
Gene Expression Linked to Reepithelialization of Human Skin Wounds

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Determination of mRNAs by NanoString
2.2. Differentially Expressed Genes (DEGs) in Human Epidermal Wounds
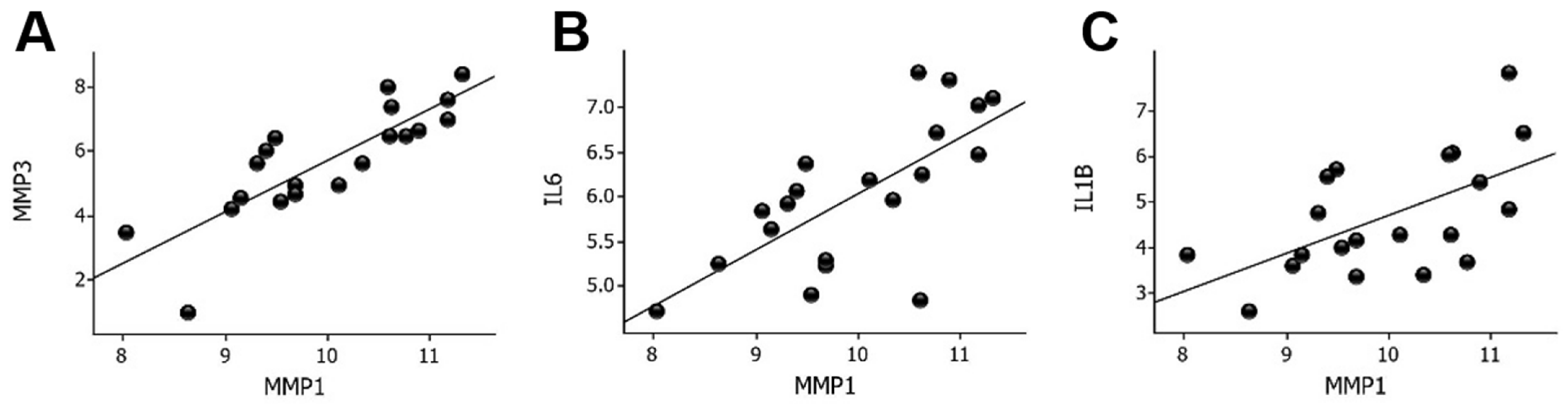
2.3. MMP1 mRNA Correlations in Wounds
2.4. Effects of Cytokines on MMP1, MMP3 and TIMP1 Gene Expression and Secretion of MMP-1 and TIMP-1 into the Medium of Normal Human Epidermal Keratinocytes (NHEKs) and Normal Human Dermal Fibroblasts (NHDFs)
2.4.1. MMP1 and TIMP1 mRNA and MMP-1 and TIMP-1 Protein Levels
2.4.2. MMP3 mRNA Levels
2.5. TIMP-1 and Collagen I Protein Expression
2.6. Proliferation Markers in Wounds
3. Discussion
4. Materials and Methods
4.1. Ethical Statements
4.2. Participants
4.3. Induction of Epidermal (Suction Blister) Wounds, Treatment and Tissue Procedures
4.4. Macrodissection and Isolation of RNA from FFPE Wound and Skin Compartments
4.5. Design of Gene Panel and nCounter Analyses
4.6. Immunohistochemical Analysis of TIMP-1 and Collagen I
4.7. Studies in NHEKs and NHDFs
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BrdU | bromodeoxyuridine |
| DEG | differentially expressed gene |
| ECM | extracellular matrix |
| ELISA | enzyme-linked immunosorbent assay |
| FBS | fetal bovine serum |
| FC | fold change |
| FDR | false discovery rate |
| FFPE | formalin-fixed, paraffin-embedded |
| KBM | keratinocyte basal medium |
| LDH | lactate dehydrogenase |
| NHDF | normal human dermal fibroblast |
| NHEK | normal human epidermal keratinocyte |
| PBS | phosphate-buffered saline |
| RT–qPCR | reverse transcription quantitative polymerase chain reaction |
| STSG | split-thickness skin graft |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank ID | |
|---|---|---|
| ACTC1 | NM_005159.4 | actin, alpha, cardiac muscle 1 |
| ACVRL1 | NM_000020.1 | activin A receptor type II-like 1 |
| ADAM17 | NM_003183.4 | ADAM metallopeptidase domain 17 |
| ADORA2A | NM_000675.3 | adenosine A2a receptor |
| AGER | NM_001136.3 | advanced glycosylation end product-specific receptor |
| ANGPT1 | NM_001146.3 | angiopoietin 1 |
| BMP1 | NM_001199.1 | bone morphogenetic protein 1 |
| CCL2 | NM_002982.3 | chemokine (C-C motif) ligand 2 |
| CCL7 | NM_006273.2 | chemokine (C-C motif) ligand 7 |
| CD36 | NM_000072.3 | thrombospondin receptor |
| CD40LG | NM_000074.2 | CD40 ligand |
| CD59 | NM_000611.4 | complement regulatory protein |
| CDH1 | NM_004360.2 | E-cadherin (epithelial) |
| COL1A1 | NM_000088.3 | collagen type I, alpha 1 chain |
| COL3A1 | NM_000090.3 | collagen type III, alpha 1 chain |
| COL4A1 | NM_001845.4 | collagen, type IV, alpha 1 chain |
| COL4A3 | NM_000091.4 | collagen, type IV, alpha 3 (Goodpasture antigen) |
| COL5A2 | NM_000393.3 | collagen, type V, alpha 2 chain |
| COL5A3 | NM_015719.3 | collagen, type V, alpha 3 chain |
| COL7A1 | NM_000094.2 | collagen type VII, alpha 1 chain |
| COL14A1 | NM_021110.1 | collagen, type XIV, alpha 1 chain |
| COL17A1 | NM_000494.3 | collagen type XVII, alpha 1 chain |
| CSF2 | NM_000758.2 | colony stimulating factor 2 (granulocyte-macrophage) |
| CSF2RA | NM_006140.3 | colony stimulating factor 2 receptor alpha subunit |
| CSF3 | NM_000759.3 | colony stimulating factor 3 (granulocyte) |
| CTGF | NM_001901.2 | connective tissue growth factor |
| CTNNB1 | NM_001098210.1 | catenin (cadherin-associated protein), beta 1, 88 kDa |
| CTSG | NM_001911.2 | cathepsin G |
| CTSK | NM_000396.2 | cathepsin K |
| CTSV | NM_001333.3 | cathepsin V |
| CXCL1 | NM_001511.1 | chemokine (C-X-C motif) ligand 1 |
