
Effect of Curcumin in Experimental Pulmonary Tuberculosis: Antimycobacterial Activity in the Lungs and Anti-Inflammatory Effect in the Brain

 ,
, 
 ,
,  ,
, 

Abstract
1. Introduction
2. Results
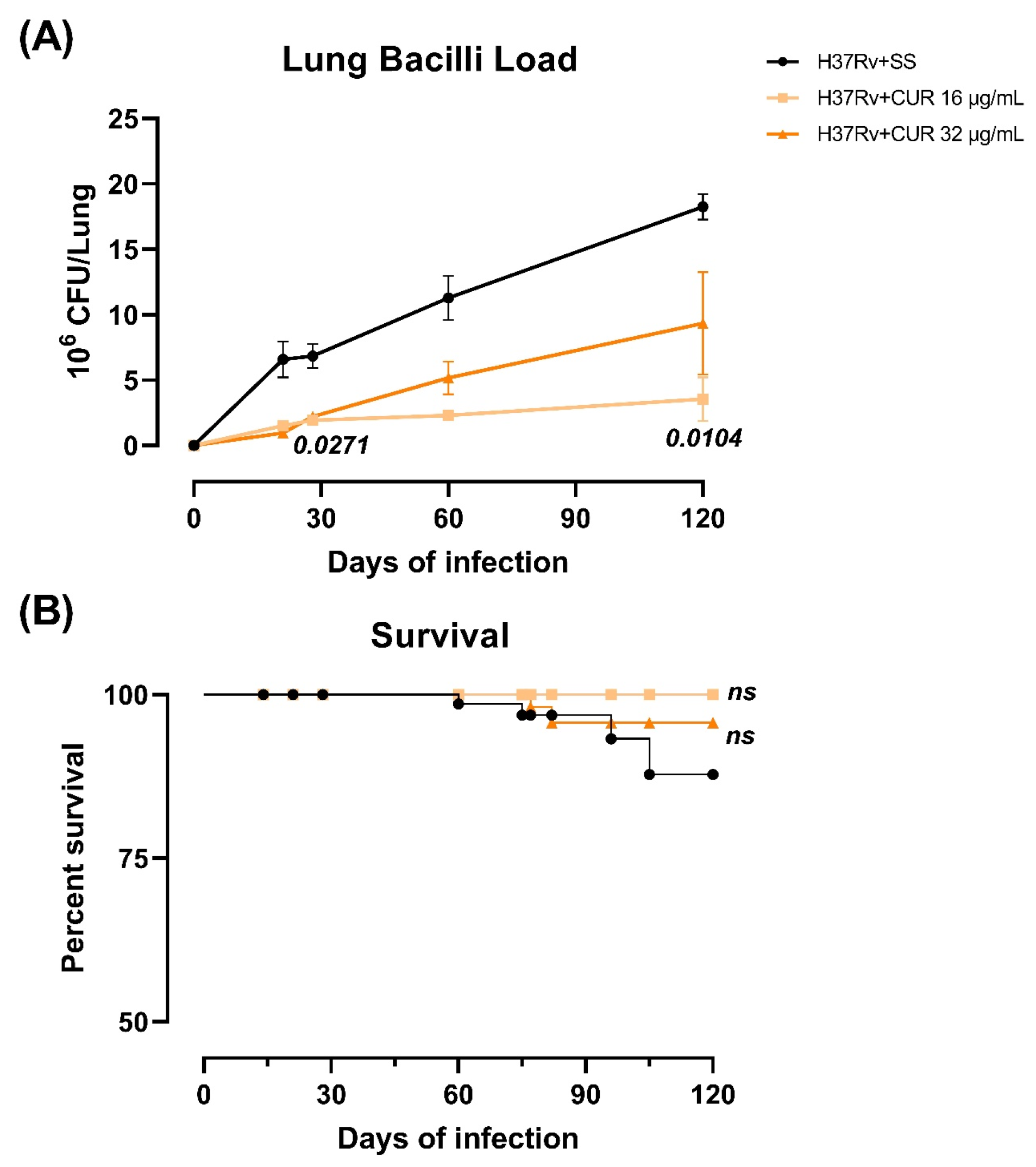
2.1. The Effect of Curcumin (CUR) Treatment on Survival, Bacilli Loads and Tissue Damage (Pneumonia) in Experimental Pulmonary Tuberculosis
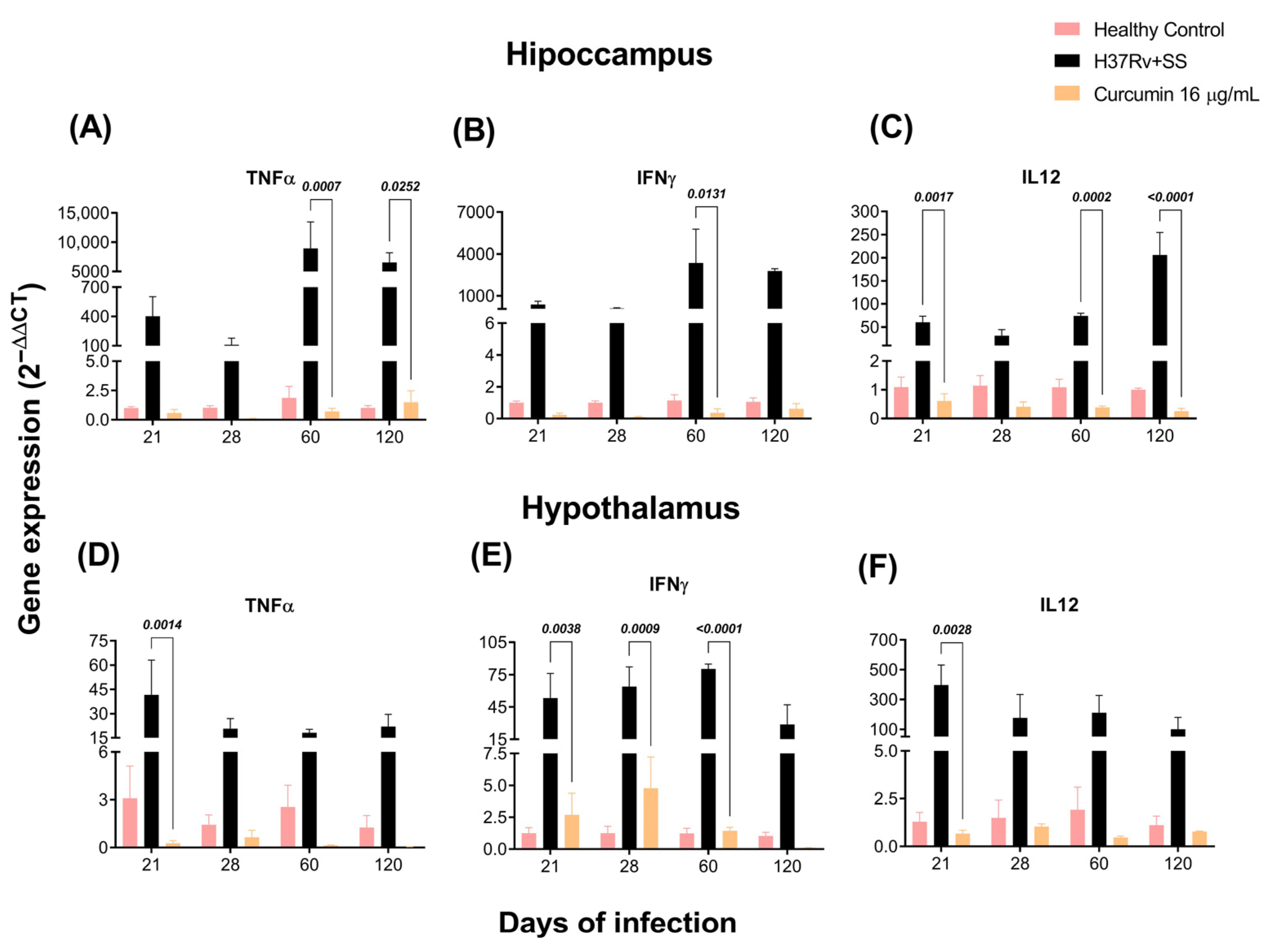
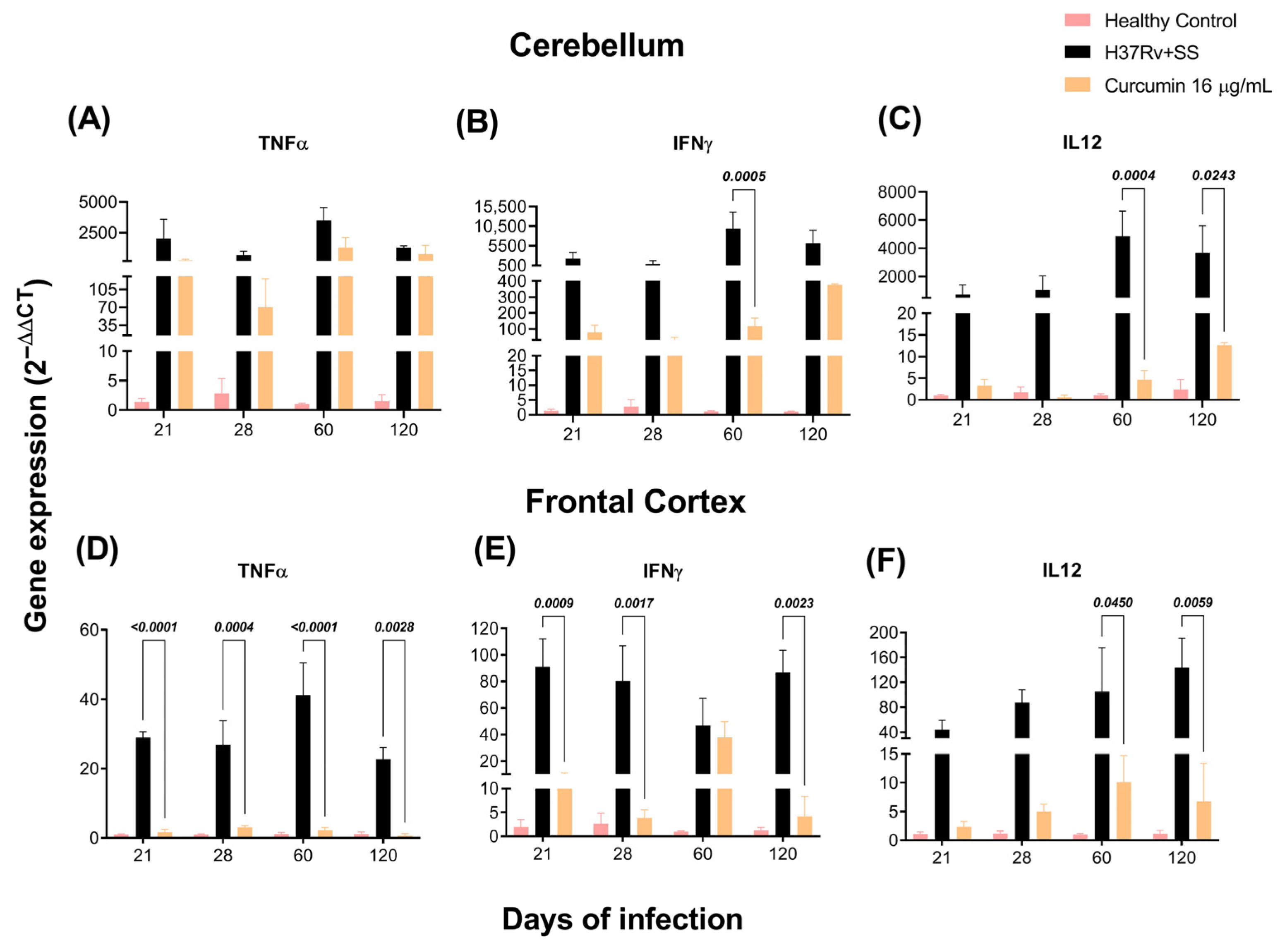
2.2. The Effect of CUR Treatment on Cytokine Expression in Distinct Brain Areas of TB Mice
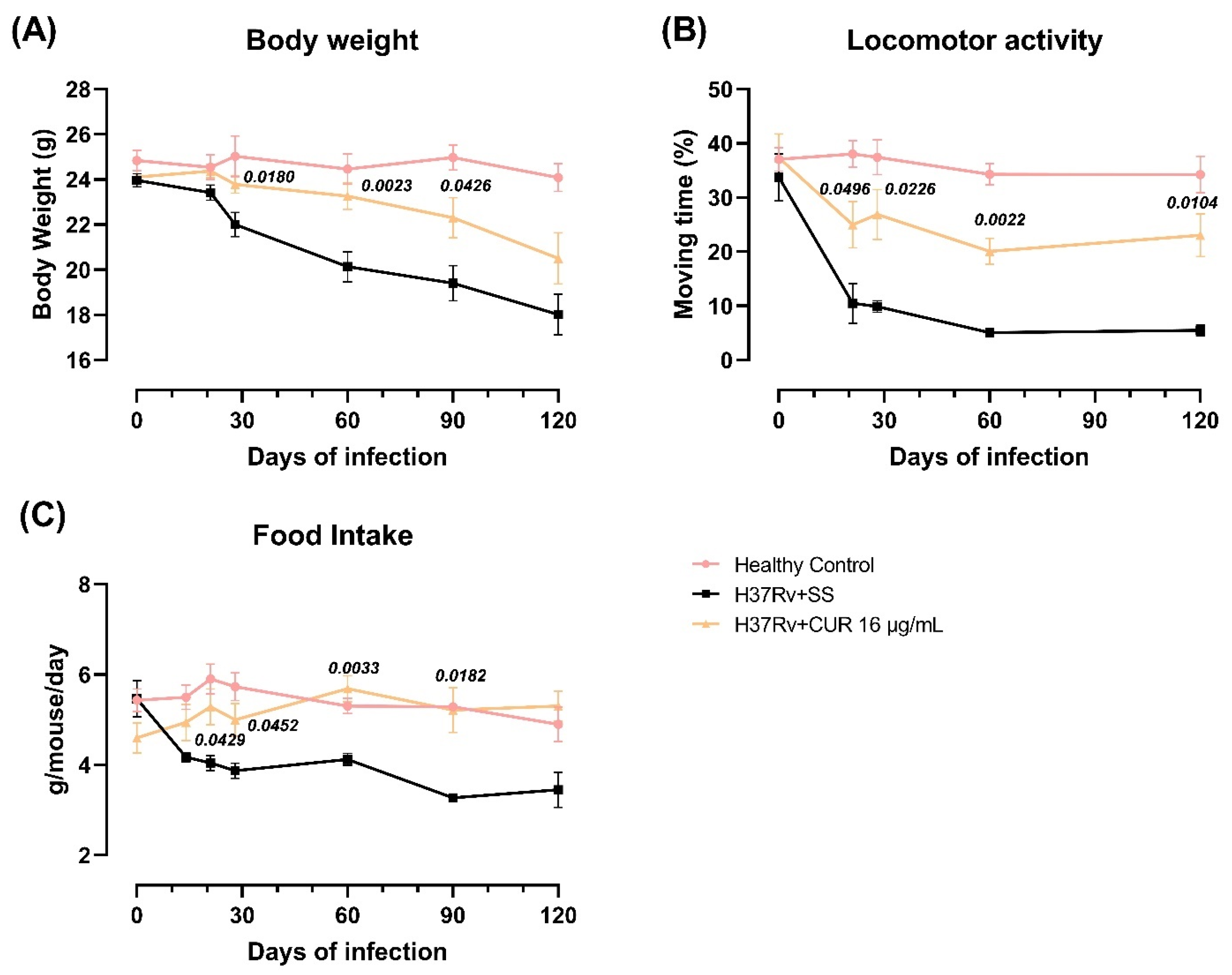
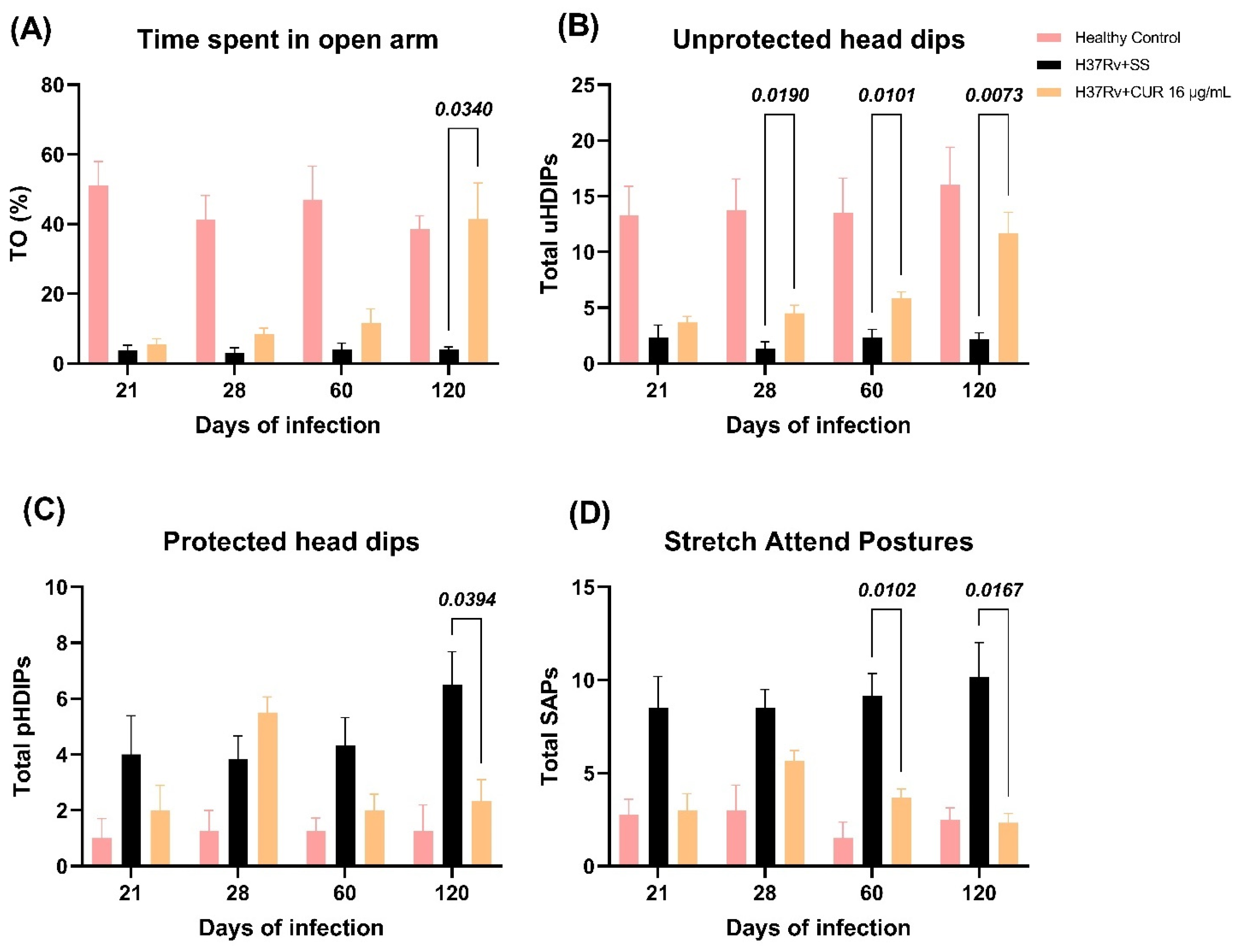
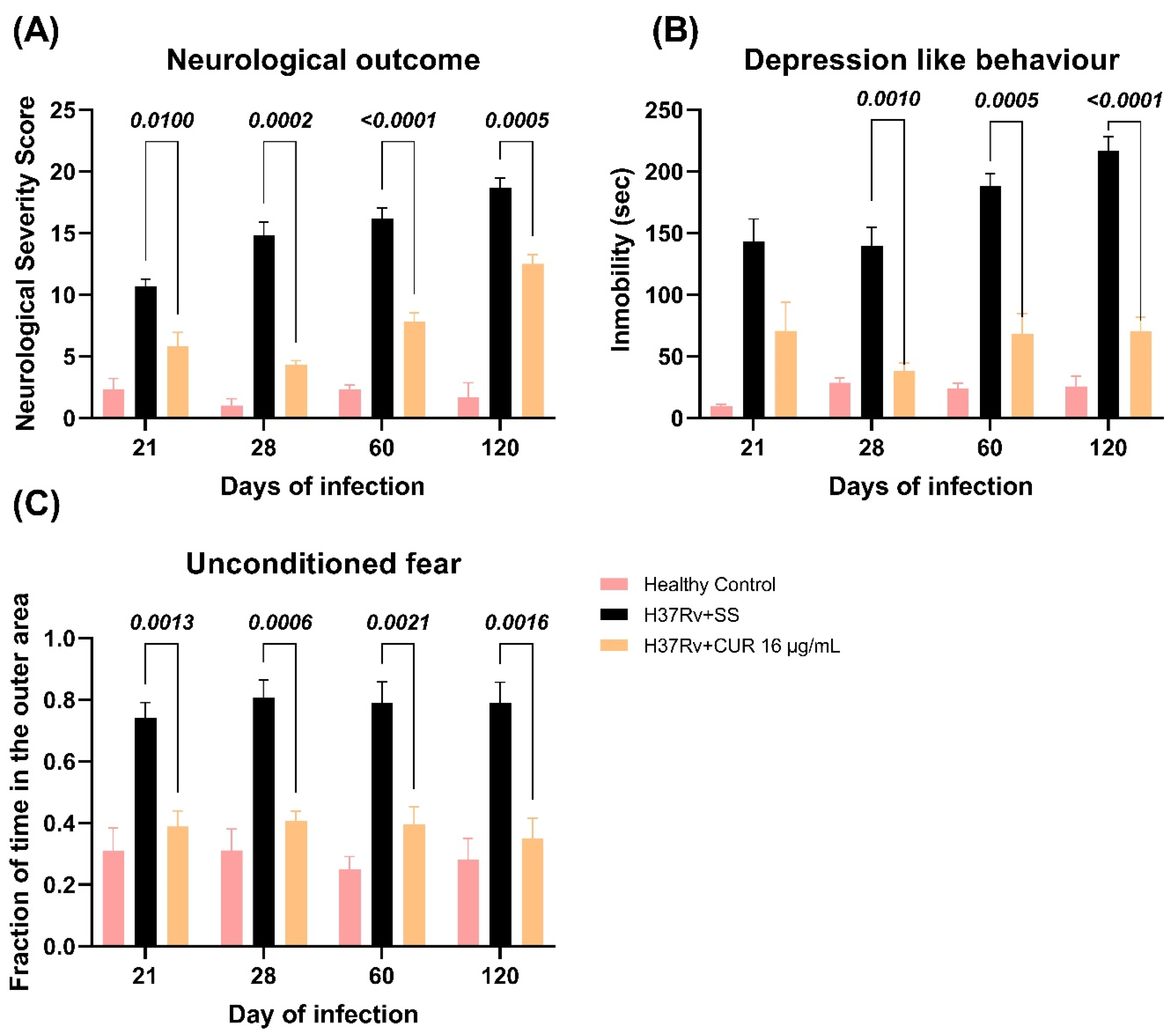
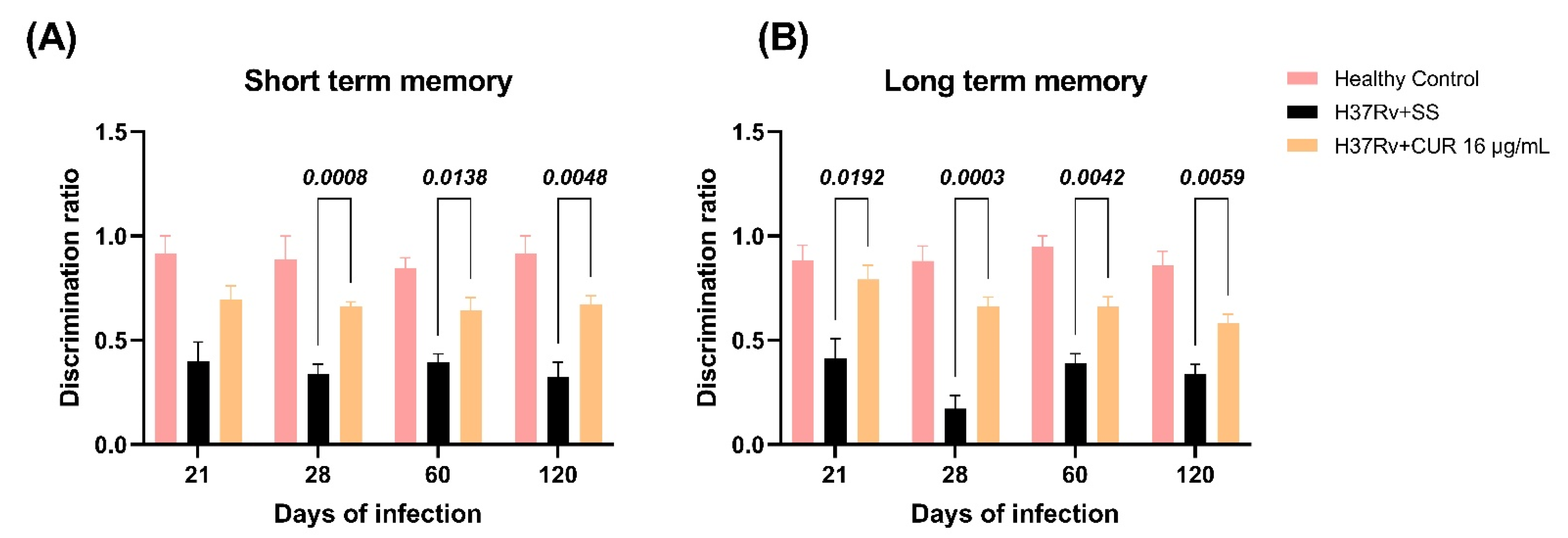
2.3. The Effect of CUR Treatment after Early TB Infection on Diverse Behavioral Abnormalities
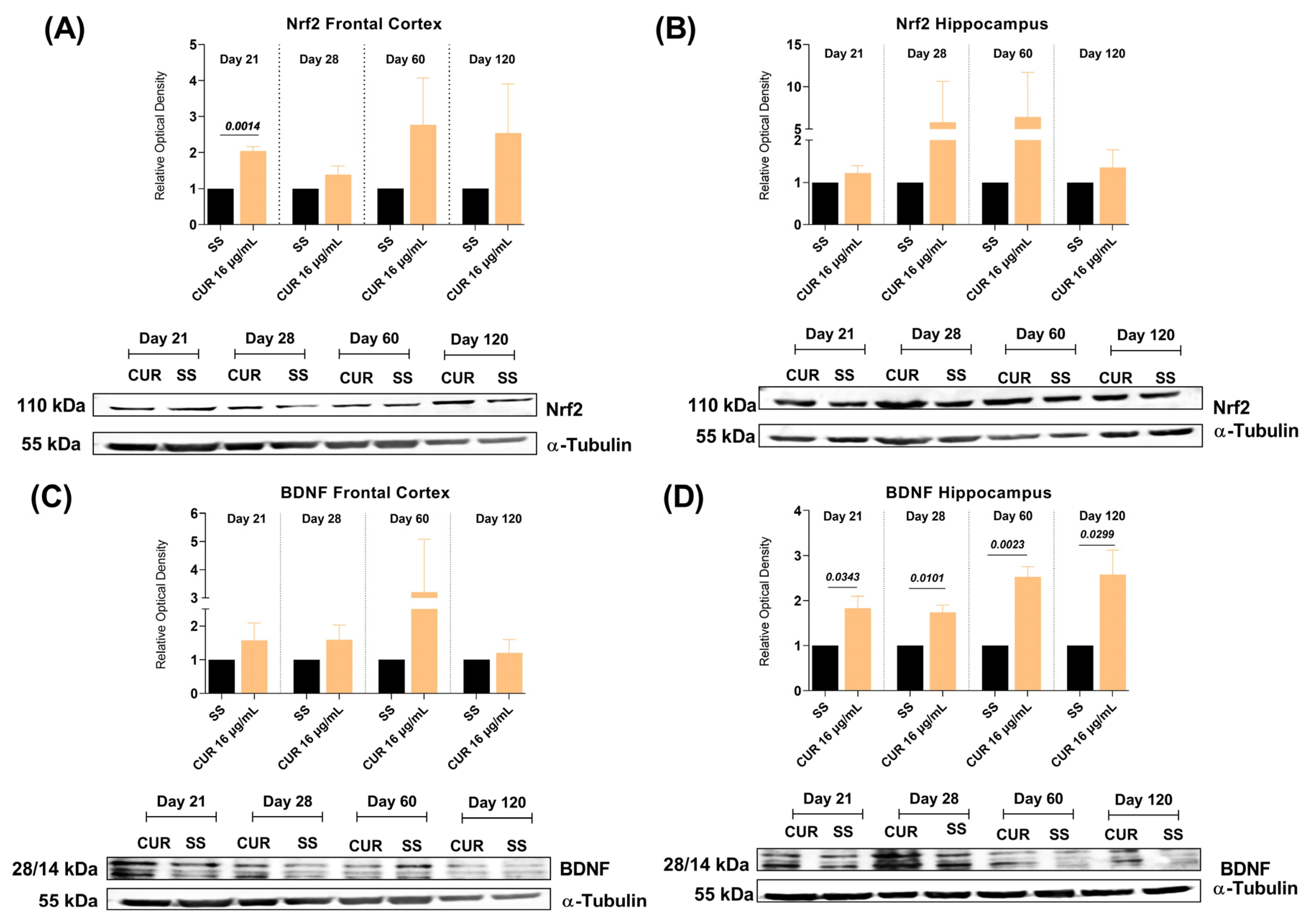
2.4. The Effect of CUR Treatment on Nrf2 and BDNF Levels in the Frontal Cortex and Hippocampus of TB MICE
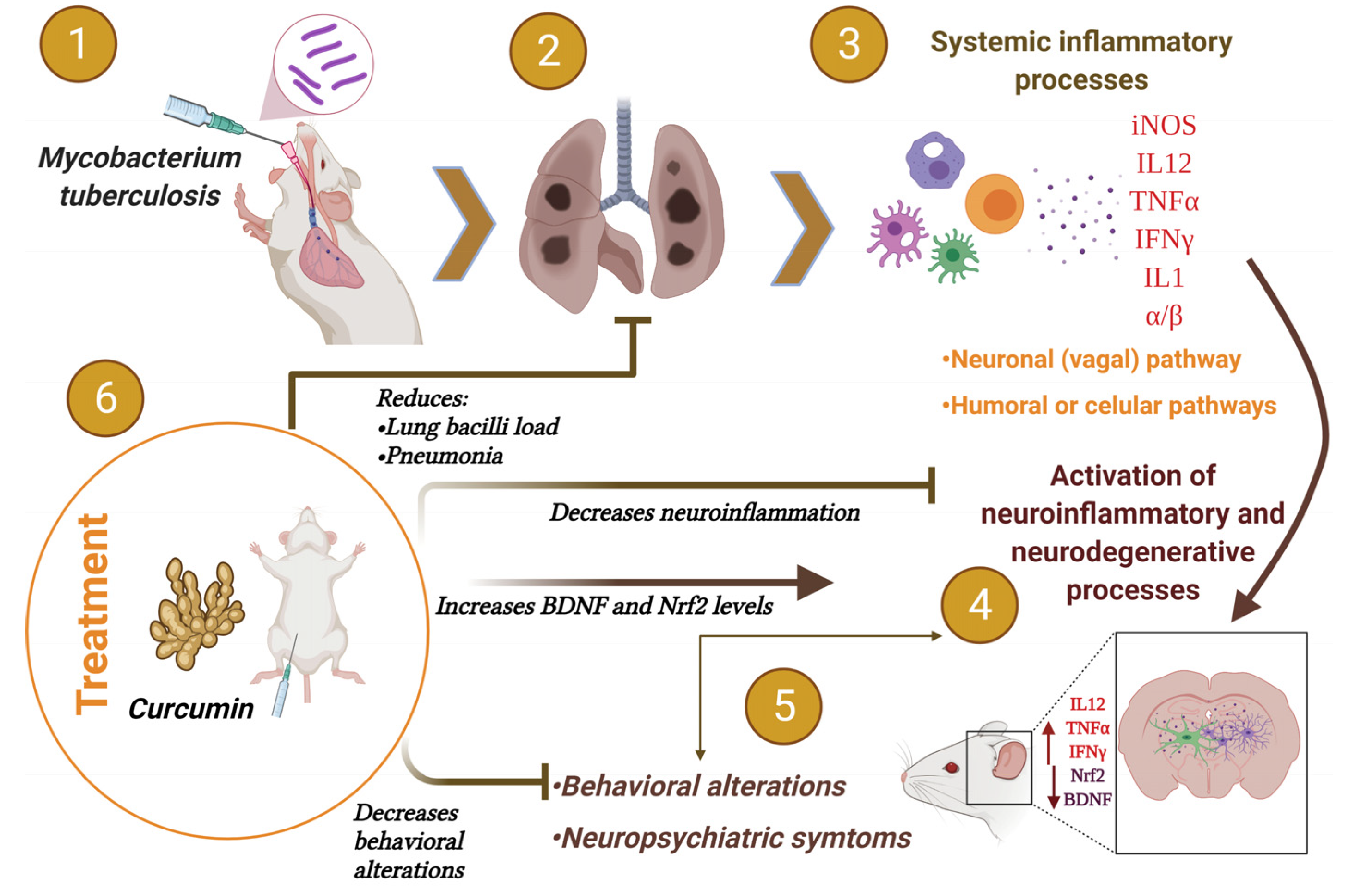
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Animals
4.3. The Experimental Murine Model of Pulmonary TB
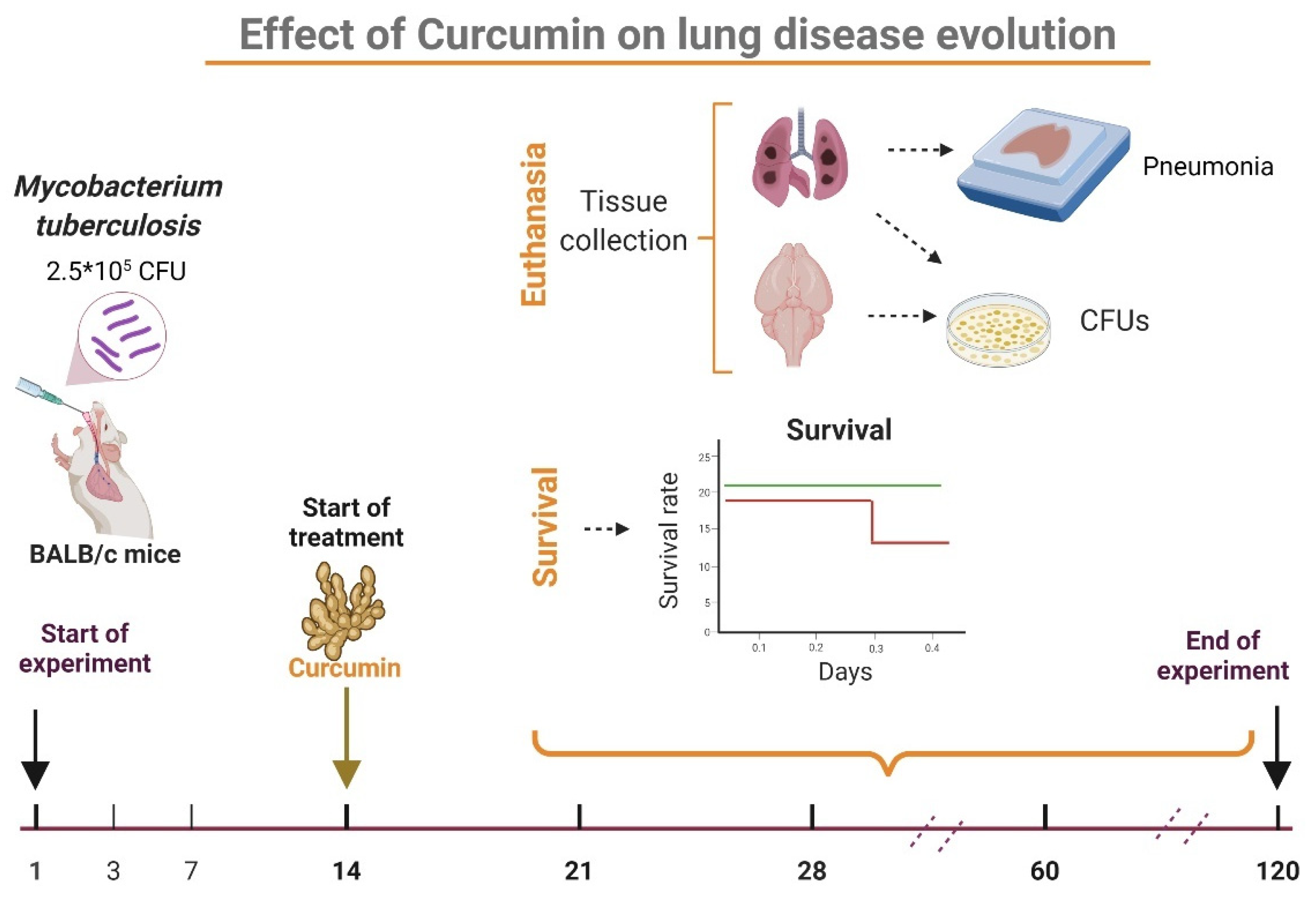
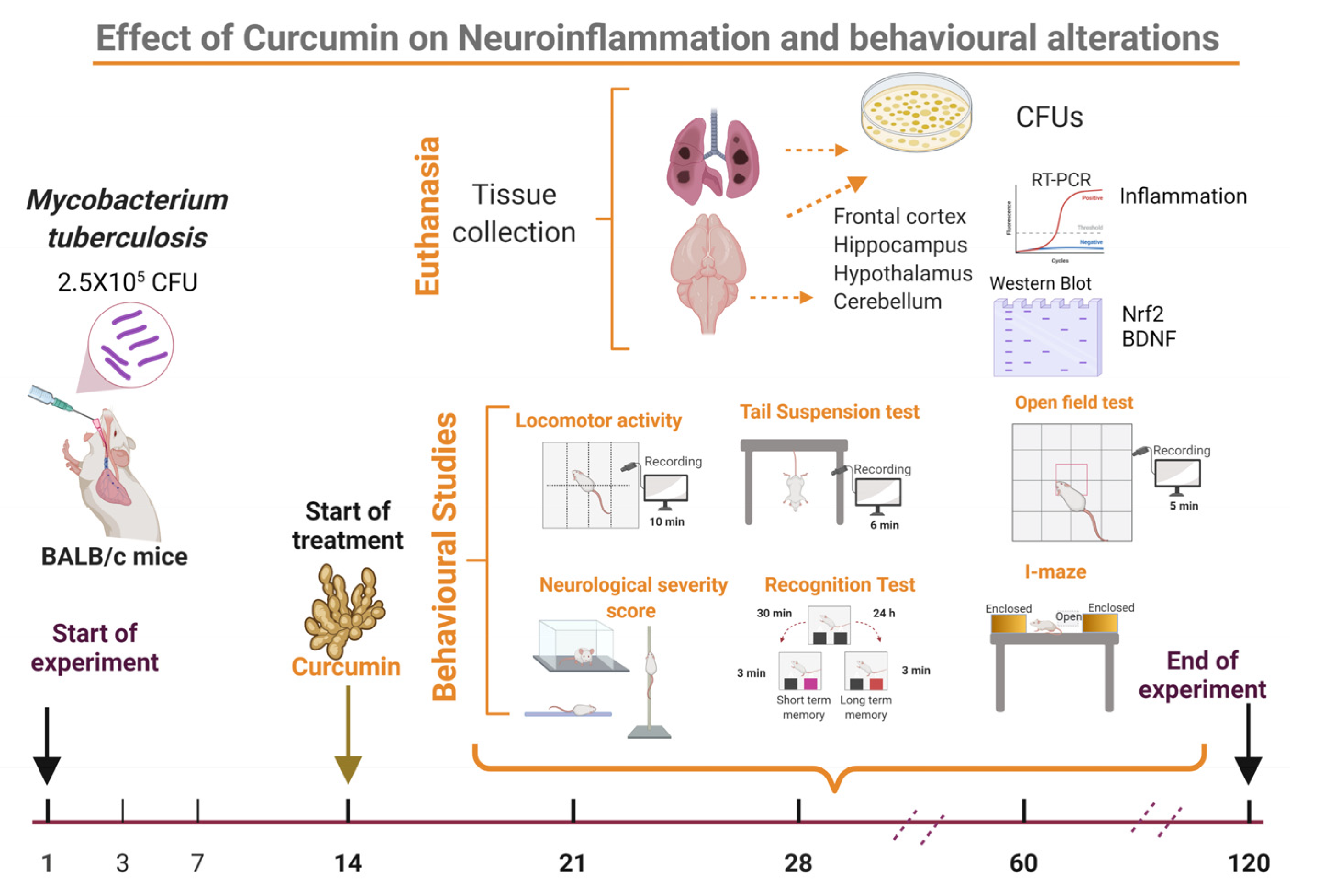
4.4. Experimental Design
4.5. Curcumin Preparation and Administration
4.6. Colony-Forming Units (CFU) in Lungs and Brain of TB Mice
4.7. Determination of Lung Affected Area by Pneumonia
4.8. Expression of Cytokine Determined by RT-PCR
4.9. Study of Nrf2 and BDNF by Western Blot Assay
4.10. Behavior Tests in Infected TB Mice
4.10.1. Sickness Behavior Study
4.10.2. Depression-like Behavior Study
4.10.3. Anxiety-like Behavior Study
4.10.4. Unconditioned Fear Study
4.10.5. Neurological Outcome Study
4.10.6. Memory Damage Study
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, I. Mycobacterium tuberculosis Pathogenesis and Molecular Determinants of Virulence Mycobacterium tuberculosis Pathogenesis and Molecular Determinants of Virulence. Society 2003, 16, 463–496. [Google Scholar] [CrossRef]
- Loddenkemper, R.; Lipman, M.; Zumla, A. Clinical aspects of adult tuberculosis. Cold Spring Harb. Perspect. Med. 2016, 6, a017848. [Google Scholar] [CrossRef] [PubMed]
- Stockdale, L.; Fletcher, H. The Future of Vaccines for Tuberculosis. Clin. Chest Med. 2019, 40, 849–856. [Google Scholar] [CrossRef]
- Schrager, L.K.; Vekemens, J.; Drager, N.; Lewinsohn, D.M.; Olesen, O.F. The status of tuberculosis vaccine development. Lancet Infect. Dis. 2020, 20, e28–e37. [Google Scholar] [CrossRef]
- Zatarain-Barrón, Z.L.; Ramos-Espinosa, O.; Marquina-Castillo, B.; Barrios-Payán, J.; Cornejo-Granados, F.; Maya-Lucas, O.; López-Leal, G.; Molina-Romero, C.; Anthony, R.M.; Ochoa-Leyva, A.; et al. Evidence for the Effect of Vaccination on Host-Pathogen Interactions in a Murine Model of Pulmonary Tuberculosis by Mycobacterium tuberculosis. Front. Immunol. 2020, 11, 930. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Jain, V.K.; Iyengar, K.P.; Samy, D.A.; Vaishya, R. Tuberculosis in the era of COVID-19 in India. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 1439–1443. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Treatment of Drug-Susceptible Tuberculosis and Patient Care, 2017 Update; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Treatment of Drug-Susceptible Tuberculosis: Rapid Communication; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Arbex, M.A.; Varella, M.d.C.L.; de Siqueira, H.R.; de Mello, F.A.F. Antituberculosis drugs: Drug interactions, adverse effects, and use in special situations. Part 2: Second-line drugs. J. Bras. Pneumol. 