
Thyroxine Regulates the Opening of the Organ of Corti through Affecting P-Cadherin and Acetylated Microtubule


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
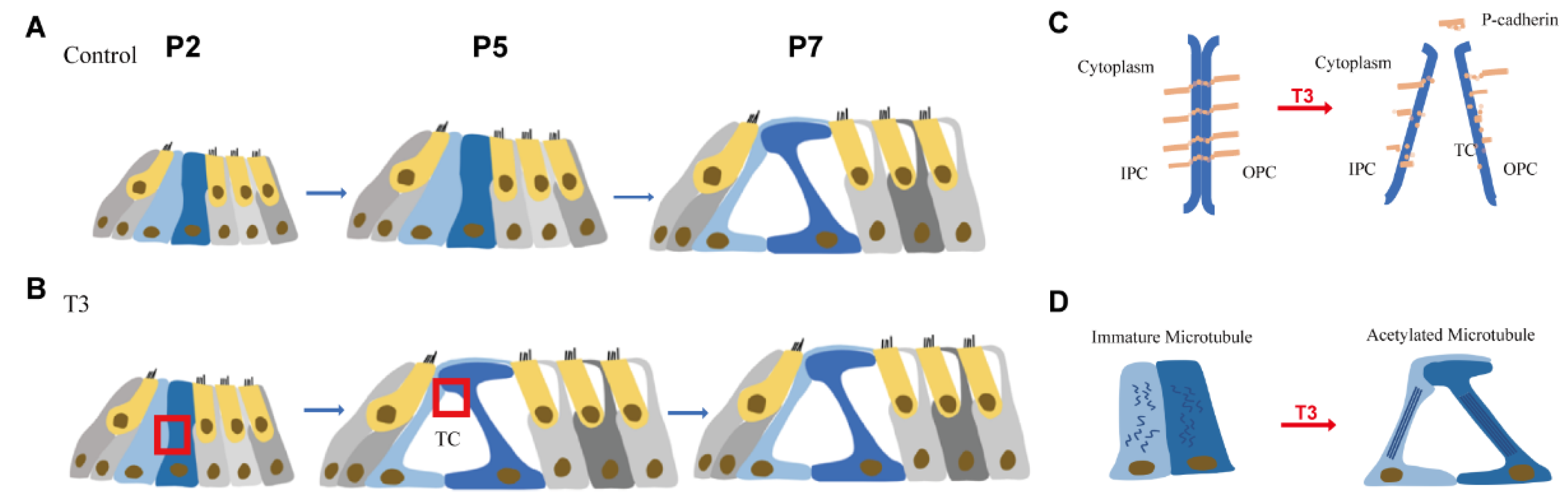
2.1. Normal Opening of the TC and Early Opening Promoted by T3
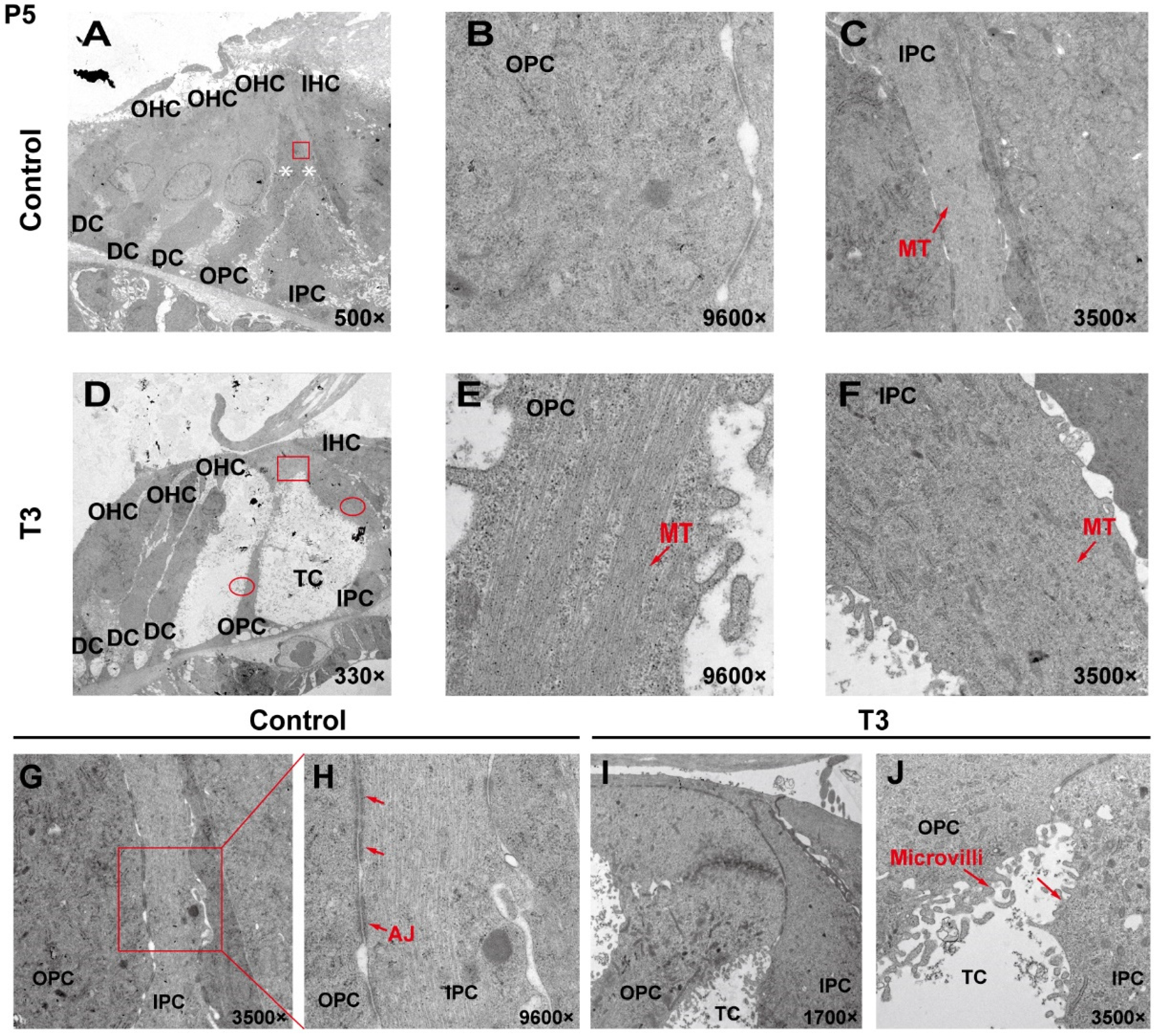
2.2. The Ultrastructure of PCs Changes Significantly after T3 Administration
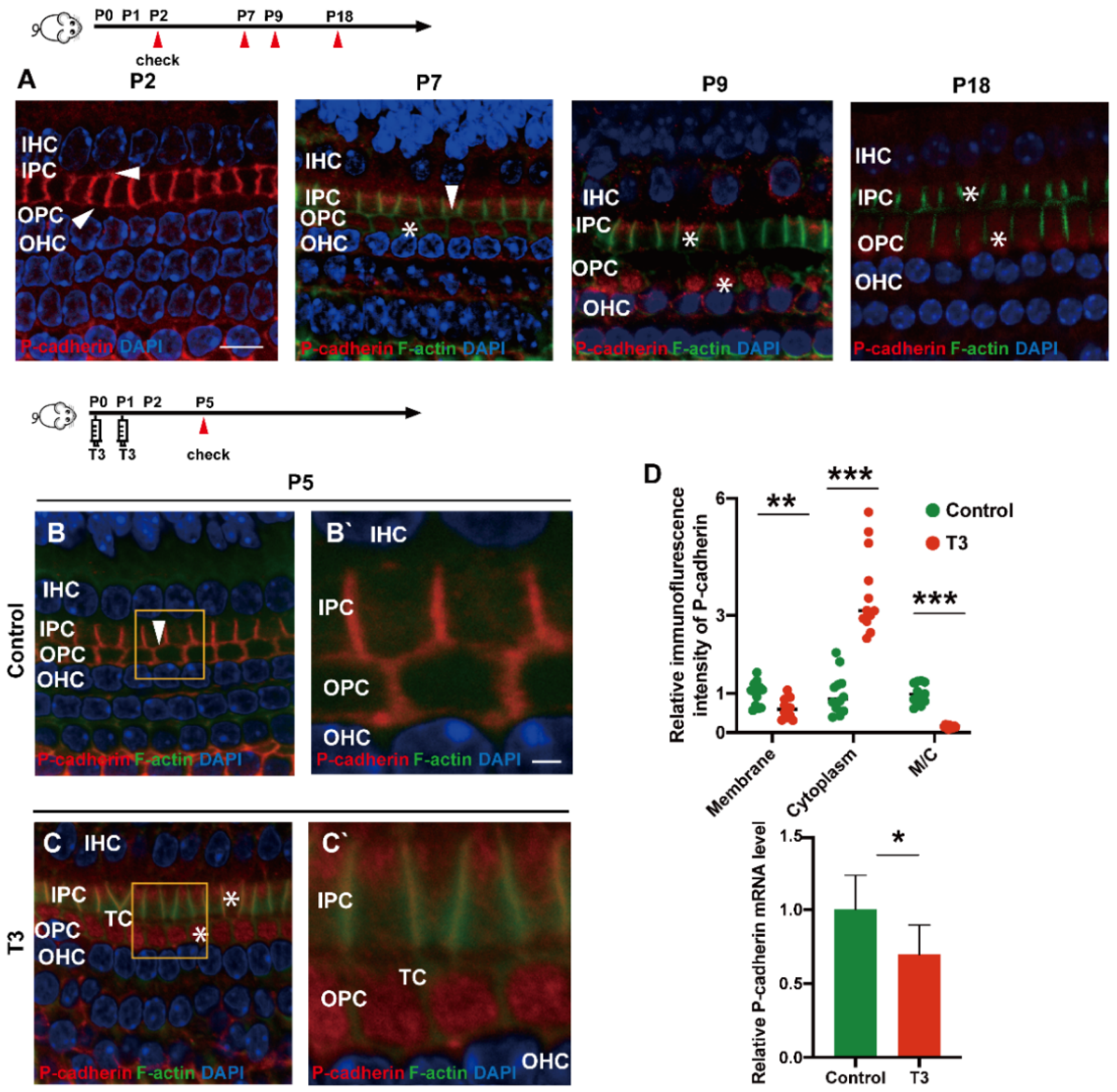
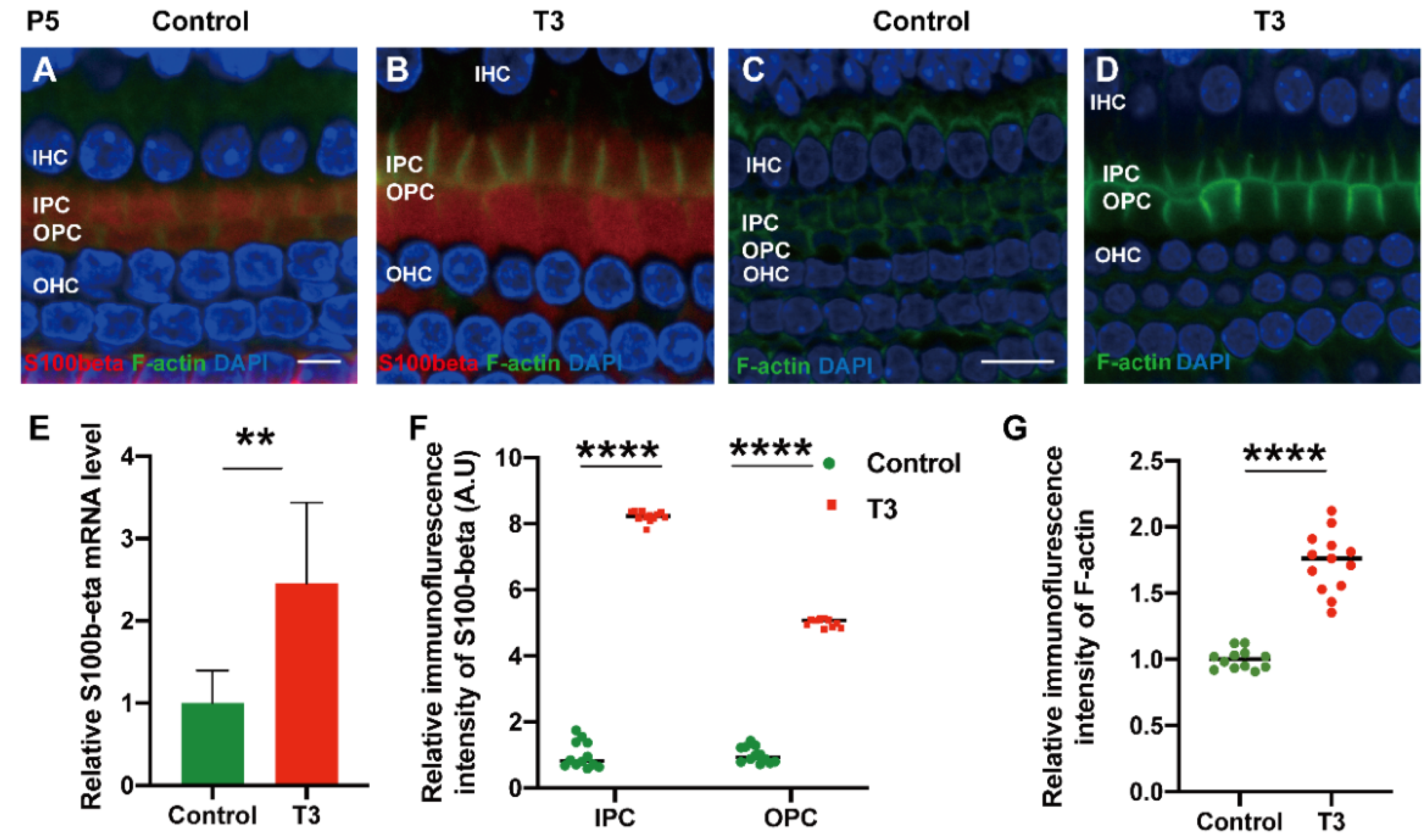
2.3. T3 Promotes the Separation of IPCs and OPCs by Affecting P-Cadherin Transfer into the Cytoplasm from the Cell Membrane
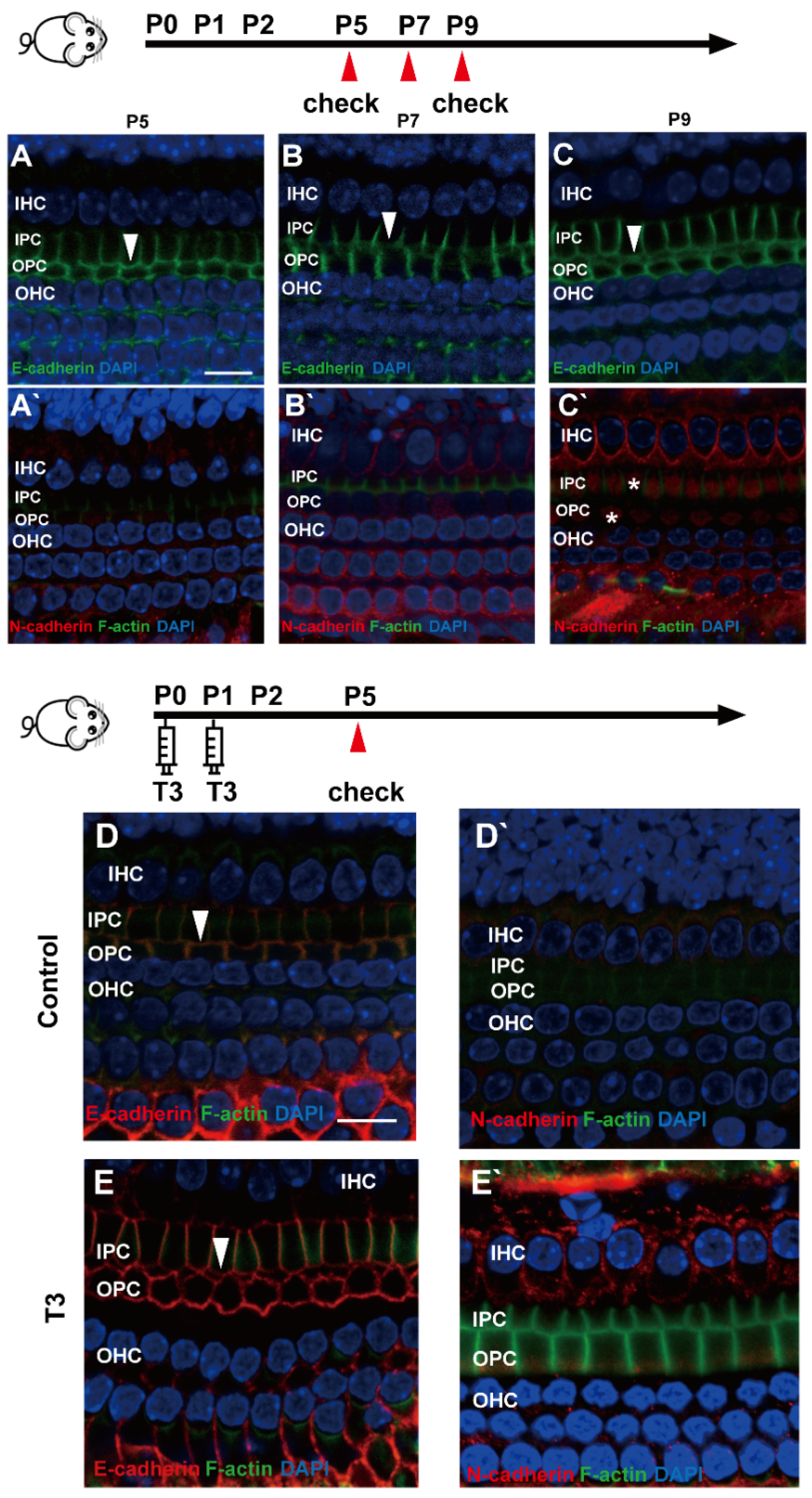
2.4. E-Cadherin and N-Cadherin Do Not Change Significantly during Opening of the TC
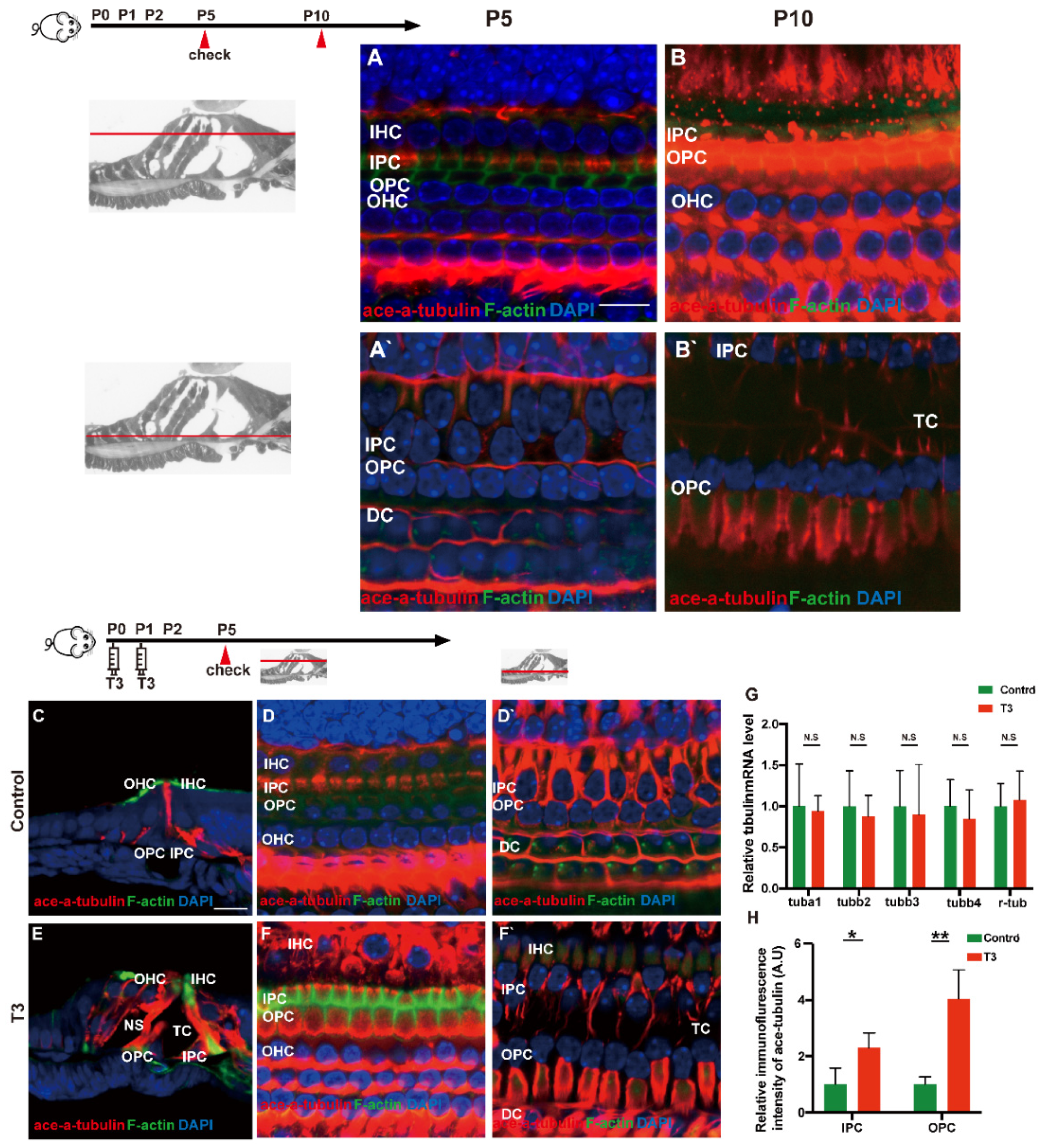
2.5. T3 influences the Growth of PCs via Acetylated α-Tubulin
3. Discussion
4. Materials and Methods
4.1. Animals and Models
4.2. Immunofluorescent Labelling and Confocal Image Analysis
4.3. Resin Sections and Transmission Electron Microscopy
4.4. RNA Preparation and Quantitative RT-PCR
