
Influence of Single Deuterium Replacement on Frequency of Hydrogen Bond Dissociation in IFNA17 under the Highest Critical Energy Range

 ,
,
Abstract
1. Introduction
2. Results
3. Discussion
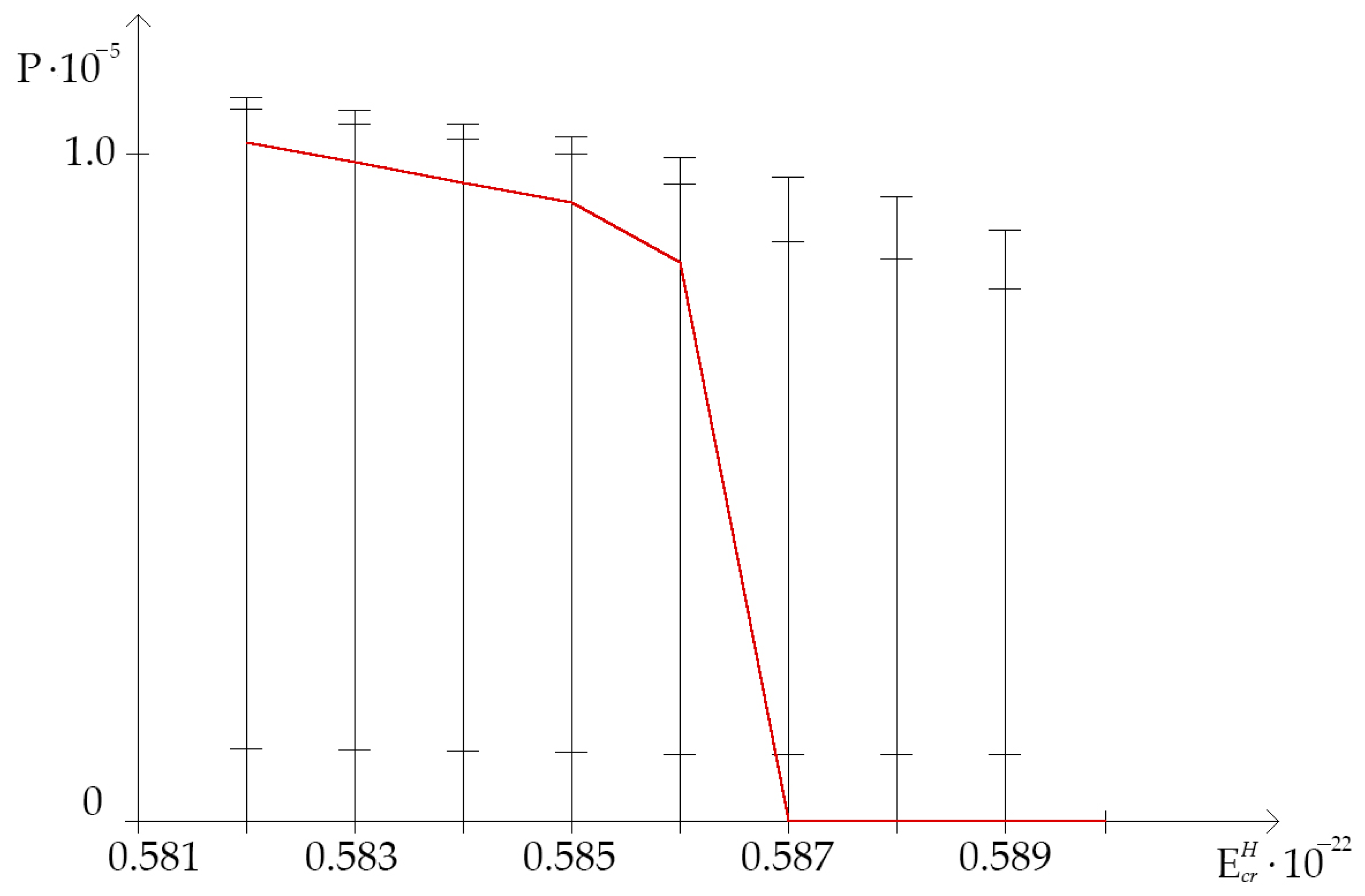
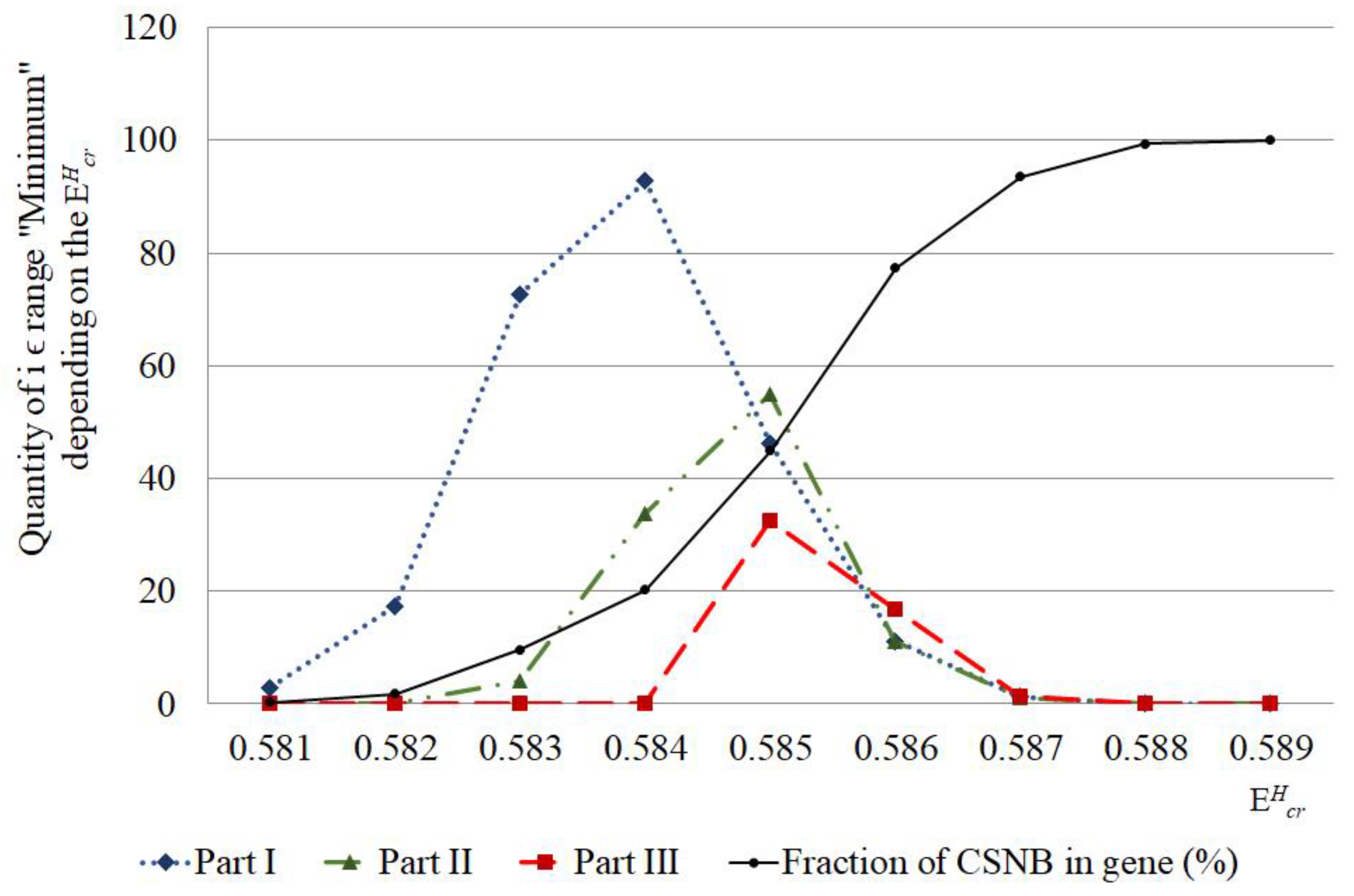
- In some cases, the single 2H/1H replacement result in positive value of OS occurrence frequency in the IFNA17 throughout diapason from N·m to N·m opposing to the OS occurrence frequency, when all hydrogen bonds in DNA nucleotides are 1H, which is always equaled to 0.0 after exceed N·m. This underlines that at least 22.7% of the total number 2H-substituted nucleobases can reduce molecular interaction in the studied gene and increase the hydrogen bond dissociation, foremost in its part from 328 to 653 nitrogenous bases;
- The counted occurrences of the OSs were much higher when the single 2H/1H replacement had taken place at nucleobases of the middle part IFNA17 (from 328 to 653 nucleotides) compared to its other parts, and this had a strong prevalence rate in the G-C pairs;
- The lowest rate of the OS occurrence was under the single deuterium substitution at the nitrogenous bases in the gene III part (from 654 to 980 nucleotides), which was also too rich in A-T pairs (72.2%) compared to the other parts of IFNA17, so that the calculated was equal to 0.0 for all of the studied critical energies (from N·m to N·m);
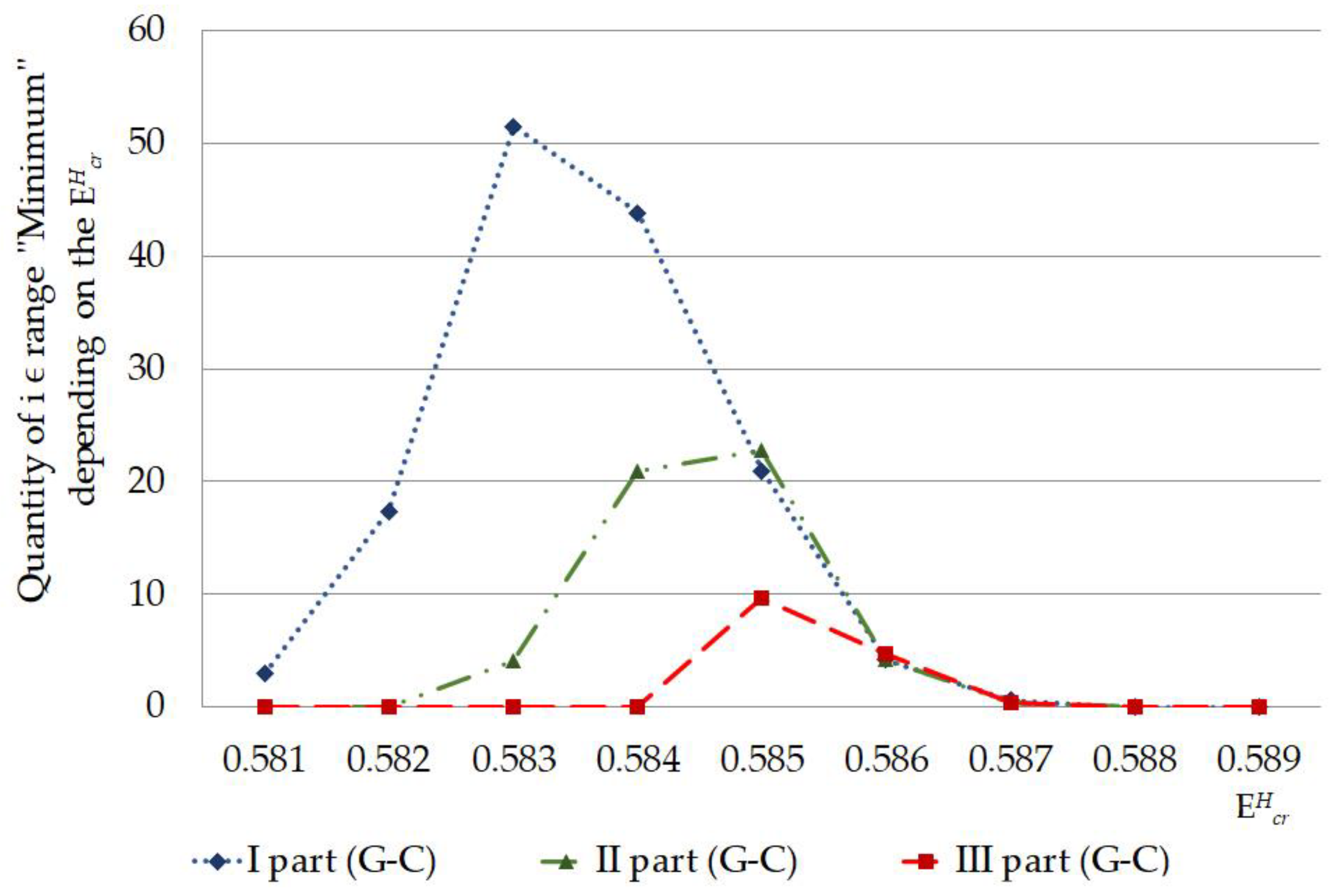
