
Elusive Trans-Acting Factors Which Operate with Type I (Poliovirus-like) IRES Elements


{kind=link}
Abstract
1. Introduction
1.1. The Concept of IRES Trans-Acting Factors
1.2. ITAFs That Interact with Poliovirus mRNA under Physiological Conditions
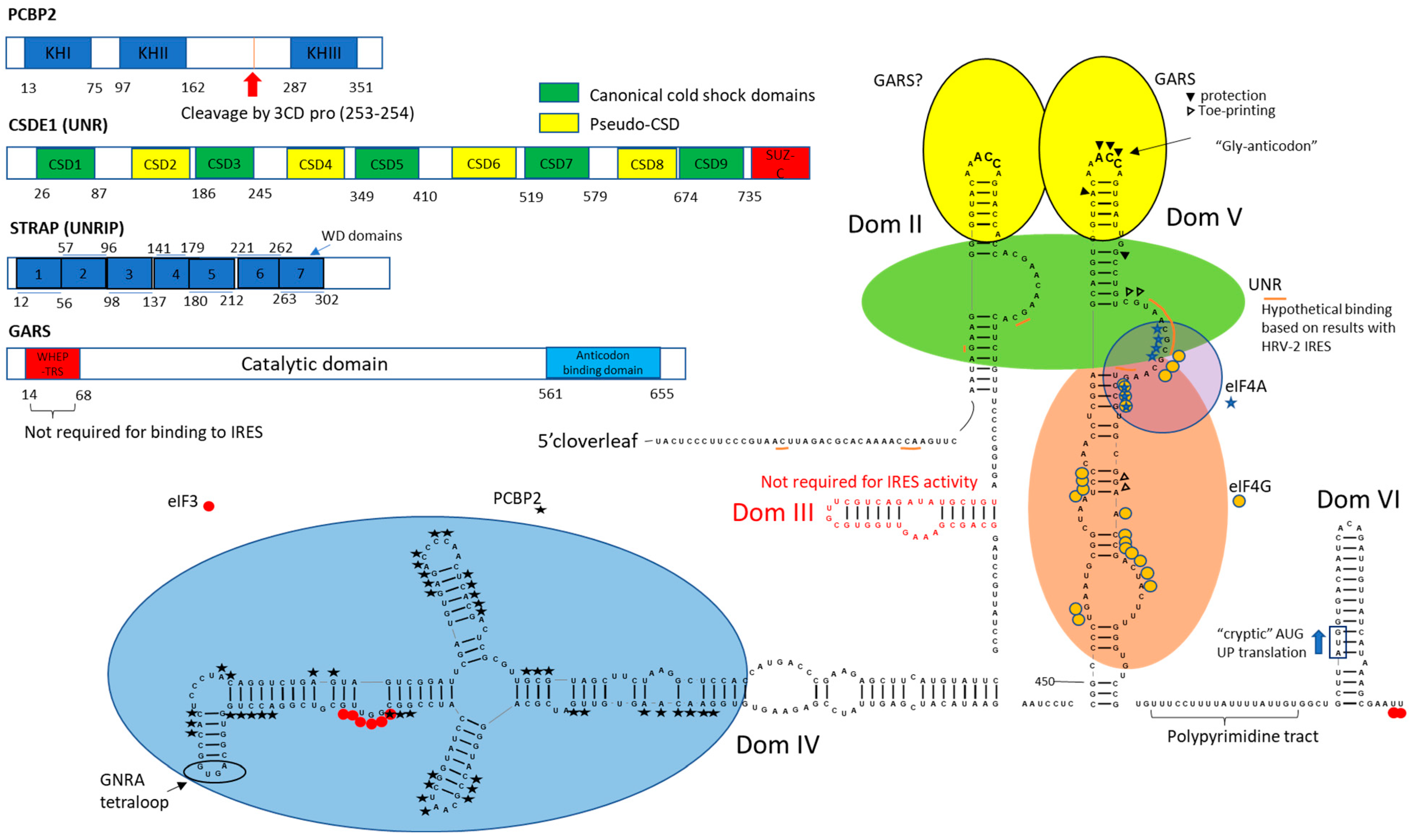
1.3. PCBP2
1.4. UNR and UNRIP
1.5. GARS
2. Perspectives
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Kozak, M. How do eucaryotic ribosomes select initiation regions in messenger RNA? Cell 1978, 15, 1109–1123. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. Evaluation of the “scanning model” for initiation of protein synthesis in eucaryotes. Cell 1980, 22, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Shirokikh, N.E.; Preiss, T. Translation initiation by cap-dependent ribosome recruitment: Recent insights and open questions. Wiley Interdiscip. Rev. RNA 2018, 9, e1473. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G.; Lorsch, J.R. The mechanism of eukaryotic translation initiation: New insights and challenges. Cold Spring Harb. Perspect. Biol. 2012, 4, a011544. [Google Scholar] [CrossRef] [PubMed]
- Meerovitch, K.; Nicholson, R.; Sonenberg, N. In vitro mutational analysis of cis-acting RNA translational elements within the poliovirus type 2 5′ untranslated region. J. Virol. 1991, 65, 5895–5901. [Google Scholar] [CrossRef] [PubMed]
- Trono, D.; Andino, R.; Baltimore, D. An RNA sequence of hundreds of nucleotides at the 5′ end of poliovirus RNA is involved in allowing viral protein synthesis. J. Virol. 1988, 62, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, R.; Pelletier, J.; Le, S.Y.; Sonenberg, N. Structural and functional analysis of the ribosome landing pad of poliovirus type 2: In vivo translation studies. J. Virol. 1991, 65, 5886–5894. [Google Scholar] [CrossRef]
- Dildine, S.L.; Semler, B.L. The deletion of 41 proximal nucleotides reverts a poliovirus mutant containing a temperature-sensitive lesion in the 5′ noncoding region of genomic RNA. J. Virol. 1989, 63, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Lulla, V.; Dinan, A.M.; Hosmillo, M.; Chaudhry, Y.; Sherry, L.; Irigoyen, N.; Nayak, K.M.; Stonehouse, N.J.; Zilbauer, M.; Goodfellow, I.; et al. An upstream protein-coding region in enteroviruses modulates virus infection in gut epithelial cells. Nat. Microbiol. 2019, 4, 280–292. [Google Scholar] [CrossRef]
- Mailliot, J.; Martin, F. Viral internal ribosomal entry sites: Four classes for one goal. Wiley Interdiscip. Rev. RNA 2018, 9, e1458. [Google Scholar] [CrossRef]
- Sorokin, I.I.; Vassilenko, K.S.; Terenin, I.M.; Kalinina, N.O.; Agol, V.I.; Dmitriev, S.E. Non-Canonical Translation Initiation Mechanisms Employed by Eukaryotic Viral mRNAs. Biochem. Biokhimiia 2021, 86, 1060–1094. [Google Scholar] [CrossRef]
