


Photocatalytic Hydrogen Production and Carbon Dioxide Reduction Catalyzed by an Artificial Cobalt Hemoprotein

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
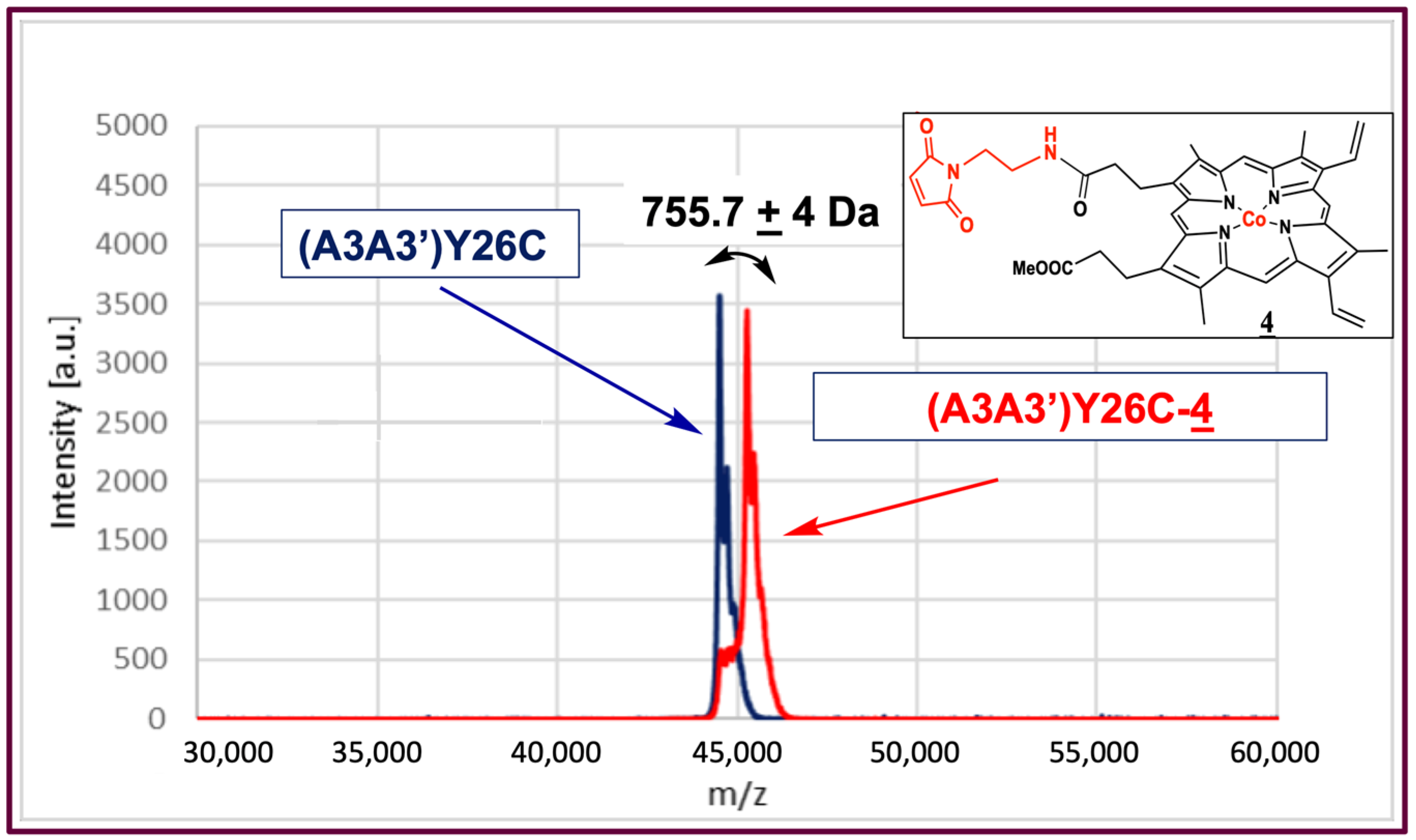
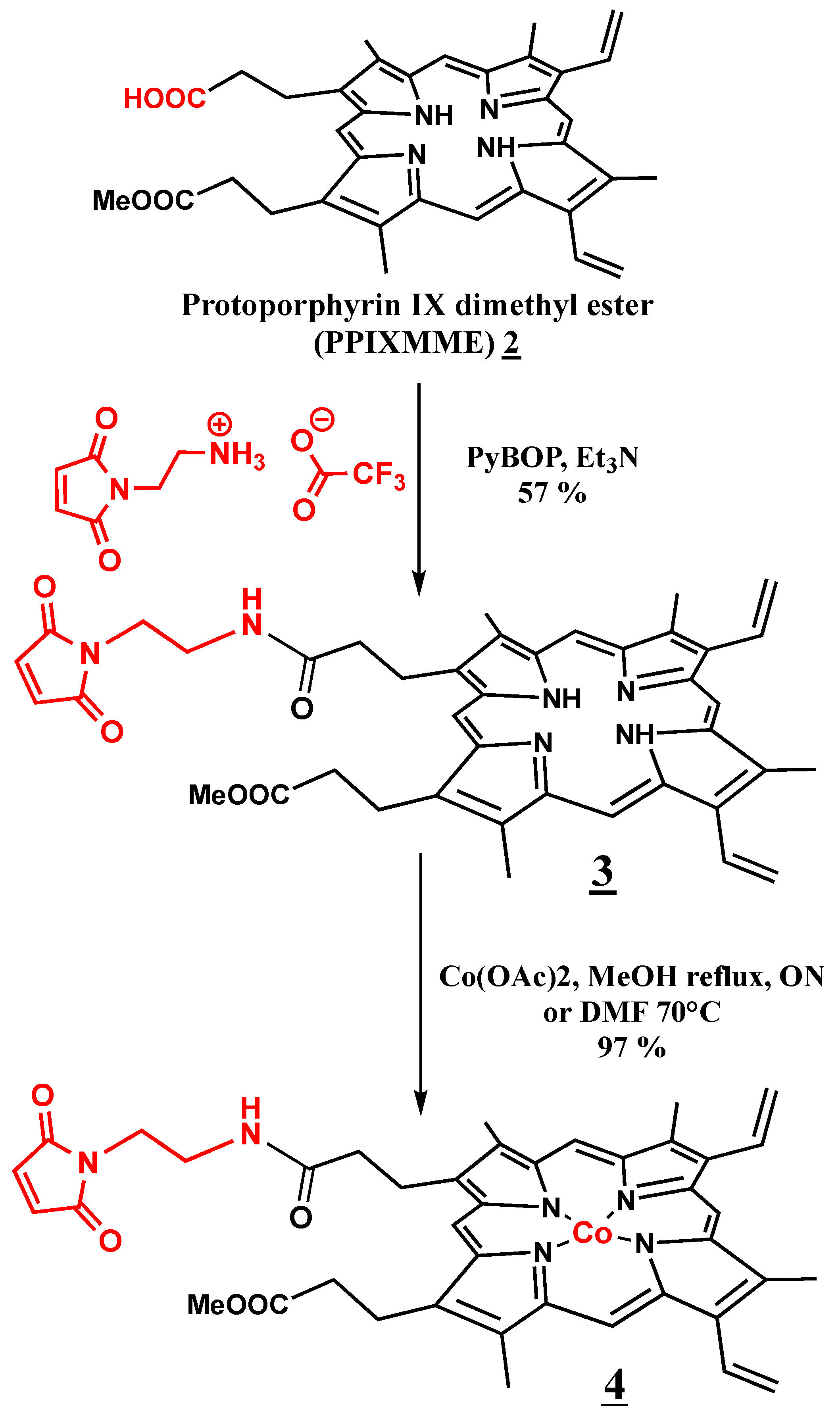
2.1. Synthesis of Co(III)Mal-PPIXMME 4
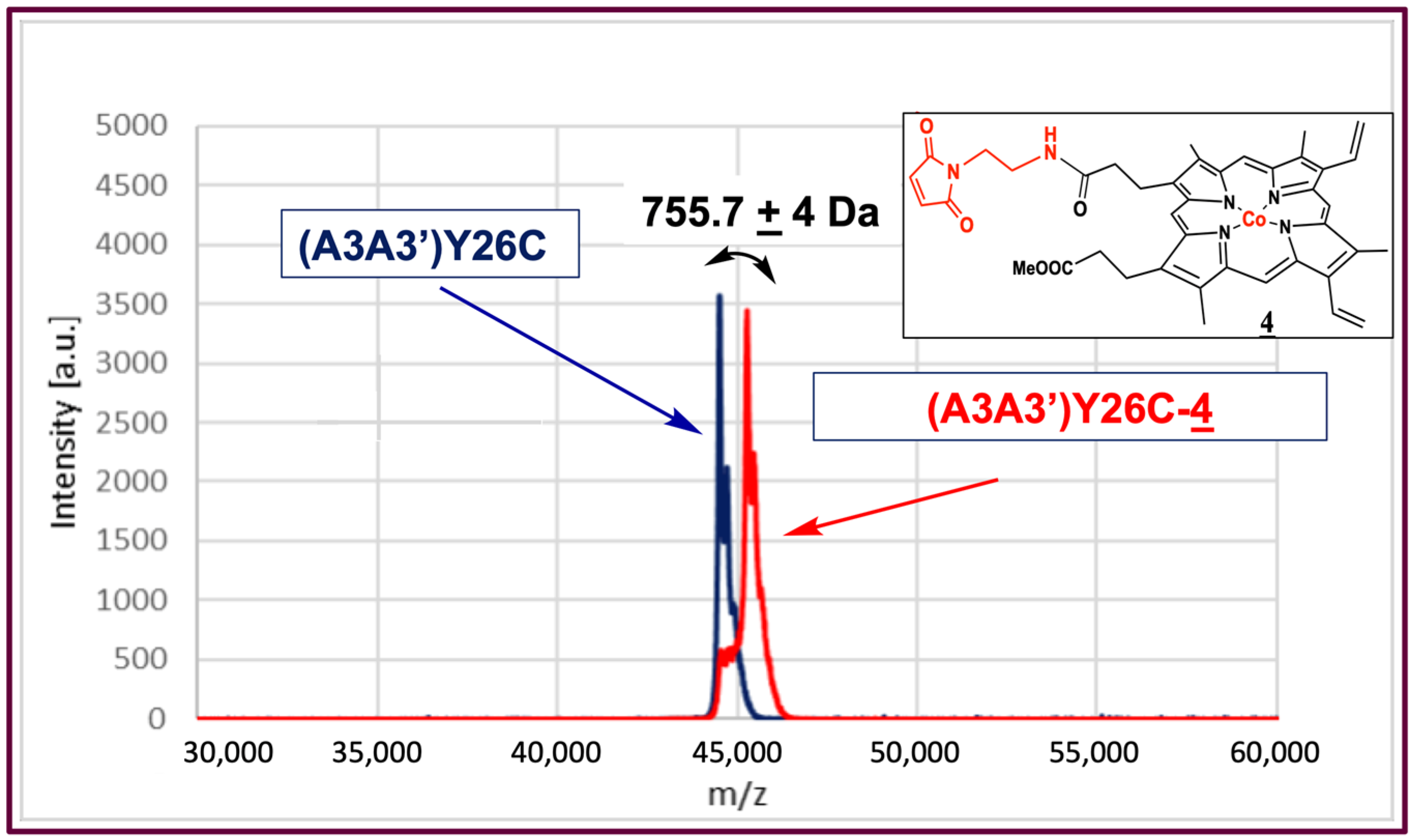
2.2. Biohybrid Preparation
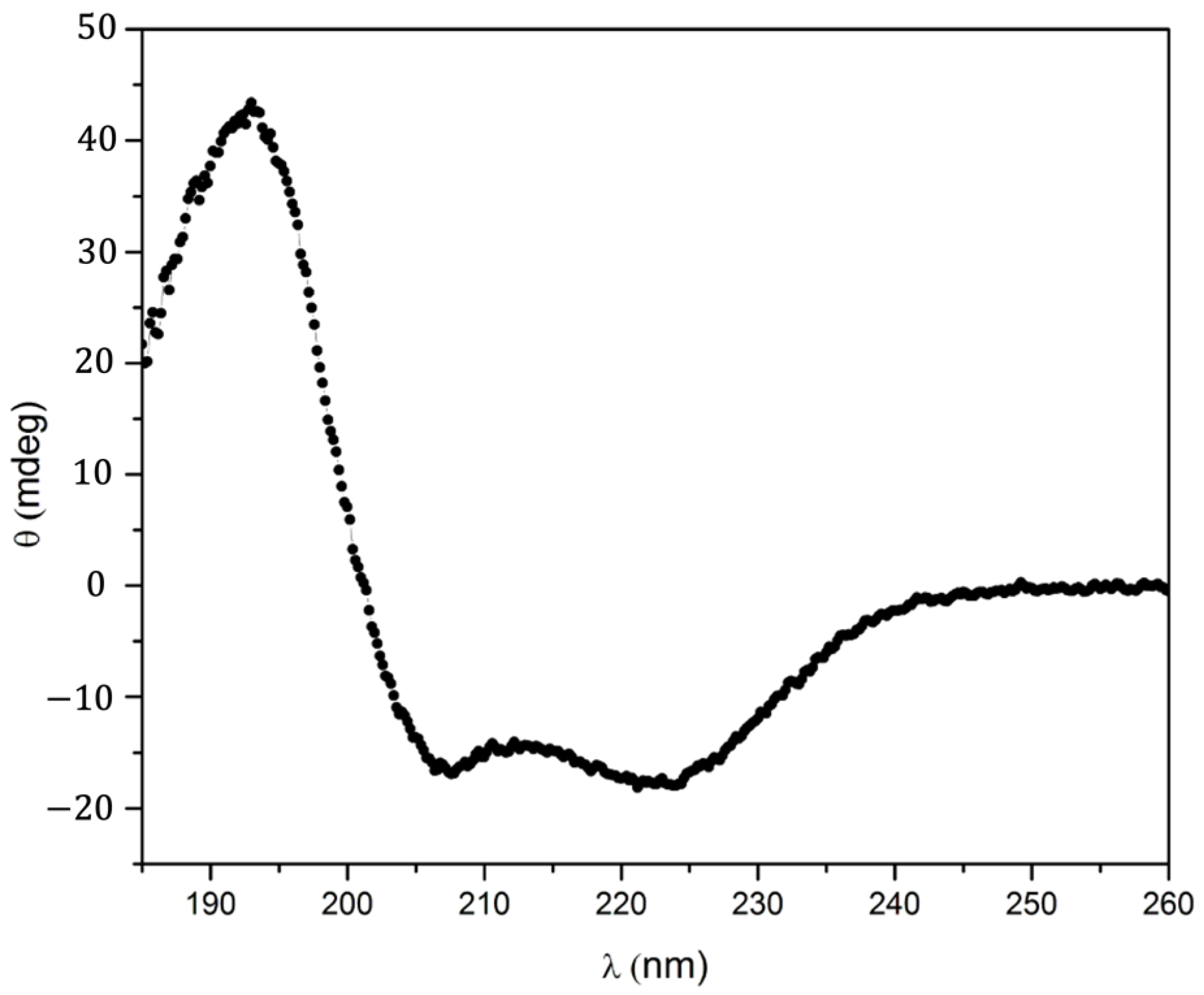
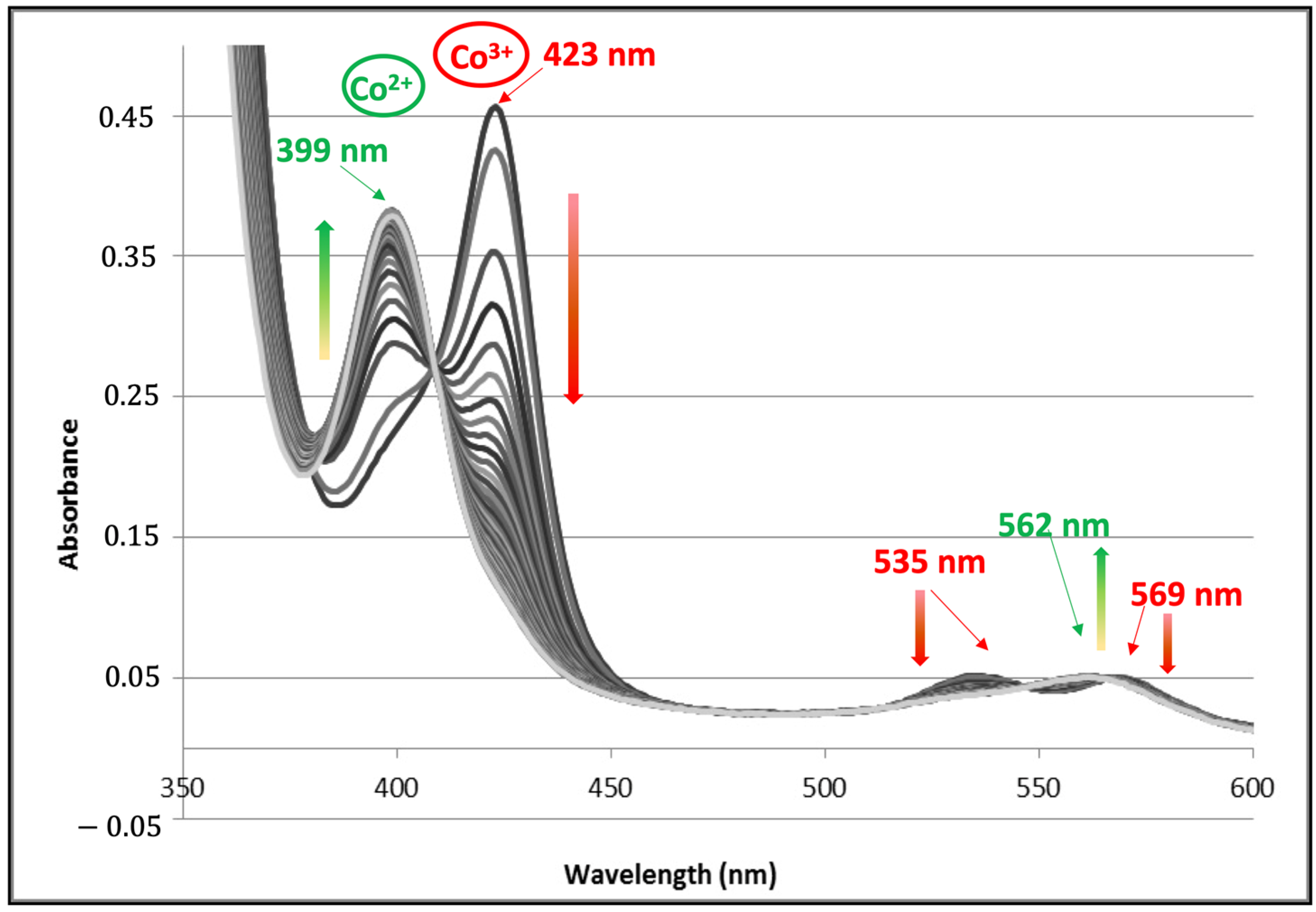
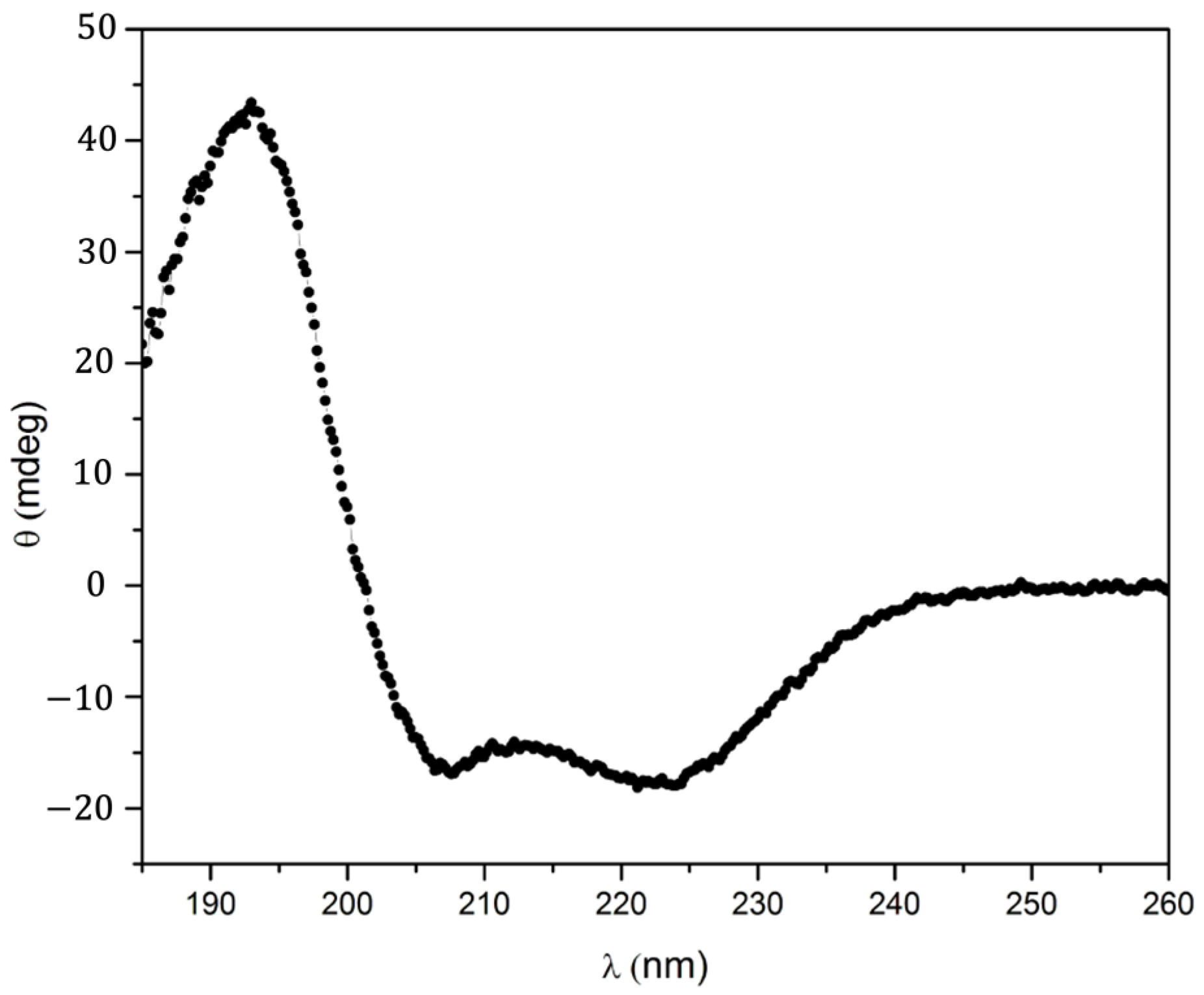
2.3. UV–Visible Characteristics of the (A3A3′)Y26C-4 Biohybrid
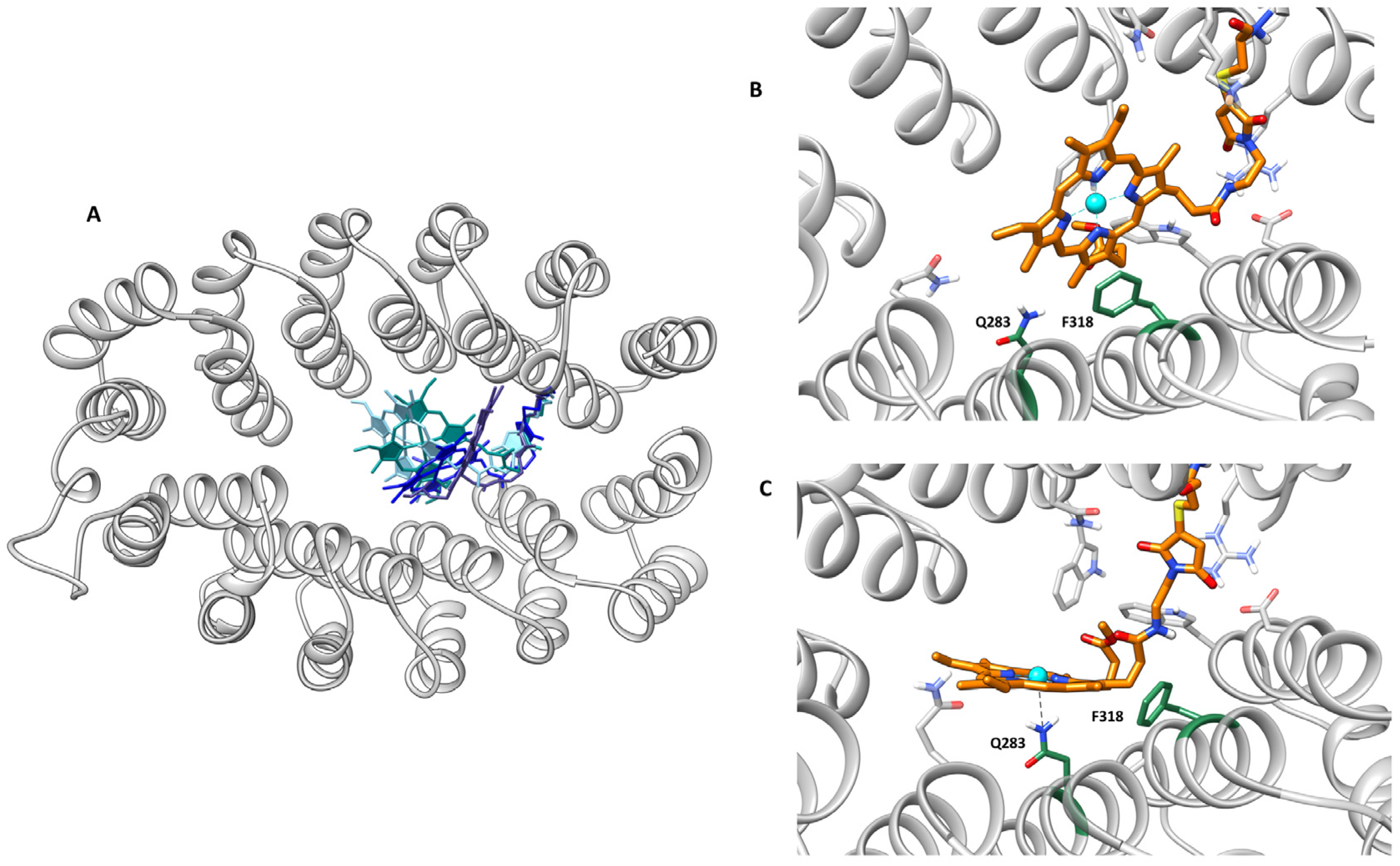
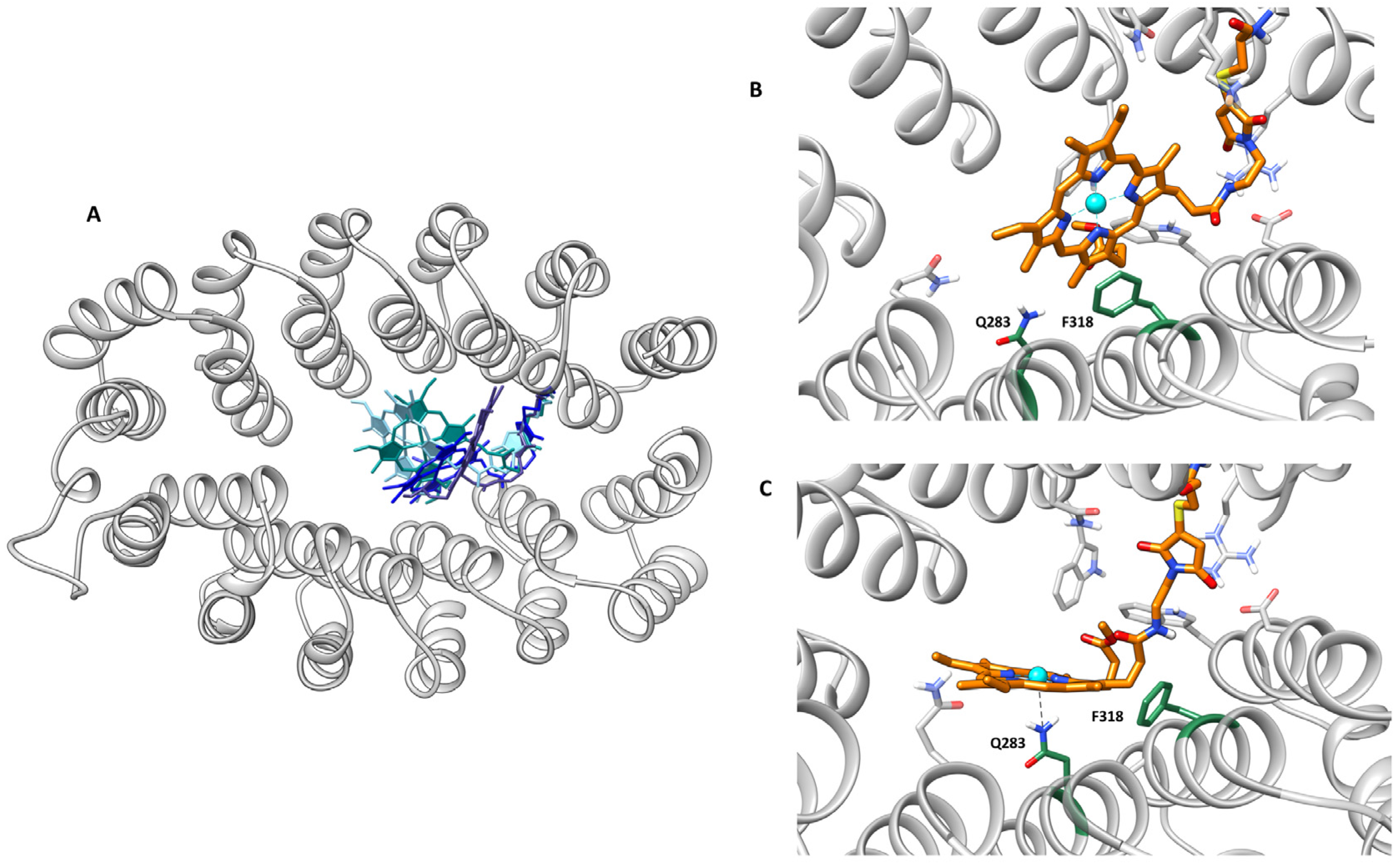
2.4. Computational Modeling Studies
2.5. Photocatalytic Activity of the (A3A3′)Y26C-4 Biohybrid
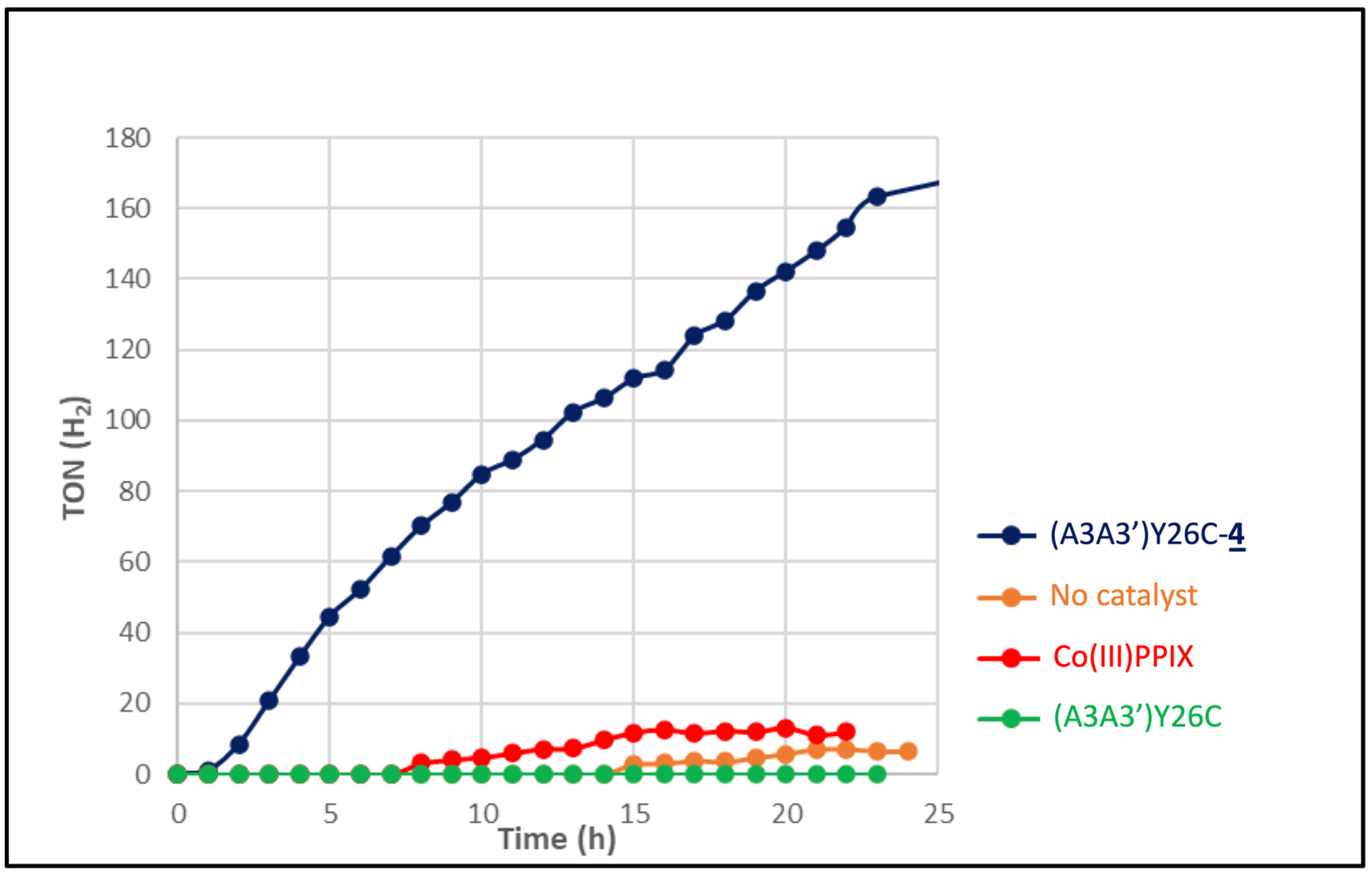
2.5.1. Light-Driven H+ Reduction
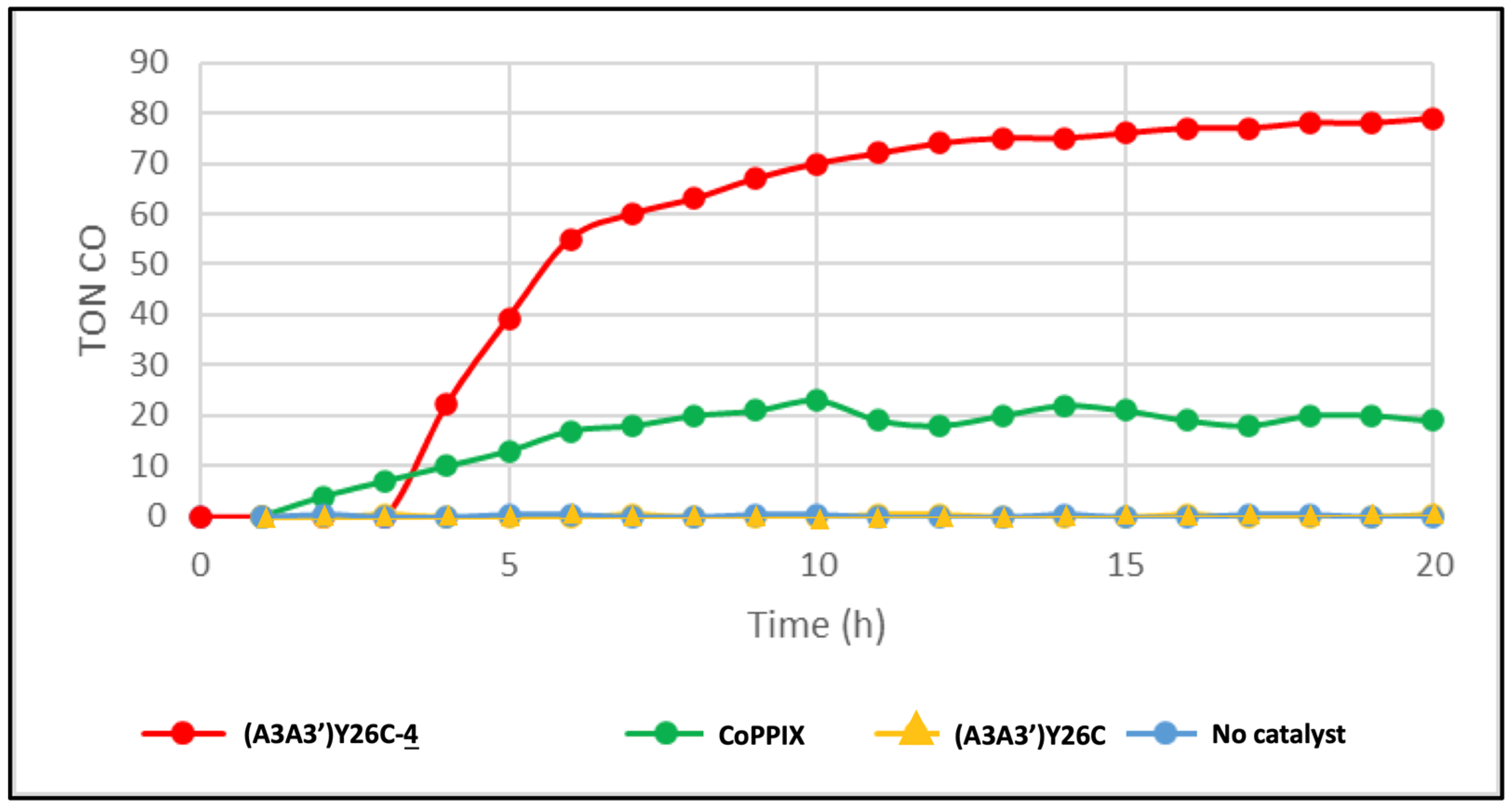
2.5.2. Light-Driven CO2 Reduction
3. Materials and Methods
3.1. Synthesis of Co(III)Mal-PPIX 4
3.1.1. Protoporphyrin IX Dimethyl Ester (PPIX DME) Hydrolysis [23]
3.1.2. Synthesis of Maleimido-Ethylamido-Protoporphyrin IX Monomethyl Ester Mal-PPIX MME 3
3.1.3. Insertion of Cobalt into Maleimido-Ethylamido-Protoporphyrin IX Monomethyl Ester Mal-PPIX MME
3.2. Production and Purification of αRep Proteins
3.2.1. Protein Production
3.2.2. Protein Purification
3.3. Covalent Anchoring of Compound 4 into (A3A3′)Y26C
3.4. Photocatalysis Experiments
3.5. Spectroscopic Characterization of the (A3A3′)Y26C-4 Biohybrid
3.6. Spectroscopic Characterization of the (A3A3′)Y26C-4 Biohybrid