- Sum of was less significant when the single 2H/1H replacement occurred at the A-T nucleobase pairs compared to the G-C ones in the I and II parts of IFNA17, and the relationship between total and at G-C in these parts for each throughout the energy diapason from N·m to N·m was strong () that proves the obvious dominant role of 2H-substitutied G-C bases in the generation compared to the A-T;
- Earliest CSNBs (n = 3) arose under equal to N·m when IFNA17 had had the single 2H-substituted cytosine or guanine nitrogenous bases in its I part (from 1 to 327 nucleotides). Moreover, throughout the range energy from N·m to N·m, the single 2H/1H replacement, leading to the CSNBs, prevailed in the I part, especially for its G-C pairs making up at least 67% of the total CSNB quantity in the whole gene. So, for IFNA17 throughout the range of from N·m to N·m the Spearman correlation coefficient between A-T/G-C ratio in the each gene part and CSNB numbers was significant and negative ();
- The highest acceleration of CSNB occurrence was observed when the single 2H/1H replacement took place at nucleotides of the III part of IFNA17 under from N·m to N·m, and throughout it they very abruptly reached the value of 100% of the nucleobases in this gene part. It indicates the obvious and higher vulnerability of IFNA17 due to the single 2H-substitution at nucleobases from 654 to 980 compared with other gene parts exposed to studied critical energies, which increased the risk of permanent disorders of converting genetic information to mRNA messenger;
- All of the above-mentioned underline clearly the significant difference in the responsiveness of each IFNA17 parts under range of critical energies because of the single 2H/1H replacement in their nucleobases and with its strong dependence on A-T/G-C ratio with the prevalent contribution of the last pair, which leads to an increase due to the 2H-substitution into its nitrogenous bases both and CSNB, especially under diapason from N·m to N·m;
- The single 2H-substitution at G-C pairs not only had the most influence on the initial arise both and in the whole gene throughout the critical energy diapason from N·m to N·m but also made possible the existence of the last OS occurrence under equals to N·at least in six cases that proves the leading effect of the isotopic 2H/1H modifications at G-C compared to A-T on the molecular dynamics of IFNA17;
- In addition, in the study was presented a modified algorithm allowing for accounting for nucleobases with the single 2H/1H replacement, which leads to occurrence of both the highest rate of Oss and CSNBs. Also, it showed the developed approach, decreasing significantly the false positive results compared to non-modified BJ-algorithm [29] due to the differentiated counting of the total sum of CSNB occurrence in the gene with relevance to the critical energy in the highest diapason.