2010, 36, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Forget, E.J.; Menzies, D. Adverse reactions to first-time antituberculosis drugs. Expert Opin. Drug Saf. 2006, 5, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Nath, H.; Ryoo, S. First– and Second–Line Drugs and Drug Resistance. Tuberc. Curr. Issues Diagn. Manag. 2013, 20, 163–180. [Google Scholar] [CrossRef]
- O’Donnell, M.R.; Daftary, A.; Frick, M.; Hirsch-Moverman, Y.; Amico, K.R.; Senthilingam, M.; Wolf, A.; Metcalfe, J.Z.; Isaakidis, P.; Davis, J.L.; et al. Re-inventing adherence: Toward a patient-centered model of care for drug-resistant tuberculosis and HIV. Int. J. Tuberc. Lung Dis. 2016, 20, 430–434. [Google Scholar] [CrossRef]
- World Health Organization. WHO Operational Handbook on Tuberculosis. Module 4: Treatment—Drug-Resistant Tuberculosis Treatment; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- García, A.; Bocanegra-García, V.; Palma-Nicolás, J.P.; Rivera, G. Recent advances in antitubercular natural products. Eur. J. Med. Chem. 2012, 49, 1–23. [Google Scholar] [CrossRef]
- Rodriguez-Takeuchi, S.Y.; Renjifo, M.E.; Medina, F.J. Extrapulmonary tuberculosis: Pathophysiology and imaging findings. RadioGraphics 2019, 39, 2023–2037. [Google Scholar] [CrossRef]
- Sherwood, E.R.; Toliver-Kinsky, T. Mechanisms of the inflammatory response. Best Pract. Res. Clin. Anaesthesiol. 2004, 18, 385–405. [Google Scholar] [CrossRef]
- Xu, W.; Larbi, A. Immunity and Inflammation: From Jekyll to Hyde. Exp. Gerontol. 2018, 107, 98–101. [Google Scholar] [CrossRef]
- Liu, C.H.; Liu, H.; Ge, B. Innate immunity in tuberculosis: Host defense vs pathogen evasion. Cell. Mol. Immunol. 2017, 14, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Sia, J.K.; Rengarajan, J. Immunology of mycobacterium tuberculosis infections. Gram-Posit. Pathog. 2019, 7, 1056–1086. [Google Scholar] [CrossRef]
- Amaral, E.P.; Vinhaes, C.L.; Oliveira-De-Souza, D.; Nogueira, B.; Akrami, K.M.; Andrade, B.B. The Interplay between Systemic Inflammation, Oxidative Stress, and Tissue Remodeling in Tuberculosis. Antioxid. Redox Signal. 2021, 34, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Mrdjen, D.; Pavlovic, A.; Hartmann, F.J.; Schreiner, B.; Utz, S.G.; Leung, B.P.; Lelios, I.; Heppner, F.L.; Kipnis, J.; Merkler, D.; et al. High-Dimensional Single-Cell Mapping of Central Nervous System Immune Cells Reveals Distinct Myeloid Subsets in Health, Aging, and Disease. Immunity 2018, 48, 380–395.e6. [Google Scholar] [CrossRef]
- Klein, R.S.; Garber, C.; Howard, N. Infectious immunity in the central nervous system and brain function. Nat. Immunol. 2017, 18, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation pathways: A general review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Rhie, S.J.; Jung, E.Y.; Shim, I. The role of neuroinflammation on pathogenesis of affective disorders. J. Exerc. Rehabil. 2020, 16, 2–9. [Google Scholar] [CrossRef]
