
Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo



Abstract
1. Introduction
2. Results
2.1. Effect of SeNP Injections into Mice on Neutrophil Counts and Their Adhesive Functions
2.2. Dose-Dependent Effect of SeNP Injections on Spontaneous and Activated ROS Production by Mouse Neutrophils
2.3. The Action of SeNP Injections into Mice Regulates the Inflammatory Status of Neutrophils
2.4. Dose-Dependent Effect of SeNPs on the Antimicrobial Activity of Mouse Neutrophils Mediated by NET Generation
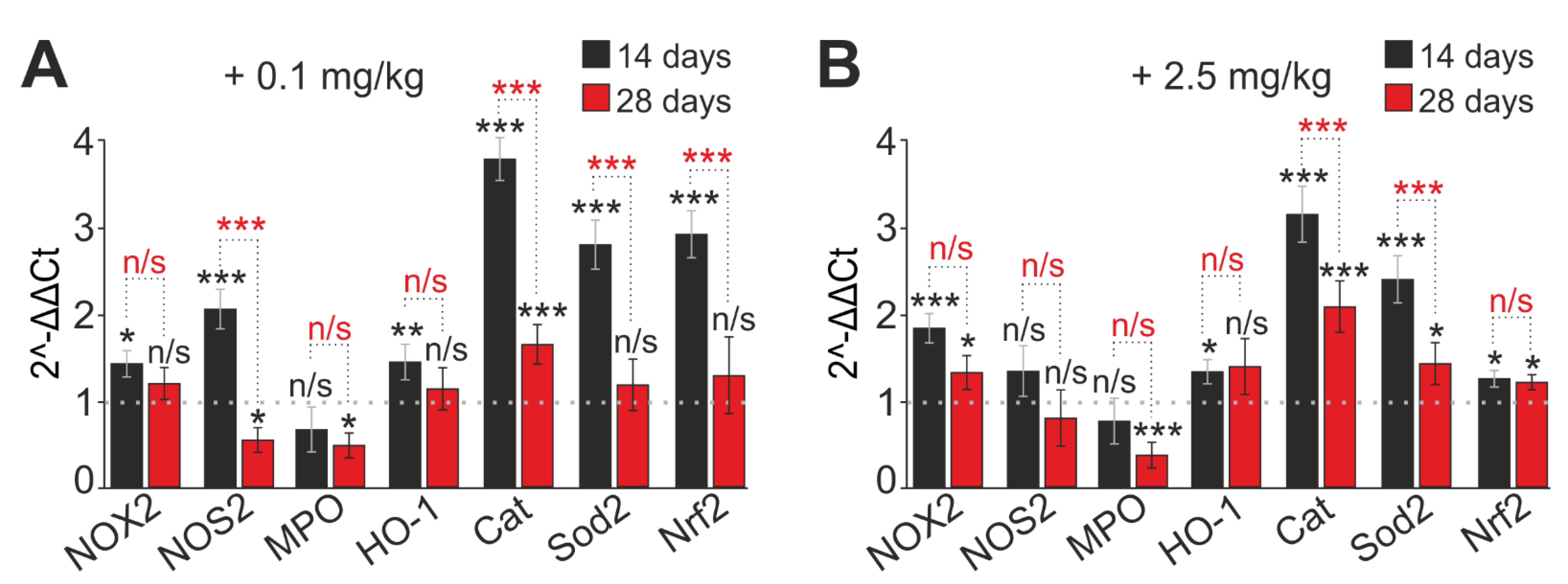
2.5. SeNP Injections into Mice Regulate the Expression of Genes Encoding Selenoproteins
3. Discussion
3.1. Effects of SeNPs and Selenium on the Dynamics of the Adhesive Abilities of Neutrophils
3.2. Effect of SeNPs on ROS Production and Release of Neutrophil Extracellular Traps (NETs)
3.3. Effect of SeNPs on the Inflammatory Status of Mouse
3.4. Effect of SeNP Injections on Selenoprotein Expression in Mouse Neutrophils
4. Materials and Methods
4.1. Experimental Animals
4.2. Preparation and Characterization of Selenium Nanoparticles
4.3. Mouse Model of SeNP Administration
4.4. Isolation of Peritoneal Neutrophils and Estimation of Their Number and Viability
4.5. Adhesion Assay
4.6. Quantification of NETs
4.7. Chemiluminescent Analysis of ROS Generation
4.8. Extraction of RNA and Real-Time Polymerase Chain Reaction (RT-qPCR)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Espinosa, O.; Rojas-Espinosa, O.; Moreno-Altamirano, M.M.; López-Villegas, E.O.; Sánchez-García, F. Metabolic requirements for neutrophil extracellular traps formation. J. Immunol. 2015, 145, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Peiseler, M.; Kubes, P. More friend than foe: The emerging role of neutrophils in tissue repair. J. Clin. Investig. 2019, 129, 2629–2639. [Google Scholar] [CrossRef] [PubMed]
- Pruchniak, M.P.; Arazna, M.; Demkow, U. Life of neutrophil: From stem cell to neutrophil extracellular trap. Respir. Physiol. Neurobiol. 2013, 187, 68–73. [Google Scholar] [CrossRef]
- Lehman, H.K.; Segal, B.H. The role of neutrophils in host defense and disease. J. Allergy. Clin. Immunol. 2020, 145, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Mouillot, P.; Witko-Sarsat, V.; Wislez, M. Neutrophil plasticity: A new key in the understanding of onco-immunology. Rev. Mal. Respir. 2022, 39, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, A.; Popp, S.K.; Fukuda, R.; Parish, C.R.; Bosi, E.; Simeonovic, C.J. The Contribution of Neutrophils and NETs to the Development of Type 1 Diabetes. Front. Immunol. 2022, 13, 930553. [Google Scholar] [CrossRef]
- Masuda, S.; Nakazawa, D.; Shida, H.; Miyoshi, A.; Kusunoki, Y.; Tomaru, U.; Ishizu, A. NETosis markers: Quest for specific, objective, and quantitative markers. Clin. Chim. Acta 2016, 459, 89–93. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- Castanheira, F.V.S.; Kubes, P. Neutrophils and NETs in modulating acute and chronic inflammation. Blood 2019, 133, 2178–2185. [Google Scholar] [CrossRef]
- Liao, C.; Mao, F.; Qian, M.; Wang, X. Pathogen-Derived Nucleases: An Effective Weapon for Escaping Extracellular Traps. Front. Immunol. 2022, 13, 899890. [Google Scholar] [CrossRef]
- Kubes, P. The enigmatic neutrophil: What we do not know. Cell Tissue Res. 2018, 371, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lv, G.; Lian, S.; Wang, J.; Wu, R. Effect of Copper, Zinc, and Selenium on the Migration of Bovine Neutrophils. Vet. Sci. 2021, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Bartneck, M.; Wang, J. Therapeutic Targeting of Neutrophil Granulocytes in Inflammatory Liver Disease. Front. Immunol. 2019, 10, 2257. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Seo, J.; Yang, E.J.; Choi, I.H. Silver Nanoparticles Induce Neutrophil Extracellular Traps Via Activation of PAD and Neutrophil Elastase. Biomolecules 2021, 11, 317. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, X.; Liu, M.; Duan, M.; Zhang, S.; Wei, X.; Liu, X. Toward improved human health: Efficacy of dietary selenium on immunity at the cellular level. Food Funct. 2021, 12, 976–989. [Google Scholar] [CrossRef]