4. Materials and Methods
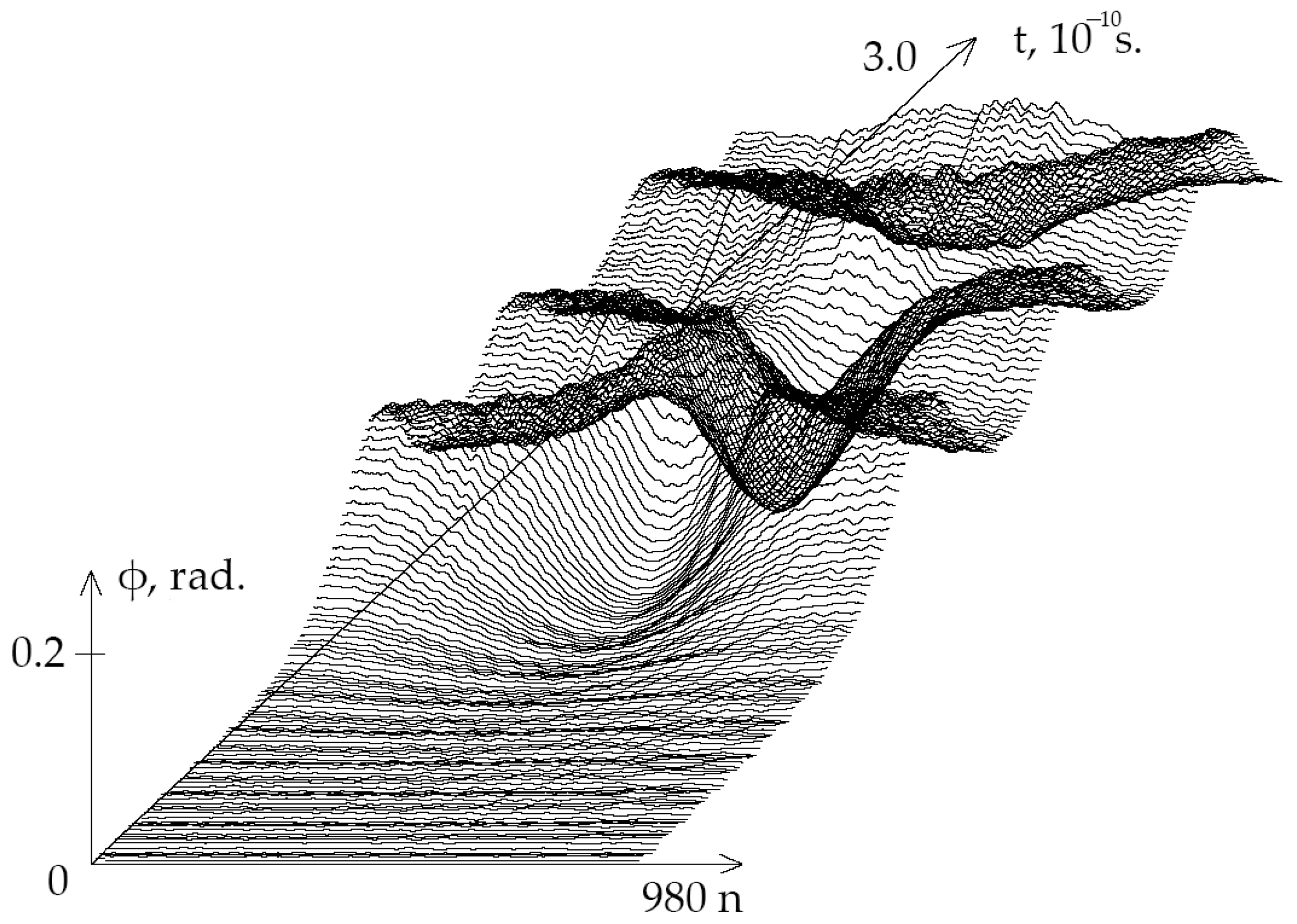
4.1. Mathematical Model
4.2. Modification of Basov–Jimack Algorithm
- (1)
- i ϵ range “Maximum” (BJ-max):
- if and :; or else:
- if and :;
- (2)
- i ϵ range “Minimum” (CSNB):
- if ;
4.3. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, F.; Inman, J.T.; Hong, Y.; Hall, P.M.; Wang, M.D. Resonator nanophotonic standing-wave array trap for single-molecule manipulation and measurement. Nat. Commun. 2022, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Kayikcioglu, T.; Ngo, T.T.M.; Ranjan, A.; Eustermann, S.; Cieza, B.; Morgan, M.T.; Hejna, M.; Rube, H.T.; Hopfner, K.P.; et al. Measuring DNA mechanics on the genome scale. Nature 2021, 589, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Jie, M.; Wang, M.D. DNA supercoiling during transcription. Biophys. Rev. 2016, 8, 75–87. [Google Scholar] [CrossRef]
- Konyshev, I.; Byvalov, A. Model systems for optical trapping: The physical basis and biological applications. Biophys. Rev. 2021, 13, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Manghi, M.; Destainville, N. Physics of base-pairing dynamics in DNA. Phys. Rep. 2016, 631, 1–41. [Google Scholar] [CrossRef]
- Chevizovich, D.; Michieletto, D.; Mvogo, A.; Zakiryanov, F.; Zdravković, S. A review on nonlinear DNA physics. R. Soc. Open Sci. 2020, 7, 200774. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Drobotenko, M.I.; Basov, A.A.; Svidlov, A.A.; Fedulova, L.V.; Lyasota, O.M.; Baryshev, M.G. Mathematical Modeling of Open State in DNA Molecule Depending on the Deuterium Concentration in the Surrounding Liquid Media at Different Values of Hydrogen Bond Disruption Energy. Dokl. Biochem. Biophys. 2018, 483, 359–362. [Google Scholar] [CrossRef]
- De Santis, D.; Guarcello, C.; Spagnolo, B.; Carollo, A.; Valenti, D. Generation of travelling sine-Gordon breathers in noisy long Josephson junctions. Chaos Solitons Fractals 2022, 158, 112039. [Google Scholar] [CrossRef]
- Grinevich, A.A.; Masulis, I.S.; Yakushevich, L.V. Mathematical Modeling of Transcription Bubble Behavior in the pPF1 Plasmid and its Modified Versions: The Link between the Plasmid Energy Profile and the Direction of Transcription. Biophysics 2021, 66, 209–217. [Google Scholar] [CrossRef]
- Svidlov, A.A.; Drobotenko, M.I.; Basov, A.A.; Gerasimenko, E.O.; Malyshko, V.V.; Elkina, A.A.; Baryshev, M.G.; Dzhimak, S.S. DNA dynamics under periodic force effects. Int. J. Mol. Sci. 2021, 22, 7873. [Google Scholar] [CrossRef]
- Pyne, A.L.B.; Noy, A.; Main, K.H.S.; Velasco-Berrelleza, V.; Piperakis, M.M.; Mitchenall, L.A.; Cugliandolo, F.M.; Beton, J.G.; Stevenson, C.E.M.; Hoogenboom, B.W.; et al. Base-pair resolution analysis of the effect of supercoiling on DNA flexibility and major groove recognition by triplex-forming oligonucleotides. Nat. Commun. 2021, 12, 1053. [Google Scholar] [CrossRef] [PubMed]
- Drobotenko, M.I.; Dzhimak, S.S.; Svidlov, A.A.; Basov, A.A.; Lyasota, O.M.; Baryshev, M.G. A Mathematical Model for Basepair Opening in a DNA Double Helix. Biophysics 2018, 63, 177–182. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Svidlov, A.A.; Basov, A.A.; Baryshev, M.G.; Drobotenko, M.I. The effect of single deuterium substitutions for protium in a DNA molecule on the occurrence of open states. Biophysics 2018, 63, 497–500. [Google Scholar] [CrossRef]
- Svidlov, A.A.; Drobotenko, M.I.; Basov, A.A.; Elkina, A.A.; Gerasimenko, E.O.; Malyshko, V.V.; Baryshev, M.G.; Dzhimak, S.S. Influence of the 2H/1H isotope composition of the water environment on the probability of denaturation bubble formation in a DNA molecule. Phys. Wave Phenom. 2021, 29, 180–185. [Google Scholar] [CrossRef]
