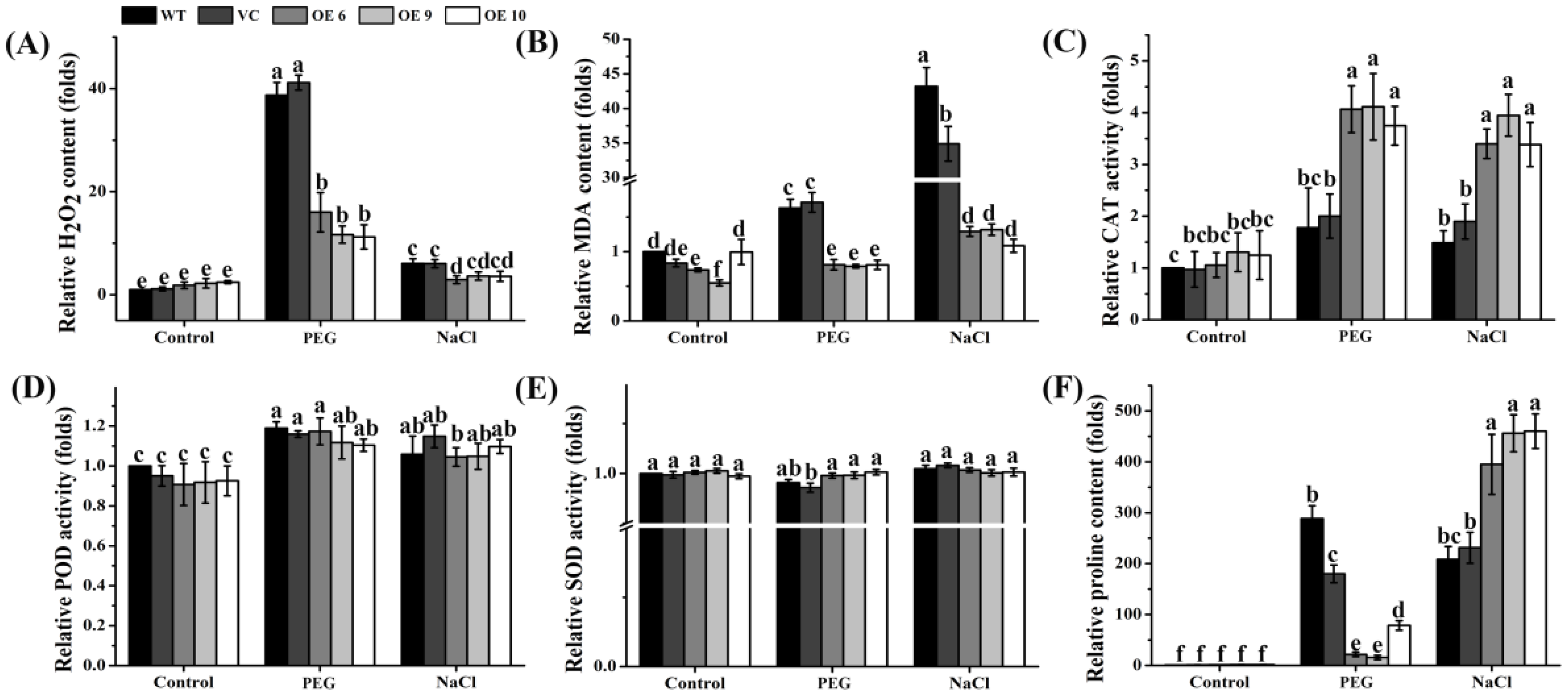
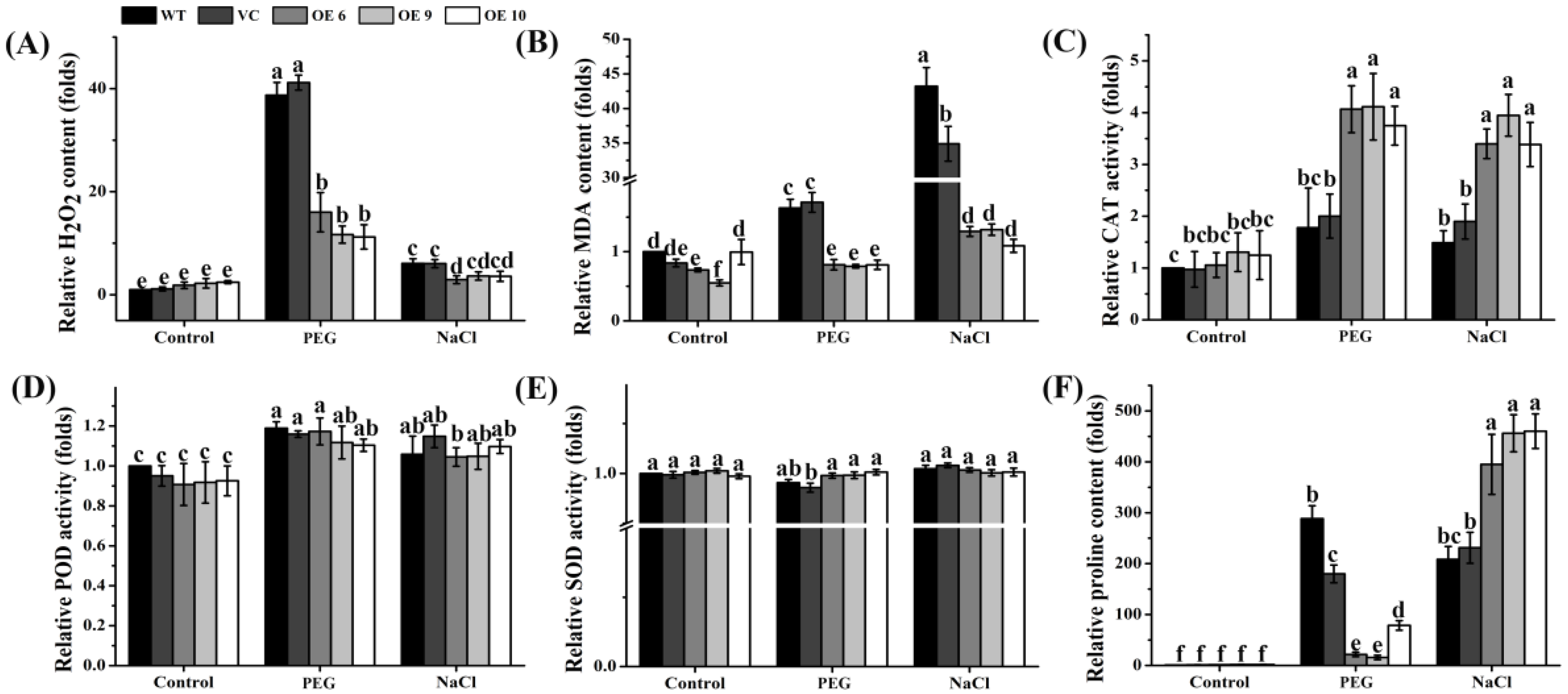
2.9. Overexpression of TaTIP4;1 Altered H2O2 Accumulation, MDA Contents, the Activity of Some Antioxidases, and Proline Levels in Arabidopsis upon Drought and Salt Stress
2.10. TaTIP4;1 Expression in Rice Affected the Levels of H2O2 and MDA, Antioxidase Activities and Proline Accumulation under Salt and Osmotic Stress
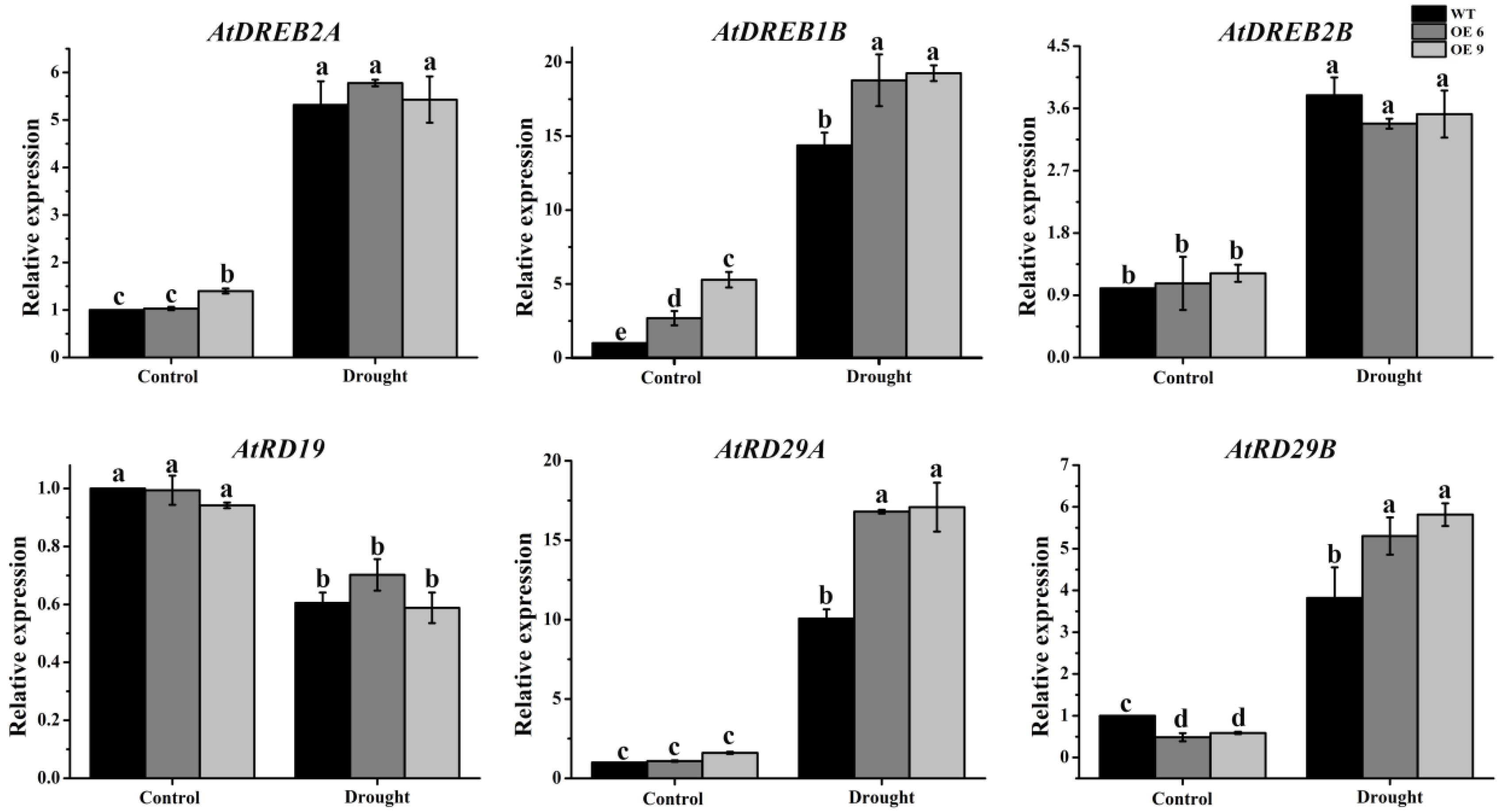
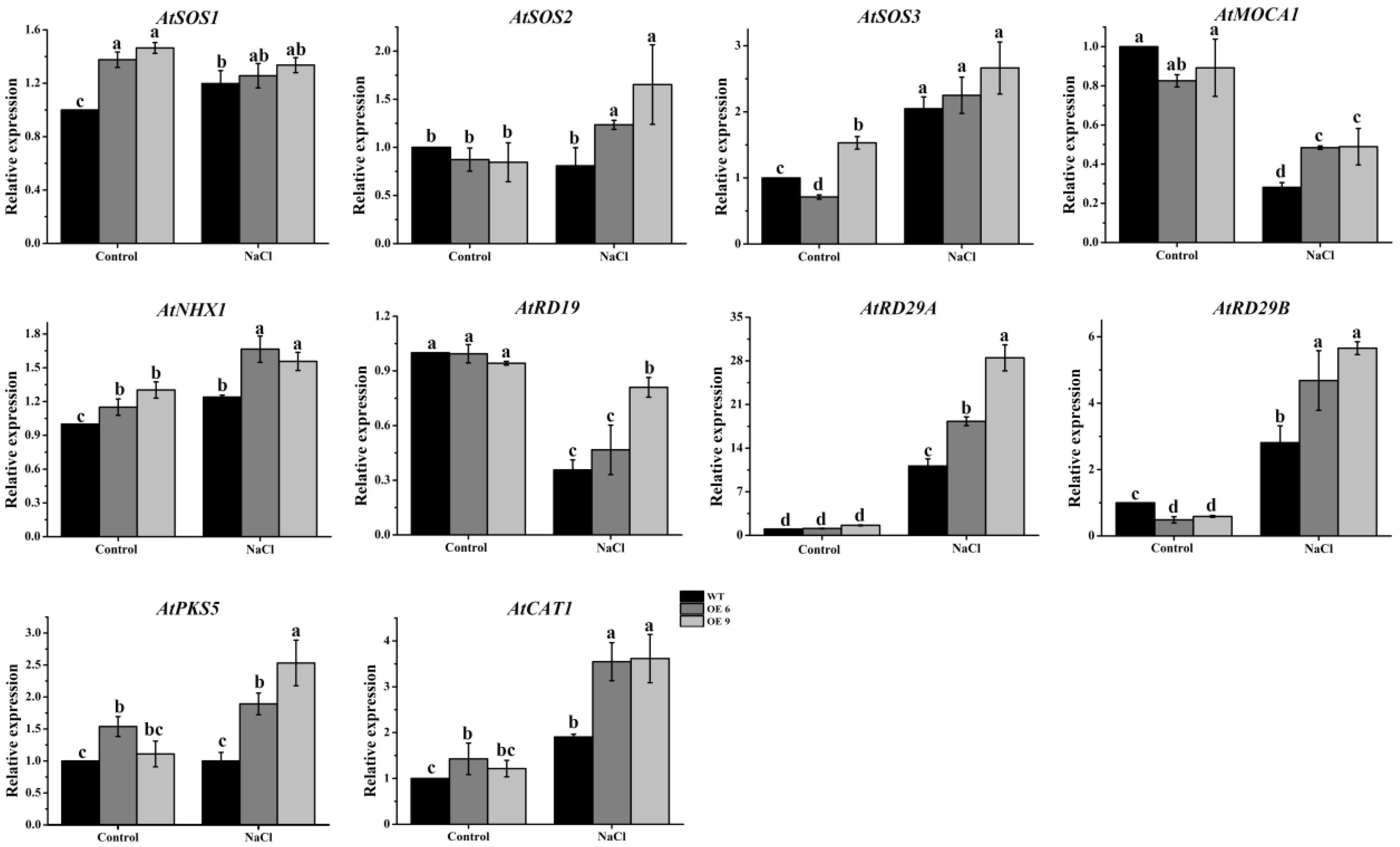
2.11. The Expression of Several Drought Responsive Genes Was Upregulated in TaTIP4;1 OE Lines of Arabidopsis and Rice under Water Stress
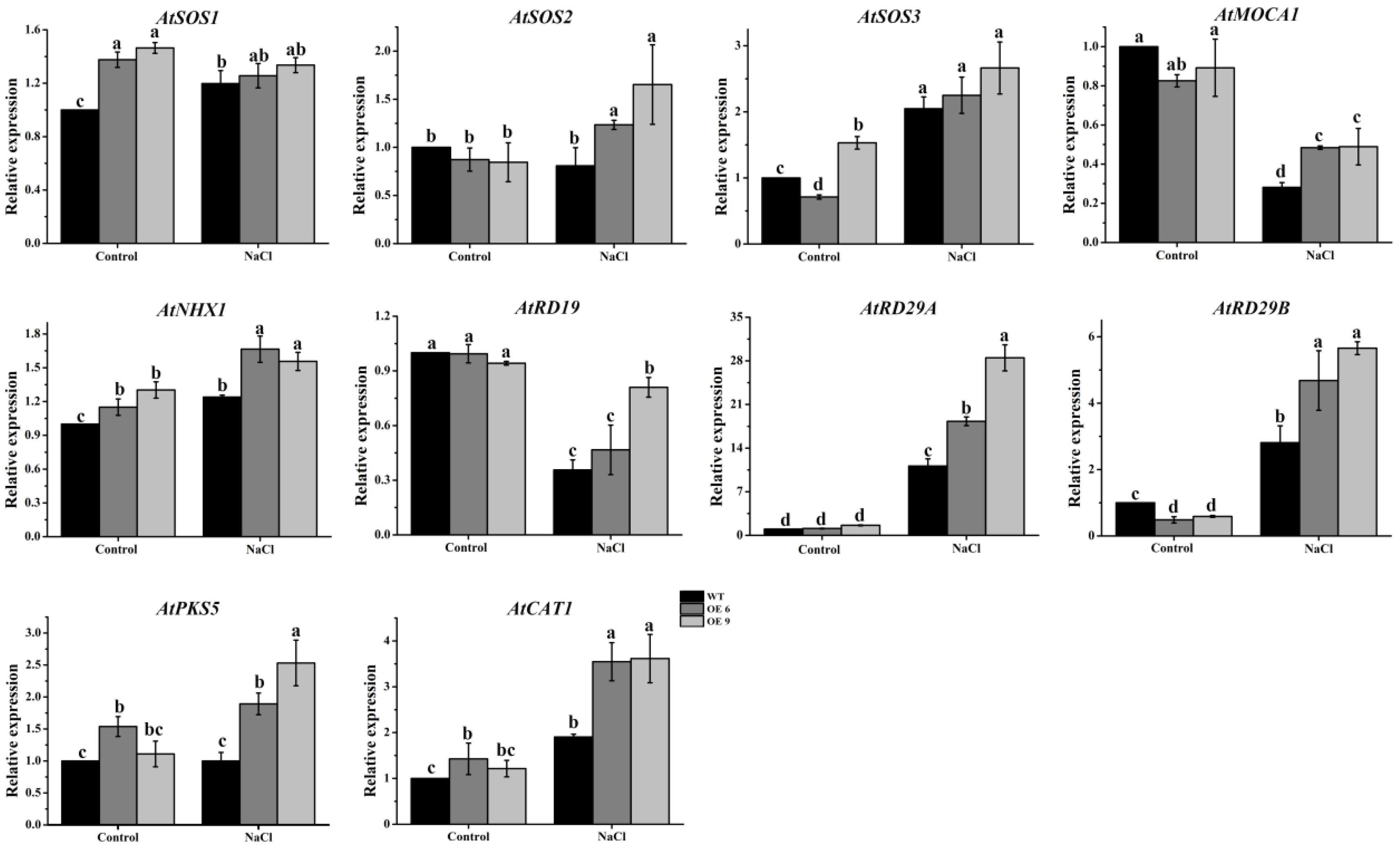
2.12. The Transcript Abundances of Multiple Salt Responsive Genes Increased in TaTIP4;1 OE Lines of Arabidopsis and Rice upon Salinity
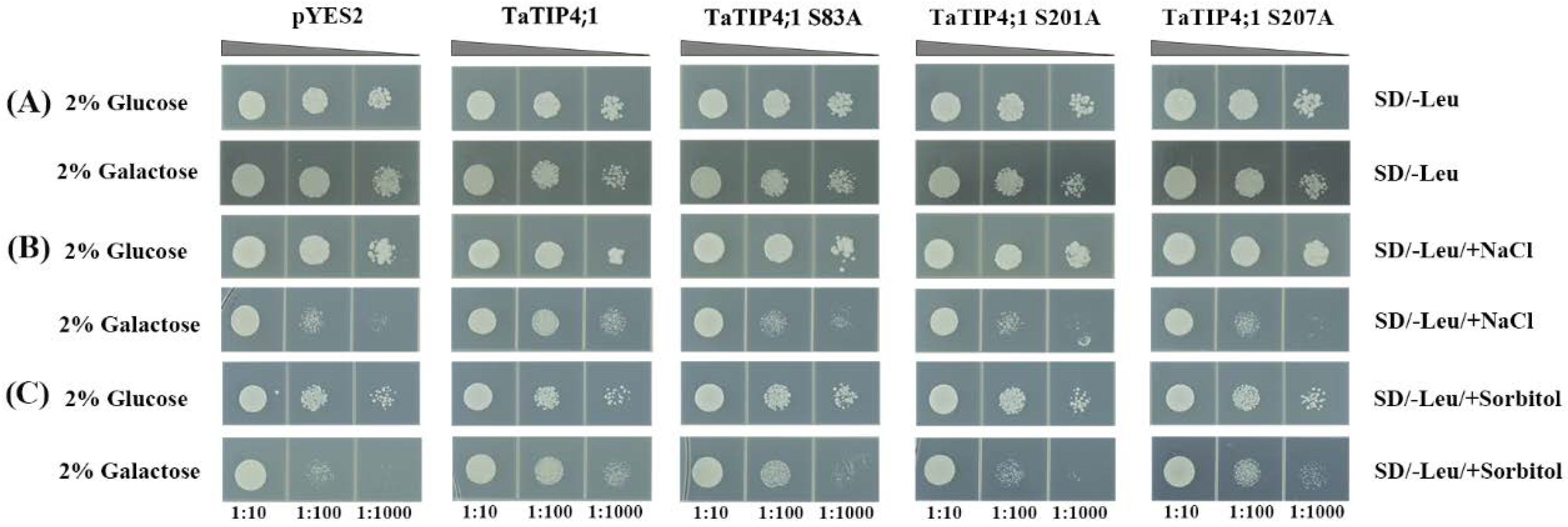
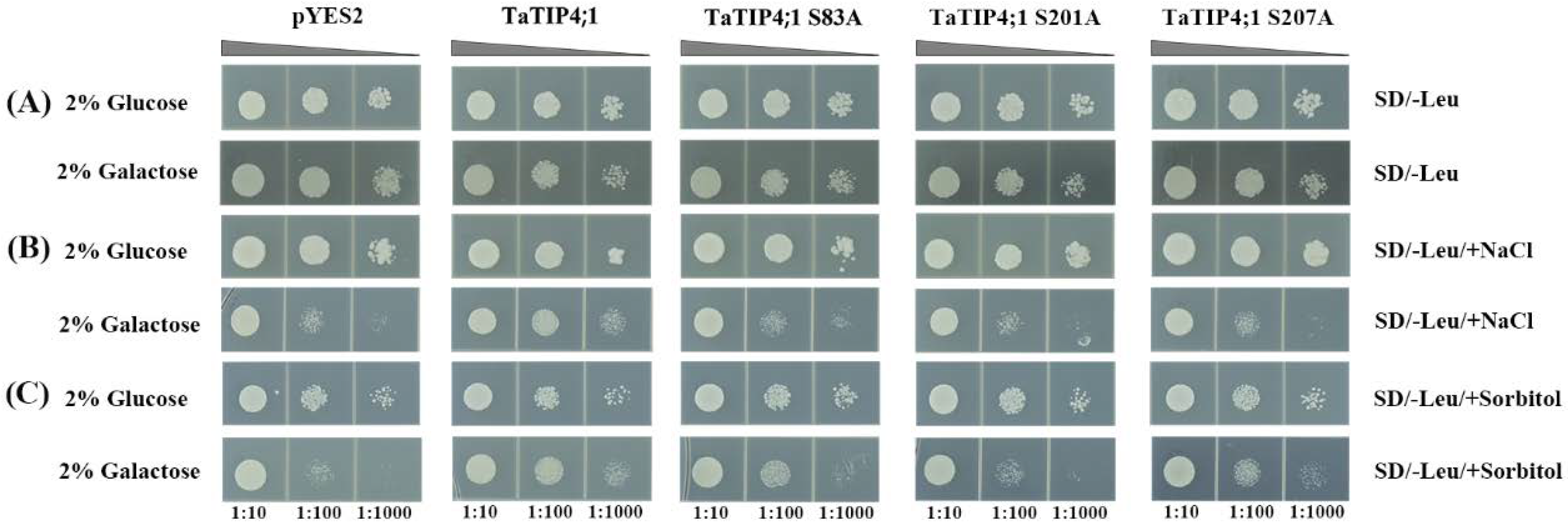
2.13. Phosphorylation Might Regulate TaTIP4;1 Activity in Adaptation to Salt and Osmotic Stress
3. Discussion
4. Materials and Methods
4.1. Analysis of TaTIP4;1 Expression in Wheat under Stresses
4.2. Generation of TaTIP4;1 Transgenic Lines of Arabidopsis and Rice