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharlin, D.S.; Ng, L.; Verrey, F.; Visser, T.J.; Liu, Y.; Olszewski, R.T.; Hoa, M.; Heuer, H.; Forrest, D. Deafness and loss of cochlear hair cells in the absence of thyroid hormone transporters Slc16a2 (Mct8) and Slc16a10 (Mct10). Sci. Rep. 2018, 8, 4403. [Google Scholar] [CrossRef] [PubMed]
- Rusch, A.; Ng, L.; Goodyear, R.; Oliver, D.; Lisoukov, I.; Vennstrom, B.; Richardson, G.; Kelley, M.W.; Forrest, D. Retardation of cochlear maturation and impaired hair cell function caused by deletion of all known thyroid hormone receptors. J. Neurosci. 2001, 21, 9792–9800. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.; Goodyear, R.J.; Woods, C.A.; Schneider, M.J.; Diamond, E.; Richardson, G.P.; Kelley, M.W.; Germain, D.L.; Galton, V.A.; Forrest, D. Hearing loss and retarded cochlear development in mice lacking type 2 iodothyronine deiodinase. Proc. Natl. Acad. Sci. USA 2004, 101, 3474–3479. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.; Liu, Y.; Liu, H.; Forrest, D. Cochlear Fibrocyte and Osteoblast Lineages Expressing Type 2 Deiodinase Identified with a Dio2CreERt2 Allele. Endocrinology 2021, 162, bqab179. [Google Scholar] [CrossRef] [PubMed]