- Balvay, L.; Soto Rifo, R.; Ricci, E.P.; Decimo, D.; Ohlmann, T. Structural and functional diversity of viral IRESes. Biochim. Biophys. Acta 2009, 1789, 542–557. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Unbehaun, A.; Spahn, C.M.T. Ribosomal Chamber Music: Toward an Understanding of IRES Mechanisms. Trends Biochem. Sci. 2017, 42, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ulloa, B.; Fuentes, Y.; Pizarro-Ortega, M.S.; Lopez-Lastra, M. RNA-Binding Proteins as Regulators of Internal Initiation of Viral mRNA Translation. Viruses 2022, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Chen, C.J.; Shih, S.R. Regulation Mechanisms of Viral IRES-Driven Translation. Trends Microbiol. 2017, 25, 546–561. [Google Scholar] [CrossRef]
- Brown, B.A.; Ehrenfeld, E. Translation of poliovirus RNA in vitro: Changes in cleavage pattern and initiation sites by ribosomal salt wash. Virology 1979, 97, 396–405. [Google Scholar] [CrossRef]
- Dorner, A.J.; Semler, B.L.; Jackson, R.J.; Hanecak, R.; Duprey, E.; Wimmer, E. In vitro translation of poliovirus RNA: Utilization of internal initiation sites in reticulocyte lysate. J. Virol. 1984, 50, 507–514. [Google Scholar] [CrossRef]
- Phillips, B.A.; Emmert, A. Modulation of the expression of poliovirus proteins in reticulocyte lysates. Virology 1986, 148, 255–267. [Google Scholar] [CrossRef]
- Svitkin, Y.V.; Pestova, T.V.; Maslova, S.V.; Agol, V.I. Point mutations modify the response of poliovirus RNA to a translation initiation factor: A comparison of neurovirulent and attenuated strains. Virology 1988, 166, 394–404. [Google Scholar] [CrossRef]
- Meerovitch, K.; Pelletier, J.; Sonenberg, N. A cellular protein that binds to the 5′-noncoding region of poliovirus RNA: Implications for internal translation initiation. Genes Dev. 1989, 3, 1026–1034. [Google Scholar] [CrossRef]
- Meerovitch, K.; Svitkin, Y.V.; Lee, H.S.; Lejbkowicz, F.; Kenan, D.J.; Chan, E.K.; Agol, V.I.; Keene, J.D.; Sonenberg, N. La autoantigen enhances and corrects aberrant translation of poliovirus RNA in reticulocyte lysate. J. Virol. 1993, 67, 3798–3807. [Google Scholar] [CrossRef] [PubMed]
- Pestova, T.V.; Hellen, C.U.; Shatsky, I.N. Canonical eukaryotic initiation factors determine initiation of translation by internal ribosomal entry. Mol. Cell. Biol. 1996, 16, 6859–6869. [Google Scholar] [CrossRef]
- Sweeney, T.R.; Abaeva, I.S.; Pestova, T.V.; Hellen, C.U. The mechanism of translation initiation on Type 1 picornavirus IRESs. EMBO J. 2014, 33, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Aviner, R.; Li, K.H.; Frydman, J.; Andino, R. Cotranslational prolyl hydroxylation is essential for flavivirus biogenesis. Nature 2021, 596, 558–564. [Google Scholar] [CrossRef]
- Makeyev, A.V.; Liebhaber, S.A. The poly(C)-binding proteins: A multiplicity of functions and a search for mechanisms. RNA 2002, 8, 265–278. [Google Scholar] [CrossRef]
- Yuan, C.; Chen, M.; Cai, X. Advances in poly(rC)-binding protein 2: Structure, molecular function, and roles in cancer. Biomed. Pharmacother. 2021, 139, 111719. [Google Scholar] [CrossRef]
