

Apoptosis, Proliferation, and Autophagy Are Involved in Local Anesthetic-Induced Cytotoxicity of Human Breast Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
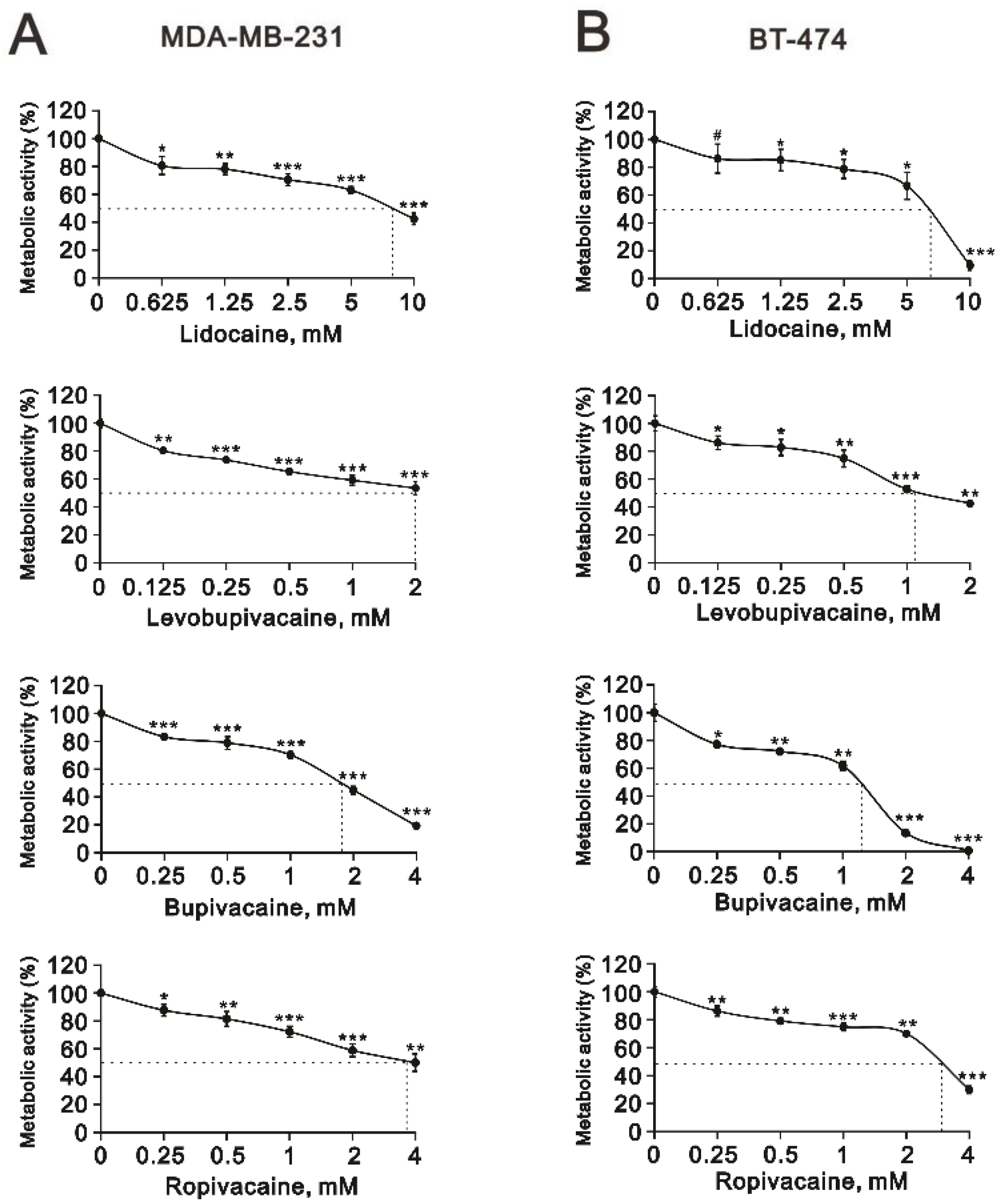
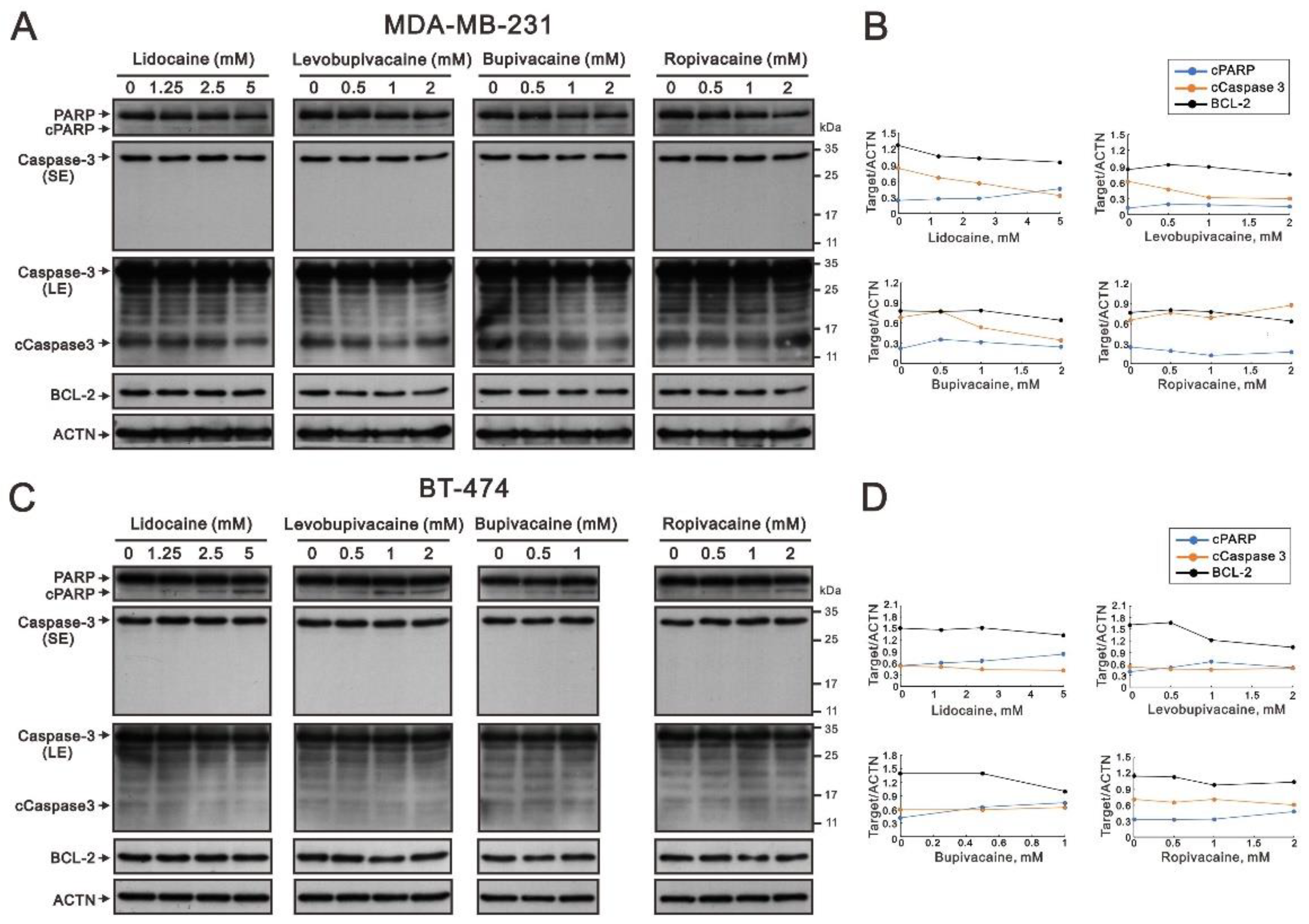
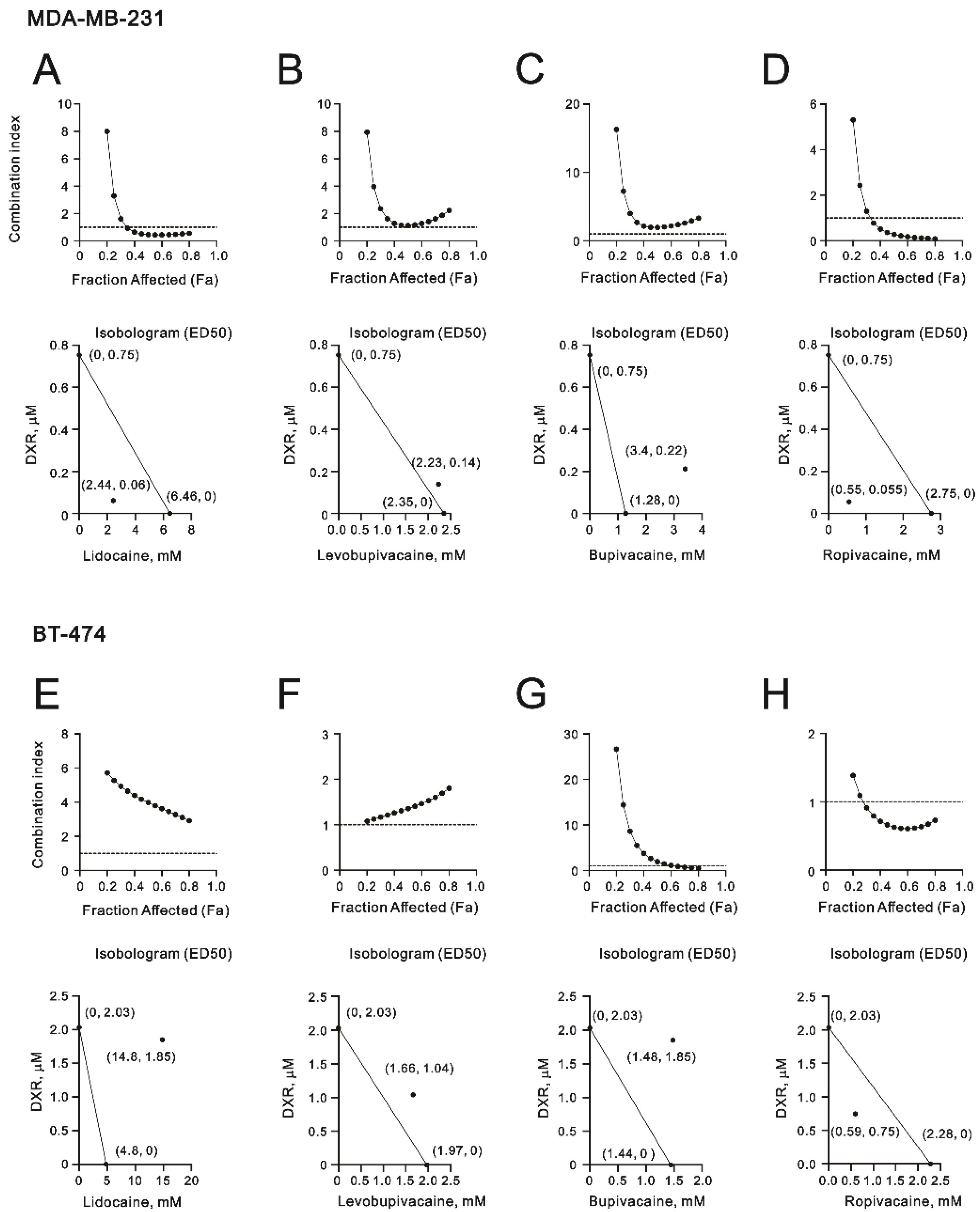
2.1. The Cytotoxic Effects of Lidocaine, Levobupivacaine, Bupivacaine, and Ropivacaine on Two Human Breast Cancer Cell Lines (MDA-MB-231 and BT-474)
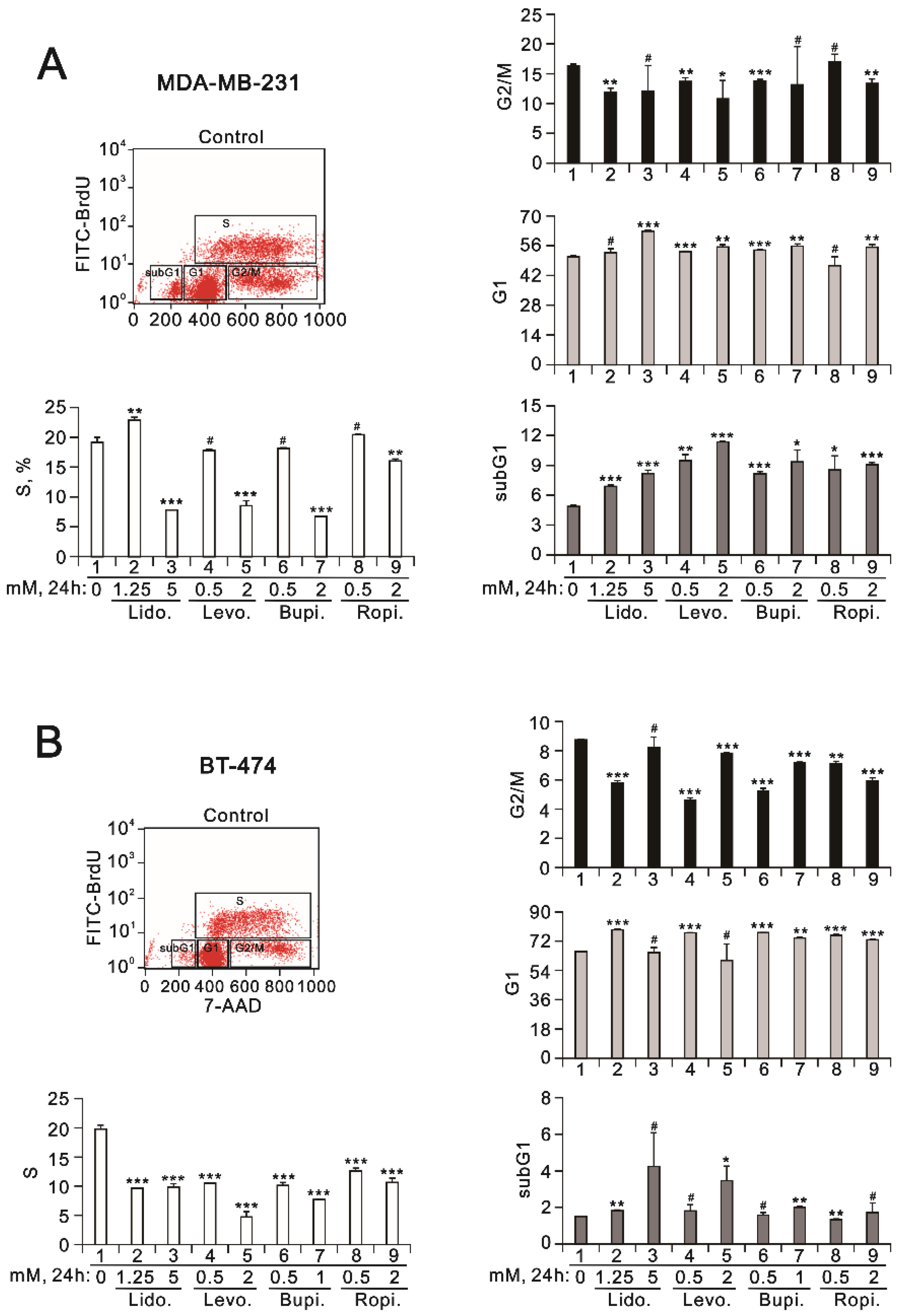
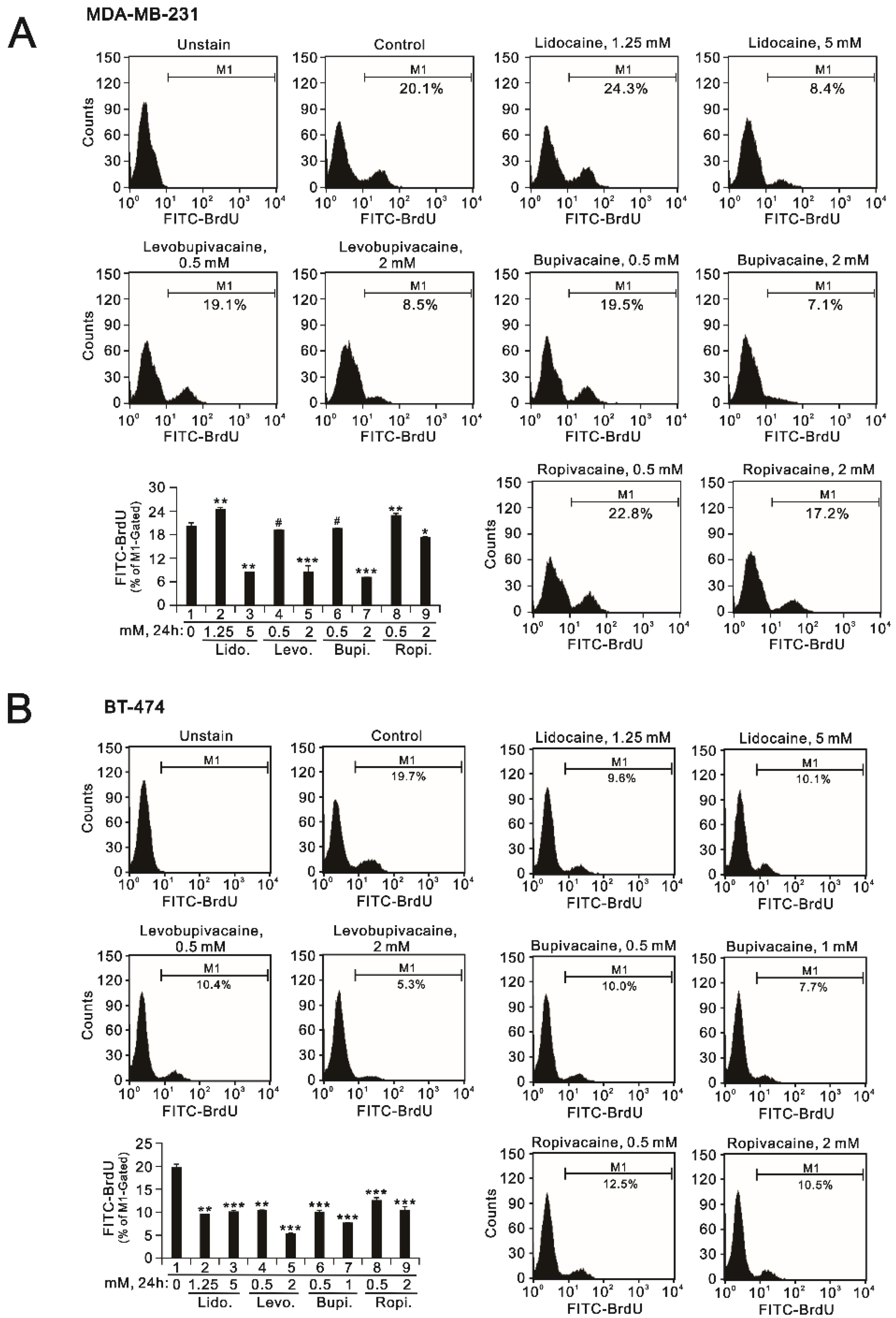
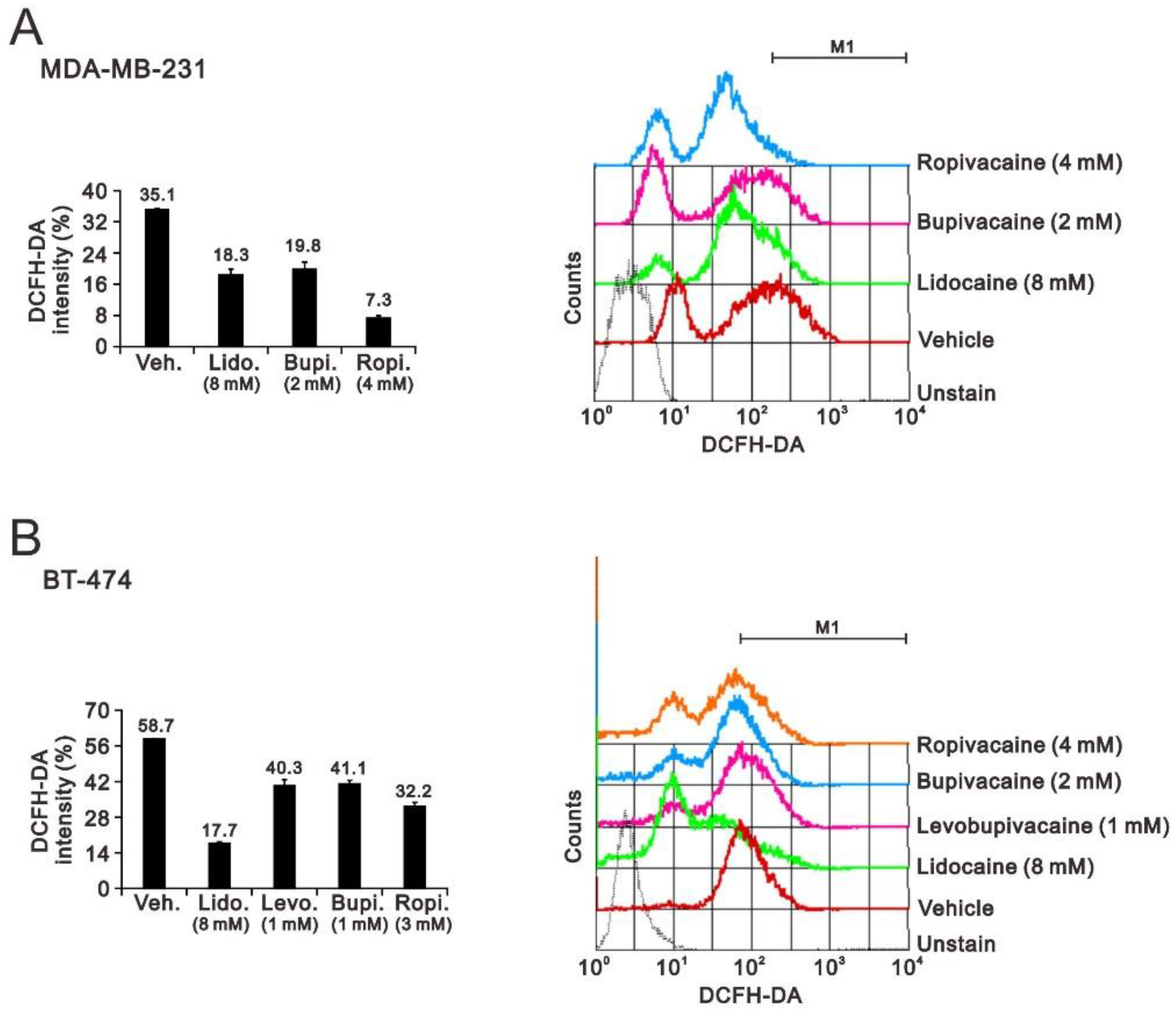
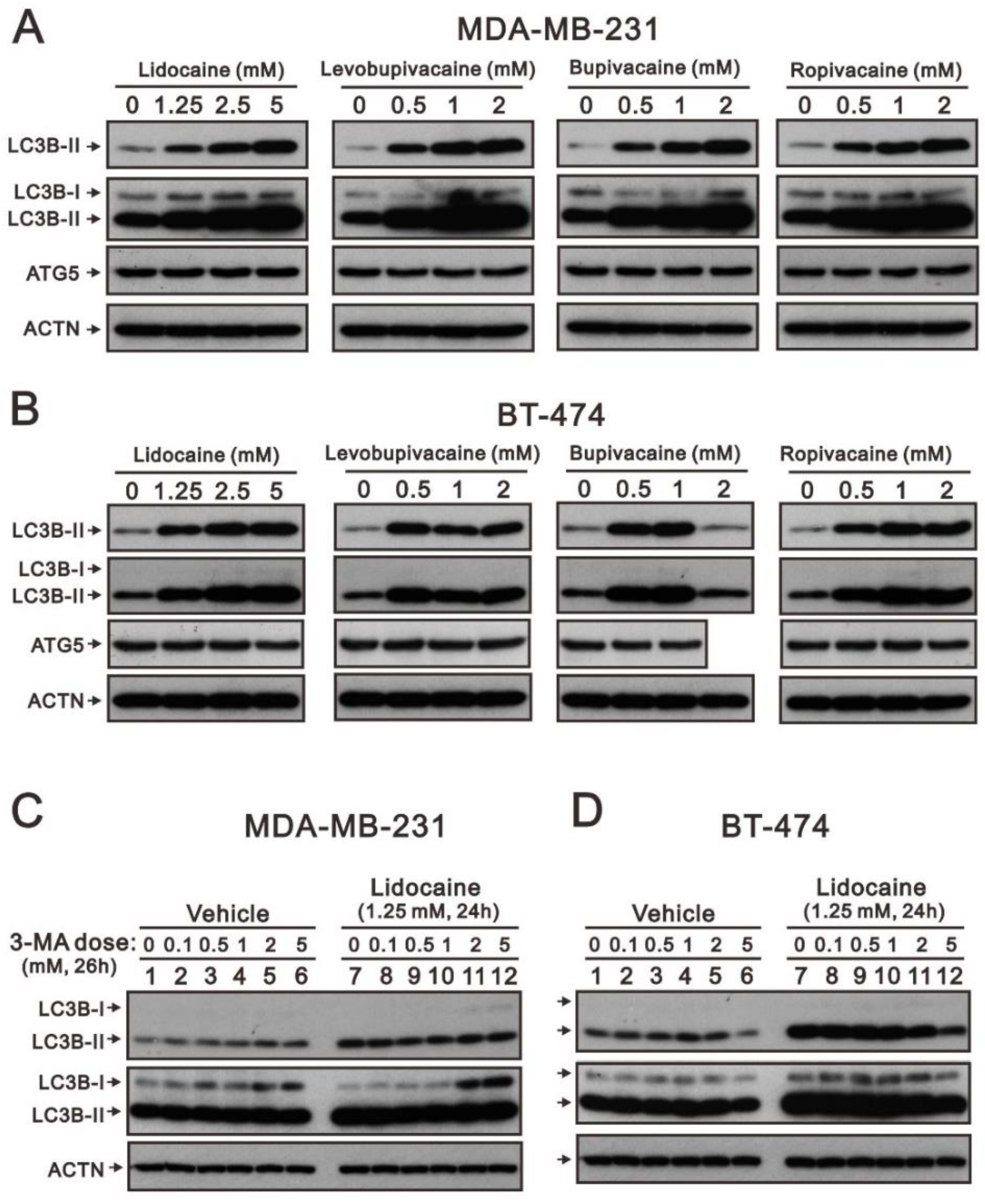
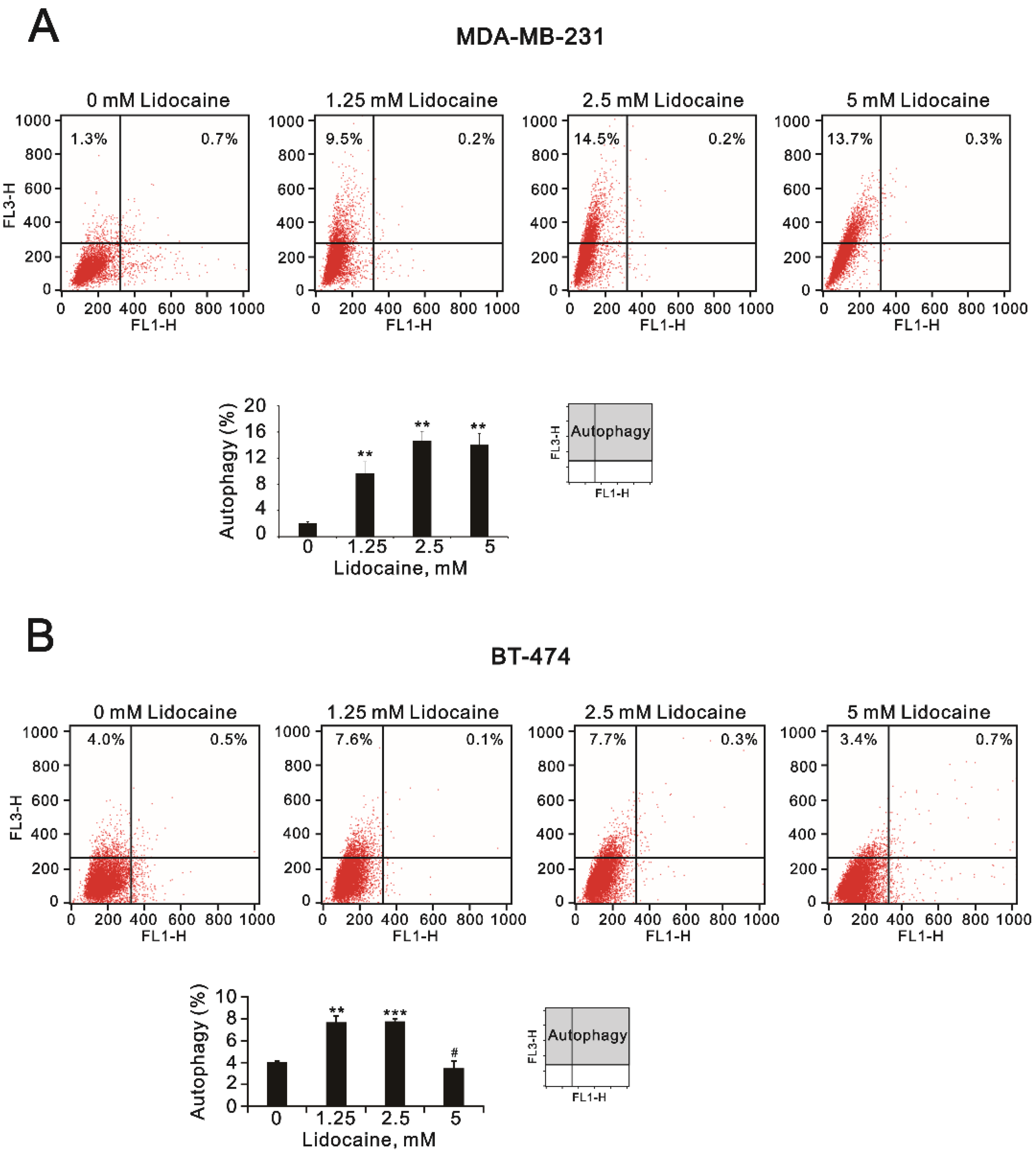
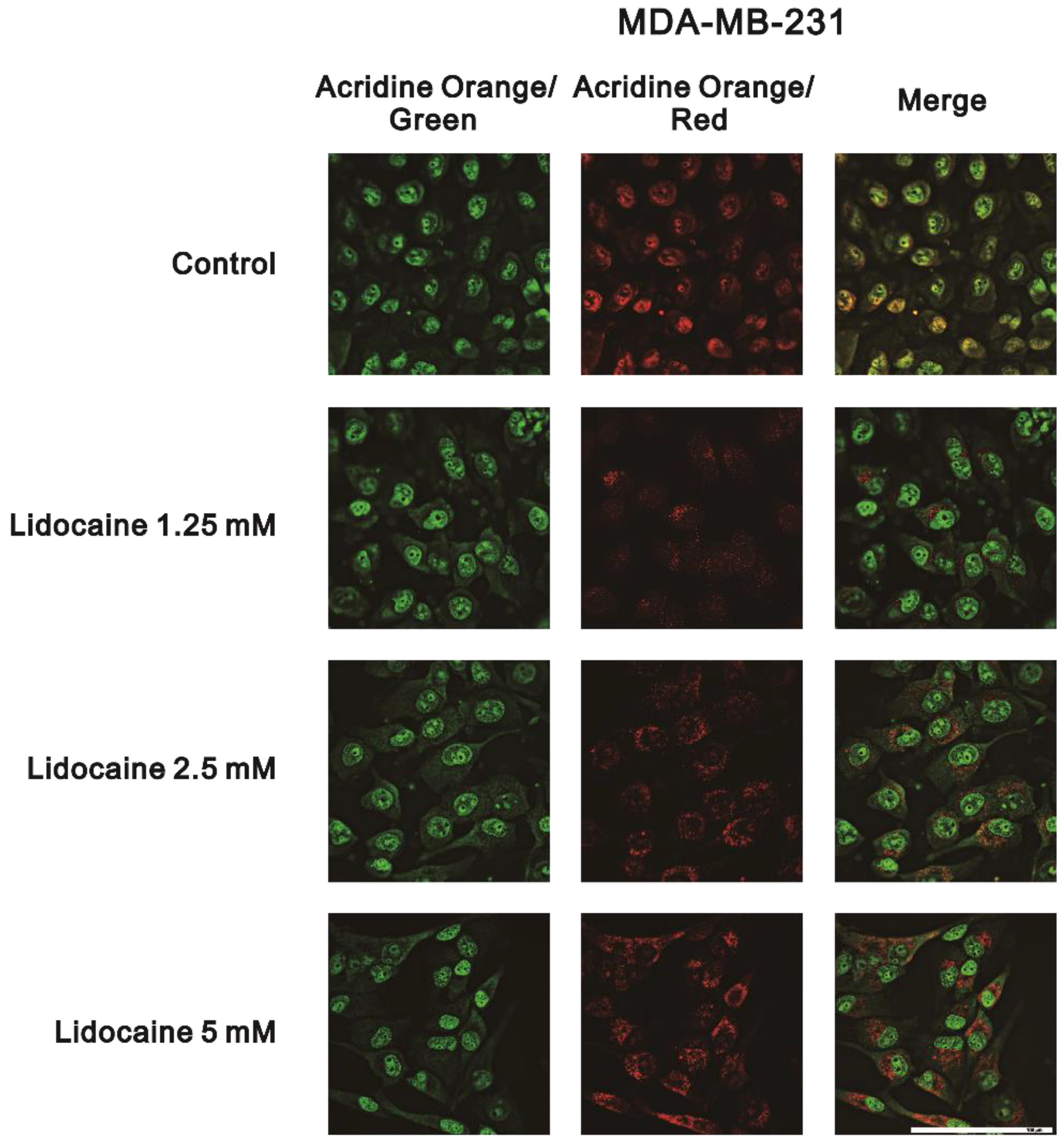
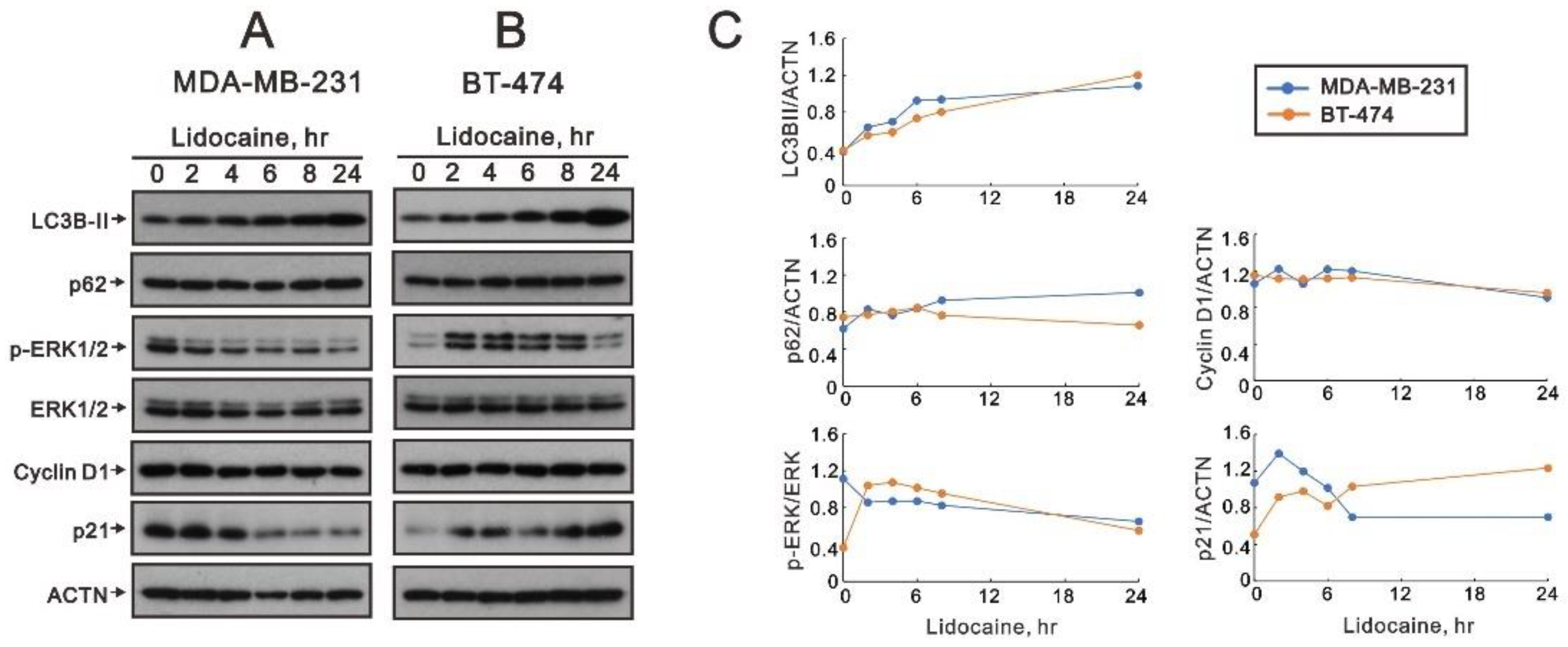
2.2. The Effects of Lidocaine, Levobupivacaine, Bupivacaine, and Ropivacaine on the Induction of Autophagy in MDA-MB-231 and BT-474 Cell Lines
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Reagents
4.2. Metabolic Activity Analysis
4.3. Western Blotting Analysis
4.4. Fluorescence-Activated Cell Sorting (FACS) for Flow Cytometry Analyses of Cell Cycle Profiles, Proliferation, and ROS
4.5. Flow Cytometric Quantification of Acidic Vesicular Organelles
4.6. Detection of Acidic Vesicular Organelles Using a Fluorescence Microscope
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast cancer. Nat. Rev. Dis. Primers 2019, 5, 66. [Google Scholar] [CrossRef]
- Liao, M.; Zhang, J.; Wang, G.; Wang, L.; Liu, J.; Ouyang, L.; Liu, B. Small-Molecule Drug Discovery in Triple Negative Breast Cancer: Current Situation and Future Directions. J. Med. Chem. 2021, 64, 2382–2418. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Sudhan, D.R.; Arteaga, C.L. Overcoming Endocrine Resistance in Breast Cancer. Cancer Cell 2020, 37, 496–513. [Google Scholar] [CrossRef] [PubMed]
