Abstract
L-Asparagine (Asn) has been regarded as one of the most economical molecules for nitrogen (N) storage and transport in plants due to its relatively high N-to-carbon (C) ratio (2:4) and stability. Although its internal function has been addressed, the biological role of exogenous Asn in plants remains elusive. In this study, different concentrations (0.5, 1, 2, or 5 mM) of Asn were added to the N-deficient hydroponic solution for poplar ‘Nanlin895’. Morphometric analyses showed that poplar height, biomass, and photosynthesis activities were significantly promoted by Asn treatment compared with the N-free control. Moreover, the amino acid content, total N and C content, and nitrate and ammonia content were dramatically altered by Asn treatment. Moreover, exogenous Asn elicited root growth inhibition, accompanied by complex changes in the transcriptional pattern of genes and activities of enzymes associated with N and C metabolism. Combined with the plant phenotype and the physiological and biochemical indexes, our data suggest that poplar is competent to take up and utilize exogenous Asn dose-dependently. It provides valuable information and insight on how different forms of N and concentrations of Asn influence poplar root and shoot growth and function, and roles of Asn engaged in protein homeostasis regulation.
1. Introduction
Nitrogen (N) is a crucial element for plant growth and development, and one of the most critical limiting factors for biomass and yield formation. Hence, the efficient utilization of N is the most central event for desirable plant growth performance and productivity [1,2]. Broadly, the source of N utilized by plants in the environment can be grouped into organic and inorganic. The choice of uptake of these nutrients by plants depends on plant species and soil abundance of N (nitrate (NO3−), ammonium (NH4+), amino acids, etc.) [3], whereas the utilization of N compounds depends on the N availability and the capacity of transporters for the uptake, and the endogenous metabolism system for the subsequent utilization [4,5,6,7,8].
Although the main route of N uptake by plants is realized through inorganic means, the use of organic N, which can be directly absorbed by plants in the form of amino acid, has been presented by many researchers [5,9,10,11,12]. Impressive results reveal that amino acids are a very efficacious source of N for plant growth, since the majority of N in cells is bound up to them [13]. Furthermore, plants’ uptake and utilization of amino acids as a source of N are more advantageous energetically during assimilation than that of NO3− and NH4+ [14]. Beyond their fertilizing properties, natural amino acids are nontoxic, biodegradable molecules and are potential chelators for soil remediation [15]. Moreover, amino acids represent one of the major categories of biostimulants, which have been reported to improve plant vigor, crop yield, quality characteristics, and tolerance to biotic and abiotic stresses [16]. Collectively, the interests in the application of amino acid-based N in agroforestry are increasing because of its environmentally friendly features and demonstrated capacities to improve plant growth performance [6,17,18,19,20,21,22,23,24,25,26]. Despite a long history being available about the role of amino acids in plants, their effects have varied depending on the type of amino acid over plant species; therefore, extensive studies are required to explain the physiological functions and mode of action of individual amino acids [27].
L-asparagine (Asn) is one of the twenty naturally occurring proteinogenic amino acids on Earth. It serves as a principal source of N for protein synthesis, especially in plants’ vigorously growing tissues [28,29]. Asn has carboxamide as the side chain’s functional group, which often plays a key role in the active site of enzymes [30]. Free Asn contains 21.2% N, and it has a relatively high ratio of N to C (2:4) and stability compared to the other amino acids [31,32]; thus, is commonly considered as a main organic nitrogenous storage, transport, and partitioning compound in many plant species, in particular when the supply of soluble carbohydrate is severely reduced [33]. In addition, Asn is crucial in coordinating general translation, modulating cellular signaling for amino acid homeostasis and regulating metabolic availability during biological processes [34,35]. Although Asn is a non-essential amino acid, it has been characterized as an essential exchange factor that regulates mTOR complex 1 (mTORC1, a central regulator of cell growth and division) activation, thereby coordinating protein and nucleotide biosynthesis [36].