3.7. Computational Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zee, D.Z.; Chantarojsiri, T.; Long, J.R.; Chang, C.J. Metal-polypyridyl catalysts for electro- and photochemical reduction of water to hydrogen. Acc. Chem. Res. 2015, 48, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Landrou, G.; Panagiotopoulos, A.A.; Ladomenou, K.; Coutsolelos, A.G. Photochemical hydrogen evolution using Sn-porphyrin as photosensitizer and a series of Cobaloximes as catalysts. J. Porphyr. Phthalocyanine 2016, 20, 534–541. [Google Scholar] [CrossRef]
- Rao, H.; Wang, Z.Y.; Zheng, H.Q.; Wang, X.B.; Pan, C.M.; Fan, Y.T.; Hou, H.Y. Photocatalytic hydrogen evolution from a cobalt/nickel complex with dithiolene ligands under irradiation with visible light. Catal. Sci. Technol. 2015, 5, 2332–2339. [Google Scholar] [CrossRef]
- Rao, H.; Yu, W.Q.; Zheng, H.Q.; Bonin, J.; Fan, Y.T.; Hou, H.Y. Highly efficient photocatalytic hydrogen evolution from nickel quinolinethiolate complexes under visible light irradiation. J. Power Sources 2016, 324, 253–260. [Google Scholar] [CrossRef]
- Dempsey, J.L.; Brunschwig, B.S.; Winkler, J.R.; Gray, H.B. Hydrogen evolution catalyzed by cobaloximes. Acc. Chem. Res. 2009, 42, 1995–2004. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Duan, L.; Zhou, A.; Thummel, R.P. First-row transition metal polypyridine complexes that catalyze proton to hydrogen reduction. Coord. Chem. Rev. 2020, 402, 2–20. [Google Scholar] [CrossRef]
- Wang, P.; Liang, G.; Reddy, M.R.; Long, M.; Driskill, K.; Lyons, C.; Donnadieu, B.; Bollinger, J.C.; Webster, C.E.; Zhao, X. Electronic and Steric Tuning of Catalytic H2 Evolution by Cobalt Complexes with Pentadentate Polypyridyl-Amine Ligands. J. Am. Chem. Soc. 2018, 140, 9219–9229. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Han, Y.; Lei, H.; Chen, M.; Cao, R. Cobalt corroles with phosphonic acid pendants as catalysts for oxygen and hydrogen evolution from neutral aqueous solution. Chem. Commun. 2017, 53, 6195–6198. [Google Scholar] [CrossRef] [PubMed]