4.3. Performance Identification of Arabidopsis Overexpressors of TaTIP4;1 upon Drought Stress
4.4. Analysis of Seed Germination of Arabidopsis and Rice under Stresses
4.5. Analysis of Seedlings Growth of Arabidopsis and Rice upon Stresses
4.6. Measurement of Na+ and K+ Contents under Salt Stress
4.7. Detection of the Levels of H2O2 and MDA, the Activities of Antioxidases and Proline Accumulation upon Stresses
4.8. Gene Expression Analysis by qRT-PCR
4.9. Analysis of TaTIP4;1 Functions in Yeast
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2021, 23, 104–119. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Lv, Z.; Liu, Y.; Li, L.; Zhang, L. Network analysis of ABA-dependent and ABA-independent drought responsive genes in Arabidopsis thaliana. Genet. Mol. Biol. 2018, 41, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Soma, F.; Takahashi, F.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Cellular Phosphorylation Signaling and Gene Expression in Drought Stress Responses: ABA-Dependent and ABA-Independent Regulatory Systems. Plants 2021, 10, 756. [Google Scholar] [CrossRef]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [Green Version]
- Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms. Int. J. Mol. Sci. 2015, 16, 15811–15851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Laxmi, A. Transcriptional regulation of drought response: A tortuous network of transcriptional factors. Front. Plant Sci. 2015, 6, 895. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.; Wani, S.; Singh, B.; Bohra, A.; Dar, Z.; Lone, A.; Pareek, A.; Singla-Pareek, S.L. Transcription Factors and Plants Response to Drought Stress: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Mao, C.; Xi, D.; Ming, F. The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 40641. [Google Scholar] [CrossRef]
- Mushtaq, Z.; Faizan, S.; Gulzar, B. Salt stress, its impacts on plants and the strategies plants are employing against it: A review. J. Appl. Biol. Biotechnol. 2020, 8, 81–91. [Google Scholar]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Liu, F.; Sun, L.; Hao, F. Versatile Roles of Aquaporins in Plant Growth and Development. Int. J. Mol. Sci. 2020, 21, 9485. [Google Scholar] [CrossRef]