Asn generally accounts for 24–76% of the xylem N in tree species [37]. The proportion of Asn in the xylem can be taken as an indicator of plant N status, wherein a decrease in N availability frequently results in a reduction in Asn and an increase in aspartate [33]. Asn is selectively accumulated in various plant tissues during normal physiological processes such as seed germination and leaf senescence, or when plants are placed in the dark [38] or exposed to a range of abiotic/biotic stresses [33,39,40,41]. The conversion of Asp to Asn in plants has been demonstrated to correlate with enhanced disease resistance [42]. Aside from the roles shown above for endogenous Asn, exogenous Asn has been found to play a part in many biological processes, such as stimulating shooting and minimizing withering of Rosa centifolia [13], alleviating salt stress in maize [43], altering root growth in Arabidopsis [44], promoting chlorophyll content, sugar metabolism and proline accumulation in cherry rootstocks [45], inducing somatic embryo formation [46], and improving Phaseolus vulgaris growth by stimulating cell elongation and division [47]. Furthermore, exogenous Asn has been reported as a growth regulator that positively improves vegetative growth, nutrient uptake, and pigment production in garden cress [48]. When combined with sucrose application, exogenous Asn stimulates additional outgrowth of the collateral buds in Rosa hybrid [49].
Poplar is a fast-growing woody plant species planted worldwide to produce wood, biomaterials, and biofuel [50,51]. Although Asn is one of the predominant translocated forms of organic N in poplar [52], the effect of Asn on poplar growth has rarely been reported so far. Previously, several studies, including ours [17,53,54], have demonstrated the ability of poplars to absorb and utilize various types of amino acids. This motivates us to conduct the present study to evaluate the effects of Asn on the vegetative growth and the physiological and biochemical traits of poplar plants.
2. Results
2.1. Exogenous Treatment of Asn Has Significant Impacts on Poplar Growth
To explore the modulation effect of exogenous Asn on poplar growth, different concentrations of Asn (i.e., 0.5, 1, 2, or 5 mM) were supplemented as a sole N source in the culture medium for poplar ‘Nanlin895’, in parallel with 0 mM and 3 mM KNO3 serving as the N0 and N+ control, respectively. Impressively, the morphology and the growth parameters of poplar ‘Nanlin895’ seedlings were significantly affected by Asn treatment (Figure 1 and Table 1). As seen from Figure 1, Asn treatment strongly influenced the morphophysiological traits of poplar ’Nanlin895’. Specifically, in the N0 group, plants underwent growth retardation, yellowing, and the easy falling off of mature leaves (Figure 1b). However, such effects were reversed by the treatment of varied concentrations of Asn (0.5, 1, or 2 mM) (Figure 1c–e), whose addition recorded an increase of seedling height, shoot fresh weight, leaf numbers, and leaf area by 22–65%, 63–201%, 54–86%, and 23–132%, respectively. Nonetheless, plants showed a decrease in root length (by 38–54%) and root-to-shoot ratio (by 49–62%) compared with the N0 control (Table 1). In addition, it was realized that the increment in Asn supply was inversely proportional to the plant’s growth, in that raising the concentration of the exogenous Asn to a concentration above 2 mM was observed to reduce various plant growth parameters, among which the root system in particular became thinner and darker (Figure 1f). While there were increases in fresh weights in plants treated with 0.5 and 1 mM Asn, the supply of 2 and 5 mM Asn recorded lower fresh weight than N0. Taken together, both N+ and Asn supplements had a profound impact on the morphology of the roots and shoots of poplar ‘Nanlin895’ seedlings, and the effect of Asn was dose-dependent. This is in accordance with findings reported by Haroun et al. (2010), which showed that the growth parameters of Phaseoulus vulgaris were induced by 1 mM Asn, but inhibited by higher concentrations of Asn (i.e., 2 mM, 3 mM, 4 mM, and 5 mM) [47]. Likewise, similar results were found by Jorkesh and Aminifard (2019) in garden cress [48].

Figure 1.
Phenotypical changes of exogenous asparagine (Asn) on poplar ‘Nanlin895’ growth. Plants were grown in (a) 3 mM KNO3 (N+), (b) N-free hydroponic solutions (N0), (c) 0.1 mM Asn, (d) 0.5 mM Asn, (e) 2 mM Asn, and (f) 5 mM Asn for two months.

Table 1.
Physiological effects of exogenous asparagine (Asn) on poplar ‘Nanlin895’ growth.
2.2. Asn Affects Photosynthesis of Poplar
As earlier indicated, N strongly influences the photosynthetic rate and is an essential indicator of plants’ material accumulation [55]. In agreement with this, adding N and Asn greatly affected poplar photosynthetic and chlorophyll fluorescence parameters (Table 2). Apart from 5 mM Asn, in all other Asn treatments, there were significant increases in stomatal conductance (gs), transpiration rate (Tr), and intercellular carbon dioxide concentration (Ci) compared with N0. However, there was an irregular trend in the changes in the photosynthetic rate (Pn), in which 1 mM Asn treatment recorded the highest Pn and 5 mM Asn recorded the lowest Pn across all the Asn treatments (Table 2). Nevertheless, the Pn, gs, Ci, and Tr values of poplar ‘Nanlin895’ seedlings grown under N+ conditions were remarkably higher than those of the Asn treatments.