- Natali, M.; Luisa, A.; Iengo, E.; Scandola, F. Efficient photocatalytic hydrogen generation from water by a cationic cobalt(II) porphyrin. Chem. Commun. 2014, 50, 1842–1844. [Google Scholar] [CrossRef] [PubMed]
- Beyene, B.B.; Mane, S.B.; Hung, C.H. Highly efficient electrocatalytic hydrogen evolution from neutral aqueous solution by a water-soluble anionic cobalt(ii) porphyrin. Chem. Commun. 2015, 51, 15067–15070. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.H.; Jelušić, J.; Chakraborty, S.; Bren, K.L. Photochemical hydrogen evolution from cobalt microperoxidase-11. J. Inorg. Biochem. 2021, 217, 111384. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.H.; Le, J.M.; Salamatian, A.A.; Peluso, N.L.; Leone, L.; Lombardi, A.; Bren, K.L. A cobalt mimochrome for photochemical hydrogen evolution from neutral water. J. Inorg. Biochem. 2022, 230, 111753. [Google Scholar] [CrossRef] [PubMed]
- Onoda, A.; Hayashi, T. Artificial hydrogenase: Biomimetic approaches controlling active molecular catalysts. Curr. Opin. Chem. Biol. 2015, 25, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Bacchi, M.; Berggren, G.; Niklas, J.; Veinberg, E.; Mara, M.W.; Shelby, M.L.; Poluektov, O.G.; Chen, L.X.; Tiede, D.M.; Cavazza, C.; et al. Cobaloxime-based artificial hydrogenases. Inorg. Chem. 2014, 53, 8071–8082. [Google Scholar] [CrossRef] [PubMed]
- Bacchi, M.; Veinberg, E.; Field, M.J.; Niklas, J.; Matsui, T.; Tiede, D.M.; Poluektov, O.G.; Ikeda-Saito, M.; Fontecave, M.; Artero, V. Artificial Hydrogenases Based on Cobaloximes and Heme Oxygenase. ChemPlusChem 2016, 81, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Soltau, S.R.; Niklas, J.; Dahlberg, P.D.; Poluektov, O.G.; Tiede, D.M.; Mulfort, K.L.; Utschig, L.M. Aqueous light driven hydrogen production by a Ru-ferredoxin-Co biohybrid. Chem. Commun. 2015, 51, 10628–10631. [Google Scholar] [CrossRef]