- Syroeshkin, A.V.; Antipova, N.V.; Zlatska, A.V.; Zlatskiy, I.A.; Skylska, M.D.; Grebennikova, T.V.; Goncharuk, V.V. The effect of the deuterium depleted water on the biological activity of the eukaryotic cells. J. Trace Elem. Med. Biol. 2018, 50, 629–633. [Google Scholar] [CrossRef]
- Basov, A.A.; Bykov, I.M.; Baryshev, M.G.; Dzhimak, S.S.; Bykov, M.I. Determination of deuterium concentration in foods and influence of water with modified isotopic composition on oxidation parameters and heavy hydrogen isotopes content in experimental animals. Vopr. Pitan. 2014, 83, 43–50. [Google Scholar]
- Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Yaglov, V.V. Response of Pituitary—Thyroid Axis to a Short-Term Shift in Deuterium Content in the Body. Bull. Exp. Biol. Med. 2021, 171, 262–264. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Basov, A.A.; Fedulova, L.V.; Naumov, G.N.; Baryshev, M.G. Correction of metabolic processes in rats during chronic endotoxicosis using isotope (D/H) exchange reactions. Biol. Bull. 2015, 42, 440–448. [Google Scholar] [CrossRef]
- Elkina, A.A.; Tumaev, E.N.; Basov, A.A.; Moiseev, A.V.; Malyshko, V.V.; Barisheva, E.V.; Churkina, A.V.; Dzhimak, S.S. The mechanisms of the interaction of stable isotopes with biological objects in the presence of an uncompensated neutron in chemical bonds. Biophysics 2020, 65, 883–888. [Google Scholar] [CrossRef]
- Schmidt, H.L.; Robins, R.J.; Werner, R.A. Multi-factorial in vivo stable isotope fractionation: Causes, correlations, consequences and applications. Isot. Environ. Health Stud. 2015, 51, 155–199. [Google Scholar] [CrossRef]
- Basov, A.A.; Kozin, S.V.; Bikov, I.M.; Popov, K.A.; Moiseev, A.V.; Elkina, A.A.; Dzhimak, S.S. Changes in prooxidant-antioxidant system indices in the blood and brain of rats with modelled acute hypoxia which consumed a deuterium-depleted drinking diet. Biol. Bull. 2019, 46, 531–535. [Google Scholar] [CrossRef]
- Xie, X.; Zubarev, R.A. Isotopic Resonance Hypothesis: Experimental Verification by Escherichia coli Growth Measurements. Sci. Rep. 2015, 5, 9215. [Google Scholar] [CrossRef] [PubMed]
- Kravtsov, A.; Kozin, S.; Basov, A.; Butina, E.; Baryshev, M.; Malyshko, V.; Moiseev, A.; Elkina, A.; Dzhimak, S. Reduction of deuterium level supports resistance of neurons to glucose deprivation and hypoxia: Study in cultures of neurons and on animals. Molecules 2022, 27, 243. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Liu, Y.; Wang, L.; Lu, F. Interaction between heavy water and single-strand DNA: A SERS study. Molecules 2022, 27, 6023. [Google Scholar] [CrossRef] [PubMed]
- Lisicin, A.B.; Barishev, M.G.; Basov, A.A.; Barisheva, E.V.; Bikov, I.M.; Didikin, A.S.; Tekutskaya, E.E.; Timakov, A.A.; Fedulova, L.V.; Chernuha, I.M.; et al. Influence of deuterium depleted water on the organism of laboratory animals in various functional conditions of nonspecific protective systems. Biophysics 2014, 59, 620–627. [Google Scholar] [CrossRef]
- Yakushevich, L.V. Nonlinear Physics of DNA; John Wiley & Sons: Hoboken, NJ, USA, 2007; p. 220. [Google Scholar] [CrossRef]
- Svidlov, A.A.; Drobotenko, M.I.; Basov, A.A.; Gerasimenko, E.O.; Elkina, A.A.; Baryshev, M.G.; Nechipurenko, Y.D.; Dzhimak, S.S. Influence of Environmental Parameters on the Stability of the DNA Molecule. Entropy 2021, 23, 1446. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Svidlov, A.A.; Elkina, A.A.; Gerasimenko, E.O.; Baryshev, M.G.; Drobotenko, M.I. Genesis of Open States Zones in a DNA Molecule Depends on the Localization and Value of the Torque. Int. J. Mol. Sci. 2022, 23, 4428. [Google Scholar] [CrossRef]