| CXCL2 | NM_002089.3 | chemokine (C-X-C motif) ligand 2 |
| CXCL5 | NM_002994.3 | chemokine (C-X-C motif) ligand 5 |
| CXCL8 | NM_000584.2 | chemokine (C-X-C motif) ligand 8 |
| CXCL11 | NM_005409.4 | chemokine (C-X-C motif) ligand 11 |
| CYR61 | NM_001554.3 | cysteine-rich angiogenic inducer 61 |
| DEFB4 | NM_004942.2 | beta-defensin 2 |
| EGF | NM_001963.4 | epidermal growth factor |
| EGFR | NM_201282.1 | epidermal growth factor receptor |
| EGR1 | NM_001964.2 | early growth response 1 |
| ENTPD1 | NM_001098175.1 | ectonucleoside triphosphate diphosphohydrolase 1 |
| EREG | NM_001432.2 | epiregulin |
| F3 | NM_001993.3 | coagulation factor III (thromboplastin, tissue factor) |
| F5 | NM_000130.2 | coagulation factor V (proaccelerin, labile factor) |
| F13A1 | NM_000129.3 | coagulation factor XIII, A1 polypeptide |
| FGF2 | NM_002006.4 | fibroblast growth factor 2 (basic) |
| FGF7 | NM_002009.3 | fibroblast growth factor 7 |
| FGF10 | NM_004465.1 | fibroblast growth factor 10 |
| FLG1 | NM_002016.1 | filaggrin |
| FLG2 | NM_001014342.2 | filaggrin family member 2 |
| FN1 | NM_212482.1 | fibronectin |
| GNAQ | NM_002072.2 | guanine nucleotide binding protein (G protein) |
| HBEGF | NM_001945.1 | heparin-binding EGF-like growth factor |
| HGF | NM_000601.4 | hepatocyte growth factor (hepapoietin A, scatter factor) |
| HIF1A | NM_001530.2 | hypoxia inducible factor 1 alpha subunit |
| IFNG | NM_000619.2 | interferon gamma |
| IGF1 | NM_000618.3 | insulin-like growth factor 1 (somatomedin C) |
| IL1A | NM_000575.3 | interleukin 1 alpha |
| IL1B | NM_000576.2 | interleukin 1 beta |
| IL2 | NM_000586.2 | interleukin 2 |
| IL4 | NM_000589.2 | interleukin 4 |
| IL6 | NM_000600.3 | interleukin 6 (interferon, beta 2) |
| IL6ST | NM_002184.2 | interleukin 6 signal transducer (gp130) |
| IL10 | NM_000572.2 | interleukin 10 |
| ITGA1 | NM_181501.1 | integrin, alpha 1 |
| ITGA2 | NM_002203.2 | integrin, alpha 2 (CD49B, alpha 2 subunit of VLA-2 receptor) |
| ITGA3 | NM_002204.2 | integrin, alpha 3 (antigen CD49C) |
| ITGA4 | NM_000885.4 | integrin, alpha 4 (antigen CD49D) |
| ITGA5 | NM_002205.2 | integrin, alpha 5 (fibronectin receptor, alpha polypeptide) |
| ITGA6 | NM_000210.1 | integrin, alpha 6 |
| ITGAV | NM_002210.2 | integrin, alpha V |
| ITGB1 | NM_002211.3 | integrin, beta 1 (fibronectin receptor, antigen CD29) |
| ITGB3 | NM_000212.2 | integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61) |
| ITGB5 | NM_002213.3 | integrin, beta 5 |
| ITGB6 | NM_000888.3 | integrin, beta 6 |
| IVL | NM_005547.2 | involucrin |
| KLK8 | NM_144507.1 | kallikrein-related peptidase 8 |
| KRT6A | NM_005554.3 | keratin 6A |
| KRT10 | NM_000421.3 | keratin 10 |
| LAMA3 | NM_000227.3 | laminin subunit alpha 3 |
| MAPK1 | NM_138957.2 | mitogen-activated protein kinase 1 |
| MAPK3 | NM_001040056.1 | mitogen-activated protein kinase 3 |
| MIF | NM_002415.1 | macrophage migration inhibitory factor |
| MKI67 | NM_002417.2 | marker of proliferation Ki-67 |
| MMP1 | NM_002421.3 | matrix metallopeptidase-1 (interstitial collagenase) |
| MMP2 | NM_004530.2 | matrix metallopeptidase-2 (gelatinase A) |
| MMP3 | NM_002422.3 | matrix metallopeptidase-3 |
| MMP7 | NM_002423.3 | matrix metallopeptidase-7 (matrilysin) |
| MMP8 | NM_002424.2 | matrix metallopeptidase-8 |
| MMP9 | NM_004994.2 | matrix metallopeptidase-9 (gelatinase B) |
| MMP10 | NM_002425.1 | matrix metallopeptidase-10 |
| MMP12 | NM_002426.3 | matrix metallopeptidase-12 |
| MMP13 | NM_002427.2 | matrix metallopeptidase-13 |
