
Nitric Oxide Extends the Postharvest Life of Water Bamboo Shoots Partly by Maintaining Mitochondrial Structure and Energy Metabolism

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
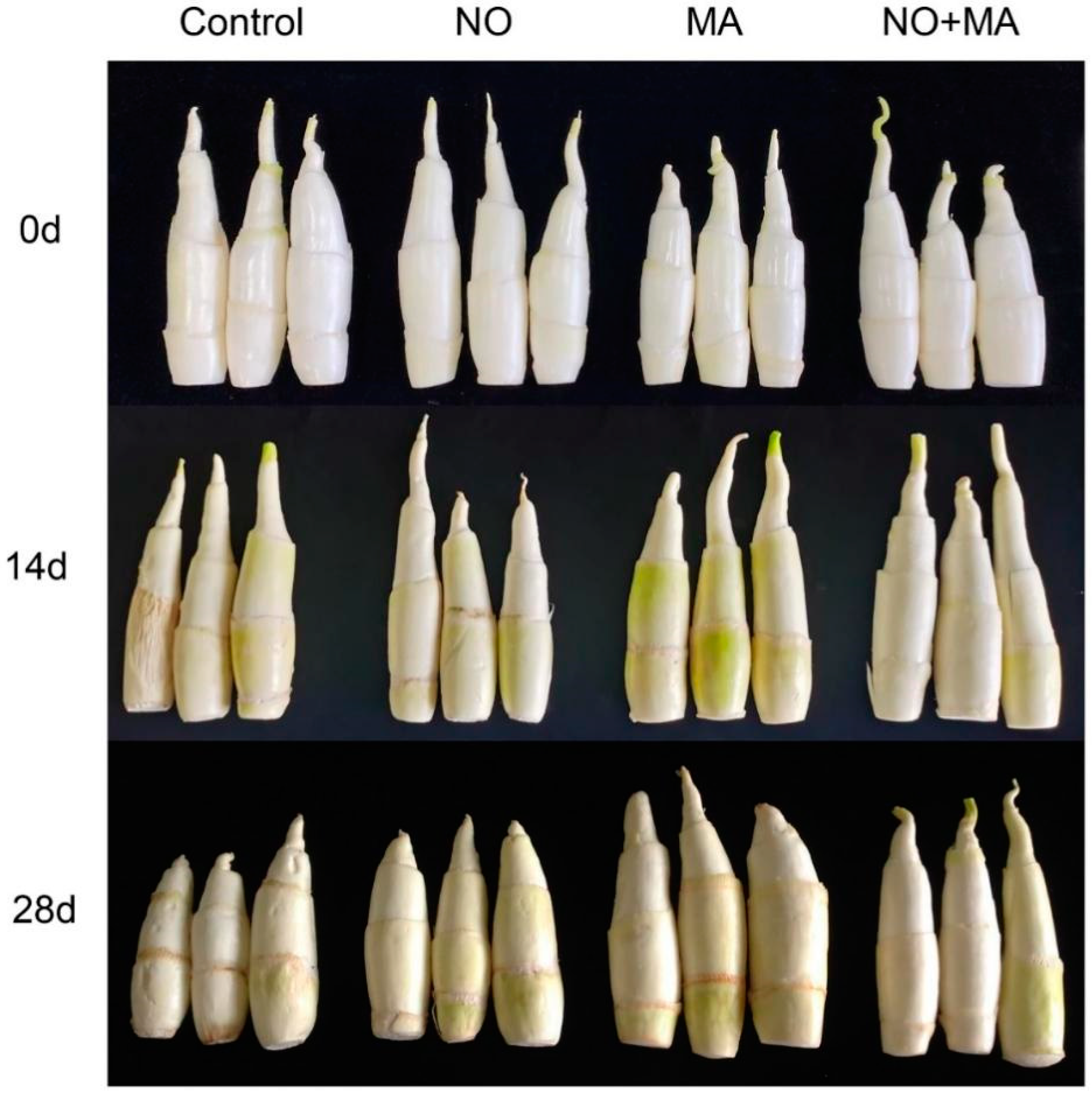
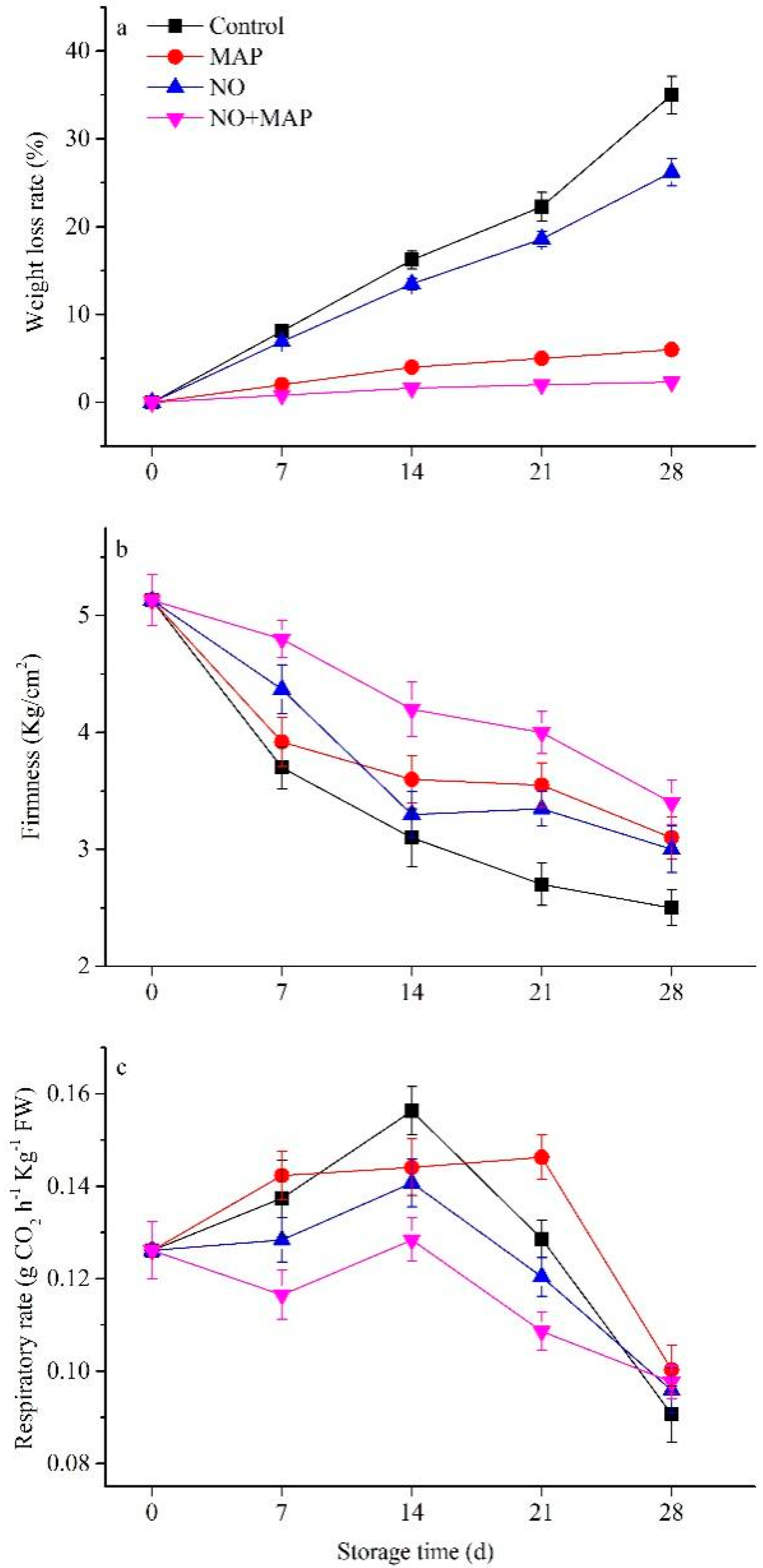
2.1. Effects of NO and MAP on Appearance, Weight Loss, and Firmness of Water Bamboo Shoots during Cold Storage
2.2. Effects of NO and MAP on Respiration Rate of Water Bamboo Shoots during Cold Storage
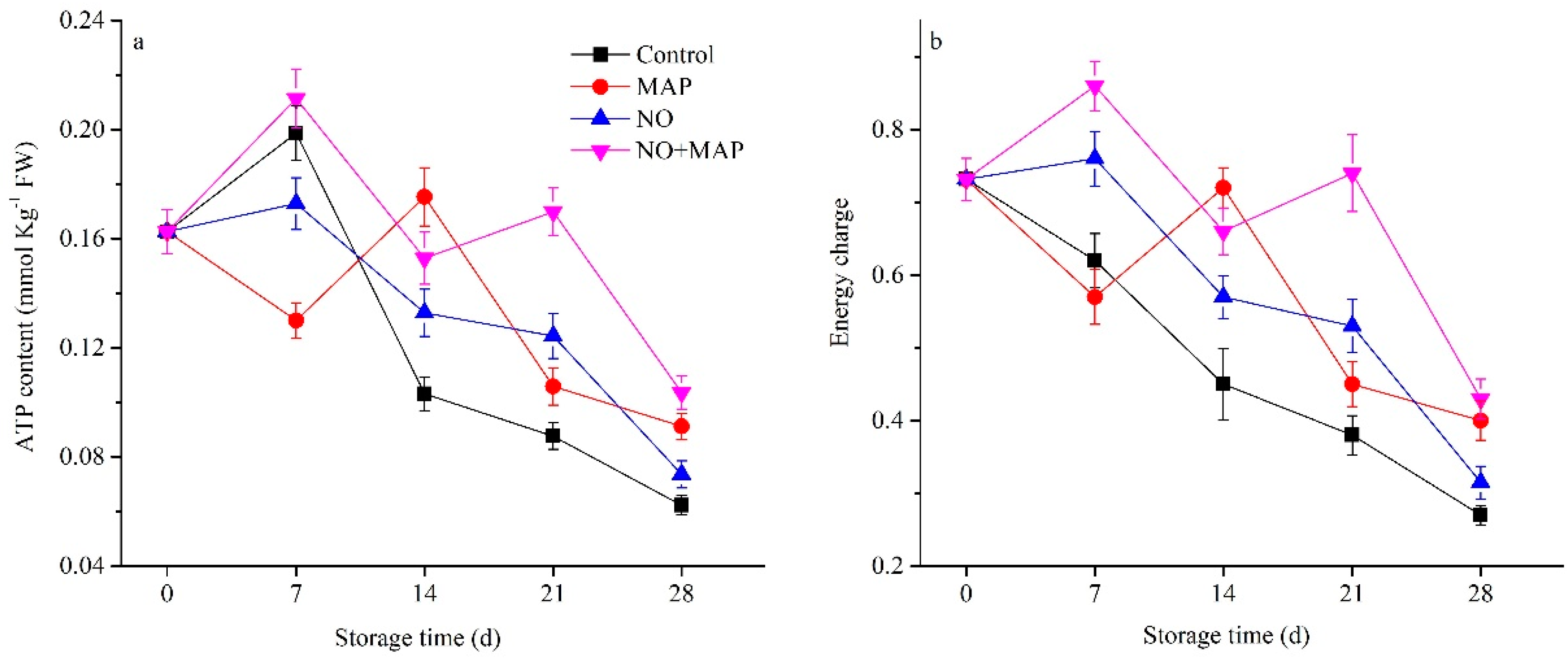
2.3. Effects of NO and MAP on ATP and Energy Charge Levels of Water Bamboo Shoots during Cold Storage
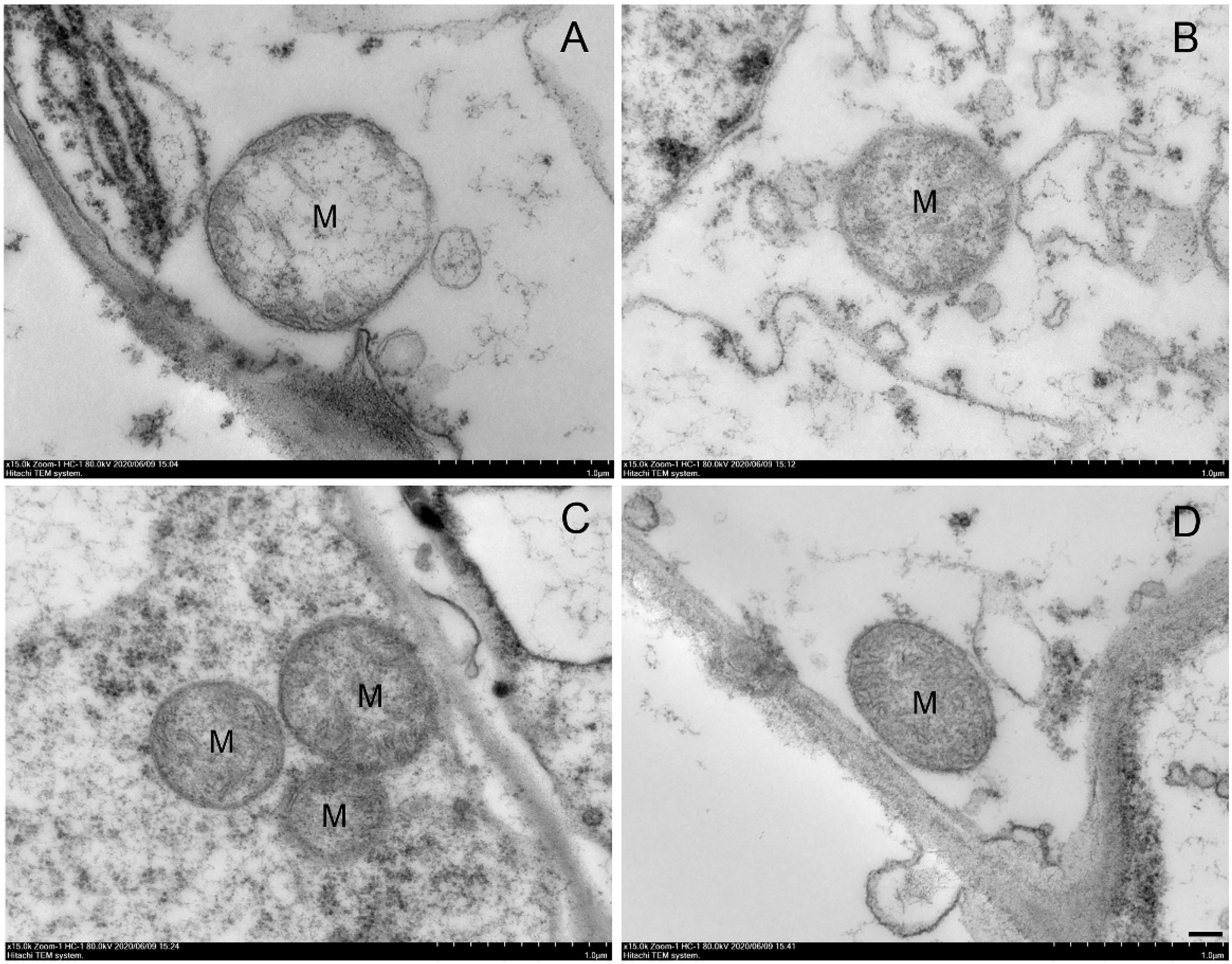
2.4. Effects of NO and MAP on Mitochondrial Ultrastructure of Water Bamboo Shoots during Cold Storage
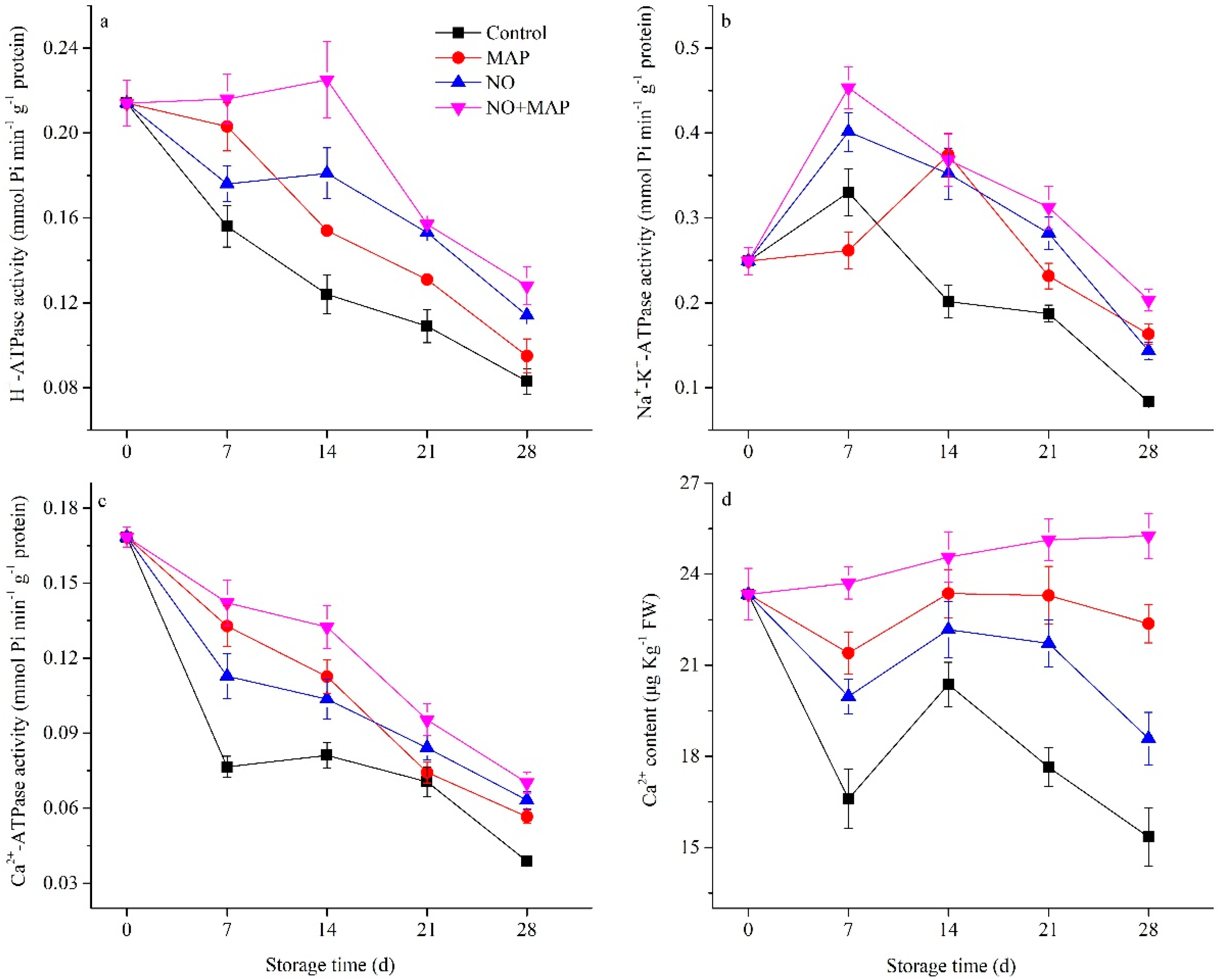
2.5. Effects of NO and MAP on ATPase Activity and Ca2+ Content of Water Bamboo Shoots during Cold Storage
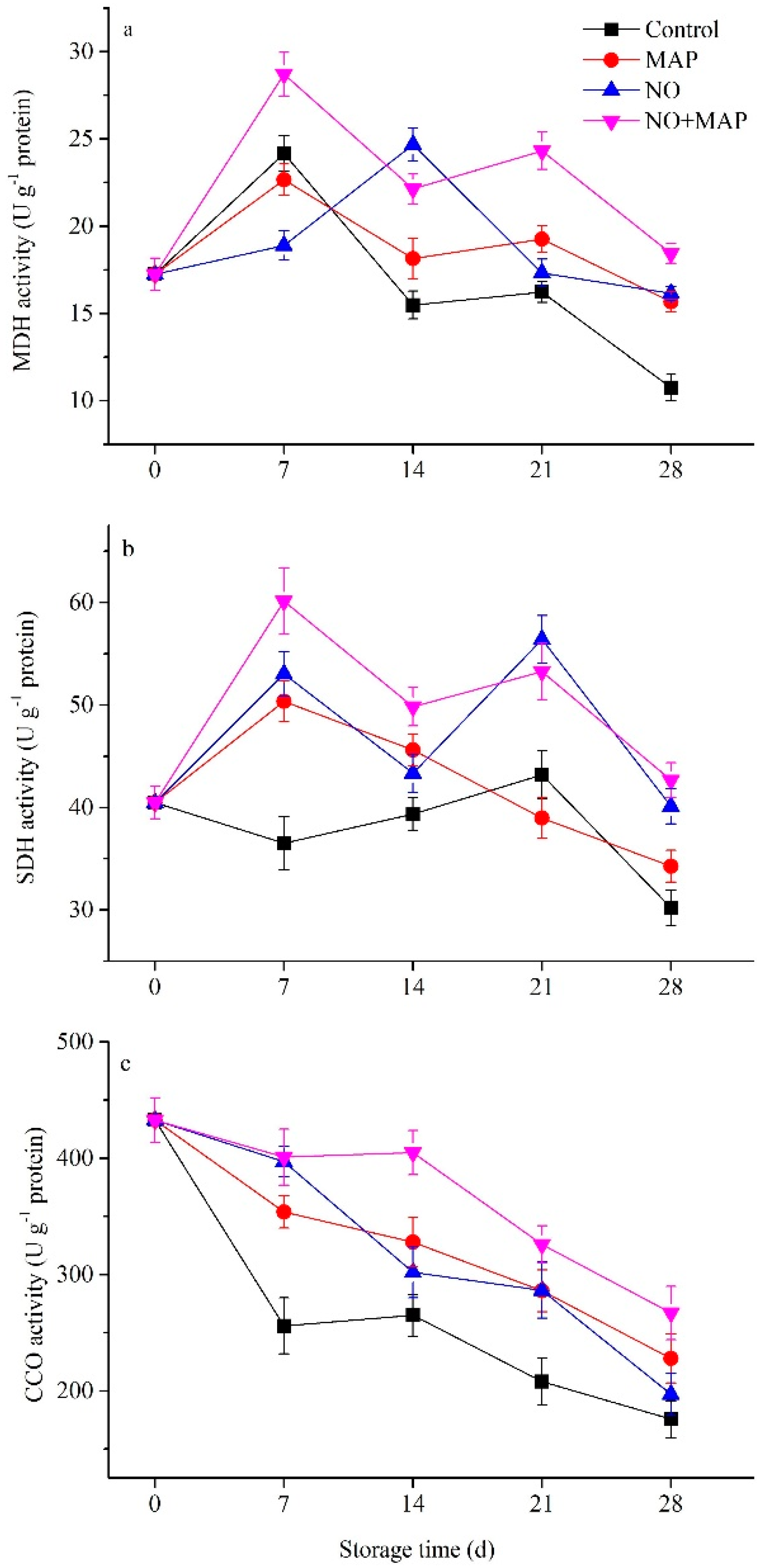
2.6. Effects of NO and MAP on Enzyme Activities of MDH, SDH, and CCO of Water Bamboo Shoots during Cold Storage
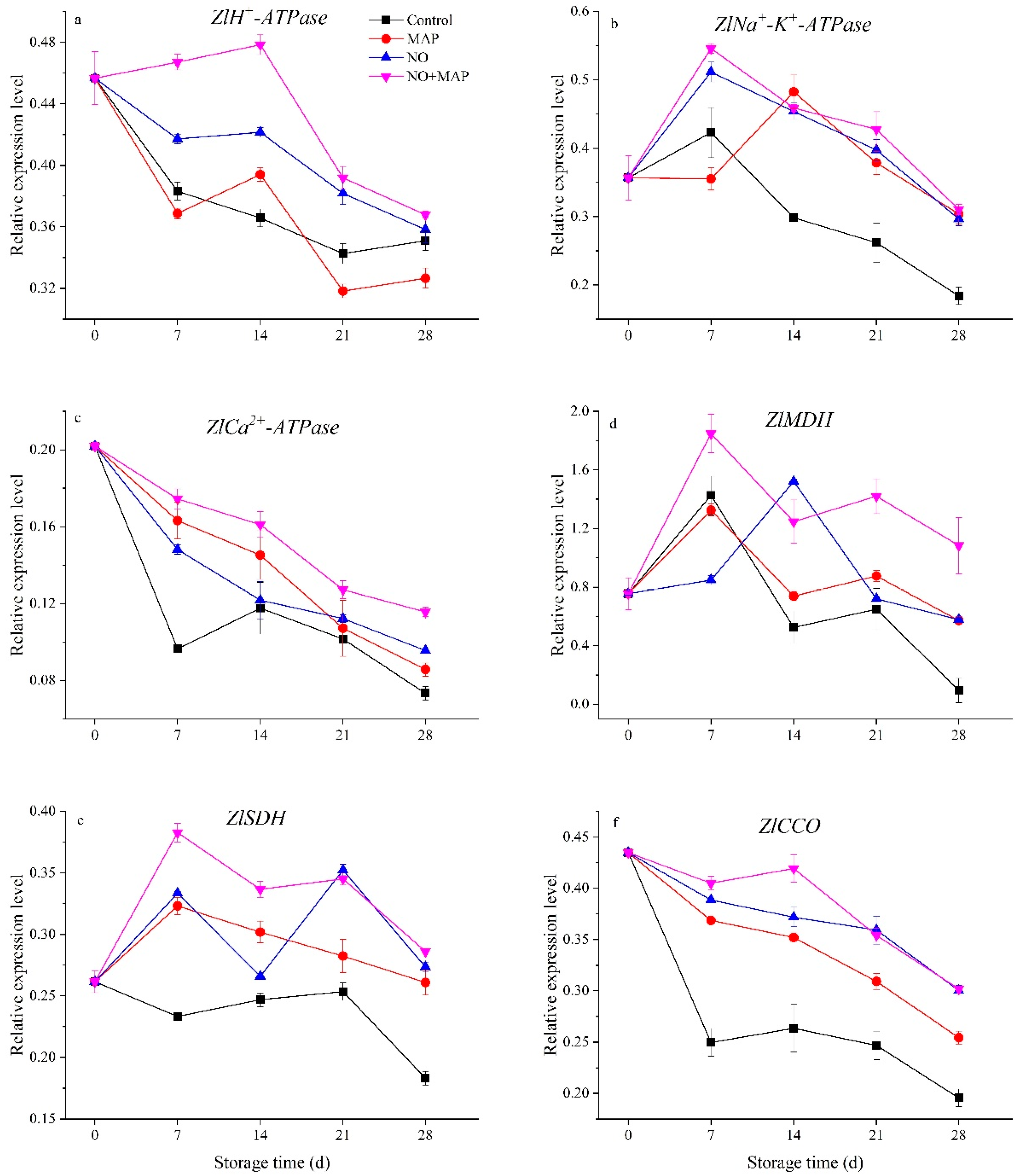
2.7. Effects of NO and MAP on the Expression of ZlH+-ATPase, ZlNa+-K+-ATPase, ZlCa2+-ATPase, ZlMDH, ZlSDH, and ZlCCO of Water Bamboo Shoots during Cold Storage
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Treatments
4.3. Weight Loss, Firmness, and Respiration Rate Assays
4.4. ATP and Energy Charge Measurement
4.5. Mitochondrial Ultrastructure
4.6. The Activities of H+-ATPase, Ca2+-ATPase, and Na+-K+- ATPase
4.7. Measurement of Ca2+ Concentration
4.8. Mitochondria Extraction and Enzyme Extraction