Table 2.
Photosynthetic parameters of the poplar ‘Nanlin895’ under different asparagine (Asn) concentrations.
The potential maximum light energy conversion efficiency (Fv/Fm) represents the original light energy conversion efficiency of PSII [56,57]. In the current study, there was no significant difference across all the treatments, except for the 2 mM Asn treatment, which reduced the Fv/Fm compared to N+ (Table 3). The photosystem II activity as measured by Fv/Fo depicts the potential photosynthetic capacity in the plants, and its result was consistent with the change of the Fv/Fm. The nonphotochemical quenching coefficient (NPQ) represents one of the most important mechanisms protecting plants against photoinhibition [58]. As shown in Table 3, Asn-fed poplars recorded lower values of NPQ relative to N0 and N+ groups, indicating that Asn inactivates photoprotection mechanisms. Furthermore, the photochemical quenching coefficient (qP) and the actual photochemical quantum efficiency (ΦPSII) were remarkably lower in Asn-treated plants than that of the N0 and N+ control, suggesting that extra Asn limits photosynthetic capacity.
Table 3.
Chlorophyll fluorescence parameters of the poplar ‘Nanlin895’ under different asparagine (Asn) treatments.
2.3. Exogenous Asn Has a Significant Influence on the Internal N Status and NUE of Poplars
The relative in vivo intensity of N and C metabolism plays an essential role in plant growth and development. To ascertain the effect of Asn treatment on poplar metabolism, the content of nitrate, ammonia, total C, total N, C/N ratio, and N use efficiency (NUE) were determined. In general, nitrate content in the shoots was higher than in the roots and was increased as Asn applied concentrations rose (Figure 2a). Moreover, in the young leaves, the treatment of Asn remarkably increased nitrate content; thus, the N+ group recorded the lowest nitrate content, followed by the N0 control. However, the opposite trends were observed in the mature leaves: the N0 group recorded the highest nitrate content, while unpredictable differences were observed in the other treatments. In the stems, the N+ control had the highest nitrate content, whereas Asn treatment reduced nitrate content to the level of the N0 control.
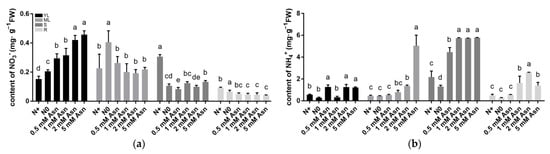
Figure 2.
The contents of nitrate (a) and ammonia (b) in various tissues of the poplar ‘Nanlin895’ under different asparagine (Asn) treatments. “N0” indicates N-free; “N+” represents the inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, 2 mM, or 5 mM, as indicated. YL, young leaves; ML, mature leaves; S, stems; R, roots. Bars indicate means ± SD (n ≥ 3), and different letters indicate significant differences (p < 0.05) based on Duncan’s test.
Nevertheless, Asn application concentration had an insignificant effect on nitrate content in the roots. Previously, it has been reported that the supply of high levels of Asn in Pisum sativum resulted in ammonia production [59]. In accordance with this, poplar ‘Nanlin895’-fed with Asn universally increased ammonium content in the stems, leaves, and roots (Figure 2b), indicating that exogenous Asn is indeed participating in the internal N metabolism process.
The effect of Asn treatment over changes in C and N metabolism in the shoots and roots of poplar ‘Nanlin895’ were shown in Figure 3. In the roots, plant N accumulation (i.e., total N content) under Asn treatment increased significantly compared to N0 and N+ control. In contrast, in the shoots except for 2 mM Asn, N accumulation was insignificantly affected by other concentrations of Asn treatment compared with the N+ control (Figure 3a). Somewhat differently, C accumulation (i.e., total C content) in both shoots and roots was higher in Asn-treated plants than the N+ control (Figure 3b), implying that Asn also confers C bonus to plants. The C/N as a whole showed an opposite trend to the plant total N content (Figure 3c), wherein the highest C/N was observed in the N0 control.
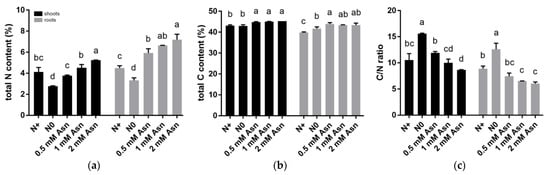
Figure 3.