| MMP14 | NM_004995.2 | matrix metallopeptidase-14 |
| MMP28 | NM_001032278.1 | matrix metallopeptidase-28 |
| MT1A | NM_005946.2 | metallothionein 1A |
| MT2A | NM_005953.3 | metallothionein 2A |
| MYC | NM_002467.3 | MYC proto-oncogene, BHLH transcription factor |
| NF1 | NM_000267.2 | neurofibromin 1 |
| NGF | NM_002506.2 | nerve growth factor |
| PDGFA | NM_002607.5 | platelet-derived growth factor alpha polypeptide |
| PF4 | NM_002619.2 | platelet factor 4 |
| PI3 | NM_002638.3 | peptidase inhibitor 3 (elafin) |
| PI3K | NM_006218.2 | phosphatidylinositide 3-kinase |
| PLAT | NM_000931.2 | plasminogen activator, tissue |
| PLAU | NM_002658.2 | plasminogen activator, urokinase |
| PLAUR | NM_001005376.2 | plasminogen activator, urokinase receptor |
| POLR1B | NM_019014.3 | polymerase (RNA) I polypeptide B, 128 kDa |
| PTGS2 | NM_000963.1 | prostaglandin-endoperoxide synthase 2 |
| RAC1 | NM_006908.4 | ras-related C3 botulinum toxin substrate 1 (rho family) |
| RHOA | NM_001664.2 | ras homolog family member A |
| S100A7 | NM_002963.3 | psoriasin |
| S100A8 | NM_002964.3 | S100 calcium binding protein A8 |
| S100A9 | NM_002965.2 | S100 calcium binding protein A9 |
| SERPINB3 | NM_006919.2 | serpin family B member 3 (serine proteinase inhibitor) |
| SERPINE1 | NM_000602.2 | serpin peptidase inhibitor, clade E (plasminogen activator inhibitor type 1) |
| SLC30A1 | NM_021194.2 | solute carrier family 30 member 1 (ZnT1) |
| SLC39A2 | NM_014579.1 | solute carrier family 39 member 2 (ZIP2) |
| SLC39A4 | NM_017767.2 | solute carrier family 39 member 4 (ZIP4) |
| SOD1 | NM_000454.4 | superoxide dismutase [Cu-Zn] |
| STAT3 | NM_003150.3 | signal transducer and activator of transcription 3 |
| TFPI | NM_001032281.2 | tissue factor pathway inhibitor |
| TGFA | NM_003236.2 | transforming growth factor, alpha |
| TGFB1 | NM_000660.3 | transforming growth factor, beta 1 |
| TGFB2 | NM_003238.2 | transforming growth factor, beta 2 |
| TGFBR3 | NM_003243.3 | transforming growth factor, beta receptor III |
| THBD | NM_000361.2 | thrombomodulin |
| TIMP1 | NM_003254.2 | TIMP metallopeptidase inhibitor 1 |
| TIMP2 | NM_003255.4 | TIMP metallopeptidase inhibitor 2 |
| TIMP3 | NM_000362.4 | TIMP metallopeptidase inhibitor 3 |
| TLR4 | NM_138554.2 | toll-like receptor 4 |
| TMPRSS6 | NM_153609.2 | transmembrane protease, serine 6 |
| TNF | NM_000594.2 | tumor necrosis factor |
| VEGFA | NM_001025366.1 | vascular endothelial growth factor A |
| VTN | NM_000638.3 | vitronectin |
| VWF | NM_000552.3 | von Willebrand factor |
| WAS | NM_000377.2 | WASP actin nucleation promoting factor |
| WISP1 | NM_080838.1 | WNT1 inducible signaling pathway protein 1 |
| WNT5A | NM_003392.3 | wingless-type MMTV integration site family, member 5A |
| ABCF11 | NM_001090.2 | ATP-binding cassette, sub-family F (GCN20), member 1 |
| GUSB1 | NM_000181.3 | glucuronidase, beta |
| HPRT11 | NM_000194.1 | hypoxanthine phosphoribosyltransferase 1 |
| LDHA1 | NM_001165414.1 | lactate dehydrogenase A |
| PTEN1 | NM_000314.3 | phosphatase and tensin homolog |
| RPLP0 1 | NM_001002.3 | ribosomal protein, large, P0 |
References
- Ågren, M.S.; Chafranska, L.; Eriksen, J.O.; Forman, J.L.; Bjerrum, M.J.; Schjerling, P.; Larsen, H.F.; Cottarelli, E.; Jorgensen, L.N.; Gjerdrum, L.M.R. Spatial expression of metallothionein, matrix metalloproteinase-1 and Ki-67 in human epidermal wounds treated with zinc and determined by quantitative immunohistochemistry: A randomised double-blind trial. Eur. J. Cell Biol. 2021, 100, 151147. [Google Scholar] [CrossRef] [PubMed]