4.9. RNA Extraction and Quantitative Real-Time PCR Analysis
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Guo, H.B.; Li, S.M.; Peng, J.; Ke, W.D. Zizania latifolia Turcz. cultivated in China. Genet. Resour. Crop Evol. 2007, 54, 1211–1217. [Google Scholar] [CrossRef]
- Wen, B.; Cheng, Z.; Hu, Y.; Boon-Ek, Y.; Wongs-Aree, C.; Supapanich, S. Ultraviolet-C treatment maintains physicochemical quality of water bamboo (Zizania latifolia) shoots during postharvest storage. Postharvest Biol. Technol. 2019, 152, 65–72. [Google Scholar] [CrossRef]
- Qi, X.H.; Ji, Z.J.; Lin, C.; Li, S.F.; Liu, J.; Kan, J.; Zhang, M.; Jin, C.H.; Qian, C.L. Nitric oxide alleviates lignification and softening of water bamboo (Zizania latifolia) shoots during postharvest storage. Food Chem. 2020, 332, 127416. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, Y.; Lin, H.; Ritenour, M.A.; Shi, J.; Zhang, S.; Chen, Y.; Wang, H. Hydrogen peroxide-induced pericarp browning of harvested longan fruit in association with energy metabolism. Food Chem. 2017, 225, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Shi, C.; Hu, Q.; Wu, Y.; Fang, D.; Pei, F.; Mariga, A.M. Nanocomposite packaging regulate respiration and energy metabolism in Flammulina velutipes. Postharvest Biol. Technol. 2019, 151, 119–126. [Google Scholar] [CrossRef]
- Li, L.; Lv, F.Y.; Guo, Y.Y.; Wang, Z.Q. Respiratory pathway metabolism and energy metabolism associated with senescence in postharvest Broccoli (Brassica oleracea L. var. italica) florets in response to O2/CO2 controlled atmospheres. Postharvest Biol. Technol. 2016, 111, 330–336. [Google Scholar] [CrossRef]