Effects of exogenous asparagine (Asn) on the nitrogen (N) and carbon (C) contents in the shoots and roots of poplar ‘Nanlin895’. (a) total N content; (b) total C content; (c) the C/N ratio. “N0” indicates N-free reference; “N+” represents the inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, and 2 mM as indicated. Bars denote means ± SD (n ≥ 3), and different letters indicate significant differences (p < 0.05) based on Duncan’s test.
The unprecedented change in plant N content suggests that Asn treatment significantly impacts poplar N metabolism. To explicit this effect, the ability of poplar to utilize N for synthesizing biomass (NUtE) and to absorb N from the soil (NUpE) under different concentrations of Asn treatment was determined. As shown in Figure 4, among all the treatments, the highest NUtE was found in 0.5 mM Asn-fed poplars, which was approximately two times higher than the N+ control. However, as exogenous Asn concentration increased, NUtE dropped sharply. Treatment with 1 mM Asn and 2 mM Asn reduced NUtE to 10.4 and 3.8 g dry weight per gram N, respectively (Figure 4a). The changing trend of NUpE was similar to that of NUtE in poplar ‘Nanlin895’ seedling, in which treatment with 0.5 mM Asn recorded two times higher NUpE than the N+, whereas NUpE of the rest of the Asn treatment fell below the level of the N0 control (Figure 4b). Taken together, different concentrations of exogenous Asn significantly influenced poplar NUE, among which 0.5 mM Asn treatment recorded the highest NUE, whereas ascending concentration of Asn in poplar ‘Nanlin895’ relatively decreased NUE. Consistently, it has been demonstrated that higher NUE is observed in plants with low N supply [60].
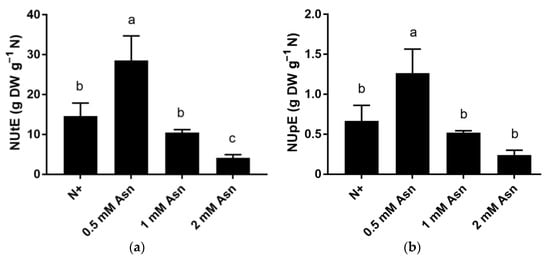
Figure 4.
Nitrogen utilization efficiency (NUtE) (a) and nitrogen uptake efficiency (NUpE) (b) of poplar ‘Nanlin895’ under different asparagine (Asn) treatment. “N0” indicates N-free; “N+” represents the inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, and 2 mM as indicated. Bars denote means ± SD (n ≥ 3), and different letters indicate significant differences (p < 0.05) based on Duncan’s test.
2.4. Effects of Exogenous Asn on the Contents of Amino Acids
The impact of exogenous Asn as a sole N source on the endogenous amino acids content in both roots and mature leaves of poplar ‘Nanlin895’ was shown in Figure 5. Generally, high amounts of Asn, arginine, glutamic acid, alanine, and aspartic acid but low contents of phosphoserine, glycine, methionine, and tyrosine were detected in all the samples. Varying the concentration of exogenous Asn dramatically affected the range of free amino acids in both roots and mature leaves of poplar ‘Nanlin895’. In brief, the content of the amino acids in the roots was only marginally affected by 0.5 mM Asn, but further increasing exogenous Asn dose augmented the content of a number of amino acids, especially the internal Asn, arginine, GABA, and alanine compared with the N+ control. Intriguingly, 5 mM Asn treatment led to a substantial accumulation of almost all amino acids, while the N0 control recorded their lowest level (Figure 5a). Overall, the application of Asn had similar effects on the amino acid content in the mature leaves compared to the roots, except that in the mature leaves the N+ control recorded the lowest level of all amino acids, while 0.5 mM Asn treatment enriched the contents of a proportion of the amino acids examined (Figure 5b).
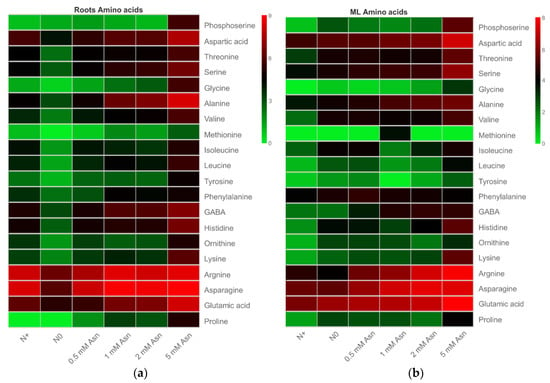
Figure 5.