- Shi, H.; Bencze, K.Z.; Stemmler, T.L.; Philpott, C.C. A cytosolic iron chaperone that delivers iron to ferritin. Science 2008, 320, 1207–1210. [Google Scholar] [CrossRef]
- Blyn, L.B.; Swiderek, K.M.; Richards, O.; Stahl, D.C.; Semler, B.L.; Ehrenfeld, E. Poly(rC) binding protein 2 binds to stem-loop IV of the poliovirus RNA 5′ noncoding region: Identification by automated liquid chromatography-tandem mass spectrometry. Proc. Natl. Acad. Sci. USA 1996, 93, 11115–11120. [Google Scholar] [CrossRef]
- Blyn, L.B.; Towner, J.S.; Semler, B.L.; Ehrenfeld, E. Requirement of poly(rC) binding protein 2 for translation of poliovirus RNA. J. Virol. 1997, 71, 6243–6246. [Google Scholar] [CrossRef]
- Andino, R.; Rieckhof, G.E.; Baltimore, D. A functional ribonucleoprotein complex forms around the 5′ end of poliovirus RNA. Cell 1990, 63, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Gamarnik, A.V.; Andino, R. Two functional complexes formed by KH domain containing proteins with the 5′ noncoding region of poliovirus RNA. RNA 1997, 3, 882–892. [Google Scholar]
- Parsley, T.B.; Towner, J.S.; Blyn, L.B.; Ehrenfeld, E.; Semler, B.L. Poly (rC) binding protein 2 forms a ternary complex with the 5′-terminal sequences of poliovirus RNA and the viral 3CD proteinase. RNA 1997, 3, 1124–1134. [Google Scholar] [PubMed]
- Gamarnik, A.V.; Andino, R. Switch from translation to RNA replication in a positive-stranded RNA virus. Genes Dev. 1998, 12, 2293–2304. [Google Scholar] [CrossRef]
- Perera, R.; Daijogo, S.; Walter, B.L.; Nguyen, J.H.; Semler, B.L. Cellular protein modification by poliovirus: The two faces of poly(rC)-binding protein. J. Virol. 2007, 81, 8919–8932. [Google Scholar] [CrossRef] [PubMed]
- Chase, A.J.; Daijogo, S.; Semler, B.L. Inhibition of poliovirus-induced cleavage of cellular protein PCBP2 reduces the levels of viral RNA replication. J. Virol. 2014, 88, 3192–3201. [Google Scholar] [CrossRef]
- Beckham, S.A.; Matak, M.Y.; Belousoff, M.J.; Venugopal, H.; Shah, N.; Vankadari, N.; Elmlund, H.; Nguyen, J.H.C.; Semler, B.L.; Wilce, M.C.J.; et al. Structure of the PCBP2/stem-loop IV complex underlying translation initiation mediated by the poliovirus type I IRES. Nucleic Acids Res. 2020, 48, 8006–8021. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Day, N.; Trifillis, P.; Kiledjian, M. An mRNA stability complex functions with poly(A)-binding protein to stabilize mRNA in vitro. Mol. Cell. Biol. 1999, 19, 4552–4560. [Google Scholar] [CrossRef]
- Bedard, K.M.; Daijogo, S.; Semler, B.L. A nucleo-cytoplasmic SR protein functions in viral IRES-mediated translation initiation. EMBO J. 2007, 26, 459–467. [Google Scholar] [CrossRef]
- Fitzgerald, K.D.; Semler, B.L. Re-localization of cellular protein SRp20 during poliovirus infection: Bridging a viral IRES to the host cell translation apparatus. PLoS Pathog. 2011, 7, e1002127. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.X.; Cui, J.J.; Wang, L.Y.; Yin, J.Y. The role of CSDE1 in translational reprogramming and human diseases. Cell Commun. Signal. CCS 2020, 18, 14. [Google Scholar] [CrossRef]