- Basov, A.A.; Drobotenko, M.I.; Svidlov, A.A.; Gerasimenko, E.O.; Malyshko, V.V.; Elkina, A.A.; Baryshev, M.G.; Dzhimak, S.S. Inequality in the Frequency of the Open States Occurrence Depends on Single 2H/1H Replacement in DNA. Molecules 2020, 25, 3753. [Google Scholar] [CrossRef]
- GeneCards: The Human Gene Database. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=IFNA17 (accessed on 19 October 2022).
- Yakushevich, L.V.; Krasnobaeva, L.A. Forced Oscillations of DNA Bases. Biophysics 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Werner, R.M.; Stivers, J.T. Kinetic isotope effect studies of the reaction catalyzed by uracil DNA glycosylase: Evidence for an oxocarbenium ion-uracil anion intermediate. Biochemistry 2000, 39, 14054–14064. [Google Scholar] [CrossRef]
- Opitz, C.; Ahrné, E.; Goldie, K.N.; Schmidt, A.; Grzesiek, S. Deuterium induces a distinctive Escherichia coli proteome that correlates with the reduction in growth rate. J. Biol. Chem. 2019, 294, 2279–2292. [Google Scholar] [CrossRef] [PubMed]
- Galagedera, S.; Flechsig, G.U. Deuterium isotope effects upon the redox-switching of the viscosity of DNA layers observed by electrochemical quartz crystal micro-balance. Electroanalysis 2019, 31, 2074–2080. [Google Scholar] [CrossRef]
- Boros, L.G.; Somlyai, I.; Kovács, B.Z.; Puskás, L.G.; Nagy, L.I.; Dux, L.; Farkas, G.; Somlyai, G. Deuterium depletion inhibits cell proliferation, RNA and nuclear membrane turnover to enhance survival in pancreatic cancer. Cancer Control 2021, 28, 1073274821999655. [Google Scholar] [CrossRef] [PubMed]
- Basov, A.A.; Fedulova, L.V.; Baryshev, M.G.; Dzhimak, S.S. Deuterium-depleted water influence on the isotope 2H/1H regulation in body and individual adaptation. Nutrients 2019, 11, 1903. [Google Scholar] [CrossRef] [PubMed]
- Kalkur, R.S.; Ballast, A.C.; Triplett, A.R.; Spendier, K. Effects of deuterium oxide on cell growth and vesicle speed in RBL-2H3 cells. PeerJ 2014, 2, e553. [Google Scholar] [CrossRef]
- Englander, S.W.; Kallenbach, N.R. Hydrogen exchange and structural dynamics of proteins and nucleic acids. Q. Rev. Biophys. 1983, 16, 521–655. [Google Scholar] [CrossRef] [PubMed]
- Sviridova, D.A.; Smirnova, S.V.; Abilev, S.K. Deuterium Oxide Enhances Expression of the ada, alkA, and luxA Genes of Escherichia coli Induced by Methyl Methanesulfonate. Russ. J. Genet. 2022, 58, 481–484. [Google Scholar] [CrossRef]
- Ullah, S.; Ishimoto, T.; Williamson, M.P.; Hansen, P.E. Ab initio calculations of deuterium isotope effects on chemical shifts of salt-bridged lysines. J. Phys. Chem. B 2011, 115, 3208–3215. [Google Scholar] [CrossRef]
- Ignatov, I.; Mosin, O.; Velikov, B.; Bauer, E. Influence of isotopic composition of water with varrying deuterium content in composition with mountain water of bulgaria on human longevity. J. Med. Physiol. Biophys. 2014, 7, 46–78. [Google Scholar]
- Hohlefelder, L.S.; Stögbauer, T.; Opitz, M.; Bayerl, T.M.; Rädler, J.O. Heavy water reduces GFP expression in prokaryotic cell-free assays at the translation level while stimulating its transcription. BioMed Res. Int. 2013, 2013, 592745. [Google Scholar] [CrossRef]
- Ignatov, I.; Mosin, O.; Bauer, E. Mathematical model of melt water and mountain water from Bulgaria obtained by IR, NES and DNES-methods. J. Med. Physiol. Biophys. 2015, 17, 30–52. [Google Scholar]
- Basov, A.A.; Fedulova, L.V.; Vasilevskaya, E.R.; Dzhimak, S.S. Possible mechanisms of biological effects observed in living systems during 2H/1H isotope fractionation and deuterium interactions with other biogenic isotopes. Molecules 2019, 24, 4101. [Google Scholar] [CrossRef] [PubMed]