- Zhang, R.R.; Guo, Q.; Liu, G.; Li, B.; Sun, X. Selenium Deficiency via the TLR4/TRIF/NF-kappa B Signaling Pathway Leading to Inflammatory Injury in Chicken Spleen. Biol. Trace Elem. Res. 2021, 199, 693–702. [Google Scholar] [CrossRef]
- Qian, F.; Misra, S.; Prabhu, K.S. Selenium and selenoproteins in prostanoid metabolism and immunity. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 484–516. [Google Scholar] [CrossRef]
- Elmadfa, I.; Meyer, A.L. The Role of the Status of Selected Micronutrients in Shaping the Immune Function. Endocr. Metab. Immune Disord. Drug. Targets 2019, 19, 1100–1115. [Google Scholar] [CrossRef]
- Minetti, G. Mevalonate pathway, selenoproteins, redox balance, immune system, Covid-19: Reasoning about connections. Med. Hypotheses 2020, 144, 110128. [Google Scholar] [CrossRef]
- Xia, H.; Zhang, L.; Dai, J.; Liu, X.; Zhang, X.; Zeng, Z.; Jia, Y. Effect of Selenium and Peroxynitrite on Immune Function of Immature Dendritic Cells in Humans. Med. Sci. Monitor. 2021, 27, 929004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xia, H.; Xia, K.; Liu, X.; Zhang, X.; Dai, J.; Zeng, Z.; Jia, Y. Selenium Regulation of the Immune Function of Dendritic Cells in Mice Through the ERK, Akt and RhoA/ROCK Pathways. Biol. Trace Elem. Res. 2021, 199, 3360–3370. [Google Scholar] [CrossRef]
- Huang, L.J.; Mao, X.T.; Li, Y.Y.; Liu, D.D.; Fan, K.Q.; Liu, R.B.; Wu, T.T.; Wang, H.L.; Zhang, Y.; Yang, B.; et al. Multiomics analyses reveal a critical role of selenium in controlling T cell differentiation in Crohn's disease. Immunity 2021, 54, 1728–1744. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, M.H.; Masoudifar, M.; Varastehmoradi, B.; Mohammadi, E.; Kheradmand, E.; Homayouni, S.; Shahverdi, A.R. Effect of Oral Supplementation of Biogenic Selenium Nanoparticles on White Blood Cell Profile of BALB/c Mice and Mice Exposed to X-ray Radiation. Avicenna J. Med. Biotechnol. 2013, 5, 158–167. [Google Scholar] [PubMed]
- Shao, C.; Zheng, M.; Yu, Z.; Jiang, S.; Zhou, B.; Song, Q.; Ma, T.; Zhou, Y.; Dong, W.; Li, D.; et al. Supplemental Dietary Selenohomolanthionine Improve Antioxidant Activity and Immune Function in Weaned Beagle Puppies. Front. Vet. Sci. 2021, 8, 728358. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox. Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef]
- Hugejiletu, H.; Bobe, G.; Vorachek, W.R.; Gorman, M.E.; Mosher, W.D.; Pirelli, G.J.; Hall, J.A. Selenium supplementation alters gene expression profiles associated with innate immunity in whole-blood neutrophils of sheep. Biol. Trace Elem. Res. 2013, 154, 28–44. [Google Scholar] [CrossRef]
- Zhu, H.; Yu, Q.; Ouyang, H.; Zhang, R.; Li, J.; Xian, R.; Wang, K.; Li, X.; Cao, C. Antagonistic Effect of Selenium on Fumonisin B1 Promotes Neutrophil Extracellular Traps Formation in Chicken Neutrophils. J. Agric. Food Chem. 2022, 70, 5911–5920. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, H.; Lian, S.; Wang, J.; Wu, R. Effect of Copper, Zinc, and Selenium on the Formation of Bovine Neutrophil Extracellular Traps. Biol. Trace Elem. Res. 2021, 199, 3312–3318. [Google Scholar] [CrossRef]
- Skalickova, S.; Milosavljevic, V.; Cihalova, K.; Horky, P.; Richtera, L.; Adam, V. Selenium nanoparticles as a nutritional supplement. Nutrition 2017, 33, 83–90. [Google Scholar] [CrossRef]