- Michopoulou, A.; Rousselle, P. How do epidermal matrix metalloproteinases support re-epithelialization during skin healing? Eur. J. Dermatol. 2015, 25 (Suppl. 1), 33–42. [Google Scholar] [CrossRef] [PubMed]
- Nuutila, K.; Siltanen, A.; Peura, M.; Bizik, J.; Kaartinen, I.; Kuokkanen, H.; Nieminen, T.; Harjula, A.; Aarnio, P.; Vuola, J.; et al. Human skin transcriptome during superficial cutaneous wound healing. Wound Repair Regen. 2012, 20, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Greco, J.A., 3rd; Pollins, A.C.; Boone, B.E.; Levy, S.E.; Nanney, L.B. A microarray analysis of temporal gene expression profiles in thermally injured human skin. Burns 2010, 36, 192–204. [Google Scholar] [CrossRef]
- Deonarine, K.; Panelli, M.C.; Stashower, M.E.; Jin, P.; Smith, K.; Slade, H.B.; Norwood, C.; Wang, E.; Marincola, F.M.; Stroncek, D.F. Gene expression profiling of cutaneous wound healing. J. Transl. Med. 2007, 5, 11. [Google Scholar] [CrossRef]
- Kutlu, N.; Svedman, P. The superficial dermal microcirculation in suction blister wounds on healthy volunteers. Vasc. Endovasc. Surg. 1992, 26, 200–212. [Google Scholar] [CrossRef]
- Oikarinen, A.; Kylmaniemi, M.; Autio-Harmainen, H.; Autio, P.; Salo, T. Demonstration of 72-kDa and 92-kDa forms of type IV collagenase in human skin: Variable expression in various blistering diseases, induction during re-epithelialization, and decrease by topical glucocorticoids. J. Investig. Dermatol. 1993, 101, 205–210. [Google Scholar] [CrossRef][Green Version]
- Lévy, J.J.; von Rosen, J.; Gassmuller, J.; Kleine Kuhlmann, R.; Lange, L. Validation of an in vivo wound healing model for the quantification of pharmacological effects on epidermal regeneration. Dermatology 1995, 190, 136–141. [Google Scholar] [CrossRef]
- Glaser, R.; Kiecolt-Glaser, J.K.; Marucha, P.T.; MacCallum, R.C.; Laskowski, B.F.; Malarkey, W.B. Stress-related changes in proinflammatory cytokine production in wounds. Arch. Gen. Psychiatry 1999, 56, 450–456. [Google Scholar] [CrossRef]
- Leivo, T.; Kiistala, U.; Vesterinen, M.; Owaribe, K.; Burgeson, R.E.; Virtanen, I.; Oikarinen, A. Re-epithelialization rate and protein expression in the suction-induced wound model: Comparison between intact blisters, open wounds and calcipotriol-pretreated open wounds. Br. J. Dermatol. 2000, 142, 991–1002. [Google Scholar] [CrossRef]
- Ågren, M.S.; Mirastschijski, U.; Karlsmark, T.; Saarialho-Kere, U.K. Topical synthetic inhibitor of matrix metalloproteinases delays epidermal regeneration of human wounds. Exp. Dermatol. 2001, 10, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Koivukangas, V.; Oikarinen, A. Suction blister model of wound healing. Methods Mol. Med. 2003, 78, 255–261. [Google Scholar] [PubMed]
- Kottner, J.; Hillmann, K.; Fimmel, S.; Seite, S.; Blume-Peytavi, U. Characterisation of epidermal regeneration in vivo: A 60-day follow-up study. J. Wound Care 2013, 22, 395–400. [Google Scholar] [CrossRef]
- Larsen, H.F.; Ahlström, M.G.; Gjerdrum, L.M.R.; Mogensen, M.; Ghathian, K.; Calum, H.; Sørensen, A.L.; Forman, J.L.; Vandeven, M.; Holerca, M.N.; et al. Noninvasive measurement of reepithelialization and microvascularity of suction-blister wounds with benchmarking to histology. Wound Repair Regen. 2017, 25, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Ahlström, M.G.; Gjerdrum, L.M.R.; Larsen, H.F.; Fuchs, C.; Sørensen, A.L.; Forman, J.L.; Ågren, M.S.; Mogensen, M. Suction blister lesions and epithelialization monitored by optical coherence tomography. Skin Res. Technol. 2018, 24, 65–72. [Google Scholar] [CrossRef]
