1. Introduction
Breast cancer is the most commonly diagnosed cancer worldwide and ranks first in terms of both prevalence and disease-related mortality in women [
1]. At the 2011 St. Gallen International Breast Cancer Conference, breast cancer was classified according to molecular subtype: luminal A [estrogen receptor (ER)+ and/or progesterone receptor (PR)+, Ki67 low, and human epidermal growth factor receptor 2 (HER2)-]; luminal B (ER+ and/or PR+, Ki-67 high and/or HER2+); HER2-positive (ER-, PR- and HER2+); triple-negative types (ER-, PR-, and HER2-) [
2]. Triple-negative breast cancer (TNBC) accounts for approximately 12–17% of all breast cancers and has the lowest survival rate compared to other subtypes [
3]. The poor treatment outcome in patients with TNBC is due to the lack of expression of the ER, PR, and HER2 during disease progression; thus, existing anticancer drugs targeting these receptors cannot be used.
Radiotherapy (RT), one of the main treatment methods for TNBC, causes DNA double-strand breaks, in turn causing cancer cell death. However, mechanisms that result in acquiring radioresistance, including dose limitations due to radiation tolerance of surrounding normal tissues, accelerated tumor cell repopulation, hypoxia, and enhanced repair of radiation damage can reduce the effectiveness of this therapeutic option [
4]. TNBC is known to be a representative radiation-resistant tumor because of its self-renewal and regeneration characteristics, similar to cancer stem cells, based on the expression of cell surface markers CD44
+CD24
− and aldehyde dehydrogenase 1 (ALDH1) [
5,
6]. Because of this characteristic, TNBC easily acquires resistance to existing standard treatment methods, including RT [
6]. Owing to this, new treatment strategies, such as the development of novel drugs or the application of various combinations of existing therapeutic strategies, are required for the management of TNBC.
In a previous study, we selected benzimidazole derivatives as potential anticancer candidates because their anthelmintic mechanisms are associated with oncogenic pathways. Consequently, these agents displayed anticancer effects in several cell lines and animal studies without causing serious side effects [
7,
8]. Further, we found that mebendazole (MBZ), a benzimidazole derivative, exhibited the most effective anticancer effects in TNBC through inducing DNA damage, cell cycle arrest, and downregulating cancer stem cell markers CD44 and OCT3/4 and cancer progression-related ESM-1 protein expression [
9]. Moreover, other studies that researched on cancer stem cells or cells resistant to approved treatment options reported that MBZ monotherapy has anticancer effects as a repurposed drug, and it also stimulated anti-tumor immune responses in synergy with ionizing radiation and various chemotherapeutic agents [
10,
11,
12]. Herein, we investigated the anticancer effect and possible mechanisms of MBZ and RT coadministration in TNBC cells in vivo and in vitro.
3. Discussion
TNBC is highly invasive, has high metastatic potential, and is prone to relapse, resulting in a poor prognosis with a mortality rate of 40% within 5 years [
14,
15]. Furthermore, TNBC does not respond to hormone- or trastuzumab-based therapies because of the lack of target receptors such as ER, PR, and HER2 in the tumor-related environments. Owing to this, a combination of surgery, chemotherapy, and RT appears to be the only available modality [
16]. Because novel treatment options are cardinal to managing the disease, we investigated the anticancer effect of combined MBZ and RT treatment.
MBZ, a benzimidazole derivative, induces various responses in vivo, and has anthelmintic effects [
17]. In the field of oncology, research is being conducted to supplement existing treatments for various cancers by harnessing the anticancer effect of anthelmintics [
8]. In a previous study, we found that MBZ alone had anticancer effects in RT-resistant TNBC by inducing apoptosis and cell cycle arrest, suppressing the expression of cancer stem cell markers (CD44 and OCT3/4), and inhibiting cancer progression-related ESM-1 proteins [
9]. Herein, we found that the coadministration of MBZ and RT increased the anticancer effect in RT-resistant TNBC compared to MBZ or RT individual regimens.