- Sommer, D.J.; Vaughn, M.D.; Ghirlanda, G. Protein secondary-shell interactions enhance the photoinduced hydrogen production of cobalt protoporphyrin IX. Chem. Commun. 2014, 50, 15852–15855. [Google Scholar] [CrossRef]
- Alcala-Torano, R.; Halloran, N.; Gwerder, N.; Sommer, D.J.; Ghirlanda, G. Light-Driven CO2 Reduction by Co-Cytochrome b 562. Front. Mol. Biosci. 2021, 8, 609–654. [Google Scholar] [CrossRef]
- Di Meo, T.; Ghattas, W.; Herrero, C.; Velours, C.; Minard, P.; Mahy, J.P.; Ricoux, R.; Urvoas, A. αRep A3: A Versatile Artificial Scaffold for Metalloenzyme Design. Chem. Eur. J. 2017, 23, 10156–10166. [Google Scholar] [CrossRef]
- Di Meo, T.; Kariyawasam, K.; Ghattas, W.; Valerio-Lepiniec, M.; Sciortino, G.; Maréchal, J.-D.; Minard, P.; Mahy, J.P.; Urvoas, A.; Ricoux, R. Functionalized Artificial Bidomain Proteins Based on an α-Solenoid Protein Repeat Scaffold: A New Class of Artificial Diels–Alderases. ACS Omega 2019, 4, 4437–4447. [Google Scholar] [CrossRef]
- Kariyawasam, K.; Di Meo, T.; Hammerer, F.; Valerio-Lepiniec, M.; Sciortino, G.; Maréchal, J.D.; Minard, P.; Mahy, J.P.; Urvoas, A.; Ricoux, R. An Artificial Hemoprotein with Inducible Peroxidase- and Monooxygenase-Like Activities. Chem. Eur. J. 2020, 26, 14929–14937. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, R.K. Chromatographic separation of milligram quantities of protoporphyrin IX monomethyl ester from protoporphyrin IX and its dimethyl ester. Anal. Biochem. 1969, 3, 377–380. [Google Scholar] [CrossRef]
