
Suitability of GRK Antibodies for Individual Detection and Quantification of GRK Isoforms in Western Blots


Abstract
:1. Introduction
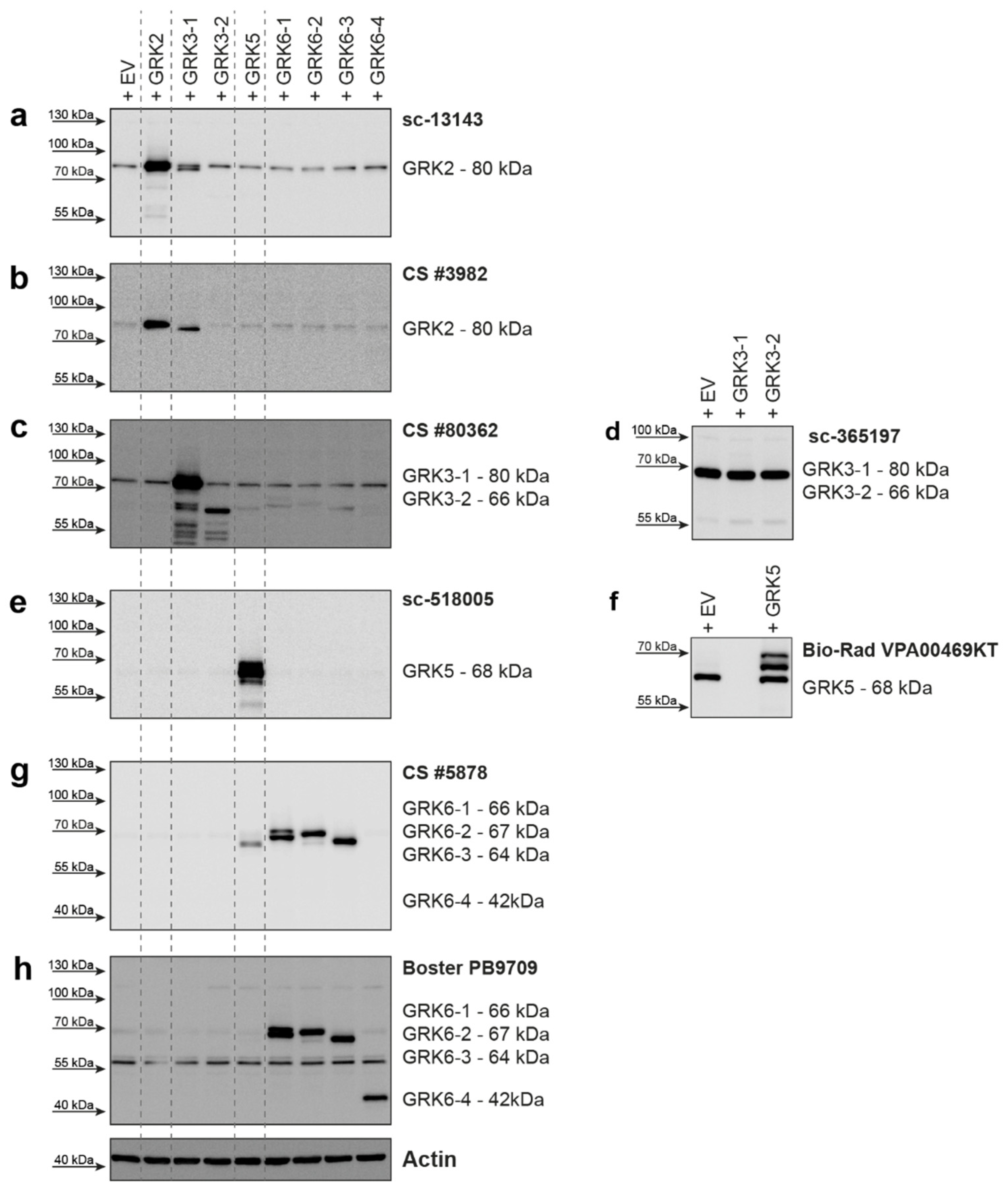
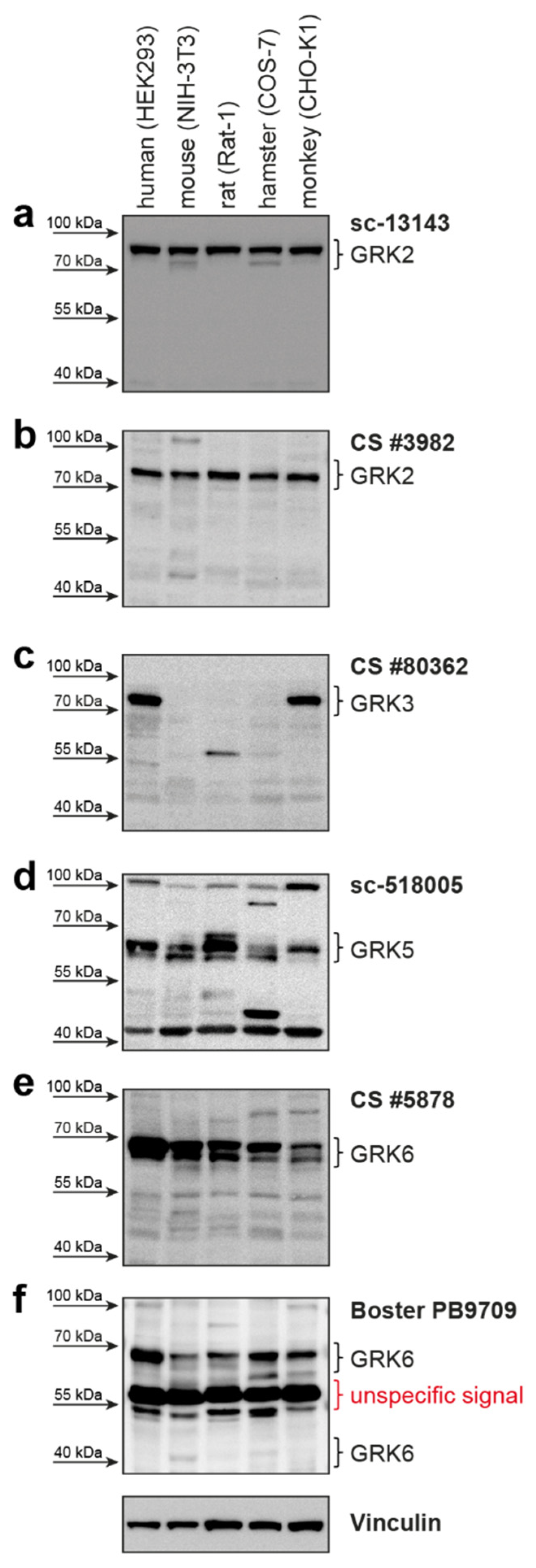
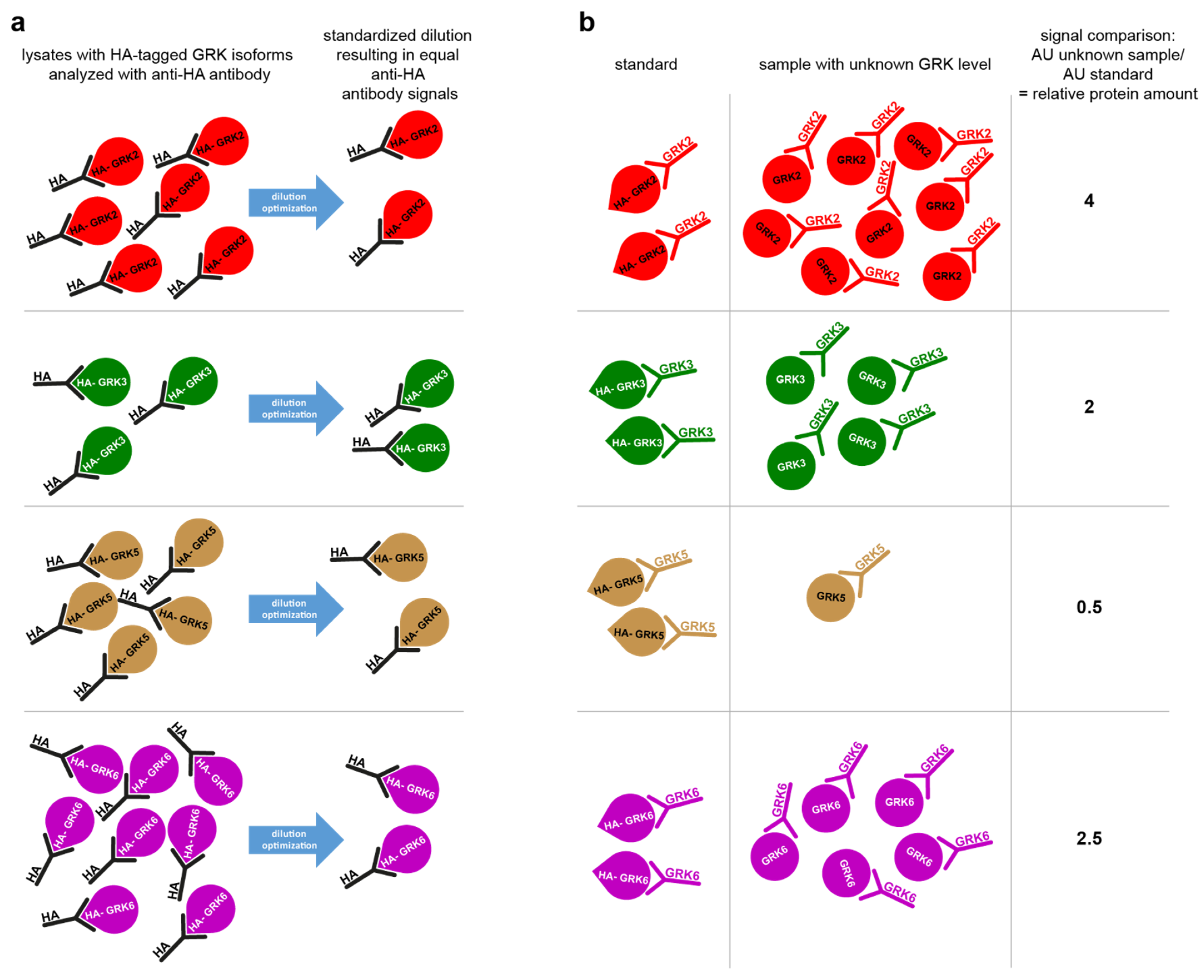
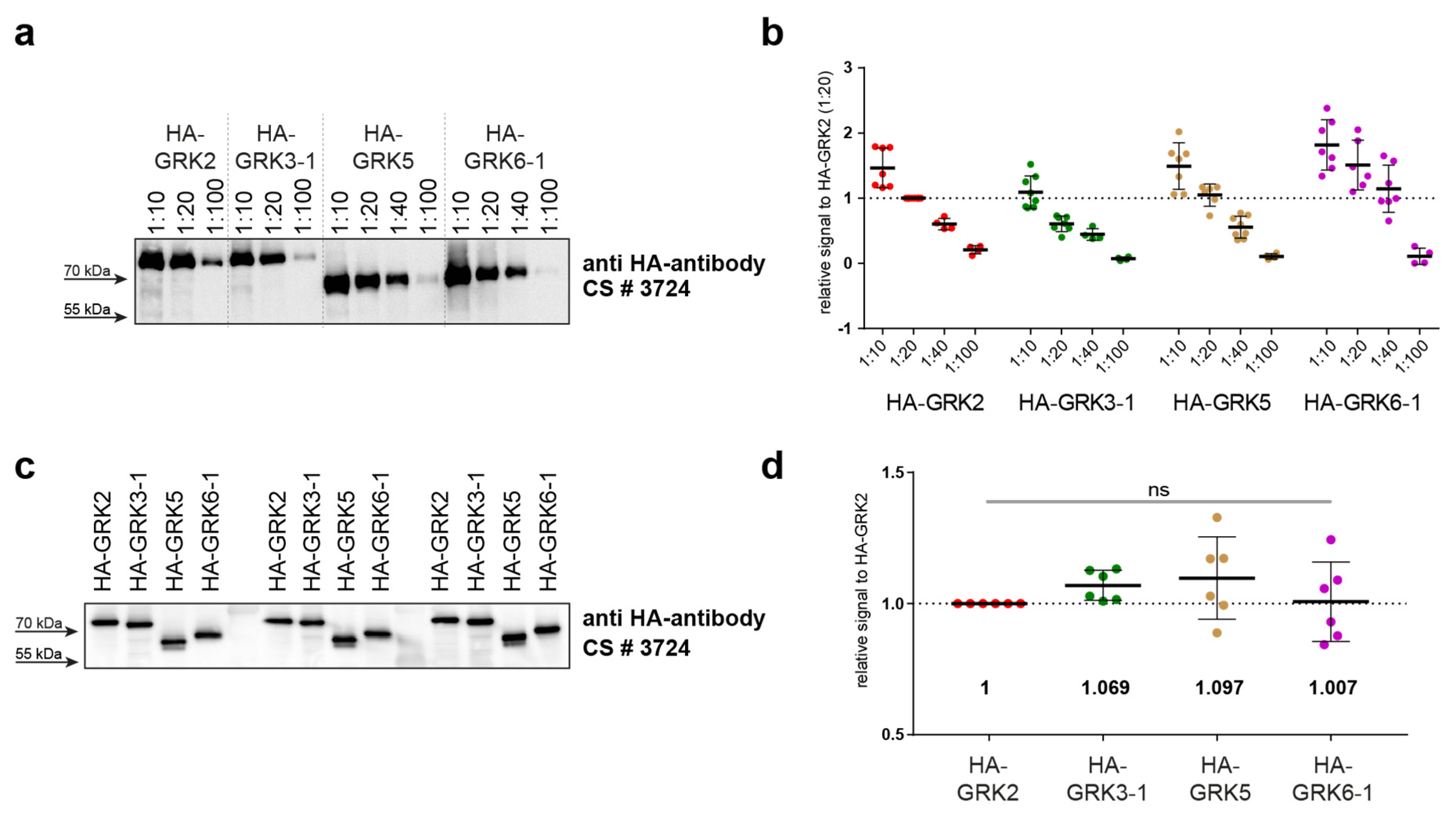
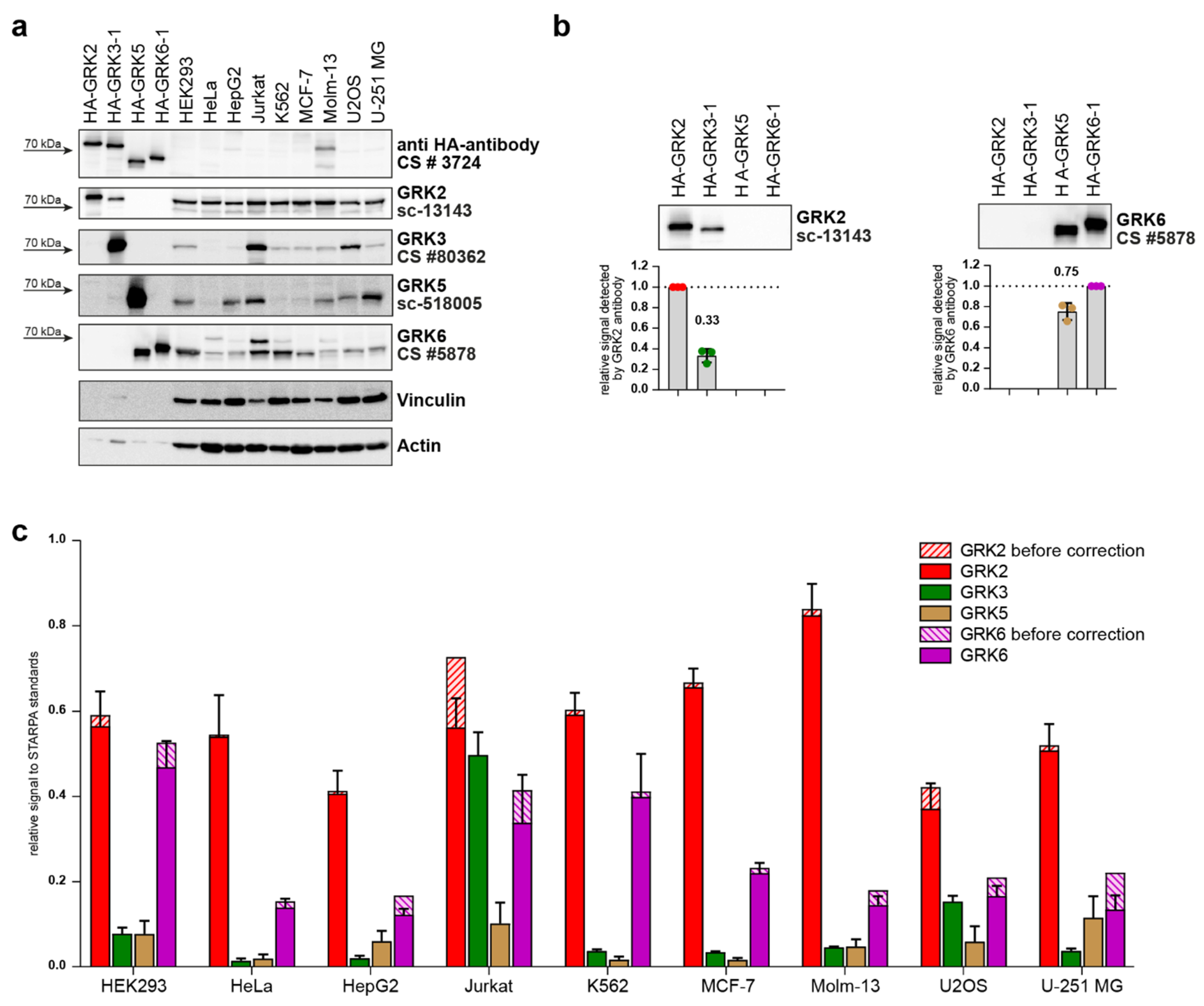
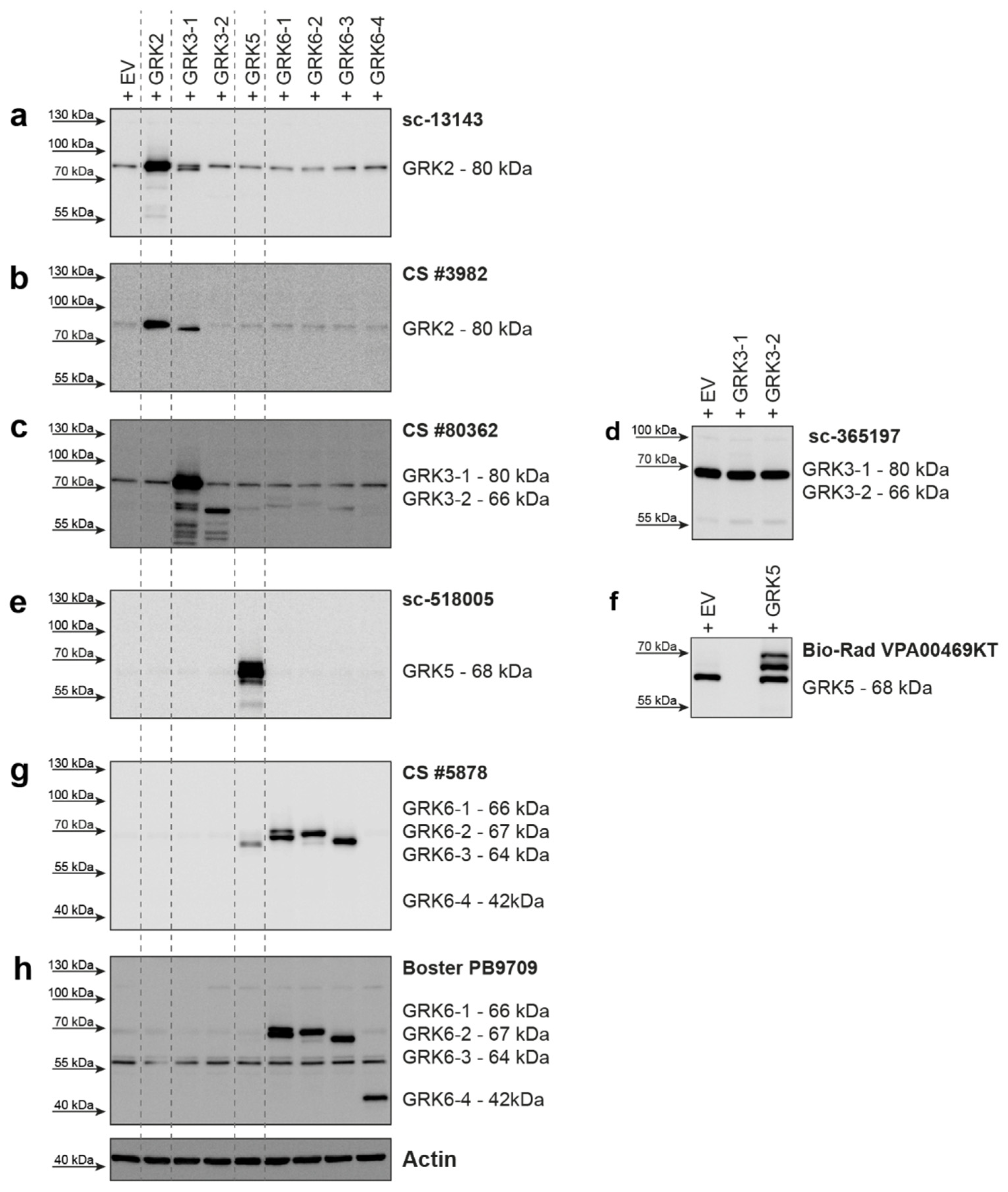
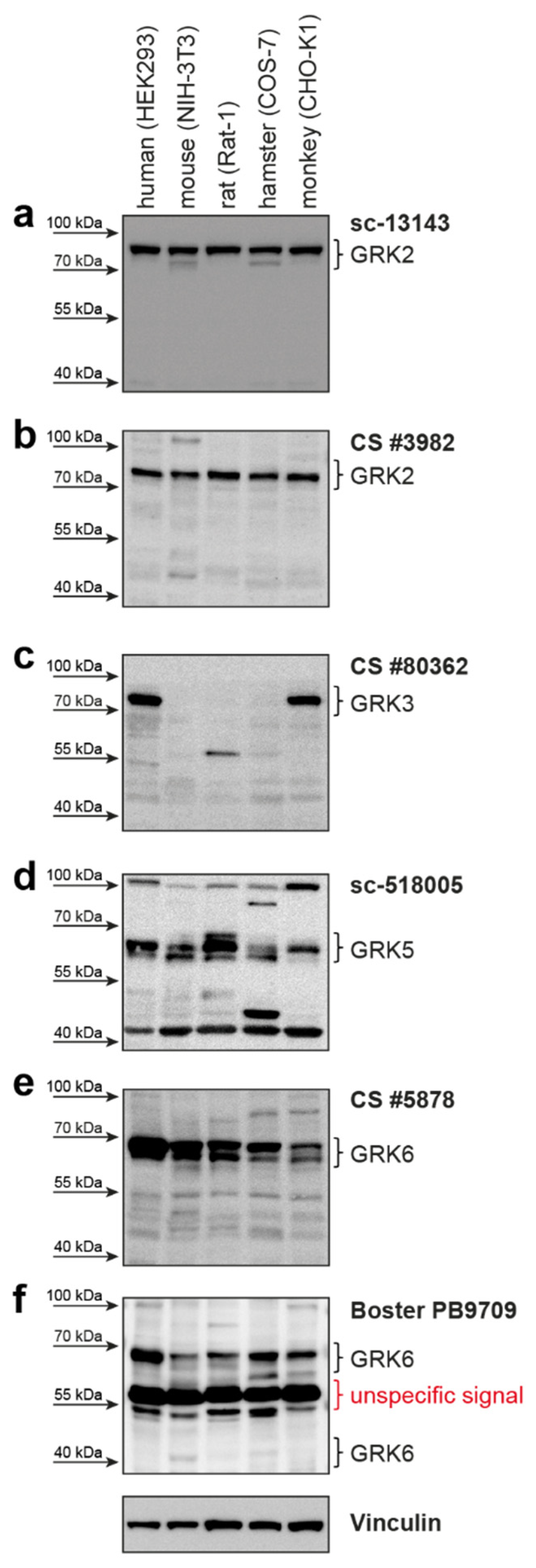
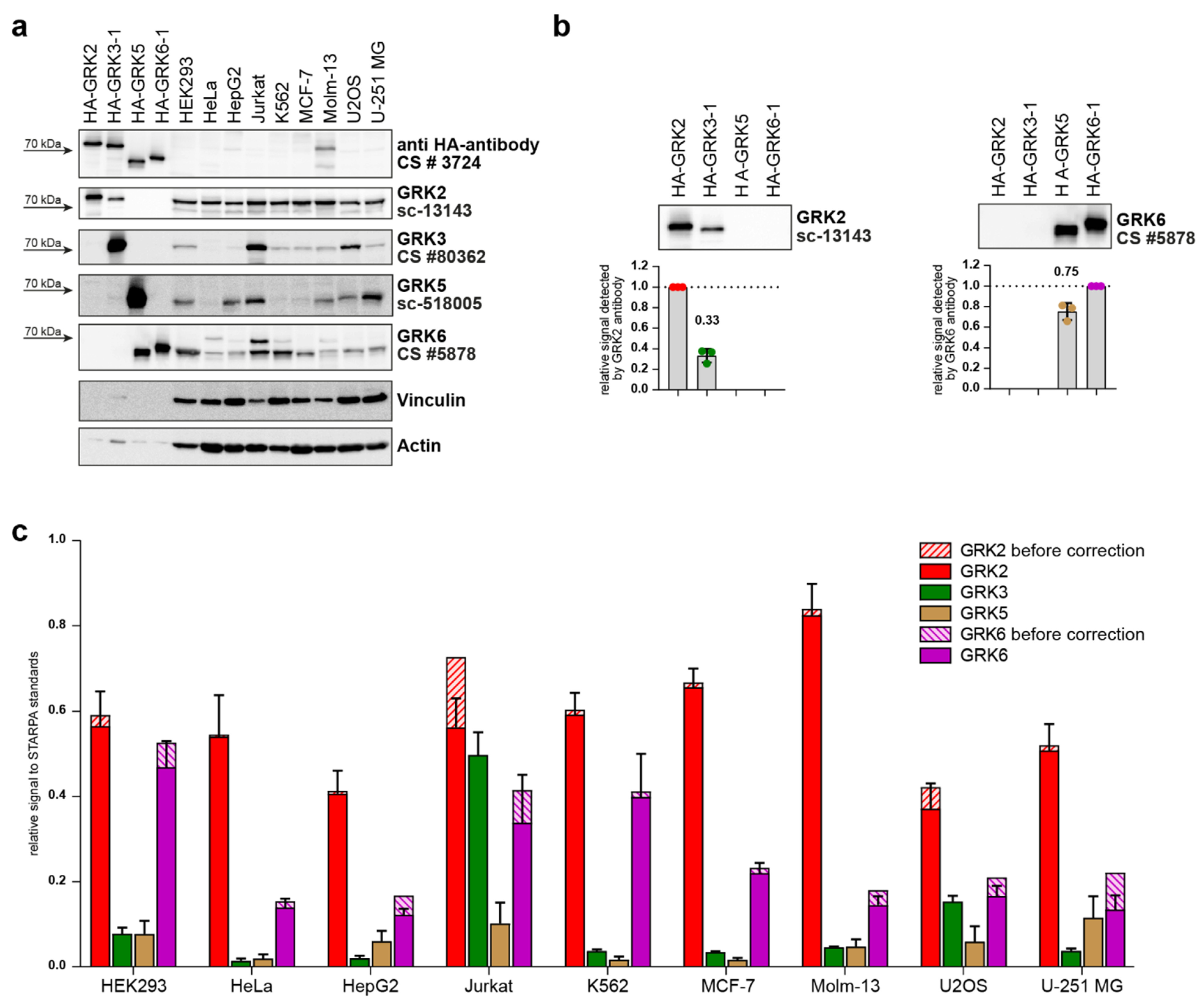
2. Results
3. Discussion
4. Materials and Methods
4.1. Expression Constructs
4.2. Cell Lines
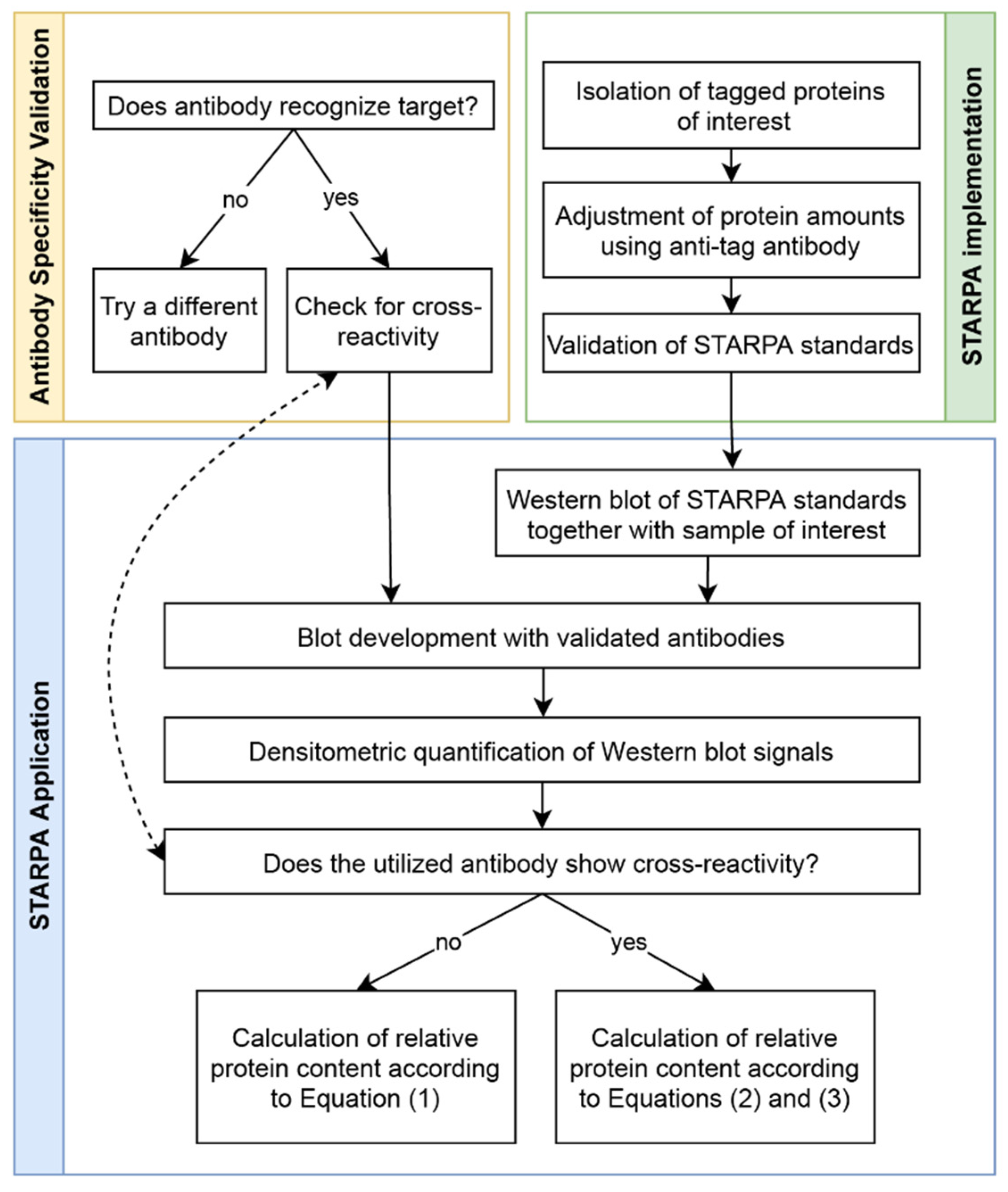
4.3. Antibody Validation
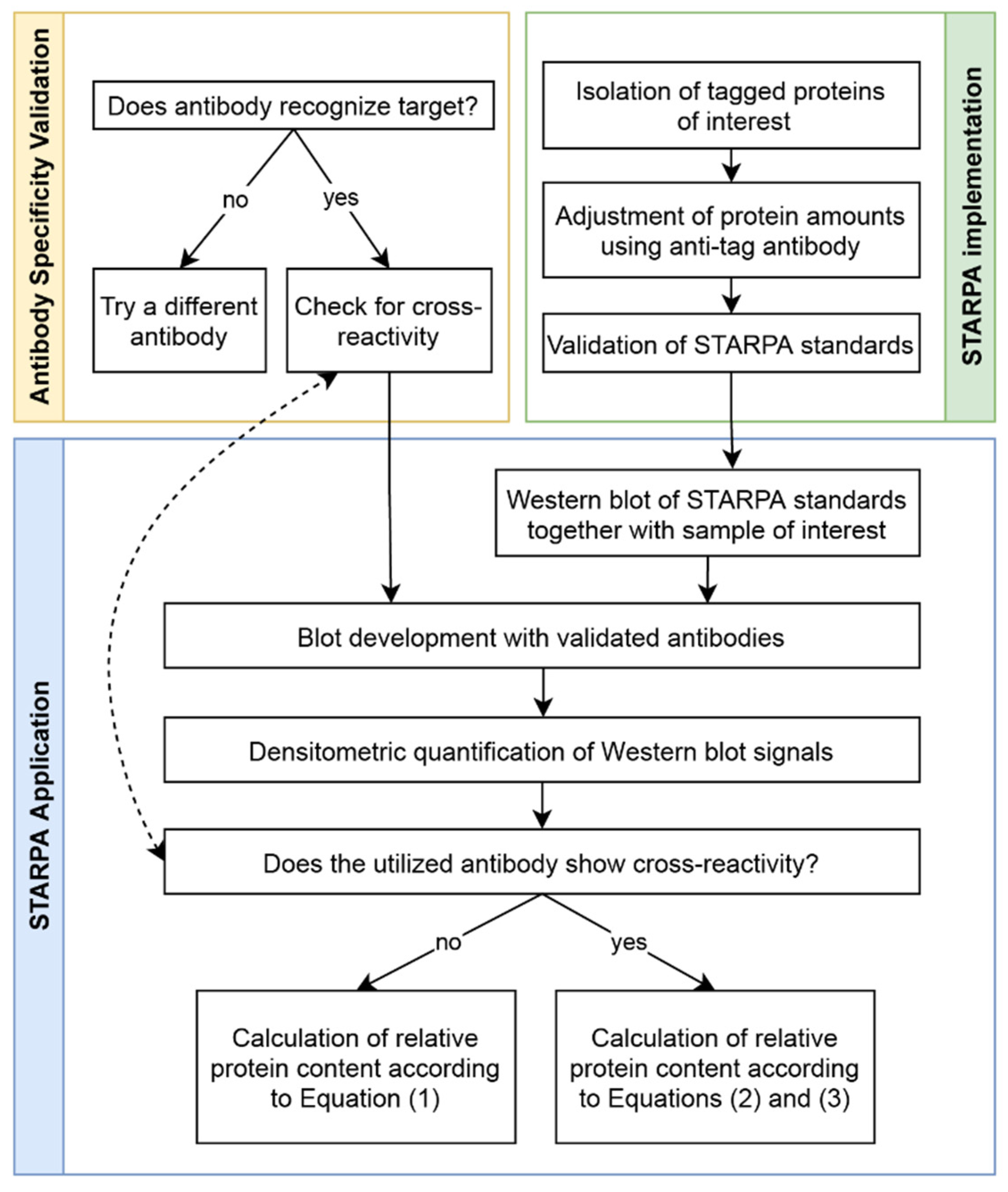
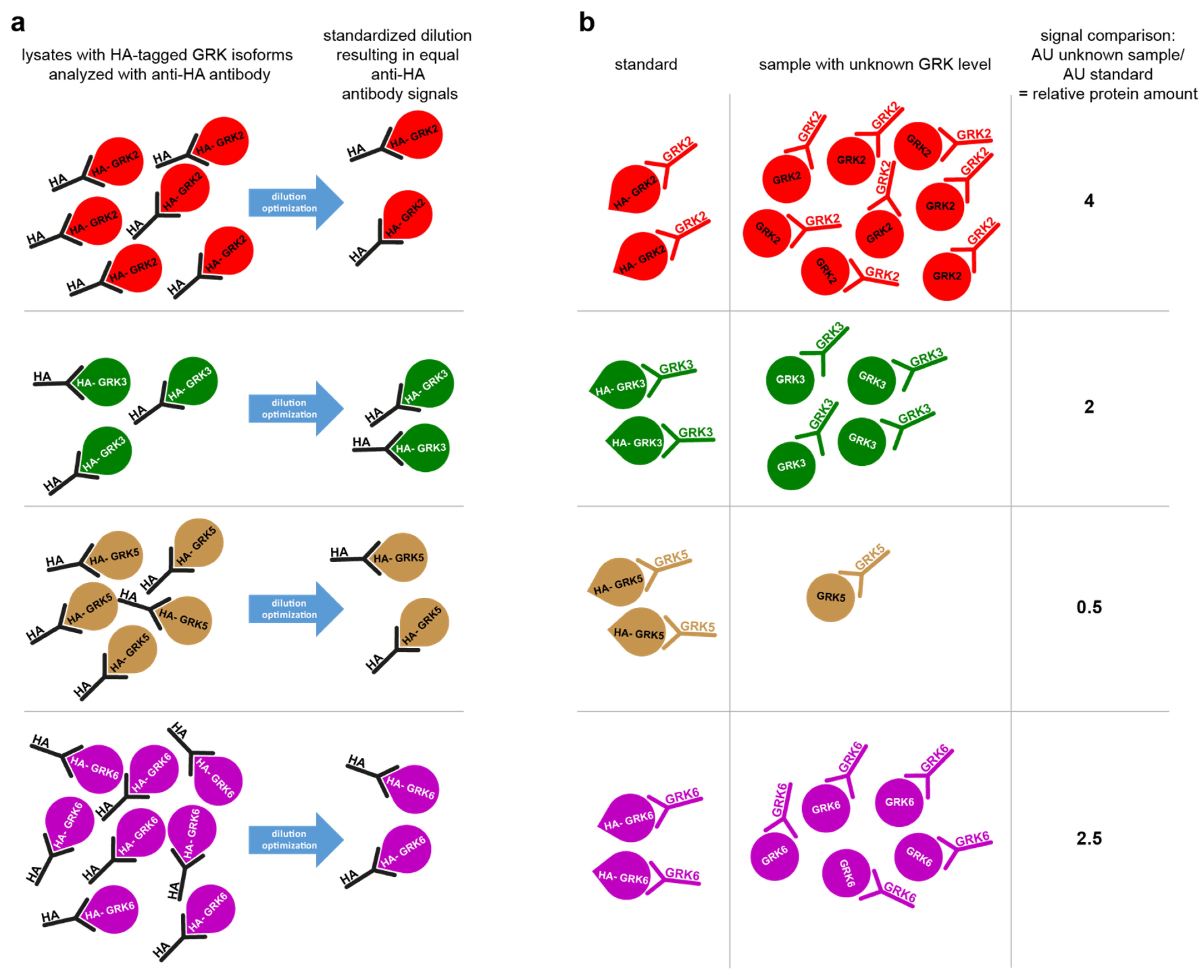
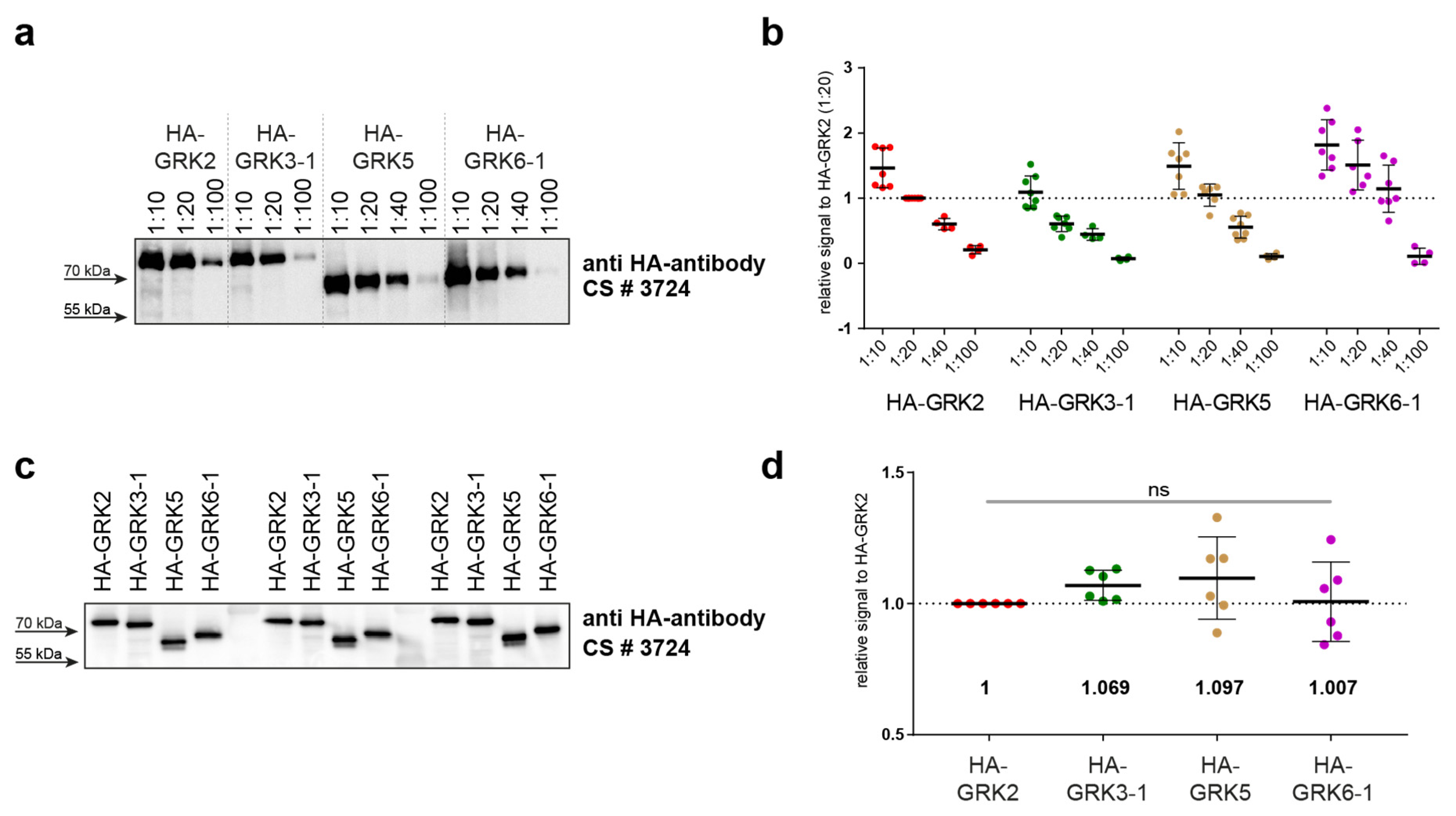
4.4. STARPA
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lohse, M.J.; Hoffmann, C. Arrestin Interactions with G Protein-Coupled Receptors. Antidepressants 2013, 219, 15–56. [Google Scholar] [CrossRef]
- Komolov, K.E.; Benovic, J.L. G protein-coupled receptor kinases: Past, present and future. Cell. Signal. 2017, 41, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, E.V.; Tesmer, J.J.; Mushegian, A.; Gurevich, V.V. G protein-coupled receptor kinases: More than just kinases and not only for GPCRs. Pharmacol. Ther. 2012, 133, 40–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penela, P.; Ribas, C.; Sánchez-Madrid, F.; Mayor, F. G protein-coupled receptor kinase 2 (GRK2) as a multifunctional signaling hub. Cell. Mol. Life Sci. 2019, 76, 4423–4446. [Google Scholar] [CrossRef] [Green Version]