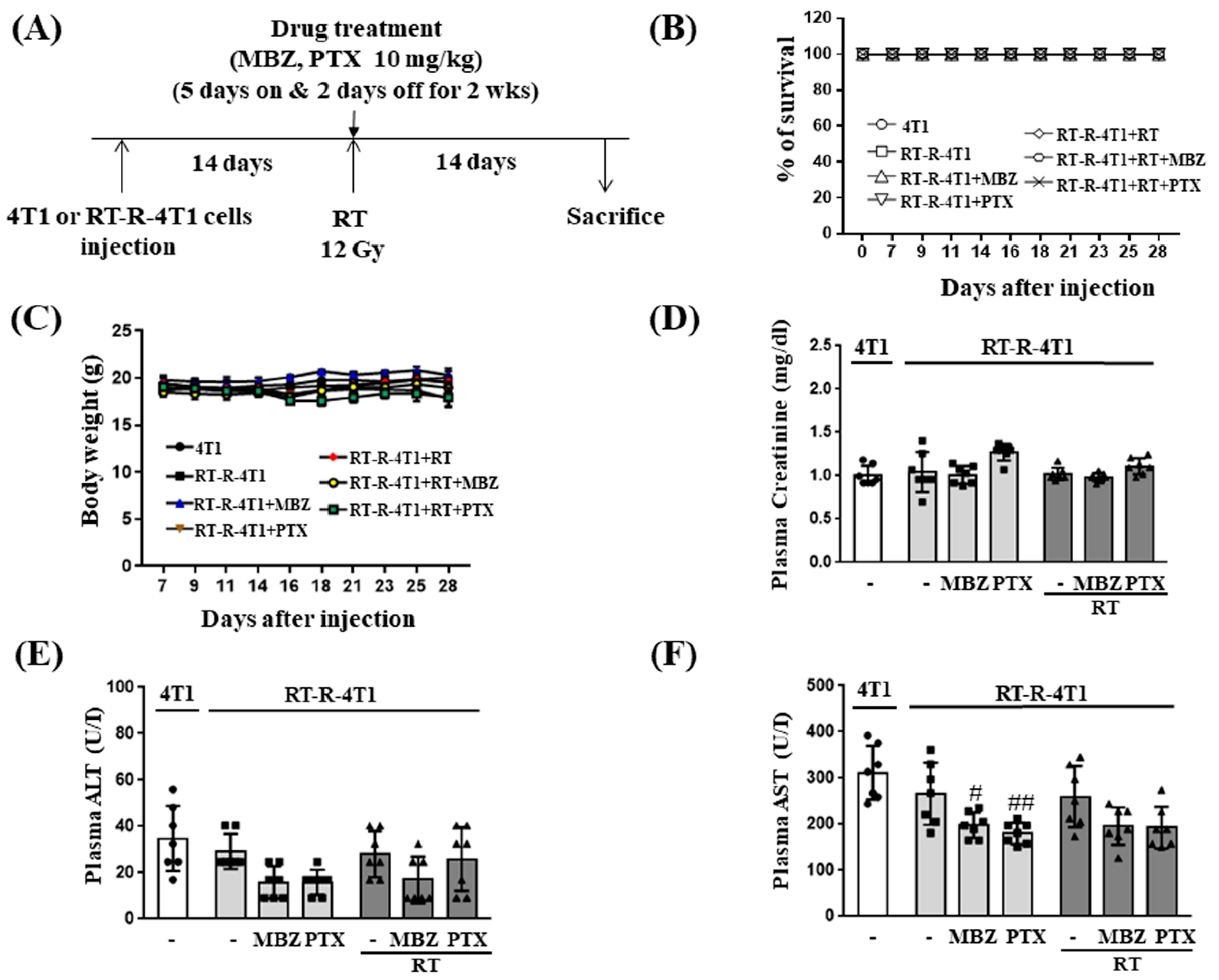
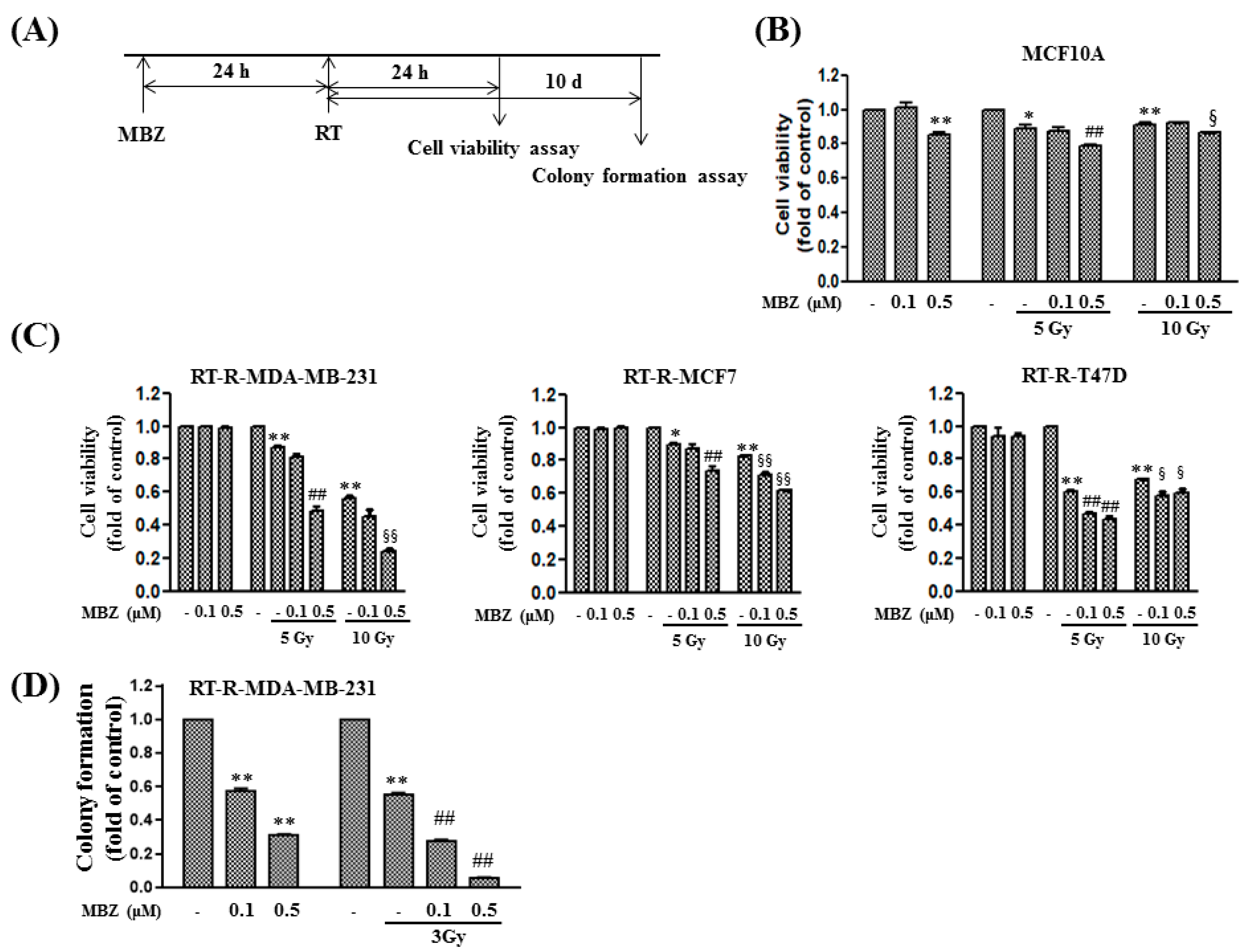
We first determined the toxicity of MBZ (10 mg/kg) + RT (12 Gy) by measuring changes in body weight, plasma creatinine, plasma ALT, and plasma AST in mice injected with RT-R-4T1 breast cancer cells and by examining the viability of MCF10A cells, a normal breast epithelial cell line. The results from these experiments demonstrated that co-treatment with MBZ (10 mg/kg) and RT (12 Gy) had little or no toxicity. Based on our findings and the proven safety profile of MBZ, the MBZ+RT combination regimen can be considered a safe treatment option that has no toxicity to normal tissues.