- Exadaktylos, A.K.; Buggy, D.J.; Moriarty, D.C.; Mascha, E.; Sessler, D.I. Can anesthetic technique for primary breast cancer surgery affect recurrence or metastasis? Anesthesiology 2006, 105, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gonzalez, O.; Cuellar-Guzman, L.F.; Soliz, J.; Cata, J.P. Impact of Regional Anesthesia on Recurrence, Metastasis, and Immune Response in Breast Cancer Surgery: A Systematic Review of the Literature. Reg. Anesth. Pain Med. 2017, 42, 751–756. [Google Scholar] [CrossRef]
- Sessler, D.I.; Riedel, B. Anesthesia and Cancer Recurrence: Context for Divergent Study Outcomes. Anesthesiology 2019, 130, 3–5. [Google Scholar] [CrossRef]
- Wong, H.Y.; Pilling, R.; Young, B.W.M.; Owolabi, A.A.; Onwochei, D.N.; Desai, N. Comparison of local and regional anesthesia modalities in breast surgery: A systematic review and network meta-analysis. J. Clin. Anesth. 2021, 72, 110274. [Google Scholar] [CrossRef]
- Haller, I.; Hausott, B.; Tomaselli, B.; Keller, C.; Klimaschewski, L.; Gerner, P.; Lirk, P. Neurotoxicity of lidocaine involves specific activation of the p38 mitogen-activated protein kinase, but not extracellular signal-regulated or c-jun N-terminal kinases, and is mediated by arachidonic acid metabolites. Anesthesiology 2006, 105, 1024–1033. [Google Scholar] [CrossRef]
- Lirk, P.; Haller, I.; Hausott, B.; Ingorokva, S.; Deibl, M.; Gerner, P.; Klimaschewski, L. The neurotoxic effects of amitriptyline are mediated by apoptosis and are effectively blocked by inhibition of caspase activity. Anesth. Analg. 2006, 102, 1728–1733. [Google Scholar] [CrossRef]