- Kjaer, M.; Frederiksen, A.K.S.; Nissen, N.I.; Willumsen, N.; van Hall, G.; Jorgensen, L.N.; Andersen, J.R.; Ågren, M.S. Multi-nutrient supplementation increases collagen synthesis during early wound repair in a randomized controlled trial in patients with inguinal hernia. J. Nutr. 2020, 150, 792–799. [Google Scholar] [CrossRef]
- Burian, E.A.; Sabah, L.; Kirketerp-Møller, K.; Gundersen, G.; Ågren, M.S. Effect of stabilized hypochlorous acid on re-epithelialization and bacterial bioburden in acute wounds: A randomized controlled trial in healthy volunteers. Acta Derm. Venereol. 2022, 102, adv00727. [Google Scholar] [CrossRef]
- Reis, P.P.; Waldron, L.; Goswami, R.S.; Xu, W.; Xuan, Y.; Perez-Ordonez, B.; Gullane, P.; Irish, J.; Jurisica, I.; Kamel-Reid, S. mRNA transcript quantification in archival samples using multiplexed, color-coded probes. BMC Biotechnol. 2011, 11, 46. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B. The validity and practicality of sun-reactive skin types I through VI. Arch. Dermatol. 1988, 124, 869–871. [Google Scholar] [CrossRef]
- Kuhns, D.B.; DeCarlo, E.; Hawk, D.M.; Gallin, J.I. Dynamics of the cellular and humoral components of the inflammatory response elicited in skin blisters in humans. J. Clin. Investig. 1992, 89, 1734–1740. [Google Scholar] [CrossRef]
- McDaniel, J.C.; Belury, M.; Ahijevych, K.; Blakely, W. Omega-3 fatty acids effect on wound healing. Wound Repair Regen. 2008, 16, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Mihara, M.; Moriya, Y.; Ohsugi, Y. IL-6-soluble IL-6 receptor complex inhibits the proliferation of dermal fibroblasts. Int. J. Immunopharmacol. 1996, 18, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; Steenfos, H.H.; Dabelsteen, S.; Hansen, J.B.; Dabelsteen, E. Proliferation and mitogenic response to PDGF-BB of fibroblasts isolated from chronic venous leg ulcers is ulcer-age dependent. J. Investig. Dermatol. 1999, 112, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Türlü, C.; Willumsen, N.; Marando, D.; Schjerling, P.; Biskup, E.; Hannibal, J.; Jorgensen, L.N.; Ågren, M.S. A human cellular model for colorectal anastomotic repair: The effect of localization and transforming growth factor-β1 treatment on collagen deposition and biomarkers. Int. J. Mol. Sci. 2021, 22, 1616. [Google Scholar] [CrossRef] [PubMed]
- Utani, A.; Momota, Y.; Endo, H.; Kasuya, Y.; Beck, K.; Suzuki, N.; Nomizu, M.; Shinkai, H. Laminin alpha 3 LG4 module induces matrix metalloproteinase-1 through mitogen-activated protein kinase signaling. J. Biol. Chem. 2003, 278, 34483–34490. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Kratz, G.; Haegerstrand, A.; Ståhle-Bäckdahl, M. Collagenase expression is rapidly induced in wound-edge keratinocytes after acute injury in human skin, persists during healing, and stops at re-epithelialization. J. Investig. Dermatol. 1995, 104, 479–483. [Google Scholar] [CrossRef]
- Vincenti, M.P.; Brinckerhoff, C.E. Transcriptional regulation of collagenase (MMP-1, MMP-13) genes in arthritis: Integration of complex signaling pathways for the recruitment of gene-specific transcription factors. Arthritis Res. Ther. 2002, 4, 157–164. [Google Scholar] [CrossRef]
- Saarialho-Kere, U.K.; Vaalamo, M.; Airola, K.; Niemi, K.M.; Oikarinen, A.I.; Parks, W.C. Interstitial collagenase is expressed by keratinocytes that are actively involved in reepithelialization in blistering skin disease. J. Investig. Dermatol. 1995, 104, 982–988. [Google Scholar] [CrossRef]