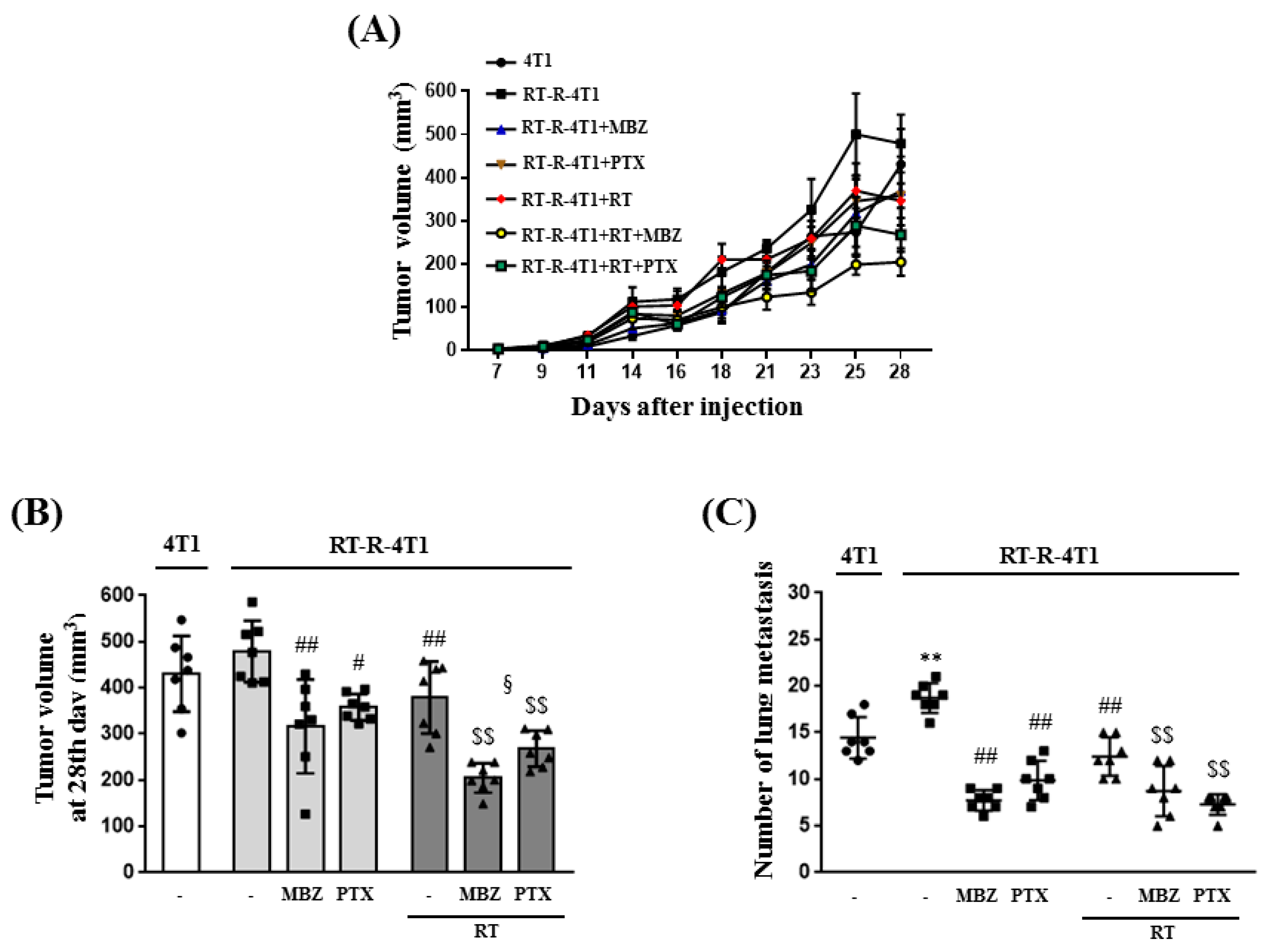
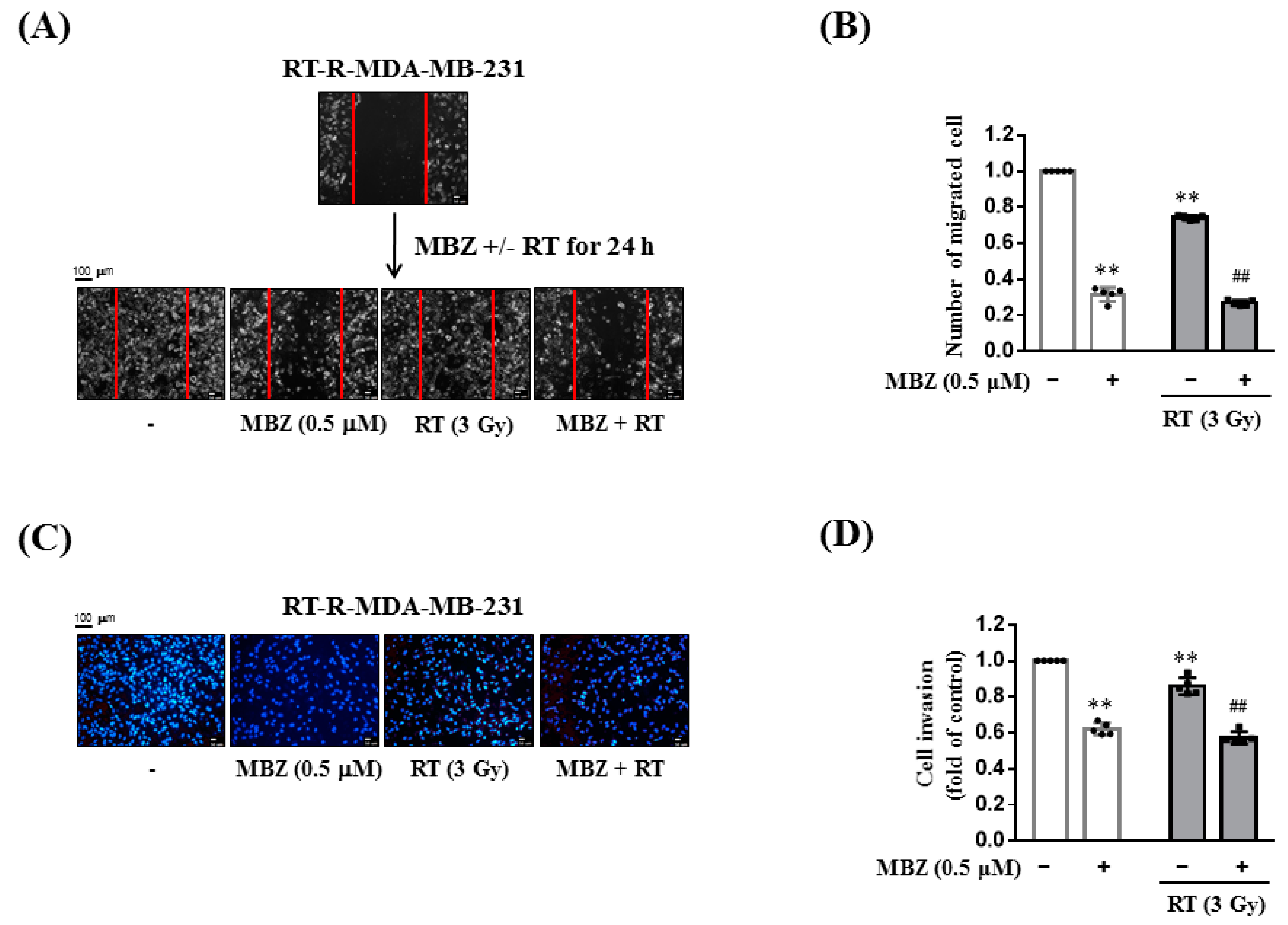
Following combination treatment with MBZ and RT at the determined safe doses, the anticancer effect of the regimen was determined in animal models and using cell line experiments. The main measurement point was whether the anticancer effect of the combined treatment was greater than that of RT. From the results of tumor volume, number of lung metastases, tumor cell viability, colony formation, tumor cell migration, and invasion, the enhanced anticancer effect of the combined regimen was demonstrated in RT-R-TNBC cells.