- Martini, J.S.; Raake, P.; Vinge, L.E.; DeGeorge, B.R.; Chuprun, J.K.; Harris, D.M.; Gao, E.; Eckhart, A.D.; Pitcher, J.A.; Koch, W.J. Uncovering G protein-coupled receptor kinase-5 as a histone deacetylase kinase in the nucleus of cardiomyocytes. Proc. Natl. Acad. Sci. USA 2008, 105, 12457–12462. [Google Scholar] [CrossRef] [Green Version]
- Raveh, A.; Cooper, A.; Guy-David, L.; Reuveny, E. Nonenzymatic Rapid Control of GIRK Channel Function by a G Protein-Coupled Receptor Kinase. Cell 2010, 143, 750–760. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, N.; Gottardo, F.L.; Alonso, M.N.; Monczor, F.; Shayo, C.; Davio, C. Roles of Phosphorylation-dependent and -independent Mechanisms in the Regulation of Histamine H2 Receptor by G Protein-coupled Receptor Kinase 2. J. Biol. Chem. 2011, 286, 28697–28706. [Google Scholar] [CrossRef] [Green Version]
- Penela, P.; Murga, C.; Ribas, C.; Lafarga, V.F.M., Jr. The complex G protein-coupled receptor kinase 2 (GRK2) interactome unveils new physiopathological targets. Br. J. Pharmacol. 2010, 160, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Nogués, L.; Palacios-García, J.; Reglero, C.; Rivas, V.; Neves, M.; Ribas, C.; Penela, P.; Mayor, F. G protein-coupled receptor kinases (GRKs) in tumorigenesis and cancer progression: GPCR regulators and signaling hubs. Semin. Cancer Biol. 2018, 48, 78–90. [Google Scholar] [CrossRef]