- Wadhwani, S.A.; Shedbalkar, U.U.; Singh, R.; Chopade, B.A. Biogenic selenium nanoparticles: Current status and future prospects. Appl. Microbiol. Biotechnol. 2016, 100, 2555–2566. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Turovsky, E.A.; Blinova, E.V. Therapeutic Potential and Main Methods of Obtaining Selenium Nanoparticles. Int. J. Mol. Sci. 2021, 22, 10808. [Google Scholar] [CrossRef] [PubMed]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J.; et al. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107–2128. [Google Scholar] [CrossRef]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Goltyaev, M.V.; Mal'tseva, V.N.; Turovsky, E.A.; Sarimov, R.M.; Simakin, A.V.; Gudkov, S.V. Mechanisms of the Cytotoxic Effect of Selenium Nanoparticles in Different Human Cancer Cell Lines. Int. J. Mol. Sci. 2021, 22, 7798. [Google Scholar] [CrossRef]
- Turovsky, E.A.; Mal'tseva, V.N.; Sarimov, R.M.; Simakin, A.V.; Gudkov, S.V.; Plotnikov, E.Y. Features of the cytoprotective effect of selenium nanoparticles on primary cortical neurons and astrocytes during oxygen-glucose deprivation and reoxygenation. Sci. Rep. 2022, 12, 1710. [Google Scholar] [CrossRef]
- Maiyo, F.; Singh, M. Selenium nanoparticles: Potential in cancer gene and drug delivery. Nanomedicine 2017, 12, 1075–1089. [Google Scholar] [CrossRef]
- Guan, B.; Yan, R.; Li, R.; Zhang, X. Selenium as a pleiotropic agent for medical discovery and drug delivery. Int. J. Nanomedicine 2018, 13, 7473–7490. [Google Scholar] [CrossRef]
- Mudgal, V.; Garg, A.K.; Dass, R.S.; Rawat, M. Selenium and copper interaction at supra-nutritional level affecting blood parameters including immune response against P. multocida antigen in Murrah buffalo (Bubalus bubalis) calves. J. Trace Elem. Med. Biol. 2018, 50, 415–423. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Gudkov, S.V.; Plotnikov, E.Y.; Turovsky, E.A. Size-Dependent Cytoprotective Effects of Selenium Nanoparticles during Oxygen-Glucose Deprivation in Brain Cortical Cells. Int. J. Mol. Sci. 2022, 23, 7464. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhang, J.; Xu, J.F.; Pi, J. The Advancing of Selenium Nanoparticles Against Infectious Diseases. Front. Pharmacol. 2021, 12, 682284. [Google Scholar] [CrossRef] [PubMed]
- Mal'tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. [Google Scholar] [CrossRef] [PubMed]
- Javdani, M.; Habibi, A.; Shirian, S.; Kojouri, G.A.; Hosseini, F. Effect of Selenium Nanoparticle Supplementation on Tissue Inflammation, Blood Cell Count, and IGF-1 Levels in Spinal Cord Injury-Induced Rats. Biol. Trace Elem. Res. 2019, 187, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ren, Z.; Bo, R.; Liu, X.; Zhang, J.; Yu, R.; Chen, S.; Meng, Z.; Xu, Y.; Ma, Y.; et al. Designing selenium polysaccharides-based nanoparticles to improve immune activity of Hericiumerinaceus. Int. J. Biol. Macromol. 2020, 143, 393–400. [Google Scholar] [CrossRef]
- Yehia, N.; Abdel Sabour, M.A.; Erfan, A.M.; Mohammed Ali, Z.; Soliman, R.A.; Samy, A.; Mohamed Soliman, M.; Abd El-Hack, M.E.; El-Saadony, M.T.; Ahmed, K.A. Saudi. Selenium nanoparticles enhance the efficacy of homologous vaccine against the highly pathogenic avian influenza H5N1 virus in chickens. J. Biol. Sci. 2022, 29, 2095–2111. [Google Scholar]
- Pi, J.; Shen, L.; Yang, E.; Shen, H.; Huang, D.; Wang, R.; Hu, C.; Jin, H.; Cai, H.; Cai, J.; et al. Macrophage-Targeted Isoniazid-Selenium Nanoparticles Promote Antimicrobial Immunity and Synergize Bactericidal Destruction of Tuberculosis Bacilli. Angew Chem. Int. Ed. Engl. 2020, 59, 3226–3234. [Google Scholar] [CrossRef]
- Gautam, P.K.; Kumar, S.; Tomar, M.S.; Singh, R.K.; Acharya, A.; Kumar, S.; Ram, B. Selenium nanoparticles induce suppressed function of tumor associated macrophages and inhibit Dalton's lymphoma proliferation. Biochem. Biophys. Rep. 2017, 12, 172–184. [Google Scholar] [CrossRef]
- Dung, N.T.; Trong, T.D.; Vu, N.T.; Binh, N.T.; Minh, T.T.L.; Luan, L.Q. Radiation Synthesis of Selenium Nanoparticles Capped with β-Glucan and Its Immunostimulant Activity in Cytoxan-Induced Immunosuppressed Mice. Nanomaterials 2021, 11, 2439. [Google Scholar] [CrossRef]