- Rajeswari, R.; Muniyandi, M.; Balasubramanian, R.; Narayanan, P.R. Perceptions of tuberculosis patients about their physical, mental and social well-being: A field report from south India. Soc. Sci. Med. 2005, 60, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Van Den Heuvel, L.; Chishinga, N.; Kinyanda, E.; Weiss, H.; Patel, V.; Ayles, H.; Harvey, J.; Cloete, K.J.; Seedat, S. Frequency and correlates of anxiety and mood disorders among TB- and HIV-infected Zambians. AIDS Care Psychol. Socio-Med. Asp. AIDS/HIV 2013, 25, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Feng, L.; Feng, L.; Nyunt, M.S.; Yap, K.B.; Ng, T.P. Systemic inflammation, depression and obstructive pulmonary function: A population-based study. Respir. Res. 2013, 14, 53. [Google Scholar] [CrossRef]
- Wells, K.B.; Golding, J.M.; Burnam, M.A. Psychiatric disorder in a sample of the general population with and without chronic medical conditions. Am. J. Psychiatry 1988, 145, 976–981. [Google Scholar] [CrossRef]
- Lara-Espinosa, J.V.; Hernández-Pando, R. Psychiatric Problems in Pulmonary Tuberculosis: Depression and Anxiety. J. Tuberc. Res. 2021, 9, 31–50. [Google Scholar] [CrossRef]
- Javaid, A.; Mehreen, S.; Khan, M.A.; Ashiq, N.; Ihtesham, M. Depression and its Associated Factors with Multidrug-Resistant Tuberculosis at Baseline. J. Depress. Anxiety 2017, 6, 1–6. [Google Scholar] [CrossRef]
- Peltzer, K.; Naidoo, P.; Matseke, G.; Louw, J.; Mchunu, G.; Tutshana, B. Prevalence of psychological distress and associated factors in tuberculosis patients in public primary care clinics in South Africa. BMC Psychiatry 2012, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Shyamala, K.K.; Sharadha Naveen, R.; Khatri, B. Depression: A neglected comorbidity in patients with tuberculosis. J. Assoc. Physicians India 2018, 66, 18–21. [Google Scholar]
- De Castro-Silva, K.M.; Carvalho, A.C.; Cavalcanti, M.T.; Da Martins, P.S.; França, J.R.; Oquendo, M.; Kritski, A.L.; Sweetland, A. Prevalence of depression among patients with presumptive pulmonary tuberculosis in Rio de Janeiro, Brazil. Braz. J. Psychiatry 2019, 41, 316–323. [Google Scholar] [CrossRef]
- Singh, L.; Pardal, P.; Prakash, J. Psychiatric morbidity in patients of pulmonary tuberculosis-an observational study. Ind. Psychiatry J. 2015, 24, 168. [Google Scholar] [CrossRef] [PubMed]
- Kruijshaar, M.E.; Lipman, M.; Essink-Bot, M.L.; Lozewicz, S.; Creer, D.; Dart, S.; Maguire, H.; Abubakar, I. Health status of UK patients with active tuberculosis. Int. J. Tuberc. Lung Dis. 2010, 14, 296–302. [Google Scholar] [PubMed]
- Rubeen, R.; Zareen, N.; Zameer, S.; Rasool, A.G.; Naqvi, S.S.N.; Iqbal, J. Anxiety and Depression in Tuberculosis Can Create Impact on Quality of Life of Patient. Acta Med. Int. 2014, 1, 93–98. [Google Scholar] [CrossRef]
- Kumar, K.; Kumar, A.; Chandra, P.; Kansal, H. A study of prevalence of depression and anxiety in patients suffering from tuberculosis. J. Fam. Med. Prim. Care 2016, 5, 150. [Google Scholar] [CrossRef] [PubMed]
- Aghanwa, H.S.; Erhabor, G.E. Demographic/socioeconomic factors in mental disorders associated with tuberculosis in southwest Nigeria. J. Psychosom. Res. 1998, 45, 353–360. [Google Scholar] [CrossRef]
- Vega, P.; Sweetland, A.; Acha, J.; Castillo, H.; Guerra, D.; Fawzi, M.C.S.; Shin, S. Psychiatric issues in the management of patients with multidrug-resistant tuberculosis. Int. J. Tuberc. Lung Dis. 2004, 8, 749–759. [Google Scholar]
- Walker, I.F.; Khan, A.M.; Khan, A.M.; Khan, N.M.; Ayub, R.M.; Ghias, K.N.; Walley, J.D. Depression among multidrug-resistant tuberculosis patients in Punjab, Pakistan: A large cross-sectional study. Int. J. Tuberc. Lung Dis. 2018, 22, 773–778. [Google Scholar] [CrossRef]