- Leoratti, F.M.D.S.; Trevelin, S.C.; Cunha, F.Q.; Rocha, B.C.; Costa, P.A.C.; Gravina, H.; Tada, M.S.; Pereira, D.; Golenbock, D.T.; Antonelli, L.; et al. Neutrophil Paralysis in Plasmodium vivax Malaria. PLoS Neglected Trop. Dis. 2012, 6, e1710. [Google Scholar] [CrossRef]
- Murga, C.; Arcones, A.C.; Cruces-Sande, M.; Briones, A.M.; Salaices, M.F.M., Jr. G Protein-Coupled Receptor Kinase 2 (GRK2) as a Potential Therapeutic Target in Cardiovascular and Metabolic Diseases. Front. Pharmacol. 2019, 10, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.R.; Bychkov, E.; Gurevich, V.V.; Benovic, J.L.; Gurevich, E.V. Altered expression and subcellular distribution of GRK subtypes in the dopamine-depleted rat basal ganglia is not normalized by l-DOPA treatment. J. Neurochem. 2007, 104, 1622–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthees, E.S.F.; Haider, R.S.; Hoffmann, C.; Drube, J. Differential Regulation of GPCRs—Are GRK Expression Levels the Key? Front. Cell Dev. Biol. 2021, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penela, P.; Ribas, C.; Mayor, F. Mechanisms of regulation of the expression and function of G protein-coupled receptor kinases. Cell. Signal. 2003, 15, 973–981. [Google Scholar] [CrossRef]
- Gilda, J.E.; Ghosh, R.; Cheah, J.X.; West, T.M.; Bodine, S.C.; Gomes, A.V. Western Blotting Inaccuracies with Unverified Antibodies: Need for a Western Blotting Minimal Reporting Standard (WBMRS). PLoS ONE 2015, 10, e0135392. [Google Scholar] [CrossRef] [PubMed]