- Tyerman, S.D.; McGaughey, S.A.; Qiu, J.; Yool, A.J.; Byrt, C.S. Adaptable and Multifunctional Ion-Conducting Aquaporins. Annu. Rev. Plant Biol. 2021, 72, 703–736. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The Roles of Aquaporins in Plant Stress Responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Johanson, U.; Karlsson, M.; Johansson, I.; Gustavsson, S.; Sjovall, S.; Fraysse, L.; Weig, A.R.; Kjellbom, P. The Complete Set of Genes Encoding Major Intrinsic Proteins in Arabidopsis Provides a Framework for a New Nomenclature for Major Intrinsic Proteins in Plants. Plant Physiol. 2001, 126, 1358–1369. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 Rice Aquaporin Genes and Analysis of Their Expression and Function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrest, K.L.; Bhave, M. The PIP and TIP aquaporins in wheat form a large and diverse family with unique gene structures and functionally important features. Funct. Integr. Genom. 2007, 8, 115–133. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, M.M. TIP aquaporins inplants: Role in abiotic stress tolerance. In Abiotic Stress in Plants; IntechOpen: London, UK, 2020. [Google Scholar]
- Gautam, A.; Pandey, A.K. Aquaporins Responses under Challenging Environmental Conditions and Abiotic Stress Tolerance in Plants. Bot. Rev. 2021, 87, 467–495. [Google Scholar] [CrossRef]
- Peng, Y.; Lin, W.; Cai, W.; Arora, R. Overexpression of a Panax ginseng tonoplast aquaporin alters salt tolerance, drought tolerance and cold acclimation ability in transgenic Arabidopsis plants. Planta 2007, 226, 729–740. [Google Scholar] [CrossRef]
- Li, J.; Cai, W. A ginseng PgTIP1 gene whose protein biological activity related to Ser128 residue confers faster growth and enhanced salt stress tolerance in Arabidopsis. Plant Sci. 2015, 234, 74–85. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Hu, Z.; Che, B.; Chen, H.; Yu, B.; Cai, W. Heterologous Expression of Panax ginseng PgTIP1 Confers Enhanced Salt Tolerance of Soybean Cotyledon Hairy Roots, Composite, and Whole Plants. Front. Plant Sci. 2017, 8, 1232. [Google Scholar] [CrossRef] [Green Version]
- An, J.; Cheng, C.; Hu, Z.; Chen, H.; Cai, W.; Yu, B. The Panax ginseng PgTIP1 gene confers enhanced salt and drought tolerance to transgenic soybean plants by maintaining homeostasis of water, salt ions and ROS. Environ. Exp. Bot. 2018, 155, 45–55. [Google Scholar] [CrossRef]