- Soto, I.C.; Fontanesi, F.; Liu, J.; Barrientos, A. Biogenesis and assembly of eukaryotic cytochrome c oxidase catalytic core. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 883–897. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Liu, M.; Zhou, J. Inhibition by nitric oxide of ethylene biosynthesis and lipoxygenase activity in peach fruit during storage. Postharvest Biol. Technol. 2006, 42, 41–48. [Google Scholar] [CrossRef]
- Wills, R.B.H.; Pristijono, P.; Golding, J.B. Browning on the surface of cut lettuce slices inhibited by short term exposure to nitric oxide (NO). Food Chem. 2008, 107, 1387–1392. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, C.; Wu, F.; Cheng, J. Effect of nitric oxide on browning and lignification of peeled bamboo shoots. Postharvest Biol. Technol. 2010, 57, 72–76. [Google Scholar] [CrossRef]
- Hao, Y.; Chen, F.; Wu, G.; Gao, W. Impact of postharvest nitric oxide treatment on lignin biosynthesis-related genes in wax apple (Syzygium samarangense) Fruit. J. Agric. Food Chem. 2016, 64, 8483–8490. [Google Scholar] [CrossRef] [PubMed]
- Steelheart, C.; Alegre, M.L.; Vera Bahima, J.; Senn, M.E.; Simontacchi, M.; Bartoli, C.G.; Gergoff Grozeff, G.E. Nitric oxide improves the effect of 1-methylcyclopropene extending the tomato (Lycopersicum esculentum L.) fruit postharvest life. Sci. Hortic. 2019, 255, 193–201. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Hou, Y.; Wang, X.; Li, X. Postharvest nitric oxide treatment delays the senescence of winter jujube (Zizyphus jujuba Mill. cv. Dongzao) fruit during cold storage by regulating reactive oxygen species metabolism. Sci. Hortic. 2020, 261, 109009. [Google Scholar] [CrossRef]