- Koshiyama, T.; Yokoi, N.; Ueno, T.; Kanamaru, S.; Nagano, S.; Shiro, Y.; Arisaka, F.; Watanabe, Y. Molecular Design of HeteroproteinAssemblies Providing a Bionanocup as a Chemical Reactor. Small 2008, 4, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Léger, C.; Di Meo, T.; Aumont-Nicaise, M.; Velours, C.; Durand, D.; Li de la Sierra-Gallay, I.; van Tilbeurgh, H.; Hildebrandt, N.; Desmadril, M.; Urvoas, A.; et al. Ligand-Induced Conformational Switch in an Artificial Bidomain Protein Scaffold. Sci. Rep. 2019, 9, 1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wen, X.; Cui, X.; Zhang, X.P. Enantioselective radical cyclization for construction of 5-membered ring structures by metalloradical C-H alkylation. J. Am. Chem. Soc. 2018, 140, 4792–4796. [Google Scholar] [CrossRef]
- Di Bilio, A.J.; Chang Bo, T.K.; Malmström, G.; Gray, H.B.; Göran Karlsson, B.; Nordling, M.; Pascher, T.; Lundberg, L.G. Electronic absorption spectra of M(II)(Met121X) azurins (MCo, Ni, Cu; XLeu, Gly, Asp, Glu): Charge-transfer energies and reduction potentials. Inorg. Chim. Acta 1992, 198–200, 145–148. [Google Scholar] [CrossRef]
- Lu, H.; Dzik, W.I.; Xu, X.; Wojtas, L.; de Bruin, B.; Zhang, P. Experimental evidence for cobalt(III)-carbene radicals: Key intermediates in cobalt(II)-based metalloradical cyclopropanation. J. Am. Chem. Soc. 2011, 133, 8518–8521. [Google Scholar] [CrossRef]
- Perkins, L.J.; Weaver, B.R.; Buller, A.R.; Burstyn, J.N. De novo biosynthesis of a nonnatural cobalt porphyrin cofactor in E. coli and incorporation into hemoproteins. Proc. Natl. Acad. Sci. USA 2021, 118, e2017625118. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E.I.; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. AMBER 2020; University of California: San Francisco, CA, USA, 2020. [Google Scholar]