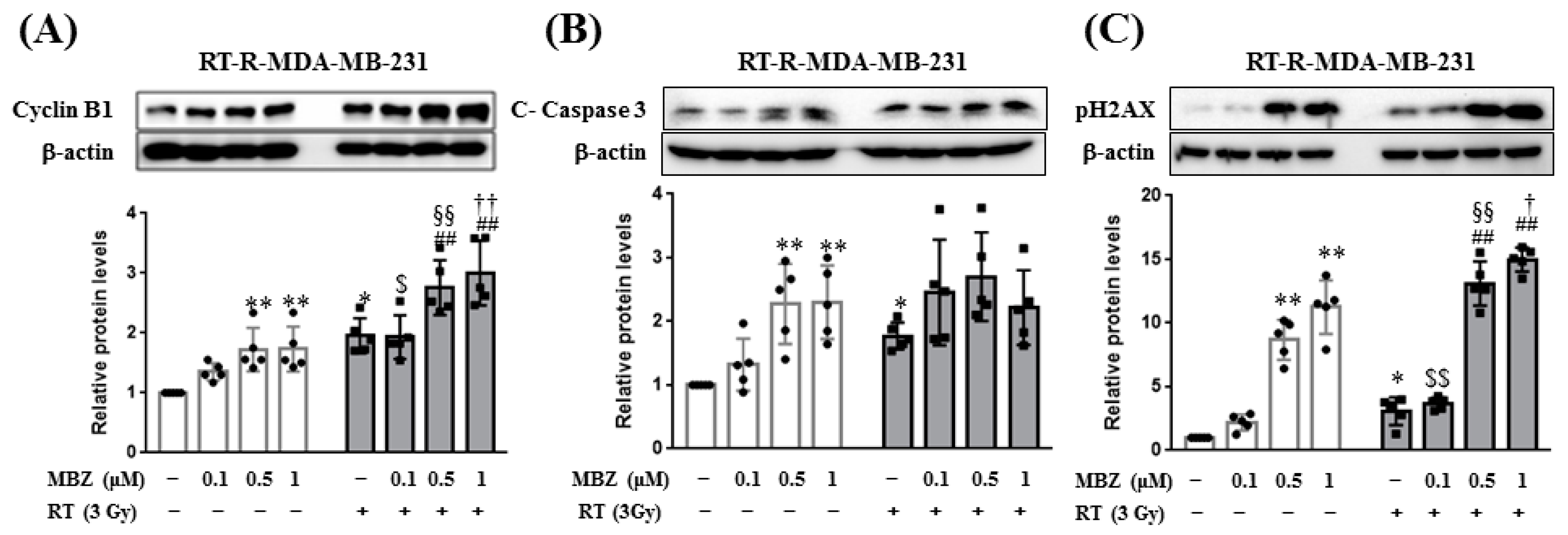
In this study, the enhanced anticancer effect of the combination regimen was associated with increased cyclin B1 expression. In a previous study, MBZ induced cell cycle arrest in the G2/M phase by increasing cyclin B1 expression and inhibiting tubulin polymerization [
9]. From this result, it can be inferred that MBZ + RT can promote cell cycle arrest in the G2/M phase through an increase in cyclin B1. In addition, in this study, PH2AX was significantly increased in the MBZ + RT combined treatment group, confirming the increased effect on DNA double-strand breaks.