- Yen, Y.F.; Chung, M.S.; Hu, H.Y.; Lai, Y.J.; Huang, L.Y.; Lin, Y.S.; Chou, P.; Deng, C.Y. Association of pulmonary tuberculosis and ethambutol with incident depressive disorder: A nationwide, population-based cohort study. J. Clin. Psychiatry 2015, 76, e505–e511. [Google Scholar] [CrossRef]
- Romeo, H.E.; Tio, D.L.; Rahman, S.U.; Chiappelli, F.; Taylor, A.N. The glossopharyngeal nerve as a novel pathway in immune-to-brain communication: Relevance to neuroimmune surveillance of the oral cavity. J. Neuroimmunol. 2001, 115, 91–100. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef]
- Maes, M.; Verkerk, R.; Bonaccorso, S.; Ombelet, W.; Bosmans, E.; Scharpé, S. Depressive and anxiety symptoms in the early puerperium are related to increased degradation of tryptophan into kynurenine, a phenomenon which is related to immune activation. Life Sci. 2002, 71, 1837–1848. [Google Scholar] [CrossRef]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress sounds the alarmin: The role of the danger-associated molecular pattern HMGB1 in stress-induced neuroinflammatory priming. Brain. Behav. Immun. 2015, 48, 1–7. [Google Scholar] [CrossRef]
- Lara-Espinosa, J.V.; Arce-Aceves, M.F.; Mata-Espinosa, D.; Barrios-Payán, J.; Marquina-Castillo, B.; Hernández-Pando, R. The therapeutic effect of intranasal administration of dexamethasone in neuroinflammation induced by experimental pulmonary tuberculosis. Int. J. Mol. Sci. 2021, 22, 5997. [Google Scholar] [CrossRef]
- Shakeri, F.; Boskabady, M.H. Anti-inflammatory, antioxidant, and immunomodulatory effects of curcumin in ovalbumin-sensitized rat. BioFactors 2017, 43, 567–576. [Google Scholar] [CrossRef]
- Avila-Rojas, S.H.; Tapia, E.; Briones-Herrera, A.; Aparicio-Trejo, O.E.; León-Contreras, J.C.; Hernández-Pando, R.; Pedraza-Chaverri, J. Curcumin prevents potassium dichromate (K2Cr2O7)-induced renal hypoxia. Food Chem. Toxicol. 2018, 121, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef] [PubMed]
- Bassani, T.B.; Turnes, J.M.; Moura, E.L.R.; Bonato, J.M.; Cóppola-Segovia, V.; Zanata, S.M.; Oliveira, R.M.M.W.; Vital, M.A.B.F. Effects of curcumin on short-term spatial and recognition memory, adult neurogenesis and neuroinflammation in a streptozotocin-induced rat model of dementia of Alzheimer’s type. Behav. Brain Res. 2017, 335, 41–54. [Google Scholar] [CrossRef]
- Izui, S.; Sekine, S.; Maeda, K.; Kuboniwa, M.; Takada, A.; Amano, A.; Nagata, H. Antibacterial Activity of Curcumin Against Periodontopathic Bacteria. J. Periodontol. 2016, 87, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, P.; Singh, M.; Kumari, H.; Kumari, A.; Mukhopadhyay, K. Bactericidal activity of curcumin I is associated with damaging of bacterial membrane. PLoS ONE 2015, 10, e0121313. [Google Scholar] [CrossRef]
- Jurenka, J.S. Anti-inflammatory Properties of Curcumin, a Major Constituent of. Altern. Med. Rev. 2009, 14, 141–154. [Google Scholar]
- Yang, C.; Zhang, X.; Fan, H.; Liu, Y. Curcumin upregulates transcription factor Nrf2, HO-1 expression and protects rat brains against focal ischemia. Brain Res. 2009, 1282, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Hurley, L.L.; Akinfiresoye, L.; Nwulia, E.; Kamiya, A.; Kulkarni, A.A.; Tizabi, Y. Antidepressant-like effects of curcumin in WKY rat model of depression is associated with an increase in hippocampal BDNF. Behav. Brain Res. 2013, 239, 27–30. [Google Scholar] [CrossRef]
