
Porphyromonas gingivalis Fimbriae Induce Osteoclastogenesis via Toll-like Receptors in RAW264 Cells
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Mfa1 and FimA Fimbriae Promote RANKL-Mediated Osteoclast Differentiation
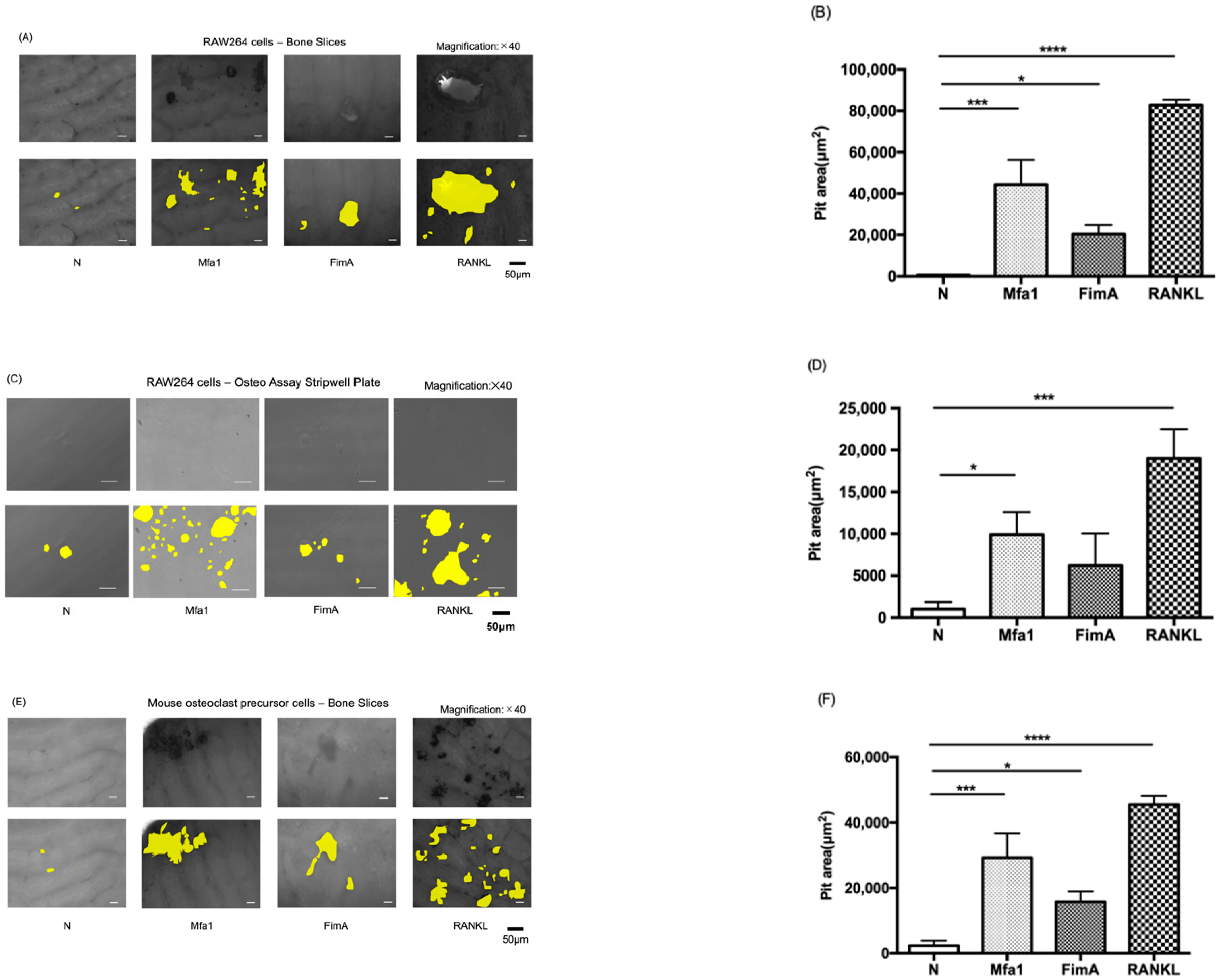
2.2. Mfa1 and FimA Fimbriae Promote RANKL-Induced Osteoclastic Bone Resorption
2.3. Mfa1 Fimbriae Synergistically Induce Expression of RANKL-Dependent Osteoclast Differentiation Markers
2.4. Mfa1 and FimA Fimbriae Increase Gene Expression of TLR2 and TLR4 in RANKL-Induced Osteoclasts
2.5. Transfection of Tlr2 and Tlr4 siRNAs into RAW264 Cells
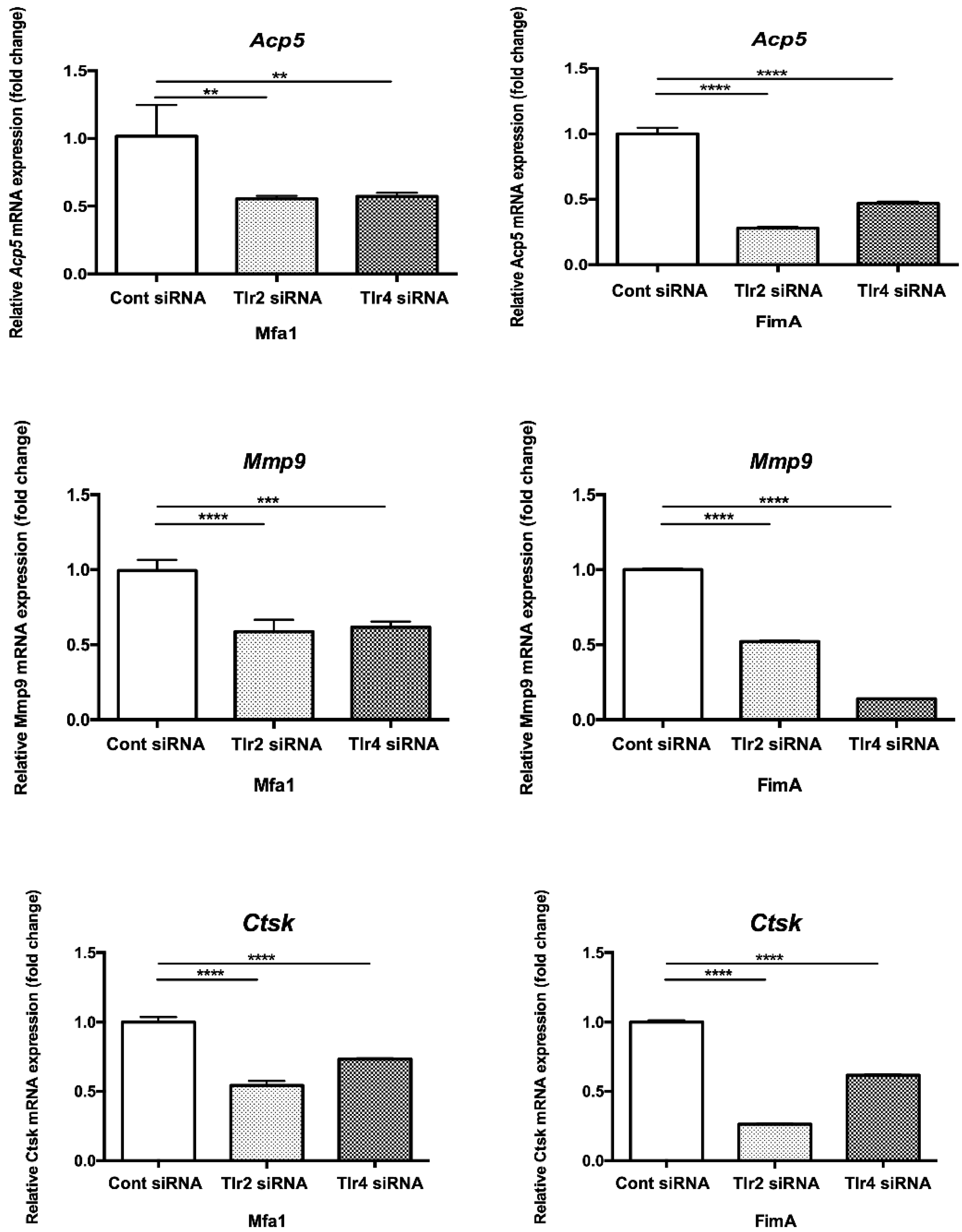
2.6. Mfa1 Fimbriae Induce Osteoclast Differentiation Primarily through Recognition by TLR2
2.7. Mfa1 Fimbriae Induce Osteoclast Differentiation Markers Primarily through Recognition by TLRs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Bacterial Strains and Growth Conditions
4.3. Purification of Fimbriae
4.4. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.5. Bone Resorption Assay
4.6. Real-Time qPCR
4.7. siRNA Transfection
4.8. Flow Cytometry
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhou, W.; Wang, H.; Liang, S. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv. Protein Chem. Struct. Biol. 2020, 120, 45–84. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, F.; Murakami, Y.; Nishikawa, K.; Hasegawa, Y.; Kawaminami, S. Surface components of Porphyromonas gingivalis. J. Periodontal. Res. 2009, 44, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Nagano, K. Porphyromonas gingivalis FimA and Mfa1 fimbriae: Current insights on localization, function, biogenesis, and genotype. Jpn. Dent. Sci Rev. 2021, 57, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Kuboniwa, M.; Inaba, H.; Amano, A. Genotyping to distinguish microbial pathogenicity in periodontitis. Periodontol. 2000 2010, 54, 136–159. [Google Scholar] [CrossRef]
- Zhao, L.; Wu, Y.F.; Meng, S.; Yang, H.; OuYang, Y.L.; Zhou, X.D. Prevalence of fimA genotypes of Porphyromonas gingivalis and periodontal health status in Chinese adults. J. Periodontal. Res. 2007, 42, 511–517. [Google Scholar] [CrossRef]