- Brini, E.; Fennell, C.J.; Fernandez-Serra, M.; Hribar-Lee, B.; Lukšič, M.; Dill, K.A. How water’s properties are encoded in its molecular structure and energies. Chem Rev. 2017, 117, 12385–12414. [Google Scholar] [CrossRef] [PubMed]
- Kleemann, J.; Reichenbach, G.; Zöller, N.; Jäger, M.; Kaufmann, R.; Meissner, M.; Kippenberger, S. Heavy water affects vital parameters of human melanoma cells in vitro. Cancer Manag. Res. 2020, 12, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Kim, N.H.; Jin, H.S.; Seo, Y.J.; Lee, J.; Lee, J.H. Base-pair opening dynamics of nucleic acids in relation to their biological function. Comput. Struct. Biotechnol. J. 2019, 17, 797–804. [Google Scholar] [CrossRef]
- Kannan, S.; Zacharias, M. Role of the closing base pair for d(GCA) hairpin stability: Free energy analysis and folding simulations. Nucleic Acids Res. 2011, 39, 8271–8280. [Google Scholar] [CrossRef]
- Bikard, D.; Loot, C.; Baharoglu, Z.; Mazel, D. Folded DNA in action: Hairpin formation and biological functions in prokaryotes. Microbiol. Mol. Biol. Rev. 2010, 74, 570–588. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, M.; Cranford, S. Ranking of molecular biomarker interaction with targeted DNA nucleobases via full atomistic molecular dynamics. Sci. Rep. 2016, 6, 18659. [Google Scholar] [CrossRef]
- Yamasaki, S.; Terada, T.; Shimizu, K.; Kono, H.; Sarai, A. A generalized conformational energy function of DNA derived from molecular dynamics simulations. Nucleic Acids Res. 2009, 37, e135. [Google Scholar] [CrossRef][Green Version]
- Brovarets, O.O.; Hovorun, D.M. Proton tunneling in the A·T Watson-Crick DNA base pair: Myth or reality? J. Biomol. Struct. Dyn. 2015, 33, 2716–2720. [Google Scholar] [CrossRef]
- Farthing, D.E.; Buxbaum, N.P.; Lucas, P.J.; Maglakelidze, N.; Oliver, B.; Wang, J.; Hu, K.; Castro, E.; Bare, C.V.; Gress, R.E. Comparing DNA enrichment of proliferating cells following administration of different stable isotopes of heavy water. Sci. Rep. 2017, 7, 4043. [Google Scholar] [CrossRef] [PubMed]
- Manalo, M.N.; Pérez, L.M.; LiWang, A. Hydrogen-bonding and pi-pi base-stacking interactions are coupled in DNA, as suggested by calculated and experimental trans-Hbond deuterium isotope shifts. J. Am. Chem. Soc. 2007, 129, 11298–11299. [Google Scholar] [CrossRef]
- Sicard, F.; Destainville, N.; Manghi, M. DNA denaturation bubbles: Free-energy landscape and nucleation/closure rates. J. Chem. Phys. 2015, 142, 034903. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, V.; Villa, A.; Hess, B. Sequence dependency of canonical base pair opening in the DNA double helix. PLoS Comput. Biol. 2017, 13, e1005463. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.G.; Bartolotti, L.; Li, L. Deuterium and its role in the machinery of evolution. J. Theor. Biol. 2006, 238, 914–918. [Google Scholar] [CrossRef] [PubMed]
- Priyakumar, U.D.; MacKerell, A.D., Jr. Computational approaches for investigating base flipping in oligonucleotides. Chem. Rev. 2006, 106, 489–505. [Google Scholar] [CrossRef]
- Grosjean, H. DNA and RNA Modification Enzymes: Structure, Mechanism, Function and Evolution, 1st ed.; CRC Press: Boca Raton, FL, USA, 2018; p. 682. [Google Scholar] [CrossRef]
- Dubois, A.; Francois, C.; Descamps, V.; Fournier, C.; Wychowski, C.; Dubuisson, J.; Castelain, S.; Duverlie, G. Enhanced anti-HCV activity of interferon alpha 17 subtype. Virology 2009, 6, 70. [Google Scholar] [CrossRef]
- GenBank: Homo Sapiens Interferon Alpha 17 (IFNA17), mRNA. Available online: http://www.ncbi.nlm.nih.gov/nuccore/NM_021268.2 (accessed on 20 October 2022).