- Miao, M.; Wang, Q.; Zhang, T.; Jiang, B. Effect of high hydrostatic pressure (HHP) treatment on texture changes of water bamboo shoots cultivated in China. Postharvest Biol. Technol. 2011, 59, 327–329. [Google Scholar] [CrossRef]
- Song, L.; Gao, H.; Chen, W.; Chen, H.; Mao, J.; Zhou, Y.; Duan, X.; Joyce, D.C. The role of 1-methylcyclopropene in lignification and expansin gene expression in peeled water bamboo shoot (Zizania caduciflora L.). J. Sci. Food Agric. 2011, 91, 2679–2683. [Google Scholar] [CrossRef]
- Song, L.; Chen, H.; Gao, H.; Fang, X.; Mu, H.; Yuan, Y.; Yang, Q.; Jiang, Y. Combined modified atmosphere packaging and low temperature storage delay lignification and improve the defense response of minimally processed water bamboo shoot. Chem. Cent. J. 2013, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liamnimitr, N.; Thammawong, M.; Techavuthiporn, C.; Fahmy, K.; Suzuki, T.; Nakano, K. Optimization of bulk modified atmosphere packaging for long-term storage of ‘Fuyu’ persimmon fruit. Postharvest Biol. Technol. 2018, 135, 1–7. [Google Scholar] [CrossRef]
- Wang, C.T.; Wang, C.T.; Cao, Y.P.; Robert Nout, M.J.; Sun, B.G.; Liu, L. Effect of modified atmosphere packaging (MAP) with low and superatmospheric oxygen on the quality and antioxidant enzyme system of golden needle mushrooms (Flammulina velutipes) during postharvest storage. Eur. Food Res. Technol. 2011, 232, 851–860. [Google Scholar] [CrossRef]