- Mammoto, T.; Higashiyama, S.; Mukai, M.; Mammoto, A.; Ayaki, M.; Mashimo, T.; Hayashi, Y.; Kishi, Y.; Nakamura, H.; Akedo, H. Infiltration anesthetic lidocaine inhibits cancer cell invasion by modulating ectodomain shedding of heparin-binding epidermal growth factor-like growth factor (HB-EGF). J. Cell Physiol. 2002, 192, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Han, B.S.; Jung, K.H.; Lee, J.E.; Yoon, Y.C.; Ko, S.; Park, M.S.; Lee, Y.J.; Kim, S.E.; Cho, Y.J.; Lee, P.; et al. Lidocaine enhances the efficacy of palbociclib in triple-negative breast cancer. Am. J. Cancer Res. 2022, 12, 3083–3098. [Google Scholar]
- Liu, H.; Dilger, J.P.; Lin, J. Effects of local anesthetics on cancer cells. Pharmacol. Ther. 2020, 212, 107558. [Google Scholar] [CrossRef]
- Cata, J.P.; Ramirez, M.F.; Perez-Gonzalez, O. Local Anesthetics: Hunting for the Holy Grail of Onco-anesthesia. Pain. Med. 2020, 21, 219–220. [Google Scholar] [CrossRef] [PubMed]
- Boland, J.W.; Pockley, A.G. Influence of opioids on immune function in patients with cancer pain: From bench to bedside. Br. J. Pharmacol. 2018, 175, 2726–2736. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Lou, A.; Li, Z.; Yang, J. Lidocaine inhibits growth, migration and invasion of gastric carcinoma cells by up-regulation of miR-145. BMC Cancer 2019, 19, 233. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Wen, X.; Li, H.; He, J.; Wang, Y.; Wu, H.; Wang, H.; Wang, X. Up-regulation of Cav3.1 expression in SH-SY5Y cells induced by lidocaine hydrochloride. Artif. Cells Nanomed. Biotechnol. 2018, 46, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, R.; Cheng, Y.; Wu, X.; Xi, S.; Sun, Y.; Jiang, H. Lidocaine inhibits the proliferation of lung cancer by regulating the expression of GOLT1A. Cell Prolif. 2017, 50, e12364. [Google Scholar] [CrossRef]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef]