- Peeters, R.P.; Ng, L.; Ma, M.; Forrest, D. The timecourse of apoptotic cell death during postnatal remodeling of the mouse cochlea and its premature onset by triiodothyronine (T3). Mol. Cell. Endocrinol. 2015, 407, 1–8. [Google Scholar] [CrossRef]
- Szarama, K.B.; Gavara, N.; Petralia, R.S.; Chadwick, R.S.; Kelley, M.W. Thyroid hormone increases fibroblast growth factor receptor expression and disrupts cell mechanics in the developing organ of corti. BMC Dev. Biol. 2013, 13, 6. [Google Scholar] [CrossRef]
- Mansour, S.L.; Li, C.; Urness, L.D. Genetic rescue of Muenke syndrome model hearing loss reveals prolonged FGF-dependent plasticity in cochlear supporting cell fates. Genes Dev. 2013, 27, 2320–2331. [Google Scholar] [CrossRef]
- Wang, Y.; Chang, Q.; Tang, W.; Sun, Y.; Zhou, B.; Li, H.; Lin, X. Targeted connexin26 ablation arrests postnatal development of the organ of Corti. Biochem. Biophys. Res. Commun. 2009, 385, 33–37. [Google Scholar] [CrossRef]
- Colvin, J.S.; Bohne, B.A.; Harding, G.W.; McEwen, D.G.; Ornitz, D.M. Skeletal overgrowth and deafness in mice lacking fibroblast growth factor receptor 3. Nat. Genet. 1996, 12, 390–397. [Google Scholar] [CrossRef]