- Lara-Espinosa, J.V.; Santana-Martínez, R.A.; Maldonado, P.D.; Zetter, M.; Becerril-Villanueva, E.; Pérez-Sánchez, G.; Pavón, L.; Mata-Espinosa, D.; Barrios-Payán, J.; López-Torres, M.O.; et al. Experimental pulmonary tuberculosis in the absence of detectable brain infection induces neuroinflammation and behavioural abnormalities in male balb/c mice. Int. J. Mol. Sci. 2020, 21, 9483. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R.; Kelley, K.W. Twenty years of research on cytokine-induced sickness behavior. Brain. Behav. Immun. 2007, 21, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Gilhotra, R.; Goel, S.; Gilhotra, N. Behavioral and biochemical characterization of elevated “I-maze” as animal model of anxiety. Beni-Suef Univ. J. Basic Appl. Sci. 2015, 4, 214–224. [Google Scholar] [CrossRef][Green Version]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of chronic oxidative stress on neuroinflammatory response mediated by CD4+T cells in neurodegenerative diseases. Front. Cell. Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef]
- Singh, S.; Nagalakshmi, D.; Sharma, K.K.; Ravichandiran, V. Natural antioxidants for neuroinflammatory disorders and possible involvement of Nrf2 pathway: A review. Heliyon 2021, 7, e06216. [Google Scholar] [CrossRef]
- Monteggia, L.M.; Björkholm, C. BDNF—A key transducer of antidepressant effects. Neuropharmacology 2016, 102, 72–79. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, Y.; Xu, Y.; Lian, Y.; Xie, N.; Wu, T.; Zhang, H.; Sun, L.; Zhang, R.; Wang, Z. Curcumin improves amyloid β-peptide (1-42) induced spatial memory deficits through BDNF-ERK signaling pathway. PLoS ONE 2015, 10, e0131525. [Google Scholar] [CrossRef]
- Changtam, C.; Hongmanee, P.; Suksamrarn, A. Isoxazole analogs of curcuminoids with highly potent multidrug-resistant antimycobacterial activity. Eur. J. Med. Chem. 2010, 45, 4446–4457. [Google Scholar] [CrossRef]
- Marathe, S.A.; Dasgupta, I.; Gnanadhas, D.P.; Marathe, S.A.; Dasgupta, I.; Gnanadhas, D.P. Multifaceted roles of curcumin: Two sides of a coin! Expert Opin. Biol. Ther. 2011, 11, 1485–1499. [Google Scholar] [CrossRef]
- Singh, A.K.; Karaulia, P.; Yadav, P.; Narender, T.; Singh, S.P.; Sashidhara, K.V.; Pandey, A.K.; Chopra, S.; Dasgupta, A. Identification of lipid metabolism-targeting compounds active against drug-resistant M. tuberculosis. J. Glob. Antimicrob. Resist. 2016, 7, 26–27. [Google Scholar] [CrossRef]
- Gupta, P.K. Inhibition of Intracellular Survival of Multi Drug Resistant Clinical Isolates of Mycobacterium tuberculosis in Macrophages by Curcumin. Open Antimicrob. Agents J. 2013, 4, 1–5. [Google Scholar] [CrossRef]
- Bai, X.; Oberley-Deegan, R.E.; Bai, A.; Ovrutsky, A.R.; Kinney, W.H.; Weaver, M.; Zhang, G.; Honda, J.R.; Chan, E.D. Curcumin enhances human macrophage control of Mycobacterium tuberculosis infection. Respirology 2016, 21, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Bhattacharya, D.; Kar, S.; Ranganathan, A.; Van Kaer, L.; Das, G. Curcumin nanoparticles enhance mycobacterium bovis BCG vaccine efficacy by modulating host immune responses. Infect. Immun. 2019, 87, e00291-19. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian Moghadamtousi, S.; Abdul Kadir, H.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. Biomed. Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Bansal, S.; Chhibber, S. Curcumin alone and in combination with augmentin protects against pulmonary inflammation and acute lung injury generated during Klebsiella pneumoniae B5055-induced lung infection in BALB/c mice. J. Med. Microbiol. 2010, 59, 429–437. [Google Scholar] [CrossRef]