- Devenish, R.J.; Klionsky, D.J. Autophagy: Mechanism and physiological relevance ‘brewed’ from yeast studies. Front. Biosci. Schol. Ed. 2012, 4, 1354–1363. [Google Scholar] [CrossRef]
- Debnath, J. The multifaceted roles of autophagy in tumors-implications for breast cancer. J. Mammary Gland Biol. Neoplasia 2011, 16, 173–187. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Baehrecke, E.H.; Brumell, J.H.; Chu, C.T.; Codogno, P.; Cuervo, A.M.; Debnath, J.; Deretic, V.; Elazar, Z.; Eskelinen, E.L.; et al. A comprehensive glossary of autophagy-related molecules and processes (2nd edition). Autophagy 2011, 7, 1273–1294. [Google Scholar] [CrossRef]
- Thorburn, A.; Debnath, J. Targeting chaperone-mediated autophagy in cancer. Sci. Transl. Med. 2011, 3, 109ps145. [Google Scholar] [CrossRef] [PubMed]
- Dower, C.M.; Wills, C.A.; Frisch, S.M.; Wang, H.G. Mechanisms and context underlying the role of autophagy in cancer metastasis. Autophagy 2018, 14, 1110–1128. [Google Scholar] [CrossRef]
- Mowers, E.E.; Sharifi, M.N.; Macleod, K.F. Autophagy in cancer metastasis. Oncogene 2017, 36, 1619–1630. [Google Scholar] [CrossRef]
- Korner, J.; Albani, S.; Sudha Bhagavath Eswaran, V.; Roehl, A.B.; Rossetti, G.; Lampert, A. Sodium Channels and Local Anesthetics-Old Friends With New Perspectives. Front. Pharmacol. 2022, 13, 837088. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Gupte, A.; Mumper, R.J. Elevated copper and oxidative stress in cancer cells as a target for cancer treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Nicco, C.; Batteux, F. ROS Modulator Molecules with Therapeutic Potential in Cancers Treatments. Molecules 2017, 23, 84. [Google Scholar] [CrossRef]
- Wang, F.; Jiao, P.; Qi, M.; Frezza, M.; Dou, Q.P.; Yan, B. Turning tumor-promoting copper into an anti-cancer weapon via high-throughput chemistry. Curr. Med. Chem. 2010, 17, 2685–2698. [Google Scholar] [CrossRef]