- Ordu, O.; Lusser, A.; Dekker, N.H. Recent insights from in vitro single-molecule studies into nucleosome structure and dynamics. Biophys. Rev. 2016, 8, 33–49. [Google Scholar] [CrossRef]
- Noy, A.; Sutthibutpong, T.; Harris, S.A. Protein/DNA interactions in complex DNA topologies: Expect the unexpected. Biophys. Rev. 2016, 8, 233–243. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part of Gene | A–T (CSNB, n) | G–C (CSNB, n) | ||
|---|---|---|---|---|
| 0.581 | 1.02 | I | 0 | 3 |
| II | 0 | 0 | ||
| III | 0 | 0 | ||
| 0.582 | I | 0 | 18 | |
| 0.99 | II | 0 | 0 | |
| III | 0 | 0 | ||
| 0.583 | 0.96 | I | 26 | 63 |
| II | 0 | 5 | ||
| III | 0 | 0 | ||
| 0.584 | I | 77 | 69 | |
| 0.93 | II | 20 | 33 | |
| III | 0 | 0 | ||
| 0.585 | 0.84 | I | 77 | 69 |
| II | 106 | 75 | ||
| III | 75 | 32 | ||
| 0.586 | 0 | I | 134 | 82 |
| II | 127 | 82 | ||
| III | 236 | 91 | ||
| 0.587 | 0 | I | 163 | 149 |
| II | 178 | 93 | ||
| III | 236 | 91 | ||
| 0.588 | 0 | I | 163 | 164 |
| II | 186 | 134 | ||
| III | 236 | 91 | ||
| 0.589 | 0 | I | 163 | 164 |
| II | 186 | 140 | ||
| III | 236 | 91 |
| Part of IFNA17 | Nucleobase Pair Quantity | A–T Ratio (%) | G–C Ratio (%) |
|---|---|---|---|
| I | 327 (from 1 to 327) | 49.8 | 50.2 |
| II | 326 (from 328 to 653) | 57.1 | 42.9 |
| III | 327 (from 654 to 980) | 72.2 | 27.8 |
| Part of Gene | |||||
|---|---|---|---|---|---|
| 0.581 | 1.02 | 1.09 | I | 0 | 0 |
| II | 1 | 14 | |||
| III | 0 | 0 | |||
| 0.582 | 0.99 | 1.07 | I | 0 | 10 |
| II | 4 | 41 | |||
| III | 0 | 0 | |||
| 0.583 | 0.96 | 1.05 | I | 0 | 2 |
| II | 4 | 39 | |||
| III | 0 | 0 | |||
| 0.584 | 0.93 | 1.03 | I | 0 | 1 |
| II | 1 | 27 | |||
| III | 0 | 0 | |||
| 0.585 | 0.84 | 1 | I | 0 | 21 |
| II | 17 | 49 | |||
| III | 0 | 0 | |||
| 0.586 | 0 | 0.97 | I | 12 | 53 |
| II | 56 | 53 | |||
| III | 0 | 0 | |||
| 0.587 | 0 | 0.94 | I | 0 | 15 |
| II | 8 | 47 | |||
| III | 0 | 0 | |||
| 0.588 | 0 | 0.89 | I | 0 | 0 |
| II | 0 | 6 | |||
| III | 0 | 0 | |||
| 0.589 | 0 | 0 | I | 0 | 0 |
| II | 0 | 0 | |||
| III | 0 | 0 |
| S | F | ||
|---|---|---|---|
| A | a | b | |
| B | c | d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basov, A.; Drobotenko, M.; Svidlov, A.; Bezhenar, M.; Gerasimenko, E.; Moiseev, A.; Malyshko, V.; Dorohova, A.; Drozdov, A.; Baryshev, M.; et al. Influence of Single Deuterium Replacement on Frequency of Hydrogen Bond Dissociation in IFNA17 under the Highest Critical Energy Range. Int. J. Mol. Sci. 2022, 23, 15487. https://doi.org/10.3390/ijms232415487
Basov A, Drobotenko M, Svidlov A, Bezhenar M, Gerasimenko E, Moiseev A, Malyshko V, Dorohova A, Drozdov A, Baryshev M, et al. Influence of Single Deuterium Replacement on Frequency of Hydrogen Bond Dissociation in IFNA17 under the Highest Critical Energy Range. International Journal of Molecular Sciences. 2022; 23(24):15487. https://doi.org/10.3390/ijms232415487
Chicago/Turabian StyleBasov, Alexander, Mikhail Drobotenko, Alexander Svidlov, Maria Bezhenar, Eugeny Gerasimenko, Arkadii Moiseev, Vadim Malyshko, Anna Dorohova, Andrey Drozdov, Mikhail Baryshev, and et al. 2022. "Influence of Single Deuterium Replacement on Frequency of Hydrogen Bond Dissociation in IFNA17 under the Highest Critical Energy Range" International Journal of Molecular Sciences 23, no. 24: 15487. https://doi.org/10.3390/ijms232415487
APA StyleBasov, A., Drobotenko, M., Svidlov, A., Bezhenar, M., Gerasimenko, E., Moiseev, A., Malyshko, V., Dorohova, A., Drozdov, A., Baryshev, M., & Dzhimak, S. (2022). Influence of Single Deuterium Replacement on Frequency of Hydrogen Bond Dissociation in IFNA17 under the Highest Critical Energy Range. International Journal of Molecular Sciences, 23(24), 15487. https://doi.org/10.3390/ijms232415487