- Nunes, F.D.; Lopez, L.N.; Lin, H.W.; Davies, C.; Azevedo, R.B.; Gow, A.; Kachar, B. Distinct subdomain organization and molecular composition of a tight junction with adherens junction features. J. Cell Sci. 2006, 119, 4819–4827. [Google Scholar] [CrossRef]
- Defourny, J.; Peuckert, C.; Kullander, K.; Malgrange, B. EphA4-ADAM10 Interplay Patterns the Cochlear Sensory Epithelium through Local Disruption of Adherens Junctions. iScience 2019, 11, 246–257. [Google Scholar] [CrossRef]
- Liu, W.; Wang, C.; Yu, H.; Liu, S.; Yang, J. Expression of acetylated tubulin in the postnatal developing mouse cochlea. Eur. J. Histochem. EJH 2018, 62. [Google Scholar] [CrossRef]
- Coopman, P.; Djiane, A. Adherens Junction and E-Cadherin complex regulation by epithelial polarity. Cell. Mol. Life Sci. CMLS 2016, 73, 3535–3553. [Google Scholar] [CrossRef]
- Priest, A.V.; Shafraz, O.; Sivasankar, S. Biophysical basis of cadherin mediated cell-cell adhesion. Exp. Cell Res. 2017, 358, 10–13. [Google Scholar] [CrossRef]
- Wu, J.; Jester, W.F., Jr.; Laslett, A.L.; Meinhardt, A.; Orth, J.M. Expression of a novel factor, short-type PB-cadherin, in Sertoli cells and spermatogenic stem cells of the neonatal rat testis. J. Endocrinol. 2003, 176, 381–391. [Google Scholar] [CrossRef]