- Machholz, E.; Mulder, G.; Ruiz, C.; Corning, B.F. Pritchett-Corning KR. Manual restraint and common compound administration routes in mice and rats. J. Vis. Exp. 2012, 26, 2771. [Google Scholar]
- Rybakova, A.V.; Makarova, M.N.; Kukharenko, A.E.; Vichare, A.S. Current Requirements for and Approaches to Dosing in Animal Studies. Bull. Sci. Cent. Expert Eval. Med. Prod. 2018, 8, 207–217. [Google Scholar] [CrossRef]
- Lawrence, S.M.; Corriden, R.; Nizet, V. How Neutrophils Meet Their End. Trends Immunol. 2020, 41, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Pillay, J.; Ramakers, B.P.; Kamp, V.M.; Loi, A.L.; Lam, S.W.; Hietbrink, F.; Leenen, L.P.; Tool, A.T.; Pickkers, P.; Koenderman, L. Functional heterogeneity and differential priming of circulating neutrophils in human experimental endotoxemia. J. Leukoc. Biol. 2010, 88, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.A.; Huo, Y.; Burcin, T.L.; Morris, M.A.; Olson, T.S.; Ley, K. Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity 2005, 22, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, X.; Xu, S. Selenium Prevents Lead-Induced Necroptosis by Restoring Antioxidant Functions and Blocking MAPK/NF-κB Pathway in Chicken Lymphocytes. Biol. Trace Elem. Res. 2020, 198, 644–653. [Google Scholar] [CrossRef]
- Kojouri, G.A.; Sadeghian, S.; Mohebbi, A.; MokhberDezfouli, M.R. The effects of oral consumption of selenium nanoparticles on chemotactic and respiratory burst activities of neutrophils in comparison with sodium selenite in sheep. Biol. Trace Elem. Res. 2012, 146, 160–166. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Novella, A.; Bergamaschi, G.; Canale, C.; Fossati, G.; Gritti, D.; Lucotti, C.; Mazzone, A.; Tonon, L.; Cazzola, M. Expression of adhesion molecules and functional stimulation in human neutrophils: Modulation by GM-CSF and role of the Bcr gene. Br. J. Haematol. 1997, 98, 621–626. [Google Scholar] [CrossRef]
- Maddox, J.F.; Aherne, K.M.; Reddy, C.C.; Sordillo, L.M. Increased neutrophil adherence and adhesion molecule mRNA expression in endothelial cells during selenium deficiency. J. Leukoc. Biol. 1999, 65, 658–664. [Google Scholar] [CrossRef]
- Algorri, M.; Wong-Beringer, A. Antibiotics Differentially Modulate Lipoteichoic Acid-Mediated Host Immune Response. Antibiotics 2020, 9, 573. [Google Scholar] [CrossRef]
- Jahnova, E.; Horvathova, M.; Gazdik, F.; Weissova, S. Effects of selenium supplementation on expression of adhesion molecules in corticoid-dependent asthmatics. Bratisl. Lek. Listy. 2002, 103, 12–16. [Google Scholar] [PubMed]
- Zarember, K.A.; Kuhns, D.B. Editorial: Will the real neutrophil please stand up? J. Leukoc. Biol. 2011, 90, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Condliffe, A.M.; Chilvers, E.R.; Haslett, C.; Dransfield, I. Priming differentially regulates neutrophil adhesion molecule expression/function. Immunology 1996, 89, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Turutin, D.V.; Kubareva, E.A.; Pushkareva, M.A.; Ullrich, V.; Sud'ina, G.F. Activation of NF-kappa B transcription factor in human neutrophils by sulphatides and L-selectin cross-linking. FEBS Lett. 2003, 536, 241–245. [Google Scholar] [CrossRef]
- Speranskii, A.I.; Kostyuk, S.V.; Kalashnikova, E.A.; Veiko, N.N. Enrichment of extracellular DNA from the cultivation medium of human peripheral blood mononuclears with genomic CpG rich fragments results in increased cell production of IL-6 and TNF-a via activation of the NF-kB signaling pathway. Biomed. Khim. 2016, 62, 331–340. [Google Scholar] [CrossRef][Green Version]
- Miguel, L.I.; Leonardo, F.C.; Torres, L.S.; Garcia, F.; Mendonça, R.; Ferreira, W.A., Jr.; Gotardo, É.M.F.; Fabris, F.C.Z.; Brito, P.L.; Costa, F.F.; et al. Heme induces significant neutrophil adhesion in vitro via an NFκB and reactive oxygen species-dependent pathway. Mol. Cell Biochem. 2021, 476, 3963–3974. [Google Scholar] [CrossRef]