- Khan, K.; Agarwal, P.; Shanware, A.; Sane, V.A. Heterologous Expression of Two Jatropha Aquaporins Imparts Drought and Salt Tolerance and Improves Seed Viability in Transgenic Arabidopsis thaliana. PLoS ONE 2015, 10, e0128866. [Google Scholar] [CrossRef]
- Wang, L.-L.; Chen, A.-P.; Zhong, N.-Q.; Liu, N.; Wu, X.-M.; Wang, F.; Yang, C.-L.; Romero, M.F.; Xia, G.-X. The Thellungiella salsuginea Tonoplast Aquaporin TsTIP1;2 Functions in Protection Against Multiple Abiotic Stresses. Plant Cell Physiol. 2014, 55, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yu, G.; Han, X.; Xin, S.; Qiang, X.; Jiang, L.; Zhang, S.; Cheng, X. TsMIP6 enhances the tolerance of transgenic rice to salt stress and interacts with target proteins. J. Plant Biol. 2015, 58, 285–292. [Google Scholar] [CrossRef]
- Sade, N.; Vinocur, B.J.; Diber, A.; Shatil, A.; Ronen, G.; Nissan, H.; Wallach, R.; Karchi, H.; Moshelion, M. Improving plant stress tolerance and yield production: Is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion? New Phytol. 