- Rosell, A.; Moser, B.; Hisada, Y.; Bs, R.C.; Lian, G.; Yang, Y.; Flick, M.J.; Mackman, N. Evaluation of different commercial antibodies for their ability to detect human and mouse tissue factor by western blotting. Res. Pract. Thromb. Haemost. 2020, 4, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Qi, L.; Chiang, H.-C.; Yuan, B.; Li, R.; Hu, Y. BRCA1 Antibodies Matter. Int. J. Biol. Sci. 2021, 17, 3239–3254. [Google Scholar] [CrossRef]
- Egelhofer, A.T.; Minoda, A.; Klugman, S.; Lee, K.; Kolasinska-Zwierz, P.; Alekseyenko, A.; Cheung, M.-S.; Day, D.S.; Gadel, S.; Gorchakov, A.; et al. An assessment of histone-modification antibody quality. Nat. Struct. Mol. Biol. 2010, 18, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Bordeaux, J.; Welsh, A.W.; Agarwal, S.; Killiam, E.; Baquero, M.T.; Hanna, J.A.; Anagnostou, V.K.; Rimm, D.L. Antibody validation. Biotechniques 2010, 48, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Michel, M.C.; Wieland, T.; Tsujimoto, G. How reliable are G-protein-coupled receptor antibodies? Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2009, 379, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palczewski, K. GTP-Binding-Protein-Coupled Receptor Kinases Two Mechanistic Models. JBIC J. Biol. Inorg. Chem. 1997, 248, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Pronin, A.N.; Benovic, J.L. Regulation of the G Protein-coupled Receptor Kinase GRK5 by Protein Kinase C. J. Biol. Chem. 1997, 272, 