Heatmap illustration of the amino acid composition of poplar ‘Nanlin895’ roots (a) and mature leaves (b) under different asparagine (Asn) treatments. “N0” indicates N-free; “N+” represents inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, 2 mM, and 5 mM as indicated. The color bar indicates low (green) expression levels to high (red).
2.5. Impacts of Asn Treatment on the Expression of Genes Involved in N and C Metabolism Pathways
The root system architecture is vital in regulating root foraging response to N fluctuation [61]. As aforementioned, the application of Asn significantly influenced poplar ‘Nanlin895’ root growth. To explore the underlying molecular basis, quantitative real-time RT-PCR (qPCR) was implemented to determine the expression levels of key genes involved in N and C metabolism pathways in the roots of poplar ‘Nanlin895’ fed with different concentrations of Asn. The results (Figure 6) showed substantial individual variability within different gene isoforms. In higher plants, glutamine synthetase (GS), the key enzyme engaged in N assimilation, is composed of two isoenzymes: the cytoplasmic GS1 (contained GS1.1, GS1.2, and GS1.3) and the chloroplast GS2 (encoded by a single gene GS2) [31]. Asn treatment significantly induced the expression level of GS1.1 and GS2 compared with both N0 and N+ control; however, the induction effect was negatively correlated with the Asn concentration applied. Similarly, the expression of GS1.3 was slightly induced by Asn except for a tentative depression by 2 mM Asn treatment. On the contrary, the expression level of GS1.2 was strongly inhibited by external Asn. Glutamate synthase (GOGAT) is another key enzyme involved in inorganic N assimilation. It has two forms in plants: ferredoxin-dependent (Fd-GOGAT) and NADH-dependent (NADH-GOGAT). Asn treatment concentration had a marked impact on the expression of both forms. While 0.5 mM Asn treatment dramatically increased Fd-GOGAT and NADH-GOGAT transcripts, 5 mM Asn treatment notably decreased them. Asparagine synthetase (AS) and asparaginase (ASPG) are the prime enzymes in plants participating in Asn synthesis and degradation, respectively [28]. The transcripts of AS1 and AS3 were positively induced by the external Asn, whereas the transcripts of AS2 were selectively induced by 0.5 mM Asn and 2 mM Asn (to a less extent) compared with the N+ control (Figure 6). Conversely, the expression of ASPG2 was repressed by 0.5 mM Asn but augmented when Asn concentration rose. Nevertheless, the expression levels of ASPG1 and ASPG3 were predominantly stimulated by 0.5 mM Asn but moderately changed among other Asn treatments, except for a decline under 2 mM Asn treatment. Likewise, nitrate reductase (NR) expression was significantly increased by different concentrations of Asn apart from 1 mM Asn. The nitrite reductase (NiR) transcriptional level was slightly influenced by Asn treatment compared with the N+ control. The cell wall invertase (CWINV) and the vacuole invertase (VI) are two sucrose breakdown enzymes important for primary carbon metabolism [62]. In contrast to the above N-related genes, the transcripts of CWINV1 were enhanced only by 0.5 mM Asn but suppressed by other concentrations of Asn treatment, while the transcripts of VI2 were strikingly induced by Asn treatment.
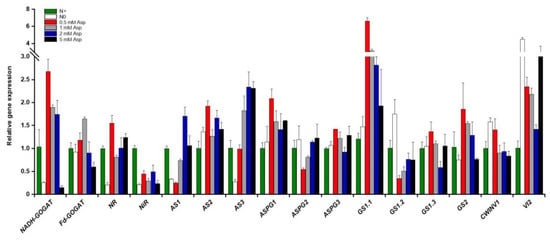
Figure 6.
Transcriptional changes of genes related to the N and C metabolism pathways in poplar ‘Nanlin895’ roots in response to asparagine (Asn) treatment. NR, nitrate reductase; GS, glutamine synthase; NADH-GOGAT, nicotinamide adenine dinucleotide-dependent glutamate synthase; Fd-GOGAT, ferredoxin-dependent glutamate synthase; AS, asparagine synthetase; ASPG, asparaginase; NiR, nitrite reductase; CWINV, cell wall apoplastic invertase; VI, vacuolar invertase. “N0” indicates N-free reference; “N+” represents the inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, 2 mM, and 5 mM as indicated.