- Shapovalov, M.V.; Dunbrack, R.L., Jr. A Smoothed Backbone-Dependent Rotamer Library for Proteins Derived from Adaptive Kernel Density Estimates and Regressions. Structure 2011, 19, 844–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, A.W.; Böhme, M.; Dapprich, S.; Gobbi, A.; Höllwarth, V.; Jonas, A.; Köhler, K.F.; Stegmann, R.; Veldkamp, A.; Frenking, G. A set of f-polarization functions for pseudo-potential basis sets of the transition metals Sc/Cu, Y/Ag and La/Au. G. Chem. Phys. Lett. 1993, 208, 111–114. [Google Scholar] [CrossRef]
- Li, P.; Merz, K.M., Jr. MCPB.py: A Python Based Metal Center Parameter Builder. J. Chem. Inf. Model. 2016, 56, 599–604. [Google Scholar] [CrossRef]
- Chen, J.; Wang, J.; Zhu, W. Zinc ion-induced conformational changes in new Delphi metallo-β-lactamase 1 probed by molecular dynamics simulations and umbrella sampling. Phys. Chem. Chem. Phys. 2017, 19, 3067–3075. [Google Scholar] [CrossRef] [PubMed]
- Teets, T.S.; Nocera, D.G. Photocatalytic hydrogen production. Chem. Commun. 2011, 47, 9268–9274. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.J.; Meyer, G.J.; Fujita, E. Molecular Approaches to the Photocatalytic Reduction of Carbon Dioxide for Solar Fuels. Acc. Chem. Res. 2009, 42, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Marianov, A.N.; Kochubei, A.S.; Roman, T.; Conquest, O.J.; Stampfl, C.; Jiang, Y. Resolving Deactivation Pathways of Co Porphyrin-Based Electrocatalysts for CO2 Reduction in Aqueous Medium. ACS. Catal. 2021, 11, 3715–3729. [Google Scholar] [CrossRef]