- Falagán, N.; Terry, L.A. Recent advances in controlled and modified atmosphere of fresh produce. Johnson Matthey Technol. Rev. 2018, 62, 107–117. [Google Scholar] [CrossRef]
- Giménez, M.; Olarte, C.; Sanz, S.; Lomas, C.; Echávarri, J.F.; Ayala, F. Relation between spoilage and microbiological quality in minimally processed artichoke packaged with different films. Food Microbiol. 2003, 20, 231–242. [Google Scholar] [CrossRef]
- Posada-Izquierdo, G.D.; Pérez-Rodríguez, F.; López-Gálvez, F.; Allende, A.; Gil, M.I.; Zurera, G. Modeling growth of Escherichia coli O157: H7 in fresh-cut lettuce treated with neutral electrolyzed water and under modified atmosphere packaging. Int. J. Food Microbiol. 2014, 177, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Siro, I.; Devlieghere, F.; Jacxsens, L.; Uyttendaele, M.; Debevere, J. The microbial safety of strawberry and raspberry fruits packaged in high-oxygen and equilibrium-modified atmospheres compared to air storage. Int. J. Food Sci. Technol. 2006, 41, 93–103. [Google Scholar] [CrossRef]
- Matar, C.; Gaucel, S.; Gontard, N.; Guilbert, S.; Guillard, V. Predicting shelf life gain of fresh strawberries ‘Charlotte cv’ in modified atmosphere packaging. Postharvest Biol. Technol. 2018, 142, 28–38. [Google Scholar] [CrossRef]
- Fahmy, K.; Nakano, K. Optimal design of modified atmosphere packaging for alleviating chilling injury in cucumber fruit. Environ. Control Biol. 2014, 52, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Qian, Z.; Ma, S.; Zhou, Y.; Patrick, J.W.; Duan, X.; Jiang, Y.; Qu, H. Energy status of ripening and postharvest senescent fruit of litchi (Litchi chinensis Sonn.). BMC Plant Biol. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lin, H.; Zhang, S.; Sun, J.; Lin, Y.; Wang, H.; Lin, M.; Shi, J. Phomopsis longanae Chi-induced disease development and pericarp browning of harvested longan fruit in association with energy metabolism. Front. Microbiol. 2018, 9, 1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, G.; Wang, Q.; Liu, J.; Li, B.; Tian, S. Proteomic analysis of changes in mitochondrial protein expression during fruit senescence. Proteomics 2009, 9, 4241–4253. [Google Scholar] [CrossRef] [PubMed]
- Kan, J.; Wang, H.M.; Jin, C.H.; Xie, H.Y. Changes of reactive oxygen species and related enzymes in mitochondria respiratory metabolism during the ripening of peach fruit. Agric. Sci. China 2010, 9, 138–146. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, S.; Su, X.; Jiang, Y. Respiratory activity and mitochondrial membrane associated with fruit senescence in postharvest peaches in response to UV-C treatment. Food Chem. 2014, 161, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Jiang, H.; Wang, Y.; Lyu, L.; Prusky, D.; Ji, Y.; Zheng, X.; Bi, Y. Effect of benzothiadiazole treatment on improving the mitochondrial energy metabolism involved in induced resistance of apple fruit during postharvest storage. Food Chem. 2020, 302, 125288. [Google Scholar] [CrossRef]