- Réus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The role of inflammation and microglial activation in the pathophysiology of psychiatric disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef]
- Kaufmann, F.N.; Gazal, M.; Bastos, C.R.; Kaster, M.P.; Ghisleni, G. Curcumin in depressive disorders: An overview of potential mechanisms, preclinical and clinical findings. Eur. J. Pharmacol. 2016, 784, 192–198. [Google Scholar] [CrossRef]
- Tiwari, V.; Chopra, K. Attenuation of oxidative stress, neuroinflammation, and apoptosis by curcumin prevents cognitive deficits in rats postnatally exposed to ethanol. Psychopharmacology 2012, 224, 519–535. [Google Scholar] [CrossRef]
- Zhang, H.J.; Xing, Y.Q.; Jin, W.; Li, D.; Wu, K.; Lu, Y. Effects of curcumin on interleukin-23 and interleukin-17 expression in rat retina after retinal ischemia-reperfusion injury. Int. J. Clin. Exp. Pathol. 2015, 8, 9223–9231. [Google Scholar]
- Liu, Z.J.; Liu, W.; Liu, L.; Xiao, C.; Wang, Y.; Jiao, J.S. Curcumin protects neuron against cerebral ischemia-induced inflammation through improving PPAR-gamma function. Evid. Based Complement. Altern. Med. 2013, 2013, 470975. [Google Scholar] [CrossRef]
- Tu, X.K.; Yang, W.Z.; Chen, J.P.; Chen, Y.; Ouyang, L.Q.; Xu, Y.C.; Shi, S.S. Curcumin Inhibits TLR2/4-NF-κB Signaling Pathway and Attenuates Brain Damage in Permanent Focal Cerebral Ischemia in Rats. Inflammation 2014, 37, 1544–1551. [Google Scholar] [CrossRef]
- Venigalla, M.; Gyengesi, E.; Münch, G. Curcumin and apigenin—Novel and promising therapeutics against chronic neuroinflammation in Alzheimer’s disease. Neural Regen. Res. 2015, 10, 1181–1185. [Google Scholar] [CrossRef]
- Wang, Y.; Yin, H.; Wang, L.; Shuboy, A.; Lou, J.; Han, B.; Zhang, X.; Li, J. Curcumin as a potential treatment for Alzheimer’s disease: A study of the effects of curcumin on hippocampal expression of glial fibrillary acidic protein. Am. J. Chin. Med. 2013, 41, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Gilani, A.H. A comparative study of curcuminoids to measure their effect on inflammatory and apoptotic gene expression in an Aβ plus ibotenic acid-infused rat model of Alzheimer’s disease. Brain Res. 2011, 1400, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin Has Potent Anti-Amyloidogenic Effects for Alzheimer’s β-Amyloid Fibrils In Vitro. J. Neurosci. Res. 2004, 75, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zheng, Z.; Li, J.; Xiao, Z.; Qi, W.; Zhang, A.; Wu, Q.; Fang, Y. Curcumin inhibits Aβ-induced microglial inflammatory responses in vitro: Involvement of ERK1/2 and p38 signaling pathways. Neurosci. Lett. 2015, 594, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Zhuang, W.; Zhou, S.; Wang, X. Plant-derived neuroprotective agents in Parkinson’s disease. Am. J. Transl. Res. 2015, 7, 1189–1202. [Google Scholar] [PubMed]
- Ojha, R.P.; Rastogi, M.; Devi, B.P.; Agrawal, A.; Dubey, G.P. Neuroprotective effect of curcuminoids against inflammation-mediated dopaminergic neurodegeneration in the mptp model of parkinson’s Disease. J. Neuroimmune Pharmacol. 2012, 7, 609–618. [Google Scholar] [CrossRef]