- Ray, S.; Catnaigh, P.O.; Anderson, E.C. Post-transcriptional regulation of gene expression by Unr. Biochem. Soc. Trans. 2015, 43, 323–327. [Google Scholar] [CrossRef]
- Grosset, C.; Chen, C.Y.; Xu, N.; Sonenberg, N.; Jacquemin-Sablon, H.; Shyu, A.B. A mechanism for translationally coupled mRNA turnover: Interaction between the poly(A) tail and a c-fos RNA coding determinant via a protein complex. Cell 2000, 103, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.A.; Spriggs, K.A.; Coldwell, M.J.; Jackson, R.J.; Willis, A.E. The Apaf-1 internal ribosome entry segment attains the correct structural conformation for function via interactions with PTB and unr. Mol. Cell 2003, 11, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.P.; Ma, S.; Bag, J. The autoregulatory translational control element of poly(A)-binding protein mRNA forms a heteromeric ribonucleoprotein complex. Nucleic Acids Res. 2005, 33, 7074–7089. [Google Scholar] [CrossRef] [PubMed]
- Dinur, M.; Kilav, R.; Sela-Brown, A.; Jacquemin-Sablon, H.; Naveh-Many, T. In vitro evidence that upstream of N-ras participates in the regulation of parathyroid hormone messenger ribonucleic acid stability. Mol. Endocrinol. 2006, 20, 1652–1660. [Google Scholar] [CrossRef]
- Tinton, S.A.; Schepens, B.; Bruynooghe, Y.; Beyaert, R.; Cornelis, S. Regulation of the cell-cycle-dependent internal ribosome entry site of the PITSLRE protein kinase: Roles of Unr (upstream of N-ras) protein and phosphorylated translation initiation factor eIF-2alpha. Biochem. J. 2005, 385, 155–163. [Google Scholar] [CrossRef]
- Abaza, I.; Coll, O.; Patalano, S.; Gebauer, F. Drosophila UNR is required for translational repression of male-specific lethal 2 mRNA during regulation of X-chromosome dosage compensation. Genes Dev. 2006, 20, 380–389. [Google Scholar] [CrossRef]
- Duncan, K.; Grskovic, M.; Strein, C.; Beckmann, K.; Niggeweg, R.; Abaza, I.; Gebauer, F.; Wilm, M.; Hentze, M.W. Sex-lethal imparts a sex-specific function to UNR by recruiting it to the msl-2 mRNA 3′ UTR: Translational repression for dosage compensation. Genes Dev. 2006, 20, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.L.; Hsuan, J.J.; Totty, N.; Jackson, R.J. Unr, a cellular cytoplasmic RNA-binding protein with five cold-shock domains, is required for internal initiation of translation of human rhinovirus RNA. Genes Dev. 1999, 13, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Boussadia, O.; Niepmann, M.; Creancier, L.; Prats, A.C.; Dautry, F.; Jacquemin-Sablon, H. Unr is required in vivo for efficient initiation of translation from the internal ribosome entry sites of both rhinovirus and poliovirus. J. Virol. 2003, 77, 3353–3359. [Google Scholar] [CrossRef]