- Kiersnowska-Rogowska, B.; Izycka, A.; Jabłońska, E.; Rogowski, F.; Parfieńczyk, A. Ocena ekspresji L-selektyny na neutrofilach oraz rozpuszczalnej formys L-selektyny w surowicy krwi u chorych na przewlekła białaczke szpikowa. Estimation of L-selectin expression on neutrophils and level of soluble L-selectin form in serum of patient with chronic myelogenic leukemia. Przegl. Lek. 2006, 63, 756–758. [Google Scholar]
- Chuluyan, H.E.; Lang, B.J.; Issekutz, A.C. Differential mechanisms of neutrophil and monocyte adhesion on neuroblastoma cells: CD18 and VLA-4 integrins mediate adhesion to SK-N-SH, but not to SK-N-MC cell line. J. Neurosci. Res. 2000, 60, 649–655. [Google Scholar] [CrossRef]
- Jadhav, S.H.; Sarkar, S.N.; Ram, G.C.; Tripathi, H.C. Immunosuppressive effect of subchronic exposure to a mixture of eight heavy metals, found as groundwater contaminants in different areas of India, through drinking water in male rats. Arch. Environ. Contam. Toxicol. 2007, 53, 450–458. [Google Scholar] [CrossRef]
- El-Benna, J.; Dang, P.M.; Gougerot-Pocidalo, M.A. Priming of the neutrophil NADPH oxidase activation: Role of p47phox phosphorylation and NOX2 mobilization to the plasma membrane. Semin Immunopathol. 2008, 30, 279–289. [Google Scholar] [CrossRef]
- Babior, B.M. The leukocyte NADPH oxidase. Isr. Med. Assoc. J. 2002, 4, 1023–1024. [Google Scholar] [PubMed]
- Saraiva, A.L.; Vieira, T.N.; Notário, A.F.O.; Luiz, J.P.M.; Silva, C.R.; Goulart, L.R.; Dantas, N.O.; Silva, A.C.A.; Espindola, F.S. CdSe magic-sized quantum dots attenuate reactive oxygen species generated by neutrophils and macrophages with implications in experimental arthritis. Nanomedicine 2022, 42, 102539. [Google Scholar] [CrossRef] [PubMed]
- Ibeagha, A.E.; Ibeagha-Awemu, E.M.; Mehrzad, J.; Baurhoo, B.; Kgwatalala, P.; Zhao, X. The effect of selenium sources and supplementation on neutrophil functions in dairy cows. Animal 2009, 3, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Malá, S.; Kovárů, F.; Misurová, L.; Pavlata, L.; Dvorák, R.; Cíz, M. Influence of selenium on innate immune response in kids. Folia Microbiol. 2009, 54, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Forceville, X.; Van Antwerpen, P.; Preiser, J.C. Selenocompounds and Sepsis: Redox Bypass Hypothesis for Early Diagnosis and Treatment: Part A-Early Acute Phase of Sepsis: An Extraordinary Redox Situation (Leukocyte/Endothelium Interaction Leading to Endothelial Damage). Antioxid. Redox Signal. 2021, 35, 113–138. [Google Scholar] [CrossRef] [PubMed]
- Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.D. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 10th ed.; Spain, 2007; pp. 552–557. [Google Scholar]
- Lee, S.; Takahashi, I.; Matsuzaka, M.; Yamai, K.; Danjo, K.; Kumagai, T.; Umeda, T.; Itai, K.; Nakaji, S. The relationship between serum selenium concentration and neutrophil function in peripheral blood. Biol. Trace Elem. Res. 2011, 144, 396–406. [Google Scholar] [CrossRef]
- Tolando, R.; Jovanovic, A.; Brigelius-Flohé, R.; Ursini, F.; Maiorino, M. Reactive oxygen species and proinflammatory cytokine signaling in endothelial cells: Effect of selenium supplementation. Free Radic. Biol. Med. 2000, 28, 979–986. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Manda-Handzlik, A.; Demkow, U. Neutrophils: The Role of Oxidative and Nitrosative Stress in Health and Disease. Adv. Exp. Med. Biol. 2015, 857, 51–60. [Google Scholar]
- Lartey, N.L.; Vargas-Robles, H.; Guerrero-Fonseca, I.M.; Nava, P.; Kumatia, E.K.; Ocloo, A.; Schnoor, M. Annickiapolycarpa extract attenuates inflammation, neutrophil recruitment, and colon damage during colitis. Immunol. Lett. 2022, 248, 99–108. [Google Scholar] [CrossRef]
- Bizerea, T.O.; Dezsi, S.G.; Marginean, O.; Stroescu, R.; Rogobete, A.; Bizerea-Spiridon, O.; Ilie, C. The Link Between Selenium, Oxidative Stress and Pregnancy Induced Hypertensive Disorders. Clin. Lab. 2018, 64, 1593–1610. [Google Scholar] [CrossRef] [PubMed]