- Schertl, P.; Braun, H.P. Respiratory electron transfer pathways in plant mitochondria. Front. Plant Sci. 2014, 5, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chumyam, A.; Shank, L.; Uthaibutra, J.; Saengnil, K. Effects of chlorine dioxide on mitochondrial energy levels and redox status of “Daw” longan pericarp during storage. Postharvest Biol. Technol. 2016, 116, 26–35. [Google Scholar] [CrossRef]
- Qi, X.H.; Li, Q.Q.; Ma, X.T.; Qian, C.L.; Wang, H.H.; Ren, N.N.; Shen, C.X.; Huang, S.M.; Xu, X.W.; Xu, Q.; et al. Waterlogging-induced adventitious root formation in cucumber is regulated by ethylene and auxin through reactive oxygen species signalling. Plant Cell Environ. 2019, 42, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Guo, M.; Liu, H.; Tian, S.; Zhao, X. Inhibition of autophagy by hydroxychloroquine enhances antioxidant nutrients and delays postharvest fruit senescence of Ziziphus jujube. Food Chem. 2019, 296, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kiroaki, H.; Zhang, R.; Wang, X.; Zhang, L.; Yu, S.; Li, Y. New insights into the chilling injury of postharvest white mushroom (Agaricus bisporus) related to mitochondria and electron transport pathway under high O2/CO2 controlled atmospheres. Postharvest Biol. Technol. 2019, 152, 45–53. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lü, G.; Liang, Y.; Wu, X.; Li, J.; Ma, W.; Zhang, Y.; Gao, H. Molecular cloning and functional characterization of mitochondrial malate dehydrogenase (mMDH) is involved in exogenous GABA increasing root hypoxia tolerance in muskmelon plants. Sci. Hortic. 2019, 258, 108741. [Google Scholar] [CrossRef]
- Ackrell, B.A.C.; Kearney, E.B.; Singer, T.P. Mammalian succinate dehydrogenase. Methods Enzymol. 1978, 53, 466–483. [Google Scholar]