- Wang, J.; Yi, J. Cancer cell killing via ROS: To increase or decrease, that is the question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012, 645460. [Google Scholar] [CrossRef]
- Denton, D.; Kumar, S. Autophagy-dependent cell death. Cell Death Differ. 2019, 26, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Kriel, J.; Loos, B. The good, the bad and the autophagosome: Exploring unanswered questions of autophagy-dependent cell death. Cell Death Differ. 2019, 26, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, B.L.; Yang, X.; Zhang, X.; Liu, J. The autophagic inhibitor 3-methyladenine potently stimulates PKA-dependent lipolysis in adipocytes. Br. J. Pharmacol. 2013, 168, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Millot, C.; Millot, J.M.; Morjani, H.; Desplaces, A.; Manfait, M. Characterization of acidic vesicles in multidrug-resistant and sensitive cancer cells by acridine orange staining and confocal microspectrofluorometry. J. Histochem. Cytochem. 1997, 45, 1255–1264. [Google Scholar] [CrossRef]
- Li, R.; Xiao, C.; Liu, H.; Huang, Y.; Dilger, J.P.; Lin, J. Effects of local anesthetics on breast cancer cell viability and migration. BMC Cancer 2018, 18, 666. [Google Scholar] [CrossRef]
- Safety Committee of Japanese Society of Anesthesiologists. Practical guide for the management of systemic toxicity caused by local anesthetics. J. Anesth. 2019, 33, 1–8. [Google Scholar] [CrossRef]
- Mather, L.E.; Chang, D.H. Cardiotoxicity with modern local anaesthetics: Is there a safer choice? Drugs 2001, 61, 333–342. [Google Scholar] [CrossRef]
- Xiong, J.; Kong, Q.; Dai, L.; Ma, H.; Cao, X.; Liu, L.; Ding, Z. Autophagy activated by tuberin/mTOR/p70S6K suppression is a protective mechanism against local anaesthetics neurotoxicity. J. Cell. Mol. Med. 2017, 21, 579–587. [Google Scholar] [CrossRef]