- Soares de Oliveira, A.R.; JayanneClímaco Cruz, K.; Beatriz Silva Morais, J.; Rocha Dos Santos, L.; Rodrigues de Sousa Melo, S.; Fontenelle, L.C.; Santos de Sousa, G.; Costa Maia, C.S.; Oliveira Duarte de Araújo, C.; Leal Mendes, I.; et al. Selenium status and oxidative stress in obese: Influence of adiposity. Eur. J. Clin. Invest. 2021, 51, 13538. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.É.L.; Dias, T.M.D.S.; Moura, M.S.B.; Soares, N.R.M.; Pierote, N.R.A.; Araújo, C.O.D.; Maia, C.S.C.; Henriques, G.S.; Barros, V.C.; Moita Neto, J.M.; et al. Relationship between selenium status and biomarkers of oxidative stress in Crohn's disease. Nutrition 2020, 74, 110762. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, T.; Moon, T.C.; St Laurent, C.D.; Puttagunta, L.; Chung, K.; Wright, E.; Yoshikawa, M.; Moriyama, H.; Befus, A.D. Expression of nitric oxide synthases in leukocytes in nasal polyps. Ann. Allergy Asthma Immunol. 2012, 108, 172–177. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Sadeghian, S.; Kojouri, G.A.; Mohebbi, A. Nanoparticles of selenium as species with stronger physiological effects in sheep in comparison with sodium selenite. Biol. Trace Elem. Res. 2012, 146, 302–308. [Google Scholar] [CrossRef]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Oxidative stress and vascular inflammation in aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar] [CrossRef]
- Zhou, J.; Zeng, L.; Zhang, Y.; Wang, M.; Li, Y.; Jia, Y.; Wu, L.; Su, P. Cadmium exposure induces pyroptosis in testicular tissue by increasing oxidative stress and activating the AIM2 inflammasome pathway. Sci. Total Environ. 2022, 20, 157500. [Google Scholar] [CrossRef]
- Yang, T.; Zhao, Z.; Liu, T.; Zhang, Z.; Wang, P.; Xu, S.; Lei, X.G.; Shan, A. Oxidative stress induced by Se-deficient high-energy diet implicates neutrophil dysfunction via Nrf2 pathway suppression in swine. Oncotarget 2017, 8, 13428–13439. [Google Scholar] [CrossRef][Green Version]
- Alhussien, M.N.; Dang, A.K. JAK3 and PI3K mediates the suppressive effects of interferon tau on neutrophil extracellular traps formation during peri-implantation period. J. Reprod. Immunol. 2021, 145, 103321. [Google Scholar] [CrossRef]
- Chi, Q.; Zhang, Q.; Lu, Y.; Zhang, Y.; Xu, S.; Li, S. Roles of selenoprotein S in reactive oxygen species-dependent neutrophil extracellular trap formation induced by selenium-deficient arteritis. Redox. Biol. 2021, 44, 102003. [Google Scholar] [CrossRef] [PubMed]
- Zenlander, R.; Havervall, S.; Magnusson, M.; Engstrand, J.; Ågren, A.; Thålin, C.; Stål, P. Neutrophil extracellular traps in patients with liver cirrhosis and hepatocellular carcinoma. Sci. Rep. 2021, 11, 18025. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Li, X.; Xing, M.; Chen, M.; Zhao, J.; Fan, R.; Zhao, X.; Cao, C.; Yang, J.; Zhang, Z.; Xu, S. Effects of selenium-lead interaction on the gene expression of inflammatory factors and selenoproteins in chicken neutrophils. Ecotoxicol. Environ. Saf. 2017, 139, 447–453. [Google Scholar] [CrossRef]
- Tin-Tin-Win-Shwe; Mitsushima, D.; Yamamoto, S.; Fukushima, A.; Funabashi, T.; Kobayashi, T.; Fujimaki, H. Changes in neurotransmitter levels and proinflammatory cytokine mRNA expressions in the mice olfactory bulb following nanoparticle exposure. Toxicol. Appl. Pharmacol. 2008, 226, 192–198. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Y.; Zeng, X.; Bo, L.; Jiang, S.; Du, X.; Xie, Y.; Jiang, R.; Zhao, J.; Song, W. Investigation of selenium pretreatment in the attenuation of lung injury in rats induced by fine particulate matters. Environ. Sci. Pollut. Res. Int. 2017, 24, 4008–4017. [Google Scholar] [CrossRef]