3806–3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drube, J.; Haider, R.S.; Matthees, E.S.F.; Reichel, M.; Zeiner, J.; Fritzwanker, S.; Ziegler, C.; Barz, S.; Klement, L.; Filor, J.; et al. GPCR kinase knockout cells reveal the impact of individual GRKs on arrestin binding and GPCR regulation. Nat. Commun. 2022; in press. [Google Scholar] [CrossRef]
- Premont, R.; Macrae, A.D.; Aparicio, S.A.; Kendall, H.E.; Welch, J.E.; Lefkowitz, R.J. The GRK4 Subfamily of G Protein-coupled Receptor Kinases. J. Biol. Chem. 1999, 274, 29381–29389. [Google Scholar] [CrossRef] [Green Version]
- Penela, P.; Rivas, V.; Salcedo, A.; Mayor, F., Jr. G protein-coupled receptor kinase 2 (GRK2) modulation and cell cycle progression. Proc. Natl. Acad. Sci. USA 2009, 107, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Geraghty, R.J.; Capes-Davis, A.; Davis, J.M.; Downward, J.; Freshney, R.I.; Knezevic, I.; Lovell-Badge, R.; Masters, J.R.W.; Meredith, J.; Stacey, G.N.; et al. Guidelines for the use of cell lines in biomedical research. Br. J. Cancer 2014, 111, 1021–1046. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Ahn, S.; Ren, X.-R.; Whalen, E.J.; Reiter, E.; Wei, H.; Lefkowitz, R.J. Functional antagonism of different G protein-coupled receptor kinases for -arrestin-mediated angiotensin II receptor signaling. Proc. Natl. Acad. Sci. USA 2005, 102, 1442–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.-R.; Reiter, E.; Ahn, S.; Kim, J.; Chen, W.; Lefkowitz, R.J. Different G protein-coupled receptor kinases govern G protein and -arrestin-mediated signaling of V2 vasopressin receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 1448–1453. [Google Scholar] [CrossRef] [Green Version]