- Kitanaka, N.; Nakano, R.; Sugiura, K.; Kitanaka, T.; Namba, S.; Konno, T.; Nakayama, T.; Sugiya, H. Interleukin-1beta promotes interleulin-6 expression via ERK1/2 signaling pathway in canine dermal fibroblasts. PLoS ONE 2019, 14, e0220262. [Google Scholar] [CrossRef]
- Dumin, J.A.; Dickeson, S.K.; Stricker, T.P.; Bhattacharyya-Pakrasi, M.; Roby, J.D.; Santoro, S.A.; Parks, W.C. Pro-collagenase-1 (matrix metalloproteinase-1) binds the alpha(2)beta(1) integrin upon release from keratinocytes migrating on type I collagen. J. Biol. Chem. 2001, 276, 29368–29374. [Google Scholar] [CrossRef]
- Li, W.; Henry, G.; Fan, J.; Bandyopadhyay, B.; Pang, K.; Garner, W.; Chen, M.; Woodley, D.T. Signals that initiate, augment, and provide directionality for human keratinocyte motility. J. Investig. Dermatol. 2004, 123, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; auf dem Keller, U. Matrix metalloproteinases: How much can they do? Int. J. Mol. Sci. 2020, 21, 2678. [Google Scholar] [CrossRef] [PubMed]
- Saus, J.; Quinones, S.; Otani, Y.; Nagase, H.; Harris, E.D., Jr.; Kurkinen, M. The complete primary structure of human matrix metalloproteinase-3. Identity with stromelysin. J. Biol. Chem. 1988, 263, 6742–6745. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; Schnabel, R.; Christensen, L.H.; Mirastschijski, U. Tumor necrosis factor-alpha-accelerated degradation of type I collagen in human skin is associated with elevated matrix metalloproteinase (MMP)-1 and MMP-3 ex vivo. Eur. J. Cell Biol. 2015, 94, 12–21. [Google Scholar] [CrossRef]
- Chakraborty, S.; Sampath, D.; Yu Lin, M.O.; Bilton, M.; Huang, C.K.; Nai, M.H.; Njah, K.; Goy, P.A.; Wang, C.C.; Guccione, E.; et al. Agrin-Matrix Metalloproteinase-12 axis confers a mechanically competent microenvironment in skin wound healing. Nat. Commun. 2021, 12, 6349. [Google Scholar] [CrossRef]
- Salonurmi, T.; Parikka, M.; Kontusaari, S.; Pirilä, E.; Munaut, C.; Salo, T.; Tryggvason, K. Overexpression of TIMP-1 under the MMP-9 promoter interferes with wound healing in transgenic mice. Cell Tissue Res. 2004, 315, 27–37. [Google Scholar] [CrossRef]
- Aplin, A.C.; Zhu, W.H.; Fogel, E.; Nicosia, R.F. Vascular regression and survival are differentially regulated by MT1-MMP and TIMPs in the aortic ring model of angiogenesis. Am. J. Physiol. Cell Physiol. 2009, 297, C471–C480. [Google Scholar] [CrossRef]
- Emonard, H.; Munaut, C.; Melin, M.; Lortat-Jacob, H.; Grimaud, J.A. Interleukin-6 does not regulate interstitial collagenase, stromelysin and tissue inhibitor of metalloproteinases synthesis by cultured human fibroblasts. Matrix 1992, 12, 471–474. [Google Scholar] [CrossRef]
- Han, R.; Smith, T.J. Induction by IL-1 beta of tissue inhibitor of metalloproteinase-1 in human orbital fibroblasts: Modulation of gene promoter activity by IL-4 and IFN-gamma. J. Immunol. 2005, 174, 3072–3079. [Google Scholar] [CrossRef]
- Rømer, J.; Bugge, T.H.; Pyke, C.; Lund, L.R.; Flick, M.J.; Degen, J.L.; Dano, K. Impaired wound healing in mice with a disrupted plasminogen gene. Nat. Med. 1996, 2, 287–292. [Google Scholar] [CrossRef]
- Lund, L.R.; Rømer, J.; Bugge, T.H.; Nielsen, B.S.; Frandsen, T.L.; Degen, J.L.; Stephens, R.W.; Danø, K. Functional overlap between two classes of matrix-degrading proteases in wound healing. EMBO J. 1999, 18, 4645–4656. [Google Scholar] [CrossRef] [PubMed]