- Shyu, A.B.; Greenberg, M.E.; Belasco, J.G. The c-fos transcript is targeted for rapid decay by two distinct mRNA degradation pathways. Genes Dev. 1989, 3, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Schiavi, S.C.; Wellington, C.L.; Shyu, A.B.; Chen, C.Y.; Greenberg, M.E.; Belasco, J.G. Multiple elements in the c-fos protein-coding region facilitate mRNA deadenylation and decay by a mechanism coupled to translation. J. Biol. Chem. 1994, 269, 3441–3448. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Yamashita, A.; Chen, C.Y.; Yamashita, Y.; Zhu, W.; Durdan, S.; Kahvejian, A.; Sonenberg, N.; Shyu, A.B. UNR, a new partner of poly(A)-binding protein, plays a key role in translationally coupled mRNA turnover mediated by the c-fos major coding-region determinant. Genes Dev. 2004, 18, 2010–2023. [Google Scholar] [CrossRef]
- Wu, J.; Bag, J. Negative control of the poly(A)-binding protein mRNA translation is mediated by the adenine-rich region of its 5′-untranslated region. J. Biol. Chem. 1998, 273, 34535–34542. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.P.; Bag, J. IMP1 interacts with poly(A)-binding protein (PABP) and the autoregulatory translational control element of PABP-mRNA through the KH III-IV domain. FEBS J. 2006, 273, 5678–5690. [Google Scholar] [CrossRef]
- Bashaw, G.J.; Baker, B.S. The regulation of the Drosophila msl-2 gene reveals a function for Sex-lethal in translational control. Cell 1997, 89, 789–798. [Google Scholar] [CrossRef]
- Kelley, R.L.; Wang, J.; Bell, L.; Kuroda, M.I. Sex lethal controls dosage compensation in Drosophila by a non-splicing mechanism. Nature 1997, 387, 195–199. [Google Scholar] [CrossRef]
- Beckmann, K.; Grskovic, M.; Gebauer, F.; Hentze, M.W. A dual inhibitory mechanism restricts msl-2 mRNA translation for dosage compensation in Drosophila. Cell 2005, 122, 529–540. [Google Scholar] [CrossRef]
- Grskovic, M.; Hentze, M.W.; Gebauer, F. A co-repressor assembly nucleated by Sex-lethal in the 3′UTR mediates translational control of Drosophila msl-2 mRNA. EMBO J. 2003, 22, 5571–5581. [Google Scholar] [CrossRef]
- Duncan, K.E.; Strein, C.; Hentze, M.W. The SXL-UNR corepressor complex uses a PABP-mediated mechanism to inhibit ribosome recruitment to msl-2 mRNA. Mol. Cell 2009, 36, 571–582. [Google Scholar] [CrossRef]
- Ray, S.; Anderson, E.C. Stimulation of translation by human Unr requires cold shock domains 2 and 4, and correlates with poly(A) binding protein interaction. Sci. Rep. 2016, 6, 22461. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.; Howell, M.T.; Patton, J.G.; Jackson, R.J. The involvement of a spliceosome component in internal initiation of human rhinovirus RNA translation. J. Gen. Virol. 1993, 74 Pt 9, 1775–1788. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.L.; Jackson, R.J. Polypyrimidine-tract binding protein (PTB) is necessary, but not sufficient, for efficient internal initiation of translation of human rhinovirus-2 RNA. RNA 1999, 5, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.C.; Hunt, S.L.; Jackson, R.J. Internal initiation of translation from the human rhinovirus-2 internal ribosome entry site requires the binding of Unr to two distinct sites on the 5′ untranslated region. J. Gen. Virol. 