- Zheng, S.; Song, H.; Gao, H.; Liu, C.; Zhang, Z.; Fu, J. The Antagonistic Effect of Selenium on Lead-Induced Inflammatory Factors and Heat Shock Protein mRNA Level in Chicken Cartilage Tissue. Biol. Trace Elem. Res. 2016, 173, 177–184. [Google Scholar] [CrossRef]
- Tan, S.; Chi, Q.; Liu, T.; Sun, Z.; Min, Y.; Zhang, Z.; Li, S. Alleviation Mechanisms of Selenium on Cadmium-Spiked Neutrophil Injury to Chicken. Biol. Trace Elem. Res. 2017, 178, 301–309. [Google Scholar] [CrossRef]
- Tamassia, N.; Cassatella, M.A.; Bazzoni, F. Fast and Accurate Quantitative Analysis of Cytokine Gene Expression in Human Neutrophils by Reverse Transcription Real-Time PCR. Methods Mol. Biol. 2020, 2087, 243–260. [Google Scholar]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef]
- Mistry, H.D.; Broughton Pipkin, F.; Redman, C.W.; Poston, L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, J.; Xiao, J. Selenoproteins and selenium status in bone physiology and pathology. Biochim. Biophys. Acta. 2014, 1840, 3246–3256. [Google Scholar] [CrossRef] [PubMed]
- Khoso, P.A.; Yang, Z.; Liu, C.; Li, S. Selenium Deficiency Downregulates Selenoproteins and Suppresses Immune Function in Chicken Thymus. Biol. Trace Elem. Res. 2015, 167, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Moschos, M.P. Selenoprotein P. Cell Mol. Life Sci. 2000, 57, 1836–1845. [Google Scholar] [CrossRef]
- Shetty, S.; Copeland, P.R. Molecular mechanism of selenoprotein P synthesis. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2506–2510. [Google Scholar] [CrossRef]
- Bevinakoppamath, S.; Saleh Ahmed, A.M.; Ramachandra, S.C.; Vishwanath, P.; Prashant, A. Chemopreventive and Anticancer Property of Selenoproteins in Obese Breast Cancer. Front. Pharmacol. 2021, 12, 618172. [Google Scholar] [CrossRef]
- Gong, T.; Hashimoto, A.C.; Sasuclark, A.R.; Khadka, V.S.; Gurary, A.; Pitts, M.W. Selenoprotein M Promotes Hypothalamic Leptin Signaling and Thioredoxin Antioxidant Activity. Antioxid. Redox Signal. 2021, 35, 775–787. [Google Scholar] [CrossRef]
- Hwang, D.Y.; Sin, J.S.; Kim, M.S.; Yim, S.Y.; Kim, Y.K.; Kim, C.K.; Kim, B.G.; Shim, S.B.; Jee, S.W.; Lee, S.H.; et al. Overexpression of human selenoprotein M differentially regulates the concentrations of antioxidants and H2O2, the activity of antioxidant enzymes, and the composition of white blood cells in a transgenic rat. Int. J. Mol. Med. 2008, 21, 169–179. [Google Scholar] [CrossRef][Green Version]
- Marciel, M.P.; Hoffmann, P.R. Molecular Mechanisms by Which Selenoprotein K Regulates Immunity and Cancer. Biol. Trace Elem. Res. 2019, 192, 60–68. [Google Scholar] [CrossRef]
- Verma, S.; Hoffmann, F.W.; Kumar, M.; Huang, Z.; Roe, K.; Nguyen-Wu, E.; Hashimoto, A.S.; Hoffmann, P.R. Selenoprotein K knockout mice exhibit deficient calcium flux in immune cells and impaired immune responses. J. Immunol. 2011, 186, 2127–2137. [Google Scholar] [CrossRef]
- Norton, R.L.; Fredericks, G.J.; Huang, Z.; Fay, J.D.; Hoffmann, F.W.; Hoffmann, P.R. Selenoprotein K regulation of palmitoylation and calpain cleavage of ASAP2 is required for efficient FcγR-mediated phagocytosis. J. Leukoc. Biol. 2017, 101, 439–448. [Google Scholar] [CrossRef]
- Xia, H.; Wang, Y.; Dai, J.; Zhang, X.; Zhou, J.; Zeng, Z.; Jia, Y. Selenoprotein K Is Essential for the Migration and Phagocytosis of Immature Dendritic Cells. Antioxidants 2022, 11, 1264. [Google Scholar] [CrossRef]
- Fredericks, G.J.; Hoffmann, F.W.; Rose, A.H.; Osterheld, H.J.; Hess, F.M.; Mercier, F.; Hoffmann, P.R. Stable expression and function of the 519 inositol 1,4,5-triphosphate receptor requires palmitoylation by a DHHC6/selenoprotein K complex. Proc. Natl. Acad. Sci. USA 2014, 111, 16478–16483. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It's role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef]
- Anouar, Y.; Lihrmann, I.; Falluel-Morel, A.; Boukhzar, L. Selenoprotein T is a key player in ER proteostasis, endocrine homeostasis and neuroprotection. Free Radic. Biol. Med. 2018, 127, 145–152. [Google Scholar] [CrossRef]