- Johnen, N.; Francart, M.E.; Thelen, N.; Cloes, M.; Thiry, M. Evidence for a partial epithelial-mesenchymal transition in postnatal stages of rat auditory organ morphogenesis. Histochem. Cell Biol. 2012, 138, 477–488. [Google Scholar] [CrossRef]
- Defourny, J.; Thiry, M. Tricellular adherens junctions provide a cell surface delivery platform for connexin 26/30 oligomers in the cochlea. Hear. Res. 2021, 400, 108137. [Google Scholar] [CrossRef]
- Whitlon, D.S. E-cadherin in the mature and developing organ of Corti of the mouse. J. Neurocytol. 1993, 22, 1030–1038. [Google Scholar] [CrossRef]
- Szarama, K.B.; Gavara, N.; Petralia, R.S.; Kelley, M.W.; Chadwick, R.S. Cytoskeletal changes in actin and microtubules underlie the developing surface mechanical properties of sensory and supporting cells in the mouse cochlea. Development 2012, 139, 2187–2197. [Google Scholar] [CrossRef]
- Szarama, K.B.; Stepanyan, R.; Petralia, R.S.; Gavara, N.; Frolenkov, G.I.; Kelley, M.W.; Chadwick, R.S. Fibroblast growth factor receptor 3 regulates microtubule formation and cell surface mechanical properties in the developing organ of Corti. Bioarchitecture 2012, 2, 214–219. [Google Scholar] [CrossRef]