- Amano, A.; Nakagawa, I.; Kataoka, K.; Morisaki, I.; Hamada, S. Distribution of Porphyromonas gingivalis strains with fimA genotypes in periodontitis patients. J. Clin. Microbiol 1999, 37, 1426–1430. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Iijima, Y.; Persson, K.; Nagano, K.; Yoshida, Y.; Lamont, R.J.; Kikuchi, T.; Mitani, A.; Yoshimura, F. Role of Mfa5 in Expression of Mfa1 Fimbriae in Porphyromonas gingivalis. J. Dent. Res. 2016, 95, 1291–1297. [Google Scholar] [CrossRef]
- Koide, M.; Kinugawa, S.; Takahashi, N.; Udagawa, N. Osteoclastic bone resorption induced by innate immune responses. Periodontol. 2000 2010, 54, 235–246. [Google Scholar] [CrossRef]
- Holt, S.C.; Ebersole, J.L. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: The "red complex", a prototype polybacterial pathogenic consortium in periodontitis. Periodontol. 2000 2005, 38, 72–122. [Google Scholar] [CrossRef]
- Hamada, S.; Amano, A.; Kimura, S.; Nakagawa, I.; Kawabata, S.; Morisaki, I. The importance of fimbriae in the virulence and ecology of some oral bacteria. Oral Microbiol. Immunol. 1998, 13, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, T.; Hamada, N. Characterization of biologically active cell surface components of a periodontal pathogen. The roles of major and minor fimbriae of Porphyromonas gingivalis. J. Periodontol. 2003, 74, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Lambris, J.D. Microbial manipulation of receptor crosstalk in innate immunity. Nat. Rev. Immunol. 2011, 11, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.; Bachrach, G.; Shapira, L.; Nussbaum, G. Cutting Edge: TLR2 is required for the innate response to Porphyromonas gingivalis: Activation leads to bacterial persistence and TLR2 deficiency attenuates induced alveolar bone resorption. J. Immunol. 2006, 177, 8296–8300. [Google Scholar] [CrossRef]
- Watanabe, N.; Yokoe, S.; Ogata, Y.; Sato, S.; Imai, K. Exposure to Porphyromonas gingivalis Induces Production of Proinflammatory Cytokine via TLR2 from Human Respiratory Epithelial Cells. J. Clin. Med. 2020, 9, 3433. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Kikuchi, T.; Hasegawa, Y.; Naiki, Y.; Goto, H.; Okada, K.; Okabe, I.; Kamiya, Y.; Suzuki, Y.; Sawada, N.; et al. Porphyromonas gingivalis Mfa1 Induces Chemokine and Cell Adhesion Molecules in Mouse Gingival Fibroblasts via Toll-Like Receptors. J. Clin. Med. 2020, 9, 4004. [Google Scholar] [CrossRef]