Le et al. conducted a study titled “Mebendazole potentiates radiation therapy in triple-negative breast cancer” [
18]. They reported that breast cancer-initiating cells (BCIC) are hierarchically constituting cells of breast cancer and are resistant to chemotherapy or RT, and TNBC is relatively rich in BCIC, making it readily resistant to conventional treatment. They also reported that non-tumorigenic breast cancer cells that survive exposure to ionizing radiation can be converted to BCIC by the activation of a dedifferentiation program. They performed high-throughput screening to identify compounds that inhibit RT-induced dedifferentiation in TNBC and confirmed MBZ as a potential agent. In their study, they argued that MBZ could induce cell cycle arrest, DNA double-strand breaks, and apoptosis, as well as act as a radiation sensitizer by reducing BCIC. This conclusion supports the enhanced anticancer effect of MBZ, as observed herein.
The normal immune system generally contributes to the recognition and removal of foreign pathogens including tumors [
19]. In particular, NK cells, a subset of innate lymphoid cells, are the most efficient effector cells among cytolytic lymphocytes, showing natural cytotoxicity against primary tumor cells and suppressing metastasis through inhibition of cancer cell proliferation, migration, and colonization to distant organs [
20]. However, interestingly, the immune cells in the tumor microenvironment (TME) exhibit a functional change and contribute to growth and metastasis rather than the removal of tumor cells. The NK cell function is also inhibited in the TME and provides conditions that are favorable for tumor progression. According to our previous study [