- Xie, L.; Chen, S.; Xu, K.; Cao, H.Y.; Du, A.N.; Bai, X.; Sun, Y.; Kong, W.J. Reduced postnatal expression of cochlear Connexin26 induces hearing loss and affects the developmental status of pillar cells in a dose-dependent manner. Neurochem. Int. 2019, 128, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Chausovsky, A.; Bershadsky, A.D.; Borisy, G.G. Cadherin-mediated regulation of microtubule dynamics. Nat. Cell Biol. 2000, 2, 797–804. [Google Scholar] [CrossRef]
- Henderson, C.G.; Tucker, J.B.; Chaplin, M.A.; Mackie, J.B.; Maidment, S.N.; Mogensen, M.M.; Paton, C.C. Reorganization of the centrosome and associated microtubules during the morphogenesis of a mouse cochlear epithelial cell. J. Cell Sci. 1994, 107 Pt 2, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Sorci, G.; Agneletti, A.L.; Donato, R. Effects of S100A1 and S100B on microtubule stability. An in vitro study using triton-cytoskeletons from astrocyte and myoblast cell lines. Neuroscience 2000, 99, 773–783. [Google Scholar] [CrossRef]
- Sorci, G.; Agneletti, A.L.; Bianchi, R.; Donato, R. Association of S100B with intermediate filaments and microtubules in glial cells. Biochim. Biophys. Acta 1998, 1448, 277–289. [Google Scholar] [CrossRef]
- Knipper, M.; Bandtlow, C.; Gestwa, L.; Köpschall, I.; Rohbock, K.; Wiechers, B.; Zenner, H.P.; Zimmermann, U. Thyroid hormone affects Schwann cell and oligodendrocyte gene expression at the glial transition zone of the VIIIth nerve prior to cochlea function. Development 1998, 125, 3709–3718. [Google Scholar] [CrossRef]
- Zaidel-Bar, R. Cadherin adhesome at a glance. J. Cell Sci. 2013, 126, 373–378. [Google Scholar] [CrossRef]
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. The role of thyroid hormone in testicular development and function. J. Endocrinol. 2008, 199, 351–365. [Google Scholar] [CrossRef]
- Nunez, J. Microtubules and brain development: The effects of thyroid hormones. Neurochem. Int. 1985, 7, 959–968. [Google Scholar] [CrossRef]
- Geiger, B. A 130K protein from chicken gizzard: Its localization at the termini of microfilament bundles in cultured chicken cells. Cell 1979, 18, 193–205. [Google Scholar] [CrossRef]