2009, 181, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-Y.; Kumar, M.; Xu, L.; Wan, Q.; Huang, Y.-H.; Xu, Z.-L.; He, X.-L.; Ma, J.-B.; Pandey, G.K.; Shao, H.-B. Genome-wide identification of Major Intrinsic Proteins in Glycine soja and characterization of GmTIP2;1 function under salt and water stress. Sci. Rep. 2017, 7, 4106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Peng, Y.; Li, G.; Arora, R.; Tang, Z.; Su, W.; Cai, W. Isolation and functional characterization of PgTIP1, a hormone-autotrophic cells-specific tonoplast aquaporin in ginseng. J. Exp. Bot. 2007, 58, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.-J.; Xu, S.-C.; Liu, N.; Zhang, G.-W.; Hu, Q.-Z.; Xu, Z.-S.; Gong, Y.-M. Identification of the AQP members involved in abiotic stress responses from Arabidopsis. Gene 2018, 646, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Fröhlich, E.; Ligr, M.; Grey, M.; Sigrist, S.J.; Wolf, D.H.; Fröhlich, K.-U. Oxygen Stress: A Regulator of Apoptosis in Yeast. J. Cell Biol. 1999, 145, 757–767. [Google Scholar] [CrossRef]
- Gao, C.; Wang, Y.; Jiang, B.; Liu, G.; Yu, L.; Wei, Z.; Yang, C. A novel vacuolar membrane H+-ATPase c subunit gene (ThVHAc1) from Tamarix hispida confers tolerance to several abiotic stresses in Saccharomyces cerevisiae. Mol. Biol. Rep. 2011, 38, 957–963. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, Y.; An, Y.; Wu, J.; He, S.; Sun, L.; Hao, F. Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice. Int. J. Mol. Sci. 2022, 23, 2085. https://doi.org/10.3390/ijms23042085
Wang Y, Zhang Y, An Y, Wu J, He S, Sun L, Hao F. Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice. International Journal of Molecular Sciences. 2022; 23(4):2085. https://doi.org/10.3390/ijms23042085
Chicago/Turabian StyleWang, Yan, Yaqi Zhang, Yinchao An, Jingyuan Wu, Shibin He, Lirong Sun, and Fushun Hao. 2022. "Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice" International Journal of Molecular Sciences 23, no. 4: 2085. https://doi.org/10.3390/ijms23042085
APA StyleWang, Y., Zhang, Y., An, Y., Wu, J., He, S., Sun, L., & Hao, F. (2022). Wheat TaTIP4;1 Confers Enhanced Tolerance to Drought, Salt and Osmotic Stress in Arabidopsis and Rice. International Journal of Molecular Sciences, 23(4), 2085. https://doi.org/10.3390/ijms23042085