2.6. Impacts of Asn Treatment on Enzymes Involved in N Metabolism
The enzymes involved in N metabolism pathways play fundamental roles during plant growth and development. To elucidate the biochemical effects of exogenous Asn at the enzyme level on poplar ‘Nanlin895’, the activities of key enzymes including GS, GOGAT, Aspartate aminotransferase (AspAT), and glutamate dehydrogenase (GDH) involved in N metabolism were analyzed in poplar roots. The AS and ASPG were omitted because reliable measurement of their enzyme activities was recalcitrant due to the presence of natural inhibitors and cross-reactions [63]. The enzyme activity assay results (Figure 7) showed that the application of Asn as a sole N source substantially altered the activities of the above enzymes. However, their regulatory responses varied with the change in Asn concentrations. While 0.5 mM Asn treatment significantly increased the activities of all four enzymes in poplar roots, 5 mM Asn treatment decreased their activities, though not to a marked degree compared with the N+ control. Among all of the four enzymes, GOGAT activity was most sensitive to Asn treatment, which was increased by more than 10 times (1 mM Asn treatment) than the N+ control, and its changing trend was consistent with the change in corresponding transcripts in 0.5 mM and 5 mM Asn-treated plants. In contrast, GS activity was less affected by Asn treatment, although strong inductions of GS1.1, GS1.3, and GS2 mRNA expression by 0.5 mM Asn and inhibition of GS2 by 5 mM Asn treatment were observed. The change in GDH activity was inversely correlated with Asn concentration, and the highest activity was found in 0.5 mM Asn-treated poplars, while the lowest activity was found in 5 mM Asn-treated poplars. Asn treatment increased AspAT activity; however, the change in AspAT activity was somewhat irrelevant to Asn treatment concentration.
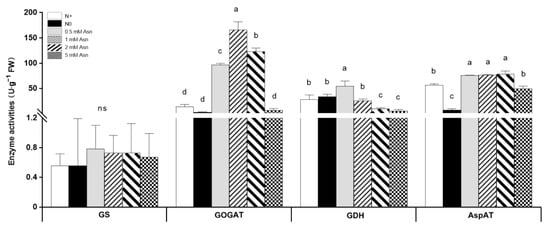
Figure 7.
Effects of exogenous asparagine (Asn) on the activities of enzymes involved in N metabolism pathways in poplar ‘Nanlin895’ roots. GS, glutamine synthase; GOGAT, glutamate synthase; GDH, glutamate dehydrogenase; AspAT, aspartate aminotransferase. “N0” indicates N-free; “N+” represents the inorganic N control. The concentration of Asn applied was 0.5 mM, 1 mM, 2 mM, and 5 mM. Bars indicate means ± SD (n ≥ 3), and different letters indicate significant differences (p < 0.05) based on Duncan’s test.
3. Discussion
Plant growth is correlated with increased synthetic demands, which require a constant supply of amino acids to support protein synthesis and matrix production. Amino acids are environmentally friendly molecules, and their uptake and utilization as an N source by plants is energetically more advantageous than inorganic N (including NO3− and NH4+ or N2) since it circumvents the mineralization pathway [14]. However, different types and concentrations of amino acids have a huge difference in their intake and utilization efficiency [17,54], which makes the effects of amino acids on plant growth vary. Asn serves as a principal amino acid for protein synthesis, transporting, and storing N in plant cells [64]. Asn-C is enriched in aspartate, malate, fumarate, and citrate, and Asn-N is enriched in aspartate, glutamate, proline, serine, and alanine [65]. Earlier works conducted in Arabidopsis and lettuce showed impressive growth results when Asn was used as a sole N source [66,67]. Moreover, the supply of Asn resulted in an improvement in relative water content, an increase in photosynthetic pigments, and a decrease in electrolyte leakage from cellular membranes, as well as a reduction in leaf proline content and activities of key oxidative relevant enzymes in salt-stressed maize plants [43]. Our prior study showed that exogenous glutamine as a single N source could promote poplar growth [53]. Given that Asn and glutamine are structurally similar since they both contain amide groups in their respective side chains, we therefore applied different concentration of Asn to poplar ‘Nanlin895’ to explore its effects on poplar biomass and its potential mode of action. In general, our study uncovers the nutritional effects and biochemical basis of exogenous Asn on poplars. It provides experimental evidence on the capacity of Asn to support poplar growth. Specifically, careful observation and comparison of plant phenotypes and physiological and biochemical analysis revealed that poplar ‘Nanlin895’ grew sturdily in Asn culture solution, which indirectly confirmed that Asn was indeed taken up and utilized by poplar. That notwithstanding, when Asn was added to the N-free nutrient solution, the N deficiency stress impacts on poplar (seen in N0 control) were relieved. Among all the concentrations of Asn applied, poplar ‘Nanlin895’ grew best at 0.5 mM Asn, as reflected by the seedlings’ height, biomass, and photosynthetic rate. Adding 1 mM Asn did not further improve poplar growth, but resulted in a low photosynthetic rate and a low conversion rate of maximized potential light energy. When exogenous Asn concentration was increased to the level of above 2 mM, it caused a high N stress response and the seedlings’ growth rate began to drop. This resultantly led to shortness in height, yellow and black roots, and a reduction in biomass and photosynthetic rate of poplar ‘Nanlin895’ seedlings. In conclusion, physiological reactions of poplar ‘Nanlin895’ seedlings to different levels of Asn were divergent based on parameters examined. Our study corroborates the dose-dependent effects of Asn on plants’ growth [47,48].