- Taneyhill, L.A.; Schiffmacher, A.T. Cadherin dynamics during neural crest cell ontogeny. Prog. Mol. Biol. Transl. Sci. 2013, 116, 291–315. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Xie, L.; Chen, S.; Qiu, Y.; Sun, Y.; Kong, W. Thyroxine Regulates the Opening of the Organ of Corti through Affecting P-Cadherin and Acetylated Microtubule. Int. J. Mol. Sci. 2022, 23, 13339. https://doi.org/10.3390/ijms232113339
Zhang H, Xie L, Chen S, Qiu Y, Sun Y, Kong W. Thyroxine Regulates the Opening of the Organ of Corti through Affecting P-Cadherin and Acetylated Microtubule. International Journal of Molecular Sciences. 2022; 23(21):13339. https://doi.org/10.3390/ijms232113339
Chicago/Turabian StyleZhang, Huimin, Le Xie, Sen Chen, Yue Qiu, Yu Sun, and Weijia Kong. 2022. "Thyroxine Regulates the Opening of the Organ of Corti through Affecting P-Cadherin and Acetylated Microtubule" International Journal of Molecular Sciences 23, no. 21: 13339. https://doi.org/10.3390/ijms232113339
APA StyleZhang, H., Xie, L., Chen, S., Qiu, Y., Sun, Y., & Kong, W. (2022). Thyroxine Regulates the Opening of the Organ of Corti through Affecting P-Cadherin and Acetylated Microtubule. International Journal of Molecular Sciences, 23(21), 13339. https://doi.org/10.3390/ijms232113339