- Tegenge, M.A.; Rajbhandari, L.; Shrestha, S.; Mithal, A.; Hosmane, S.; Venkatesan, A. Curcumin protects axons from degeneration in the setting of local neuroinflammation. Exp. Neurol. 2014, 253, 102–110. [Google Scholar] [CrossRef]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [PubMed]
- Esmaily, H.; Sahebkar, A.; Iranshahi, M.; Ganjali, S.; Mohammadi, A.; Ferns, G.; Ghayour-Mobarhan, M. An investigation of the effects of curcumin on anxiety and depression in obese individuals: A randomized controlled trial. Chin. J. Integr. Med. 2015, 21, 332–338. [Google Scholar] [CrossRef]
- Haider, S.; Naqvi, F.; Batool, Z.; Tabassum, S.; Sadir, S.; Liaquat, L.; Naqvi, F.; Zuberi, N.A.; Shakeel, H.; Perveen, T. Pretreatment with curcumin attenuates anxiety while strengthens memory performance after one short stress experience in male rats. Brain Res. Bull. 2015, 115, 1–8. [Google Scholar] [CrossRef]
- Abu-Taweel, G.M.; Ajarem, J.S.; Ahmad, M. Protective Effect of Curcumin on Anxiety, Learning Behavior, Neuromuscular Activities, Brain Neurotransmitters and Oxidative Stress Enzymes in Cadmium Intoxicated Mice. J. Behav. Brain Sci. 2013, 3, 74–84. [Google Scholar] [CrossRef]
- Fusar-Poli, L.; Vozza, L.; Gabbiadini, A.; Vanella, A.; Concas, I.; Tinacci, S.; Petralia, A.; Signorelli, M.S.; Aguglia, E. Curcumin for depression: A meta-analysis. Crit. Rev. Food Sci. Nutr. 2020, 60, 2643–2653. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Q.; Huang, H.; Liu, Z. The efficacy and acceptability of curcumin for the treatment of depression or depressive symptoms: A systematic review and meta-analysis. J. Affect. Disord. 2021, 282, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.A.; Adlimoghaddam, A.; Albensi, B.C. Role of Nrf2 in Synaptic Plasticity and Memory in Alzheimer’s Disease. Cells 2021, 10, 1884. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, Q.; Wang, X.; Yu, S.; Li, L.; Wu, X.; Chen, Y.; Zhao, J.; Zhao, Y. Neuroprotection by Curcumin in Ischemic Brain Injury Involves the Akt/Nrf2 Pathway. PLoS ONE 2013, 8, e59843. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Yang, B.; Wang, L.; Li, B.; Guo, X.; Zhang, M.; Jiang, Z.; Fu, J.; Pi, J.; Guan, D.; et al. Curcumin plays neuroprotective roles against traumatic brain injury partly via Nrf2 signaling. Toxicol. Appl. Pharmacol. 2018, 346, 28–36. [Google Scholar] [CrossRef]
- Aske, K.C.; Waugh, C.A. Expanding the 3R principles. EMBO Rep. 2017, 18, 1490–1492. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Pando, R.; Orozcoe, H.; Sampieri, A.; Pavón, L.; Velasquillo, C.; Larriva-Sahd, J.; Alcocer, J.M.; Madrid, M. V Correlation between the kinetics of Th1, Th2 cells and pathology in a murine model of experimental pulmonary tuberculosis. Immunology 1996, 89, 26–33. [Google Scholar]
- Hernandez Pando, R.; Aguilar, D.; Cohen, I.; Guerrero, M.; Ribon, W.; Acosta, P.; Orozco, H.; Marquina, B.; Salinas, C.; Rembao, D.; et al. Specific bacterial genotypes of Mycobacterium tuberculosis cause extensive dissemination and brain infection in an experimental model. Tuberculosis 2010, 90, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.B.J.; Paxinos, G. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2013; Volume 246. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analysing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Silva-Islas, C.A.; Chánez-Cárdenas, M.E.; Barrera-Oviedo, D.; Ibarra-Rubio, M.E.; Maldonado, P.D. Acute expression of the transcription factor Nrf2 after treatment with quinolinic acid is not induced by oxidative stress in the rat striatum. Neurotoxicology 2019, 73, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. 10 NIH Image to ImageJ 25 years of image analysis. Nat. Methods 2012, 9, 671–676. [Google Scholar] [CrossRef]