13], RT-R-TNBC cells which exhibit more aggressive properties in terms of tumor growth, invasion, and metastasis suppressed NK cell cytotoxicity by regulating ligands for NK cell activation. RT-R-MDA-MB-231 decreased the expression levels of MICA/B, the ligands for the activating NKG2D receptor, but increased the expression of HLA-E, the ligand for the inhibitory NKG2A receptor. Moreover, NK-92MI cells cocultured with RT-R-MDA-MB-231 cells decreased the levels of secreted perforin and granzyme B more than NK-92MI cells cultured with MDA-MB-231 cells. Therefore, in this study, we further analyzed whether the combination of MBZ and RT affects the cytotoxic function of NK cells, with one of the white blood cells responsible for the innate immune system. Although NK cells mainly act to dissolve cells infected with viruses or bacteria, they can directly inhibit the development, proliferation, and metastasis of cancer cells and effectively eliminate cancer stem cells, which are important for recurrence [
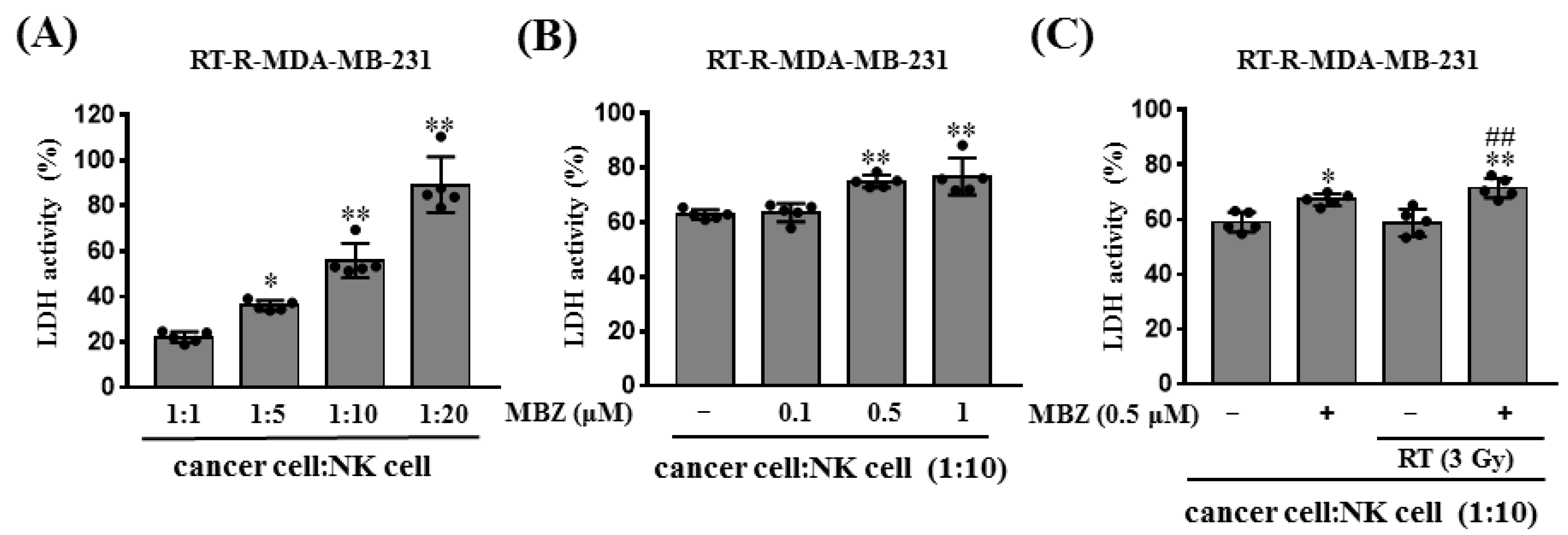
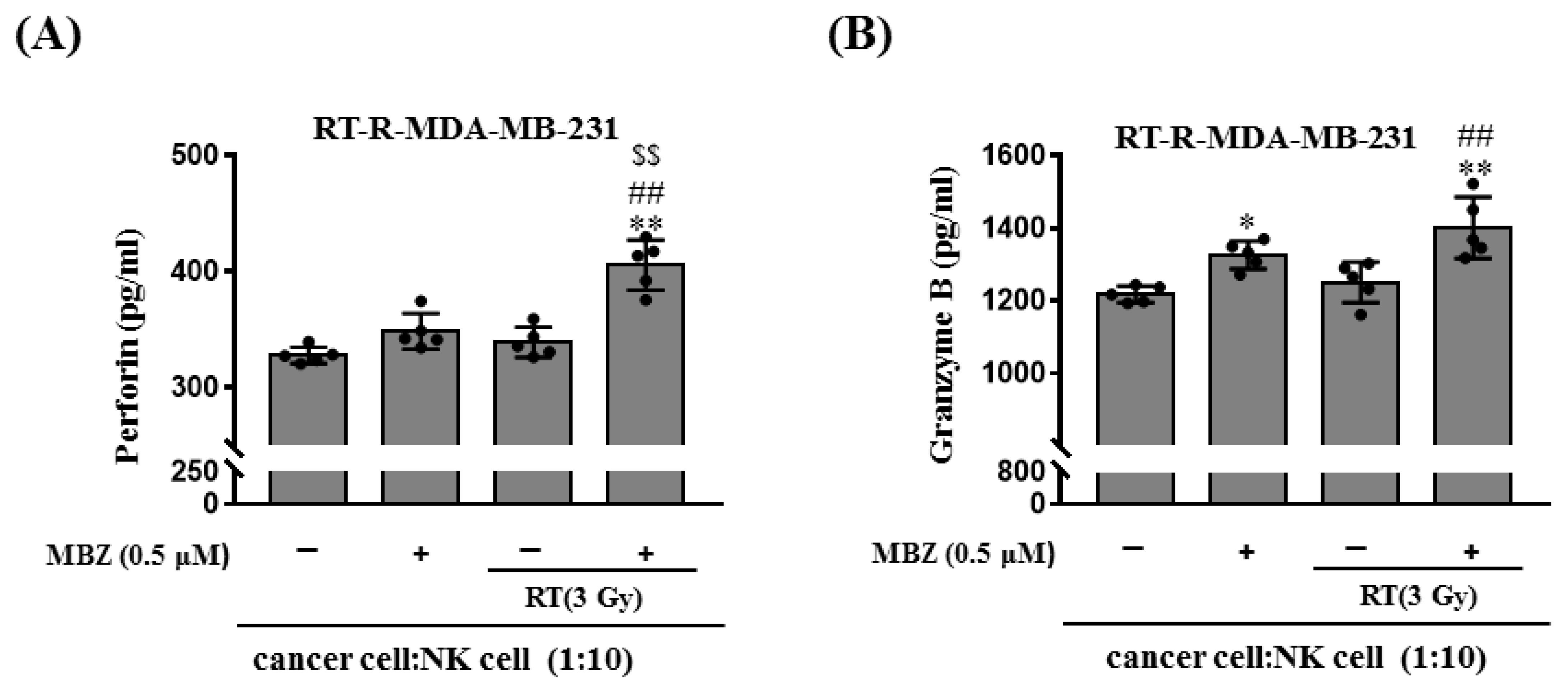
21]. In this study, MBZ + RT significantly increased NK cell-mediated cytotoxicity. In addition, perforin and granzyme B, which are proteins secreted from activated NK cells that play a major role in destroying target cells, were significantly increased in the MBZ + RT group. In contrast, no further increase in cleaved caspase-3 expression was observed. Thus, it could be speculated that NK cells are partly involved in the enhanced anticancer effect of MBZ + RT, and among associated actions, the granule-mediated mechanism of NK cells may be involved. In addition, we postulate the involvement of a caspase-independent pathway involving the mitochondria for cell destruction. Although studies have shown that MBZ or RT individually activate NK cells as an immunomodulator [
22,
23], this study is the first to show an additional enhanced effect from coadministration.