- Pfleger, J.; Gresham, K.; Koch, W.J. G protein-coupled receptor kinases as therapeutic targets in the heart. Nat. Rev. Cardiol. 2019, 16, 612–622. [Google Scholar] [CrossRef]
- Lu, R.-M.; Hwang, Y.-C.; Liu, I.-J.; Lee, C.-C.; Tsai, H.-Z.; Li, H.-J.; Wu, H.-C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef]
- Kaur, H.; Carvalho, J.; Looso, M.; Singh, P.; Chennupati, R.; Preussner, J.; Günther, S.; Albarran-Juarez, J.; Tischner, D.; Classen, S.; et al. Single-cell profiling reveals heterogeneity and functional patterning of GPCR expression in the vascular system. Nat. Commun. 2017, 8, 15700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriram, K.; Moyung, K.; Corriden, R.; Carter, H.; Insel, P.A. GPCRs show widespread differential mRNA expression and frequent mutation and copy number variation in solid tumors. PLoS Biol. 2019, 17, e3000434. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of aa and MW | Supplier’s Information | Our Findings | |||||
|---|---|---|---|---|---|---|---|
| Ordering no. and Species | Supplier | Antigen | Species Reactivity | Dilution | Performance | ||
| GRK2 | 689 aa 80 kDa | sc-13143 (C-9) mouse mc | Santa Cruz Biotechnology | raised against amino acids 468-689 of human GRK2 | human, mouse, rat | 1:500 | good detection of endogenous GRK2 in HEK293; cross-reactivity with GRK3 |