- Schweizer, U.; Bohleber, S.; Zhao, W.; Fradejas-Villar, N. The Neurobiology of Selenium: Looking Back and to the Future. Front. Neurosci. 2021, 15, 652099. [Google Scholar] [CrossRef]
- Pothion, H.; Jehan, C.; Tostivint, H.; Cartier, D.; Bucharles, C.; Falluel-Morel, A.; Boukhzar, L.; Anouar, Y.; Lihrmann, I. Selenoprotein T: An Essential Oxidoreductase Serving as a Guardian of Endoplasmic Reticulum Homeostasis. Antioxid. Redox Signal. 2020, 33, 1257–1275. [Google Scholar] [CrossRef]
- Ma, C.; Martinez-Rodriguez, V.; Hoffmann, P.R. Roles for Selenoprotein I and Ethanolamine Phospholipid Synthesis in T Cell Activation. Int. J. Mol. Sci. 2021, 22, 11174. [Google Scholar] [CrossRef]
- Safronova, V.G.; Vulfius, C.A.; Shelukhina, I.V.; Mal’tseva, V.N.; Berezhnov, A.V.; Fedotova, E.I.; Miftahova, R.G.; Kryukova, E.V.; Grinevich, A.A.; Tsetlin, V.I. Nicotinic receptor involvement in regulation of functions of mouse neutrophils from inflammatory site. Immunobiology 2016, 221, 761–772. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAPDH | Forward 5′-tccactcacggcaaattcaac-3′ Reverse 5′-cggcatcgaaggtggaagag-3′ |
| CD62L | Forward 5′-gcagagaactggggtgctgg-3′ Reverse 5′-caactcaggggcctccaaagg-3′ |
| CD11b | Forward 5′-gaagggcatggggctctgaag-3′ Reverse 5′-cagtttgttcccaaaggaggcat-3′ |
| NOX2 | Forward 5′-ttgggtcagcactggctctgg-3′ Reverse 5′-caggcccccttcagggttcttg-3′ |
| NOS-2 | Forward 5′-gctgcaggtgttcgatgccc-3′ Reverse 5′-ccaaggtagagccatctggctgctt-3′ |
| MPO | Forward 5′-ccttcgtggacgccagcg-3′ Reverse 5′-aagagggtgtgcatggaggtg-3′ |
| HO-1 | Forward 5′-ggtcaggtgtccagggaaggct-3′ Reverse 5′-ccctgagaggtcacccaggtagc-3′ |
| Catalase | Forward 5′-gctgacacagttcgtgaccctcg-3′ Reverse 5′- acaggcaagtttttgatgccctggt-3′ |
| SOD2 | Forward 5′-ctcccggcacaagcacagc-3′ Reverse 5′-tcctttgggttctccaccaccct -3′ |
| Nrf2 | Forward 5′-tcctggacgggactattgaaggctg-3′ Reverse 5′-cacattgggattcacgcataggagcact-3′ |
| TNFa | Forward 5′-acggcatggatctcaaagacaac-3′ Reverse 5′-tcctggtatgagatagcaaatcgg-3′ |
| IL1b | Forward 5′-aatctcgcagcagcacatcaaca-3′ Reverse 5′-tccacgggaaagacacaggtagc-3′ |
| Il6 | Forward 5′-aaactctaattcatatcttcaac-3′ Reverse 5′-gtccacaaactgatatgcttag-3′ |
| IL10 | Forward 5′-tgtcatcgatttctcccctgtga-3′ Reverse 5′-cattcatggccttgtagacaccttg-3′ |
| NF-kB | Forward 5′-aagtgcaaaggaaacgccagaa-3′ Reverse 5′-actaccgaacatgcctccacca-3′ |
| SOCS3 | Forward 5′-aagaacctacgcatccagtgtga-3′ Reverse 5′-atgtagtggtgcaccagcttgag-3′ |
| NE | Forward 5′-catgggctggggcaggttg-3′ Reverse 5′-ggcgaaggcatctgggtacaa-3′ |
| H2A.1 | Forward 5′-сcgcaagggcaactactcgg-3′ Reverse 5′-ctcggtcttcttgggcagcag-3′ |
| H2B | Forward 5′-caagtgcaccccgacaccg-3′ Reverse 5′-ctccgacaccgcgtgct-3′ |
| H3 | Forward 5′-ccagaagtcgaccgagctgc-3′ Reverse 5′-gatgtccttgggcatgatggtga-3′ |
| SELENOS | Forward 5′-tgggacagcatgcaagaag-3′ Reverse 5′-gcgtccaggtctccagg-3′ |
| SELENOM | Forward 5′-agcctcctgttgcctccgc-3′ Reverse 5′-aggtcagcgtggtccgaag-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mal’tseva, V.N.; Gudkov, S.V.; Turovsky, E.A. Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo. Int. J. Mol. Sci. 2022, 23, 13651. https://doi.org/10.3390/ijms232113651
Mal’tseva VN, Gudkov SV, Turovsky EA. Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo. International Journal of Molecular Sciences. 2022; 23(21):13651. https://doi.org/10.3390/ijms232113651
Chicago/Turabian StyleMal’tseva, Valentina N., Sergey V. Gudkov, and Egor A. Turovsky. 2022. "Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo" International Journal of Molecular Sciences 23, no. 21: 13651. https://doi.org/10.3390/ijms232113651
APA StyleMal’tseva, V. N., Gudkov, S. V., & Turovsky, E. A. (2022). Modulation of the Functional State of Mouse Neutrophils by Selenium Nanoparticles In Vivo. International Journal of Molecular Sciences, 23(21), 13651. https://doi.org/10.3390/ijms232113651