- Doukov, T.I.; Blasiak, L.C.; Seravalli, J.; Ragsdale, S.W.; Drennan, C.L. Xenon in and at the End of the Tunnel of Bifunctional Carbon Monoxide Dehydrogenase/Acetyl-CoA Synthase. Biochemistry 2008, 47, 3474–3483. [Google Scholar] [CrossRef] [Green Version]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision C.01; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal Solvation Model Based on Solute Electron Density and on a Continuum Model of the Solvent Defined by the Bulk Dielectric Constant and Atomic Surface Tensions. J. Phys. Chem. B 2009, 113, 6378–6396. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, G.; Garribba, E.; Maréchal, J.-D. Validation and Applications of Protein–Ligand Docking Approaches Improved for Metalloligands with Multiple Vacant Sites. Inorg. Chem. 2019, 58, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Cornell, W.D.; Cieplak, P.; Bayly, C.I.; Gould, I.R.; Merz, K.M.; Ferguson, D.M.; Spellmeyer, D.C.; Fox, T.; Caldwell, J.W.; Kollman, P.A. A Second-Generation Force Field for the Simulation of Proteins, Nucleic Acids, and Organic Molecules. J. Am. Chem. Soc. 1995, 117, 5179–5197. [Google Scholar] [CrossRef] [Green Version]
- Seminario, J.M. Calculation of intramolecular force fields from second-derivative tensors. Int. J. Quant. Chem. 1996, 60, 1271–1277. [Google Scholar] [CrossRef]
- Bayly, C.I.; Cieplak, P.; Cornell, W.; Kollman, P.A. A well-behaved electrostatic potential based method using charge restraints for deriving atomic charges: The RESP model. J. Phys. Chem. 1993, 97, 10269–10280. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]


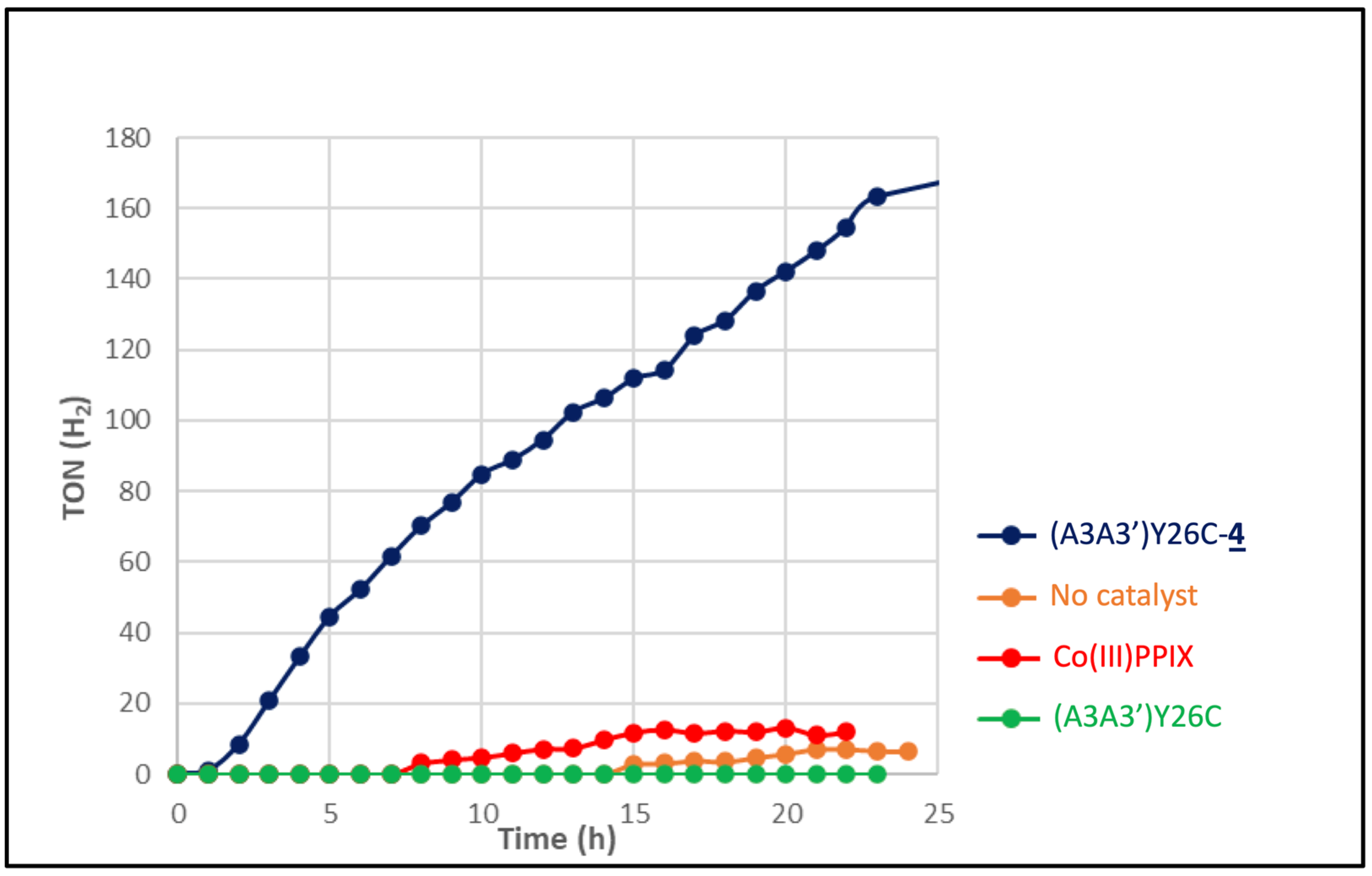
 ) as a catalyst or without catalyst (•) under white LED light, λ > 405 nm irradiation. The experiments were carried out in duplicate.