| GRK2 | 689 aa 80 kDa | CS #3982 rabbit pc | Cell Signaling Technology | immunizing animals with a synthetic peptide corresponding to the amino-terminal residues of human GRK2 | human, mouse, rat, hamster, monkey | 1:1000 | cross-reactivity with GRK3; only weak detection of endogenous GRK2 in HEK293 cells |
| GRK3-1 | 688 aa 80 kDa | sc-365197 (C-11) mouse mc | Santa Cruz Biotechnology | raised against amino acids 646-688 of human GRK3 | human, mouse, rat | 1:250 | does not detect overexpressed human GRK3 isoforms—only background band |
| GRK3-2 | 575 aa 66 kDa | ||||||
| GRK3-1 | 688 aa 80 kDa | CS #80362 (D8G6V) rabbit mc | Cell Signaling Technology | immunizing animals with a synthetic peptide corresponding to residues surrounding Lys454 of human GRK3 | human, mouse | 1:250 | slight cross-reactivity with overexpressed GRK6 isoforms |
| GRK3-2 | 575 aa 66 kDa | ||||||
| GRK5 | 590 aa 68 kDa | sc-518005 (D-9) mouse mc | Santa Cruz Biotechnology | raised against amino acids 94-157 of human GRK5 | human, mouse, rat | 1:250 | good detection of endogenous GRK5 in HEK293 |
| GRK5 | 590 aa 68 kDa | VPA00469KT rabbit pc | Bio-Rad | synthetic peptide directed towards the middle region of human GRK5 | human, mouse | 1:1000 | strong background band; reported to supplier, was then discontinued |
| GRK6-1 | 576 aa 66 kDa | CS #5878 (D1A4) rabbit mc | Cell Signaling Technology | synthetic peptide corresponding to residues surrounding Glu89 of human GRK6 | human, mouse, rat | 1:1000 | cross-reactivity with GRK5 |
| GRK6-2 | 589 aa 67 kDa | ||||||