4. Materials and Methods
4.1. Materials
MBZ (
Figure 8) and paclitaxel (PTX) were purchased from Sigma-Aldrich (St. Louis, MO, USA). MBZ was resuspended in 0.25% sodium carboxymethyl cellulose (CMC) solution. RPMI 1640 medium, fetal bovine serum (FBS), and antibiotics (penicillin/streptomycin) were purchased from Cytiva (Marlborough, MA, USA). Anti-cyclin B1 antibody was purchased from Abcam (Cambridge, UK). Anti-pH2AX and cleaved caspase-3 antibody were purchased from Cell Signaling Technology (Beverly, MA, USA). The BD Matrigel basement membrane matrix was supplied by BD Biosciences (San Diego, CA, USA). The Cyto Tox 96
® Non-Radioactive Cytotoxicity Assay Kit was obtained from Promega (Madison, WI, USA). The human perforin and granzyme B ELISA Kits were obtained from MyBioSource (San Diego, CA, USA). The enhanced chemiluminescence (ECL) western blotting detection reagent was obtained from Bio-Rad (Hercules, CA, USA). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
4.2. Animal Experiments
Female athymic nude mice (6-weeks-old) were purchased from Orient Bio (Gyeonggi-do, Republic of Korea). Mice were housed under the following constant ambient conditions: 22–26 °C, 40–60% relative humidity, 12 h light/dark cycles, and free access to sterilized food and water. The experimental protocol was approved by the Institutional Animal Care and Use Committee of the Gyeongsang National University (approval number: GNU-200603-N0030). The mice were injected into the right thigh with 4T1 or RT-resistant (RT-R)-4T1 cells (5 × 104 cells/100 μL). Seven days after the injection, body weight and tumor volume were measured three times per week. Fourteen days after injection, the mice were divided into seven groups (n = 7 per group): (1) mice injected with 4T1 cells (4T1 group), (2) mice injected with RT-R-4T1 cells (RT-R-4T1 group), (3) RT-R-4T1 + MBZ group, (4) RT-R-4T1 + PTX group, (5) RT-R-4T1 + RT group, (6) RT-R-4T1 + RT + MBZ group, and (7) RT-R-4T1 + RT + PTX group. For RT, the mice were anesthetized by intramuscular injection of Zoletil and irradiated with a single 12 Gy dose of X-rays to the right thigh tumor using a 6 MV photon beam from a linear accelerator (21EX, Varian, Palo Alto, CA, USA). On the day of irradiation, MBZ (10 mg/kg) or PTX (10 mg/kg) was administered by oral gavage and intraperitoneal injection, respectively, followed by daily treatment for two consecutive weeks on a “5 days on and 2 days off” schedule. The mice were killed on the 28th day after the initial injection, tumor volumes and lung metastases were measured, and blood was collected. Blood plasma was separated by centrifuging at 3000× g for 15 min. Plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were measured using assay kits from the IVD Lab (Uiwang, Republic of Korea) and a spectrophotometer (Shimadzu UV-1800 spectrophotometer, Tokyo, Japan). Plasma creatinine levels were measured directly using the colorimetric Jaffe method.
4.3. Cell Cultures
Human breast cancer cell lines, MDA-MB-231, MCF7, and T47D, were obtained from the Korea Cell Line Bank (Seoul, Republic of Korea). The non-malignant breast epithelial cell line, MCF10A, mouse breast cancer cell line, 4T1, human umbilical endothelial cell line, EA.hy926, and human NK cell line, NK-92MI, were obtained from American Type Culture Collection (ATCC; Manassas, VA, USA). RT-R-MDA-MB-231, RT-R-MCF7, RT-R-T47D, and RT-R-4T1 cells were generated by repetitively applying a 2 Gy dose of X-rays until a final dose of 50 Gy was achieved. All cancer cell lines were cultured in RPMI 1640 supplemented with 10% FBS, 100 IU/mL penicillin, and 10 μg/mL streptomycin, and incubated at 37 °C in a humidified atmosphere containing 5% CO2. NK-92MI cells were cultured in alpha-MEM containing 2 mM L-glutamine, 1.5 g/L sodium bicarbonate (GIBCO; Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 0.2 mM inositol (Sigma-Aldrich, St. Louis, WA, USA), 0.1 mM 2-mercaptoethanol (Sigma-Aldrich, St. Louis, WA, USA), 0.02 mM folic acid (Sigma-Aldrich, St. Louis, WA, USA), 12.5% horse serum (GIBCO), and 12.5% FBS without ribonucleosides and deoxyribonucleosides at 37 °C in a humidified atmosphere containing 5% CO2.
4.4. Cell Viability Assay
Cells were seeded at a density of 1 × 104 cells/well in 24-well plates, treated with MBZ (0.1 or 0.5 μM) for 24 h, and then irradiated with different doses of X-rays (5 Gy, 10 Gy). After 24 h, cells were treated with 10 μL/well CCK-8 reagent (Dongin Biotech, Seoul, Republic of Korea) and incubated for 30 min at 37 °C in the dark. The optical density of each well was measured at 450 nm using a microplate reader (Molecular Devices VersaMax, Sunnyvale, CA, USA).
4.5. Colony Formation Assay
RT-R-MDA-MB-231 cells were cultured in 6-well plates. The cells were treated with MBZ (0.1 or 0.5 μM) for 24 h at 37 °C and then irradiated with a 3 Gy dose of X-rays. After 24 h, the cells were trypsinized, centrifuged at 1000 rpm for 2 min, and resuspended in RPMI 1640 medium. The cells were seeded in 6-well plates (1 × 103 cells/well) and the culture medium was replaced with a completely new medium every 2–3 days. After 10 days, the medium was discarded, and the cells were washed with phosphate-buffered saline (PBS). The colonies were fixed in 100% methanol for 10 min at 25 °C, stained with 0.1% Giemsa staining solution for 30 min at the same temperature, and visible colonies were counted.