- Rømer, J.; Lund, L.R.; Eriksen, J.; Ralfkiaer, E.; Zeheb, R.; Gelehrter, T.D.; Danø, K.; Kristensen, P. Differential expression of urokinase-type plasminogen activator and its type-1 inhibitor during healing of mouse skin wounds. J. Investig. Dermatol. 1991, 97, 803–811. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Gerasi, L.; Blasi, F. uPAR-deficient mouse keratinocytes fail to produce EGFR-dependent laminin-5, affecting migration in vivo and in vitro. J. Cell. Sci. 2008, 121 Pt 23, 3922–3932. [Google Scholar] [CrossRef] [PubMed]
- Simone, T.M.; Longmate, W.M.; Law, B.K.; Higgins, P.J. Targeted inhibition of PAI-1 activity impairs epithelial migration and wound closure following cutaneous injury. Adv. Wound Care 2015, 4, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, E.; Toksoy, A.; Goebeler, M.; Debus, S.; Brocker, E.B.; Gillitzer, R. Chemokines IL-8, GROalpha, MCP-1, IP-10, and Mig are sequentially and differentially expressed during phase-specific infiltration of leukocyte subsets in human wound healing. Am. J. Pathol. 1998, 153, 1849–1860. [Google Scholar] [CrossRef]
- Devalaraja, R.M.; Nanney, L.B.; Du, J.; Qian, Q.; Yu, Y.; Devalaraja, M.N.; Richmond, A. Delayed wound healing in CXCR2 knockout mice. J. Investig. Dermatol. 2000, 115, 234–244. [Google Scholar] [CrossRef]
- Müller-Decker, K.; Scholz, K.; Neufang, G.; Marks, F.; Furstenberger, G. Localization of prostaglandin-H synthase-1 and -2 in mouse skin: Implications for cutaneous function. Exp. Cell Res. 1998, 242, 84–91. [Google Scholar] [CrossRef]
- Stoll, S.W.; Rittie, L.; Johnson, J.L.; Elder, J.T. Heparin-binding EGF-like growth factor promotes epithelial-mesenchymal transition in human keratinocytes. J. Investig. Dermatol. 2012, 132, 2148–2157. [Google Scholar] [CrossRef]
- Du, H.; Zhou, Y.; Suo, Y.; Liang, X.; Chai, B.; Duan, R.; Huang, X.; Li, Q. CCN1 accelerates re-epithelialization by promoting keratinocyte migration and proliferation during cutaneous wound healing. Biochem. Biophys. Res. Commun. 2018, 505, 966–972. [Google Scholar] [CrossRef]
- Chmielowiec, J.; Borowiak, M.; Morkel, M.; Stradal, T.; Munz, B.; Werner, S.; Wehland, J.; Birchmeier, C.; Birchmeier, W. c-Met is essential for wound healing in the skin. J. Cell Biol. 2007, 177, 151–162. [Google Scholar] [CrossRef]
- Kiwanuka, E.; Hackl, F.; Caterson, E.J.; Nowinski, D.; Junker, J.P.; Gerdin, B.; Eriksson, E. CCN2 is transiently expressed by keratinocytes during re-epithelialization and regulates keratinocyte migration in vitro by the ras-MEK-ERK signaling pathway. J. Surg. Res. 2013, 185, e109–e119. [Google Scholar] [CrossRef]
- Marchese, C.; Chedid, M.; Dirsch, O.R.; Csaky, K.G.; Santanelli, F.; Latini, C.; LaRochelle, W.J.; Torrisi, M.R.; Aaronson, S.A. Modulation of keratinocyte growth factor and its receptor in reepithelializing human skin. J. Exp. Med. 1995, 182, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, R.; Sato, C.; Kurita, Y.; Ron, D.; Rubin, J.S.; Ogawa, H. Keratinocyte growth factor (FGF-7) stimulates migration and plasminogen activator activity of normal human keratinocytes. J. Investig. Dermatol. 1993, 101, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Nauroy, P.; Nyström, A. Kallikreins: Essential epidermal messengers for regulation of the skin microenvironment during homeostasis, repair and disease. Matrix Biol. Plus 2020, 6, 100019. [Google Scholar] [CrossRef] [PubMed]
- Kishibe, M.; Bando, Y.; Tanaka, T.; Ishida-Yamamoto, A.; Iizuka, H.; Yoshida, S. Kallikrein-related peptidase 8-dependent skin wound healing is associated with upregulation of kallikrein-related peptidase 6 and PAR2. J. Investig. Dermatol. 2012, 132, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; Phothong, N.; Burian, E.A.; Mogensen, M.; Haedersdal, M.; Jorgensen, L.N. Topical zinc oxide assessed in two human wound-healing models. Acta Derm. Venereol. 2021, 101, adv00465. [Google Scholar] [CrossRef]