- Zhang, C.; Xie, C.; Lu, Y. Local Anesthetic Lidocaine and Cancer: Insight Into Tumor Progression and Recurrence. Front. Oncol. 2021, 11, 669746. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Wang, L.; Cui, Q.; Iftikhar, R.; Xia, Y.; Xu, P. Repositioning Lidocaine as an Anticancer Drug: The Role Beyond Anesthesia. Front. Cell. Dev. Biol. 2020, 8, 565. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhu, G.; Xu, W.; Dan, J.; Xia, R.; Liu, W. Bupivacaine inhibits angiogenesis through oxidative stress-dependent inhibition of Akt/mTOR and activation of AMPK. Fundam. Clin. Pharmacol. 2020, 34, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Matkar, S.; An, C.; Hua, X. Kinase inhibitors of HER2/AKT pathway induce ERK phosphorylation via a FOXO-dependent feedback loop. Am. J. Cancer Res. 2017, 7, 1476–1485. [Google Scholar]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Myhre, O.; Sterri, S.H.; Bogen, I.L.; Fonnum, F. Erk1/2 phosphorylation and reactive oxygen species formation via nitric oxide and Akt-1/Raf-1 crosstalk in cultured rat cerebellar granule cells exposed to the organic solvent 1,2,4-trimethylcyclohexane. Toxicol. Sci. 2004, 80, 296–303. [Google Scholar] [CrossRef]
- Call, T.R.; Pace, N.L.; Thorup, D.B.; Maxfield, D.; Chortkoff, B.; Christensen, J.; Mulvihill, S.J. Factors associated with improved survival after resection of pancreatic adenocarcinoma: A multivariable model. Anesthesiology 2015, 122, 317–324. [Google Scholar] [CrossRef]
- Royds, J.; Khan, A.H.; Buggy, D.J. An Update on Existing Ongoing Prospective Trials Evaluating the Effect of Anesthetic and Analgesic Techniques During Primary Cancer Surgery on Cancer Recurrence or Metastasis. Int. Anesthesiol. Clin. 2016, 54, e76–e83. [Google Scholar] [CrossRef]