- Nestler, E.J.; Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef]
- Multani, P.K.; Saini, N.; Kaur, R.; Sharma, P. Biomarkers for Drugs of Abuse and Neuropsychiatric Disorders: Models and Mechanisms; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780124046306. [Google Scholar]
- Stahel, P.F.; Shohami, E.; Younis, F.M.; Kariya, K.; Otto, V.I.; Lenzlinger, P.M.; Grosjean, M.B.; Eugster, H.P.; Trentz, O.; Kossmann, T.; et al. Experimental closed head injury: Analysis of neurological outcome, blood-brain barrier dysfunction, in-tracranial neutrophil infiltration, and neuronal cell death in mice deficient in genes for pro-inflammatory cytokines. J. Cereb. Blood Flow Metab. 2000, 20, 369–380. [Google Scholar] [CrossRef]
- Ennaceur, A.; Neave, N.; Aggleton, J.P. Spontaneous object recognition and object location memory in rats: The effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Exp. Brain Res. 1997, 113, 509–519. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | 5′-CATTGTGGAAGGGCTATGA-3′ | 5′-GGAAGGCCATGCCAGTGAGC-3′ |
| TNFα | 5′-GCCGAGAAAGGCTGCTTG-3′ | 5′-TGTGGCTTCGACCTCTACCTC-3′ |
| IFNγ | 5′-CCTCAACTTGGCAATCTCATGA-3′ | 5′-GGTGACATGAAAATCCTGCAG-3′ |
| IL12 | 5′-GGATGGAAGAGTCCCCCAAA-3′ | 5′-GCTCTGCGGGCATTTAACAT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lara-Espinosa, J.V.; Arce-Aceves, M.F.; López-Torres, M.O.; Lozano-Ordaz, V.; Mata-Espinosa, D.; Barrios-Payán, J.; Silva-Islas, C.A.; Maldonado, P.D.; Marquina-Castillo, B.; Hernández-Pando, R. Effect of Curcumin in Experimental Pulmonary Tuberculosis: Antimycobacterial Activity in the Lungs and Anti-Inflammatory Effect in the Brain. Int. J. Mol. Sci. 2022, 23, 1964. https://doi.org/10.3390/ijms23041964
Lara-Espinosa JV, Arce-Aceves MF, López-Torres MO, Lozano-Ordaz V, Mata-Espinosa D, Barrios-Payán J, Silva-Islas CA, Maldonado PD, Marquina-Castillo B, Hernández-Pando R. Effect of Curcumin in Experimental Pulmonary Tuberculosis: Antimycobacterial Activity in the Lungs and Anti-Inflammatory Effect in the Brain. International Journal of Molecular Sciences. 2022; 23(4):1964. https://doi.org/10.3390/ijms23041964
Chicago/Turabian StyleLara-Espinosa, Jacqueline V., María Fernanda Arce-Aceves, Manuel O. López-Torres, Vasti Lozano-Ordaz, Dulce Mata-Espinosa, Jorge Barrios-Payán, Carlos Alfredo Silva-Islas, Perla D. Maldonado, Brenda Marquina-Castillo, and Rogelio Hernández-Pando. 2022. "Effect of Curcumin in Experimental Pulmonary Tuberculosis: Antimycobacterial Activity in the Lungs and Anti-Inflammatory Effect in the Brain" International Journal of Molecular Sciences 23, no. 4: 1964. https://doi.org/10.3390/ijms23041964
APA StyleLara-Espinosa, J. V., Arce-Aceves, M. F., López-Torres, M. O., Lozano-Ordaz, V., Mata-Espinosa, D., Barrios-Payán, J., Silva-Islas, C. A., Maldonado, P. D., Marquina-Castillo, B., & Hernández-Pando, R. (2022). Effect of Curcumin in Experimental Pulmonary Tuberculosis: Antimycobacterial Activity in the Lungs and Anti-Inflammatory Effect in the Brain. International Journal of Molecular Sciences, 23(4), 1964. https://doi.org/10.3390/ijms23041964