NUE basically depends on plants’ efficient uptake and use of N to increase biomass [68,69]. Earlier on, it had been demonstrated that the application of phenylalanine improved NUE in Populus X canescens [17], and the supply of glutamine (0.5 mM) increased NUE in poplar ‘Nanlin895’ [53]. Accordingly, we found that poplar ‘Nanlin895’ feeding with 0.5 mM Asn had higher NUtE and NUpE than the N+ control. Nevertheless, increased concentration of Asn was inversely related to the NUtE and NUpE ascribed to N wastage, which led to the overreading of N input and a decline in N utilization. By contrast, adding 0.5 mM Asn may sustain the optimum N/C ratio (Figure 3) and amino acid homeostasis for the effective absorption, utilization, and efficient N transformation within plants. Collectively, under the current experiment conditions, 0.5 mM Asn was the optimal treatment concentration for poplar ‘Nanlin895’ growth. Although Asn application is not essential for poplar, our results demonstrate that the combination of fertilization and biostimulation effect of Asn can be aligned with the effective use of conventional inorganic N.
It has been known that N content significantly affects the level of free amino acids in plants [70,71]. The feeding of Asn [U-14C] to rice seedlings resulted in the predominant labeling of glutamine, Asn, and glutamic acid in the shoots, as well as Asn and γ-Aminobutyric acid (GABA) in the roots [72]. Accordingly, in our study, the application of Asn as a sole N source in the hydroponic nutrient solution dramatically influenced the content of asparatic acid, glutamate, alanine, GABA, arginine, and endogenous Asn in both roots and mature leaves of poplar ‘Nanlin895’ (Figure 5). In particular, the contents of the amino acids Asn, arginine, and GABA, which are involved in environmental response [41,73,74], were profoundly enhanced in poplar, showing a positive changing trend in proportion to Asn application dose. This result indicates that the exogenous application of Asn is actually utilized by poplar, and it promotes the transformation of amino acids within the plant. However, a high negative correlation between amino acid accumulation (Figure 5) and root growth (i.e., root biomass and length) (Figure 1), along with an increase in Asn applied dose, was found. Meanwhile, it was also observed that the enzymes associated with N metabolism—GS, GOGAT, GDH, and AspAT—were increased by 0.5 mM Asn treatment but declined by 5 mM Asn. Previously, it was reported that the primary use of Asn in mammalian cells is in protein synthesis [75]. Consistently, a [14C] labeling study found that Asn was used for protein synthesis on a quantitatively significant scale in plants [76]. Given that the presence of Asn in large excess is a good marker of protein synthesis and breakdown [77], it is therefore reasonable to assume that the accumulation of amino acids observed in our study in poplar roots in response to high concentration of exogenous Asn (5 mM) is relevant to protein catabolism. In agreement with this possibility, it has been shown that Asn accumulation suppresses the well-studied eukaryotic translation initiation factor-2 alpha (eIF2α) kinase GCN (general control nonderepressible) and reduces the activating transcription factor 4 (ATF4), thereby impeding protein synthesis and cell growth, whereas Asn dissipation activates GCN and increases ATF4, and hence supports the translation of protein and maintains cell growth [75].
Overall, in our study, the root morphology of poplar ‘Nanlin895’ was greatly affected by Asn, implying that the root was the main site where Asn regulation took effect under the current experimental conditions. Our results indicate that Asn may function not only as a metabolism but also as a signaling/regulatory candidate as a part of the plant adaptation process, activating multiple responses related to N assimilation [39]. The possible mode of action of Asn is illustrated in Figure 8. It can be proposed that under high doses of Asn treatment, protein degradation rather than protein synthesis occurs in poplar ‘Nanlin895’; that is, an overdose of Asn (5 mM) disrupts protein synthesis probably through the GCN2/ATF4 pathway, followed by amino acid accumulation, a reduction in enzyme activity of GDH and GOGAT, and in turn, poplar growth (root length and biomass in particular) depression, whereas the application of an appropriate level of Asn (i.e., 0.5 mM) may facilitate nascent protein synthesis as a result of poplar growth promotion. As the transcriptional change of GCN2/ATF4 pathway genes was only marginal by qPCR analysis, we deduce that Asn regulation occurs at the post-translation level, and phosphorylation of eukaryotic translation initiation factors is likely engaged in this process [75,76,78].