2007, 88, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Schimmel, P. Essential nontranslational functions of tRNA synthetases. Nat. Chem. Biol. 2013, 9, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Dreher, T.W. Viral tRNAs and tRNA-like structures. Wiley Interdiscip. Rev. RNA 2010, 1, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Andreev, D.E.; Hirnet, J.; Terenin, I.M.; Dmitriev, S.E.; Niepmann, M.; Shatsky, I.N. Glycyl-tRNA synthetase specifically binds to the poliovirus IRES to activate translation initiation. Nucleic Acids Res. 2012, 40, 5602–5614. [Google Scholar] [CrossRef] [PubMed]
- Nikonova, E.Y.; Mihaylina, A.O.; Lekontseva, N.V.; Nikonov, O.S.; Klyashtorny, V.G.; Kravchenko, O.V.; Andreev, D.E.; Shatsky, I.N.; Garber, M.B. Determination of the Minimal Fragment of the Poliovirus IRES Necessary for the Formation of a Specific Complex with the Human Glycyl-tRNA Synthetase. Biofizika 2016, 61, 277–285. [Google Scholar] [CrossRef]
- Jackson, R.J.; Hunt, S.L.; Gibbs, C.L.; Kaminski, A. Internal initiation of translation of picornavirus RNAs. Mol. Biol. Rep. 1994, 19, 147–159. [Google Scholar] [CrossRef]
- Lee, S.W.; Cho, B.H.; Park, S.G.; Kim, S. Aminoacyl-tRNA synthetase complexes: Beyond translation. J. Cell Sci. 2004, 117, 3725–3734. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Baleanu-Gogonea, C.; Willard, B.; Gogonea, V.; Fox, P.L. 3-Dimensional architecture of the human multi-tRNA synthetase complex. Nucleic Acids Res. 2020, 48, 8740–8754. [Google Scholar] [CrossRef] [PubMed]
- Antonellis, A.; Lee-Lin, S.Q.; Wasterlain, A.; Leo, P.; Quezado, M.; Goldfarb, L.G.; Myung, K.; Burgess, S.; Fischbeck, K.H.; Green, E.D. Functional analyses of glycyl-tRNA synthetase mutations suggest a key role for tRNA-charging enzymes in peripheral axons. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 10397–10406. [Google Scholar] [CrossRef] [PubMed]
- Park, M.C.; Kang, T.; Jin, D.; Han, J.M.; Kim, S.B.; Park, Y.J.; Cho, K.; Park, Y.W.; Guo, M.; He, W.; et al. Secreted human glycyl-tRNA synthetase implicated in defense against ERK-activated tumorigenesis. Proc. Natl. Acad. Sci. USA 2012, 109, E640–E647. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Kim, H.J.; Yang, S.R.; Park, C.H.; Lee, H.Y.; Hong, I.S. A novel endogenous damage signal, glycyl tRNA synthetase, activates multiple beneficial functions of mesenchymal stem cells. Cell Death Differ. 2018, 25, 2023–2036. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.T.; Chong, Y.E.; Guo, M.; Yang, X.L. Crystal structures and biochemical analyses suggest a unique mechanism and role for human glycyl-tRNA synthetase in Ap4A homeostasis. J. Biol. Chem. 2009, 284, 28968–28976. [Google Scholar] [CrossRef]
- Qin, X.; Deng, X.; Chen, L.; Xie, W. Crystal Structure of the Wild-Type Human GlyRS Bound with tRNA(Gly) in a Productive Conformation. J. Mol. Biol. 2016, 428, 3603–3614. [Google Scholar] [CrossRef]