- Sessler, D.I.; Pei, L.; Huang, Y.; Fleischmann, E.; Marhofer, P.; Kurz, A.; Mayers, D.B.; Meyer-Treschan, T.A.; Grady, M.; Tan, E.Y.; et al. Recurrence of breast cancer after regional or general anaesthesia: A randomised controlled trial. Lancet 2019, 394, 1807–1815. [Google Scholar] [CrossRef]
- Singh, N.P.; Makkar, J.K.; Kuberan, A.; Guffey, R.; Uppal, V. Efficacy of regional anesthesia techniques for postoperative analgesia in patients undergoing major oncologic breast surgeries: A systematic review and network meta-analysis of randomized controlled trials. Can. J. Anaesth. 2022, 69, 527–549. [Google Scholar] [CrossRef]
- Hung, K.C.; Ko, C.C.; Hsu, C.W.; Pang, Y.L.; Chen, J.Y.; Sun, C.K. Association of peripheral nerve blocks with patient-reported quality of recovery in female patients receiving breast cancer surgery: A systematic review and meta-analysis of randomized controlled studies. Can. J. Anaesth. 2022, 69, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, L.; Buggy, D.J. The role of regional anaesthesia in the emerging subspecialty of onco-anaesthesia: A state-of-the-art review. Anaesthesia 2021, 76 (Suppl. 1), 148–159. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, G.; Saporito, A.; Cecchinato, V.; Silvestri, Y.; Borgeat, A.; Anselmi, L.; Uguccioni, M. Lidocaine inhibits cytoskeletal remodelling and human breast cancer cell migration. Br. J. Anaesth. 2018, 121, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Sanderson, C.R.; Xuan, W.; Agar, M. Lidocaine for Cancer Pain in Adults: A Systematic Review and Meta-Analysis. J. Palliat. Med. 2019, 22, 326–334. [Google Scholar] [CrossRef]