4.6. Scratch Migration Assay
RT-R-MDA-MB-231 cells were cultured in 6-well plates and scratched with a sterile 200 μL pipette tip. The cells were treated with MBZ (0.5 μM) for 24 h at 37 °C and then irradiated with a 3 Gy dose of X-rays. After 24 h, the cells were washed with PBS and images were taken using an Olympus photomicroscope at 0 and 48 h after scratching. The number of cells that migrated to the scratched area was determined.
4.7. Matrigel Invasion Assay
RT-R-MDA-MB-231 cells were cultured in 6-well plates and the cells were treated with MBZ (0.5 μM) at 37 °C for 24 h, followed by irradiation with a 3 Gy dose of X-rays. After 24 h, the cells were trypsinized, centrifuged at 1000 rpm for 2 min, and resuspended in RPMI 1640 medium. The upper chambers of the inserts were coated with 100 μL of Matrigel (1 mg/mL), and endothelial cells (Ecs) (2 × 105 cells) were added to the Matrigel-coated inserts. Resuspended RT-R-MDA-MB-231 cells (2 × 105 cells/insert) were added to the upper chambers in serum-free media, and the lower chambers were filled with 500 µL of RPMI-1640 containing 10% FBS. The cells were grown at 37 °C for 16 h in a culture incubator. To remove non-invasive cells, the upper surface of the insert membranes was scrubbed with a wet swab. Subsequently, the cells on the lower insert membranes were stained with DAPI (Sigma-Aldrich), and the invading cells were counted under a fluorescence microscope (Eclipse Ti-U, Nikon, Tokyo, Japan).
4.8. Western Blot Analysis
RT-R-MDA-MB-231 cells were treated with MBZ (0.1, 0.5, and 1 μM) for 1 h and then irradiated with a 3 Gy dose of X-rays. After 24 h, the cells were harvested and lysed in RIPA buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5% sodium deoxycholate, and protease inhibitors. The samples were centrifuged at 13,000 rpm for 20 min at 4 °C and the supernatants were collected. Protein concentration was determined using the Bradford method. Equal amounts of protein were subjected to between 8–10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto Hybond-P + polyvinylidene difluoride membranes (Amersham, Buckinghamshire, UK). The membranes were blocked with 5% non-fat milk in Tris-buffered saline containing 0.05% Tween-20 (TBS-T) for 1 h at 25 °C and then incubated with the following primary antibodies: anti-cyclin B1 (ab32053; 1:1000), anti-cleaved caspase-3 (9661S; 1:1000), and anti-p-H2AX (2577S; 1:1000). The bound antibodies were detected using horseradish peroxidase-conjugated secondary antibodies and an ECL (Bio-Rad, Hercules, CA, USA) western blotting detection reagent. The relative protein levels were normalized to that of β-actin (MA5-15739, 1:5000, Thermo Fisher Scientific, Waltham, MA, USA), which was used as a loading control.
4.9. Cytotoxicity Assay
NK cell cytotoxicity against cancer cells was assessed using the Cyto Tox 96
® Non-Radioactive Cytotoxicity Assay Kit (Promega). RT-R-MDA-MB-231 cells (5 × 10
3 cells/60 μL) were seeded in a round-bottom 96-well plate and NK-92MI cells were added. After co-incubation for 4 h at 37 °C, the culture medium was centrifuged at 1000 rpm for 1 min, and the supernatant was obtained. To determine the effect of MBZ on NK cell-mediated cytotoxicity against cancer cells, RT-R-MDA-MB-231 cells were treated with MBZ (0.1, 0.5, and 1 μM) for 24 h, and then the cells (5 × 10
3 cells/60 μL) were seeded in a round-bottom 96-well plate. NK-92MI cells were added at a 1:10 ratio (cancer cell: NK cell) in the 96-well plate containing RT-R-MDA-MB-231 cells. For MBZ + RT co-treatment, RT-R-MDA-MB-231 cells were treated with MBZ (0.5 μM) for 1 h and then irradiated with a 3 Gy dose of X-rays. After 24 h, the cells (5 × 10
3 cells/60 μL) were seeded in a round-bottom 96-well plate and NK-92MI cells were added at a 1:10 ratio (cancer cell: NK cell) in the 96-well plate containing RT-R-MDA-MB-231 cells. The levels of lactate dehydrogenase (LDH) released into the supernatant were determined as described in the manufacturer’s protocol. Cytotoxicity was quantified as follows:
4.10. Measurement of Extracellular Perforin and Granzyme B Levels
The extracellular levels of perforin and granzyme B were measured using Human perforin and granzyme ELISA Kits (MyBioSource, San Diego, CA, USA), respectively. RT-R-MDA-MB-231 cells were pretreated with MBZ (0.5 μM) for 1 h before being irradiated with a 3 Gy dose of X-rays. After 24 h, the cells (5 × 103 cells/60 μL) were seeded in a round-bottom 96-well plate, and NK-92MI cells (cancer cell: NK cell ratio = 1:10) were added. After co-incubation for 4 h at 37 °C, supernatants were obtained from the culture medium by centrifugation at 1000 rpm for 1 min. Perforin and granzyme B levels in cell supernatants were measured according to the manufacturer’s instructions.
4.11. Statistical Analysis
All data were statistically analyzed using GraphPad Prism 7 software (GraphPad Software, San Diego, CA, USA). One-way ANOVA followed by Tukey’s multiple comparison test was performed to compare differences among groups. Data are presented as mean ± standard deviation (SD).


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