- Takahashi, Y.; Cueno, M.E.; Kamio, N.; Iinuma, T.; Hasegawa, Y.; Imai, K. Porphyromonas gingivalis Mfa1 fimbria putatively binds to TLR2 and induces both IL-6 and IL-8 production in human bronchial epithelial cells. Biochem. Biophys. Res. Commun. 2022, 589, 35–40. [Google Scholar] [CrossRef]
- Hiramine, H.; Watanabe, K.; Hamada, N.; Umemoto, T. Porphyromonas gingivalis 67-kDa fimbriae induced cytokine production and osteoclast differentiation utilizing TLR2. FEMS Microbiol. Lett. 2003, 229, 49–55. [Google Scholar] [CrossRef]
- Kawata, Y.; Hanazawa, S.; Amano, S.; Murakami, Y.; Matsumoto, T.; Nishida, K.; Kitano, S. Porphyromonas gingivalis fimbriae stimulate bone resorption in vitro. Infect. Immun. 1994, 62, 3012–3016. [Google Scholar] [CrossRef]
- Shoji, M.; Shibata, S.; Sueyoshi, T.; Naito, M.; Nakayama, K. Biogenesis of Type V pili. Microbiol. Immunol. 2020, 64, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Jimi, E.; Akiyama, S.; Tsurukai, T.; Okahashi, N.; Kobayashi, K.; Udagawa, N.; Nishihara, T.; Takahashi, N.; Suda, T. Osteoclast differentiation factor acts as a multifunctional regulator in murine osteoclast differentiation and function. J. Immunol. 1999, 163, 434–442. [Google Scholar] [PubMed]
- Kobayashi, K.; Takahashi, N.; Jimi, E.; Udagawa, N.; Takami, M.; Kotake, S.; Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; et al. Tumor necrosis factor alpha stimulates osteoclast differentiation by a mechanism independent of the ODF/RANKL-RANK interaction. J. Exp. Med. 2000, 191, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Suda, K.; Woo, J.T.; Takami, M.; Sexton, P.M.; Nagai, K. Lipopolysaccharide supports survival and fusion of preosteoclasts independent of TNF-alpha, IL-1, and RANKL. J. Cell. Physiol. 2002, 190, 101–108. [Google Scholar] [CrossRef]
- Akatsu, T.; Takahashi, N.; Debari, K.; Morita, I.; Murota, S.; Nagata, N.; Takatani, O.; Suda, T. Prostaglandins promote osteoclast like cell formation by a mechanism involving cyclic adenosine 3',5'-monophosphate in mouse bone marrow cell cultures. J. Bone Miner. Res. 1989, 4, 29–35. [Google Scholar] [CrossRef]
- Takami, M.; Kim, N.; Rho, J.; Choi, Y. Stimulation by toll-like receptors inhibits osteoclast differentiation. J. Immunol. 2002, 169, 1516–1523. [Google Scholar] [CrossRef]