- Errede, B.J.; Kamen, M.D.; Hatefi, Y. Preparation and properties of complex IV (Ferrocytochrome c: Oxygen oxidoreductase EC 1.9.3.1). Methods Enzymol. 1978, 53, 40–47. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, C.; Ji, Z.; Lin, C.; Zhang, M.; Zhang, J.; Kan, J.; Liu, J.; Jin, C.; Xiao, L.; Qi, X. Nitric Oxide Extends the Postharvest Life of Water Bamboo Shoots Partly by Maintaining Mitochondrial Structure and Energy Metabolism. Int. J. Mol. Sci. 2022, 23, 1607. https://doi.org/10.3390/ijms23031607
Qian C, Ji Z, Lin C, Zhang M, Zhang J, Kan J, Liu J, Jin C, Xiao L, Qi X. Nitric Oxide Extends the Postharvest Life of Water Bamboo Shoots Partly by Maintaining Mitochondrial Structure and Energy Metabolism. International Journal of Molecular Sciences. 2022; 23(3):1607. https://doi.org/10.3390/ijms23031607
Chicago/Turabian StyleQian, Chunlu, Zhengjie Ji, Chen Lin, Man Zhang, Jixian Zhang, Juan Kan, Jun Liu, Changhai Jin, Lixia Xiao, and Xiaohua Qi. 2022. "Nitric Oxide Extends the Postharvest Life of Water Bamboo Shoots Partly by Maintaining Mitochondrial Structure and Energy Metabolism" International Journal of Molecular Sciences 23, no. 3: 1607. https://doi.org/10.3390/ijms23031607
APA StyleQian, C., Ji, Z., Lin, C., Zhang, M., Zhang, J., Kan, J., Liu, J., Jin, C., Xiao, L., & Qi, X. (2022). Nitric Oxide Extends the Postharvest Life of Water Bamboo Shoots Partly by Maintaining Mitochondrial Structure and Energy Metabolism. International Journal of Molecular Sciences, 23(3), 1607. https://doi.org/10.3390/ijms23031607