- Song, Y.; Tzima, E.; Ochs, K.; Bassili, G.; Trusheim, H.; Linder, M.; Preissner, K.T.; Niepmann, M. Evidence for an RNA chaperone function of polypyrimidine tract-binding protein in picornavirus translation. RNA 2005, 11, 1809–1824. [Google Scholar] [CrossRef]
- Ochs, K.; Zeller, A.; Saleh, L.; Bassili, G.; Song, Y.; Sonntag, A.; Niepmann, M. Impaired binding of standard initiation factors mediates poliovirus translation attenuation. J. Virol. 2003, 77, 115–122. [Google Scholar] [CrossRef]
- de Breyne, S.; Yu, Y.; Unbehaun, A.; Pestova, T.V.; Hellen, C.U. Direct functional interaction of initiation factor eIF4G with type 1 internal ribosomal entry sites. Proc. Natl. Acad. Sci. USA 2009, 106, 9197–9202. [Google Scholar] [CrossRef]
- Bergamini, G.; Preiss, T.; Hentze, M.W. Picornavirus IRESes and the poly(A) tail jointly promote cap-independent translation in a mammalian cell-free system. RNA 2000, 6, 1781–1790. [Google Scholar] [CrossRef]
- Svitkin, Y.V.; Imataka, H.; Khaleghpour, K.; Kahvejian, A.; Liebig, H.D.; Sonenberg, N. Poly(A)-binding protein interaction with elF4G stimulates picornavirus IRES-dependent translation. RNA 2001, 7, 1743–1752. [Google Scholar] [PubMed]
- Etchison, D.; Milburn, S.C.; Edery, I.; Sonenberg, N.; Hershey, J.W. Inhibition of HeLa cell protein synthesis following poliovirus infection correlates with the proteolysis of a 220,000-dalton polypeptide associated with eucaryotic initiation factor 3 and a cap binding protein complex. J. Biol. Chem. 1982, 257, 14806–14810. [Google Scholar] [CrossRef] [PubMed]
- Kuyumcu-Martinez, N.M.; Van Eden, M.E.; Younan, P.; Lloyd, R.E. Cleavage of poly(A)-binding protein by poliovirus 3C protease inhibits host cell translation: A novel mechanism for host translation shutoff. Mol. Cell. Biol. 2004, 24, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.V.; Wimmer, E. Initiation of protein-primed picornavirus RNA synthesis. Virus Res. 2015, 206, 12–26. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreev, D.E.; Niepmann, M.; Shatsky, I.N. Elusive Trans-Acting Factors Which Operate with Type I (Poliovirus-like) IRES Elements. Int. J. Mol. Sci. 2022, 23, 15497. https://doi.org/10.3390/ijms232415497
Andreev DE, Niepmann M, Shatsky IN. Elusive Trans-Acting Factors Which Operate with Type I (Poliovirus-like) IRES Elements. International Journal of Molecular Sciences. 2022; 23(24):15497. https://doi.org/10.3390/ijms232415497
Chicago/Turabian StyleAndreev, Dmitry E., Michael Niepmann, and Ivan N. Shatsky. 2022. "Elusive Trans-Acting Factors Which Operate with Type I (Poliovirus-like) IRES Elements" International Journal of Molecular Sciences 23, no. 24: 15497. https://doi.org/10.3390/ijms232415497
APA StyleAndreev, D. E., Niepmann, M., & Shatsky, I. N. (2022). Elusive Trans-Acting Factors Which Operate with Type I (Poliovirus-like) IRES Elements. International Journal of Molecular Sciences, 23(24), 15497. https://doi.org/10.3390/ijms232415497