- Yang, J.; Ryu, Y.H.; Yun, C.H.; Han, S.H. Impaired osteoclastogenesis by staphylococcal lipoteichoic acid through Toll-like receptor 2 with partial involvement of MyD88. J. Leukoc. Biol. 2009, 86, 823–831. [Google Scholar] [CrossRef]
- Zou, W.; Bar-Shavit, Z. Dual modulation of osteoclast differentiation by lipopolysaccharide. J. Bone Miner. Res. 2002, 17, 1211–1218. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Zhang, P.; Said-Al-Naief, N.; Michalek, S.M.; Feng, X. Molecular mechanism of the bifunctional role of lipopolysaccharide in osteoclastogenesis. J. Biol. Chem. 2009, 284, 12512–12523. [Google Scholar] [CrossRef]
- Hamada, N.; Watanabe, K.; Arai, M.; Hiramine, H.; Umemoto, T. Cytokine production induced by a 67-kDa fimbrial protein from Porphyromonas gingivalis. Oral Microbiol. Immunol. 2002, 17, 197–200. [Google Scholar] [CrossRef]
- Kukita, A.; Ichigi, Y.; Takigawa, I.; Watanabe, T.; Kukita, T.; Miyamoto, H. Infection of RANKL-primed RAW-D macrophages with Porphyromonas gingivalis promotes osteoclastogenesis in a TNF-alpha-independent manner. PLoS ONE 2012, 7, e38500. [Google Scholar] [CrossRef]
- Sasaki, H.; Watanabe, K.; Toyama, T.; Koyata, Y.; Hamada, N. Porphyromonas gulae 41-kDa fimbriae induced osteoclast differentiation and cytokine production. J. Vet. Med. Sci. 2015, 77, 265–271. [Google Scholar] [CrossRef]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptors 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef]
- Cai, J.; Chen, J.; Guo, H.; Pan, Y.; Zhang, Y.; Zhao, W.; Li, X.; Li, Y. Recombinant fimbriae protein of Porphyromonas gingivalis induces an inflammatory response via the TLR4/NFkappaB signaling pathway in human peripheral blood mononuclear cells. Int. J. Mol. Med. 2019, 43, 1430–1440. [Google Scholar] [CrossRef]
- Aoki, Y.; Tabeta, K.; Murakami, Y.; Yoshimura, F.; Yamazaki, K. Analysis of immunostimulatory activity of Porphyromonas gingivalis fimbriae conferred by Toll-like receptor 2. Biochem. Biophys. Res. Commun. 2010, 398, 86–91. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Nagano, K.; Murakami, Y.; Lamont, R.J. Purification of Native Mfa1 Fimbriae from Porphyromonas gingivalis. Methods Mol. Biol. 2021, 2210, 75–86. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Iwami, J.; Sato, K.; Park, Y.; Nishikawa, K.; Atsumi, T.; Moriguchi, K.; Murakami, Y.; Lamont, R.J.; Nakamura, H.; et al. Anchoring and length regulation of Porphyromonas gingivalis Mfa1 fimbriae by the downstream gene product Mfa2. Microbiology 2009, 155, 3333–3347. [Google Scholar] [CrossRef]