| GRK6-3 | 560 aa 64 kDa | ||||||
| GRK6-1 | 576 aa 66 kDa | PB9709 rabbit pc | Boster Biological Technology | C-terminus of human GRK6 (amino acids 382-417) | human, rat | 1:1000 | strong background around 55kDa; can detect human isoform 6-4 |
| GRK6-2 | 589 aa 67 kDa | ||||||
| GRK6-3 | 560 aa 64 kDa | ||||||
| GRK6-4 | 366 aa 42 kDa | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reichel, M.; Weitzel, V.; Klement, L.; Hoffmann, C.; Drube, J. Suitability of GRK Antibodies for Individual Detection and Quantification of GRK Isoforms in Western Blots. Int. J. Mol. Sci. 2022, 23, 1195. https://doi.org/10.3390/ijms23031195
Reichel M, Weitzel V, Klement L, Hoffmann C, Drube J. Suitability of GRK Antibodies for Individual Detection and Quantification of GRK Isoforms in Western Blots. International Journal of Molecular Sciences. 2022; 23(3):1195. https://doi.org/10.3390/ijms23031195
Chicago/Turabian StyleReichel, Mona, Verena Weitzel, Laura Klement, Carsten Hoffmann, and Julia Drube. 2022. "Suitability of GRK Antibodies for Individual Detection and Quantification of GRK Isoforms in Western Blots" International Journal of Molecular Sciences 23, no. 3: 1195. https://doi.org/10.3390/ijms23031195
APA StyleReichel, M., Weitzel, V., Klement, L., Hoffmann, C., & Drube, J. (2022). Suitability of GRK Antibodies for Individual Detection and Quantification of GRK Isoforms in Western Blots. International Journal of Molecular Sciences, 23(3), 1195. https://doi.org/10.3390/ijms23031195