- Lirk, P.; Hollmann, M.W.; Fleischer, M.; Weber, N.C.; Fiegl, H. Lidocaine and ropivacaine, but not bupivacaine, demethylate deoxyribonucleic acid in breast cancer cells in vitro. Br. J. Anaesth. 2014, 113 (Suppl. 1), i32–i38. [Google Scholar] [CrossRef]
- Lin, C.K.; Liu, S.T.; Wu, Z.S.; Wang, Y.C.; Huang, S.M. Mechanisms of Cisplatin in Combination with Repurposed Drugs against Human Endometrial Carcinoma Cells. Life 2021, 11, 160. [Google Scholar] [CrossRef]
- Chen, S.Y.; Chang, Y.L.; Liu, S.T.; Chen, G.S.; Lee, S.P.; Huang, S.M. Differential Cytotoxicity Mechanisms of Copper Complexed with Disulfiram in Oral Cancer Cells. Int. J. Mol. Sci. 2021, 22, 3711. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.-L.; Liu, S.-T.; Huang, S.-M.; Wu, Z.-F. Apoptosis, Proliferation, and Autophagy Are Involved in Local Anesthetic-Induced Cytotoxicity of Human Breast Cancer Cells. Int. J. Mol. Sci. 2022, 23, 15455. https://doi.org/10.3390/ijms232415455
Chen J-L, Liu S-T, Huang S-M, Wu Z-F. Apoptosis, Proliferation, and Autophagy Are Involved in Local Anesthetic-Induced Cytotoxicity of Human Breast Cancer Cells. International Journal of Molecular Sciences. 2022; 23(24):15455. https://doi.org/10.3390/ijms232415455
Chicago/Turabian StyleChen, Jia-Lin, Shu-Ting Liu, Shih-Ming Huang, and Zhi-Fu Wu. 2022. "Apoptosis, Proliferation, and Autophagy Are Involved in Local Anesthetic-Induced Cytotoxicity of Human Breast Cancer Cells" International Journal of Molecular Sciences 23, no. 24: 15455. https://doi.org/10.3390/ijms232415455
APA StyleChen, J.-L., Liu, S.-T., Huang, S.-M., & Wu, Z.-F. (2022). Apoptosis, Proliferation, and Autophagy Are Involved in Local Anesthetic-Induced Cytotoxicity of Human Breast Cancer Cells. International Journal of Molecular Sciences, 23(24), 15455. https://doi.org/10.3390/ijms232415455