- Nyström, A.; Velati, D.; Mittapalli, V.R.; Fritsch, A.; Kern, J.S.; Bruckner-Tuderman, L. Collagen VII plays a dual role in wound healing. J. Clin. Investig. 2013, 123, 3498–3509. [Google Scholar] [CrossRef]
- Saarialho-Kere, U.; Kerkela, E.; Jahkola, T.; Suomela, S.; Keski-Oja, J.; Lohi, J. Epilysin (MMP-28) expression is associated with cell proliferation during epithelial repair. J. Investig. Dermatol. 2002, 119, 14–21. [Google Scholar] [CrossRef]
- Inoue, Y.; Hasegawa, S.; Ban, S.; Yamada, T.; Date, Y.; Mizutani, H.; Nakata, S.; Tanaka, M.; Hirashima, N. ZIP2 protein, a zinc transporter, is associated with keratinocyte differentiation. J. Biol. Chem. 2014, 289, 21451–21462. [Google Scholar] [CrossRef]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122 Pt 9, 1285–1294. [Google Scholar] [CrossRef]
- Makino, T.; Mizawa, M.; Yamakoshi, T.; Takaishi, M.; Shimizu, T. Expression of filaggrin-2 protein in the epidermis of human skin diseases: A comparative analysis with filaggrin. Biochem. Biophys. Res. Commun. 2014, 449, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Choi, D.; Chun, Y.J.; Noh, M. Keratinocyte-derived IL-24 plays a role in the positive feedback regulation of epidermal inflammation in response to environmental and endogenous toxic stressors. Toxicol. Appl. Pharmacol. 2014, 280, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kirketerp-Møller, K.; Doerfler, P.; Schoeffmann, N.; Wolff-Winiski, B.; Niazi, O.; Pless, V.; Karlsmark, T.; Ågren, M.S. Biomarkers of skin graft healing in venous leg ulcers. Acta Derm. Venereol. 2022, 102, adv00749. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]






| Cell Type | Control | IL-6 | IL-1β | IL-1β + IL-6 | p Value 1 |
|---|---|---|---|---|---|
| NHEKs | 25.9 ± 7.4 | 20.9 ± 6.3 | 19.1 ± 6.5 | 24.3 ± 8.4 | 0.370 |
| NHDFs | 14.7 ± 0.6 | 14.5 ± 0.9 | 13.7 ± 0.8 | 14.2 ± 1.5 | 0.377 |
| Gene | GenBank ID | Sense (Forward) | Antisense (Reverse) |
|---|---|---|---|
| MMP1 | NM_002421.4 | CGAATTTGCCGACAGAGATGAAG | GGGAAGCCAAAGGAGCTGTAGATG |
| MMP3 | NM_002422.5 | GATCCTGCTTTGTCCTTTGATGCTGT | CTGAGGGATTTGCGCCAAAAGTG |
| TIMP1 | NM_003254.3 | CGGGGCTTCACCAAGACCTACA | TGGTCCGTCCACAAGCAATGA |
| GAPDH | NM_002046.4 | CCTCCTGCACCACCAACTGCTT | GAGGGGCCATCCACAGTCTTCT |
| RPLP0 | NM_053275.3 | GGAAACTCTGCATTCTCGCTTCCT | CCAGGACTCGTTTGTACCCGTTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ågren, M.S.; Litman, T.; Eriksen, J.O.; Schjerling, P.; Bzorek, M.; Gjerdrum, L.M.R. Gene Expression Linked to Reepithelialization of Human Skin Wounds. Int. J. Mol. Sci. 2022, 23, 15746. https://doi.org/10.3390/ijms232415746
Ågren MS, Litman T, Eriksen JO, Schjerling P, Bzorek M, Gjerdrum LMR. Gene Expression Linked to Reepithelialization of Human Skin Wounds. International Journal of Molecular Sciences. 2022; 23(24):15746. https://doi.org/10.3390/ijms232415746
Chicago/Turabian StyleÅgren, Magnus S., Thomas Litman, Jens Ole Eriksen, Peter Schjerling, Michael Bzorek, and Lise Mette Rahbek Gjerdrum. 2022. "Gene Expression Linked to Reepithelialization of Human Skin Wounds" International Journal of Molecular Sciences 23, no. 24: 15746. https://doi.org/10.3390/ijms232415746
APA StyleÅgren, M. S., Litman, T., Eriksen, J. O., Schjerling, P., Bzorek, M., & Gjerdrum, L. M. R. (2022). Gene Expression Linked to Reepithelialization of Human Skin Wounds. International Journal of Molecular Sciences, 23(24), 15746. https://doi.org/10.3390/ijms232415746