) as a catalyst or without catalyst (•) under white LED light, λ > 405 nm irradiation. The experiments were carried out in duplicate.
) as a catalyst or without catalyst (•) under white LED light, λ > 405 nm irradiation. The experiments were carried out in duplicate.
) as a catalyst or without catalyst (•) under white LED light, λ > 405 nm irradiation. The experiments were carried out in duplicate.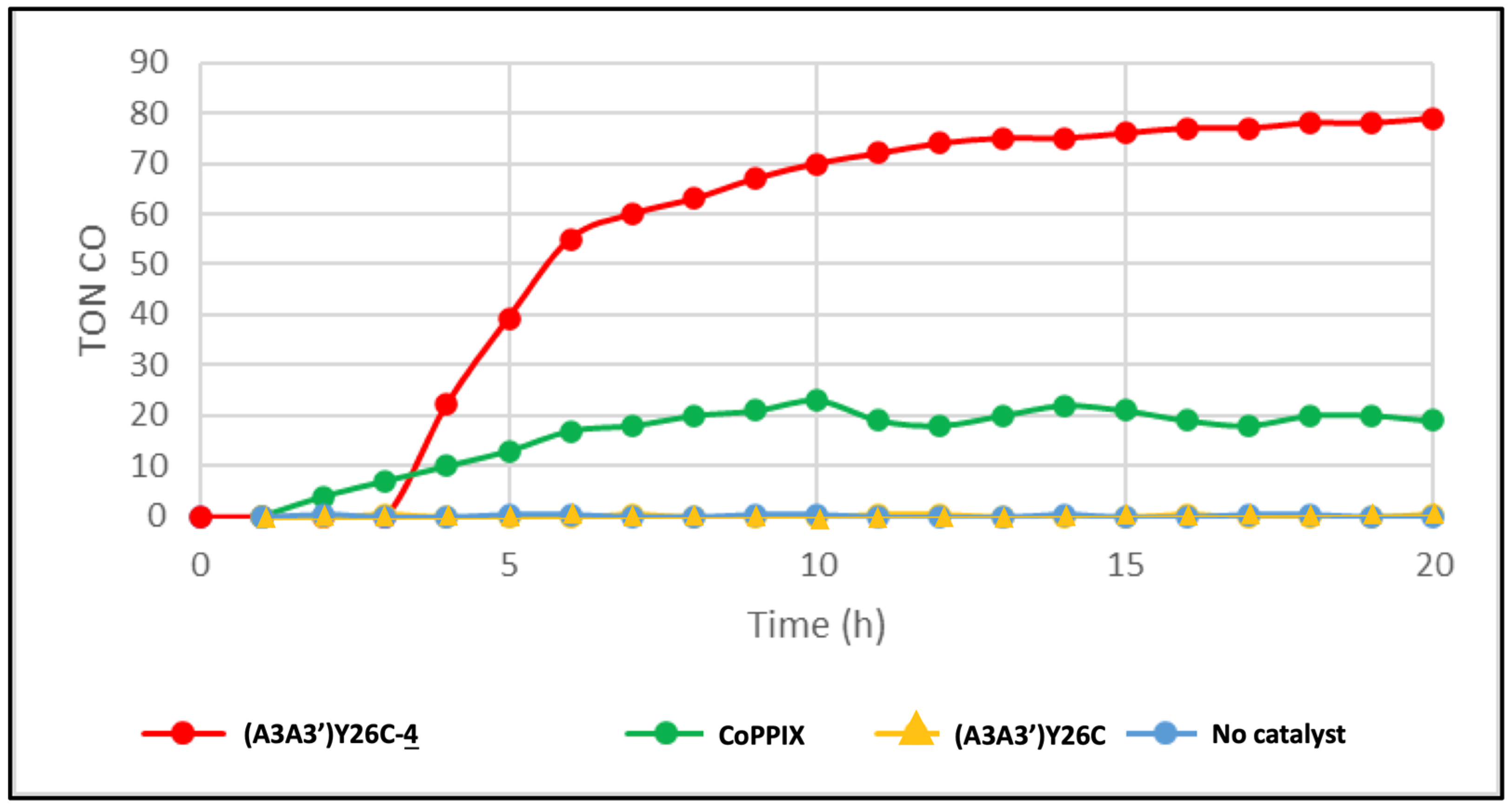

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Co(III) λMax (nm) | Co(II) λMax (nm) | |||||||
|---|---|---|---|---|---|---|---|---|
| Protein | Ligands | Soret | β | α | Ligand | Soret | α/β | Ref. |
| Mb | His/H2O | 424 | 535 | 572 | His | 406 | 558 | [24] |
| MbH64VV68A | His/H2O | 428 | 540 | 572 | His | 406 | 555 | [25] |
| P450Cam | Cys/H2O | 422 | 538 | 570 | Cys | 404 | 556 | [26] |
| CYP119 | Cys/H2O | 422 | 539 | 566 | Cys | 410 | 558 | [27] |
| Oxd | His/H2O | 429 | 542 | 574 | His | 402 | 559 | [28] |
| HRP | His/H2O | 421 | 533 | 565 | His | 401 | 553 | [24] |
| DYP | His/H2O | 422 | 534 | 567 | His | 403 | 564 | [28] |
| Cat | His/? | 427 | 537 | 570 | His | 403 | 561 | [28] |
| (A3A3′)Y26C-4 | ? | 423 | 535 | 569 | ? | 399 | 562 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Udry, G.A.O.; Tiessler-Sala, L.; Pugliese, E.; Urvoas, A.; Halime, Z.; Maréchal, J.-D.; Mahy, J.-P.; Ricoux, R. Photocatalytic Hydrogen Production and Carbon Dioxide Reduction Catalyzed by an Artificial Cobalt Hemoprotein. Int. J. Mol. Sci. 2022, 23, 14640. https://doi.org/10.3390/ijms232314640
Udry GAO, Tiessler-Sala L, Pugliese E, Urvoas A, Halime Z, Maréchal J-D, Mahy J-P, Ricoux R. Photocatalytic Hydrogen Production and Carbon Dioxide Reduction Catalyzed by an Artificial Cobalt Hemoprotein. International Journal of Molecular Sciences. 2022; 23(23):14640. https://doi.org/10.3390/ijms232314640
Chicago/Turabian StyleUdry, Guillermo A. Oliveira, Laura Tiessler-Sala, Eva Pugliese, Agathe Urvoas, Zakaria Halime, Jean-Didier Maréchal, Jean-Pierre Mahy, and Rémy Ricoux. 2022. "Photocatalytic Hydrogen Production and Carbon Dioxide Reduction Catalyzed by an Artificial Cobalt Hemoprotein" International Journal of Molecular Sciences 23, no. 23: 14640. https://doi.org/10.3390/ijms232314640
APA StyleUdry, G. A. O., Tiessler-Sala, L., Pugliese, E., Urvoas, A., Halime, Z., Maréchal, J.-D., Mahy, J.-P., & Ricoux, R. (2022). Photocatalytic Hydrogen Production and Carbon Dioxide Reduction Catalyzed by an Artificial Cobalt Hemoprotein. International Journal of Molecular Sciences, 23(23), 14640. https://doi.org/10.3390/ijms232314640