- Lamont, R.J.; El-Sabaeny, A.; Park, Y.; Cook, G.S.; Costerton, J.W.; Demuth, D.R. Role of the Streptococcus gordonii SspB protein in the development of Porphyromonas gingivalis biofilms on streptococcal substrates. Microbiology 2002, 148, 1627–1636. [Google Scholar] [CrossRef]
- Park, Y.; Simionato, M.R.; Sekiya, K.; Murakami, Y.; James, D.; Chen, W.; Hackett, M.; Yoshimura, F.; Demuth, D.R.; Lamont, R.J. Short fimbriae of Porphyromonas gingivalis and their role in coadhesion with Streptococcus gordonii. Infect. Immun. 2005, 73, 3983–3989. [Google Scholar] [CrossRef]
- Yoshimura, F.; Takahashi, K.; Nodasaka, Y.; Suzuki, T. Purification and characterization of a novel type of fimbriae from the oral anaerobe Bacteroides gingivalis. J. Bacteriol. 1984, 160, 949–957. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, Y.; Kikuchi, T.; Goto, H.; Takayanagi, Y.; Kawamura, S.; Sawada, N.; Naiki, Y.; Kondo, H.; Hayashi, J.-i.; Hasegawa, Y.; et al. Porphyromonas gingivalis Fimbriae Induce Osteoclastogenesis via Toll-like Receptors in RAW264 Cells. Int. J. Mol. Sci. 2022, 23, 15293. https://doi.org/10.3390/ijms232315293
Suzuki Y, Kikuchi T, Goto H, Takayanagi Y, Kawamura S, Sawada N, Naiki Y, Kondo H, Hayashi J-i, Hasegawa Y, et al. Porphyromonas gingivalis Fimbriae Induce Osteoclastogenesis via Toll-like Receptors in RAW264 Cells. International Journal of Molecular Sciences. 2022; 23(23):15293. https://doi.org/10.3390/ijms232315293
Chicago/Turabian StyleSuzuki, Yuki, Takeshi Kikuchi, Hisashi Goto, Yuhei Takayanagi, Shotaro Kawamura, Noritaka Sawada, Yoshikazu Naiki, Hisataka Kondo, Jun-ichiro Hayashi, Yoshiaki Hasegawa, and et al. 2022. "Porphyromonas gingivalis Fimbriae Induce Osteoclastogenesis via Toll-like Receptors in RAW264 Cells" International Journal of Molecular Sciences 23, no. 23: 15293. https://doi.org/10.3390/ijms232315293
APA StyleSuzuki, Y., Kikuchi, T., Goto, H., Takayanagi, Y., Kawamura, S., Sawada, N., Naiki, Y., Kondo, H., Hayashi, J.-i., Hasegawa, Y., & Mitani, A. (2022). Porphyromonas gingivalis Fimbriae Induce Osteoclastogenesis via Toll-like Receptors in RAW264 Cells. International Journal of Molecular Sciences, 23(23), 15293. https://doi.org/10.3390/ijms232315293