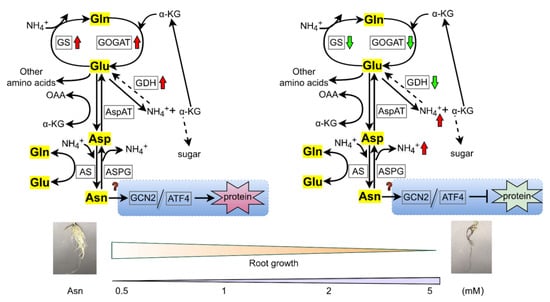
Figure 8.
Schematic model of Asn dose-dependent effects on poplar roots. Gln, glutamine; Glu, glutamic acid; Asp, aspartic acid; α-KG, α-ketoglutarate. GS, glutamine synthase; GOGAT, glutamate synthase; AS, asparagine synthetase; ASPG, asparaginase; GDH, glutamate dehydrogenase; AspAT, aspartate aminotransferase; GCN, general control nonderepressible; ATF, activating transcription factor. The up arrow indicates induction and the down arrow indicates depression. The box stands for the enzyme.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Plantlets of poplar ‘Nanlin895’ (Populus deltoides × P. euramericana) clones that had been tissue-cultured in 1/2 MS medium for one month were used throughout the experiment, unless otherwise indicated. All seedlings were initially kept in pure water to get rid of internal N for 10 days, then treated with inorganic-N (KNO3) or various concentrations of Asn-N in the N-free 1/2 MS nutrient solution. There were six groups of treatments in total: the standard inorganic-N control (3 mM KNO3; N+), the N-free control (0 mM N; N0), 0.5 mM Asn, 1 mM Asn, 2 mM Asn, and 5 mM Asn. The nutrient solution was refreshed twice a week for two months. Poplar seedlings were kept in disposable plastic water cups for the hydroponic treatment, one seedling per cup. Each cup contained 250 mL nutrient solution fixed with a black foam board. Each treatment included at least six individual plants. The whole experiment was repeated three times. The seedlings were grown under 24 °C light/22 °C dark, 5000 Lux light intensity for 16 hours with 40–60% humidity. At the end of the treatment, tissues from young leaves (YL), mature leaves (ML), stems (S), and roots (R) were collected, snap-frozen in liquid nitrogen, and stored at −80 °C for further analysis.
4.2. Analysis of Growth, Gas Exchange, and Chlorophyll Fluorescence Characteristics
Growth parameters including plant height, root length, and fresh shoot weight, as well as fresh root weight, were measured before harvest. LI-6400 Portable photosynthesis system (LI-COR, Lincoln, NE, USA) was used to measure the net photosynthetic rate (Pn, μmol CO2·m−2·s−1), intercellular carbon dioxide concentration (Ci, μmol·mol−1), stomatal conductance (Gs, mol H2O·m−2·s−1), and transpiration rate (Tr, mmol H2O·m−2·s−1). During the measurement, environmental conditions were set at 800 μmol·m−2·s−1 of light intensity and 400 μL·L−1 of carbon dioxide. The nonphotochemical quenching coefficient (NPQ), photochemical quenching coefficient (qP), the actual photochemical quantum efficiency of PSII (ΦPSII), the potential maximum light energy conversion efficiency (Fv/Fm), and photosystem II activity (Fv/Fo) were measured using chlorophyll fluorescence imaging system (CF Imager, Nanjing, China).
4.3. Determination of Activities of Enzymes Involved in N Metabolism
The activities of key enzymes—including nitrate reductase (NR, EC 1.6.6.1), glutamine synthetase (GS, EC 6.3.1.2), glutamate synthase (GOGAT, EC 1.4.7.1), aspartate aminotransferase (AspAT, EC 2.6.1.1), and glutamate dehydrogenase (GDH, EC 1.4.1.2)—that are involved in plant N metabolism pathways [68] were analyzed in poplar ‘Nanlin895’ leaves, stems, and shoots following methods described in earlier studies [69,79,80].
4.4. Measurement of the Content of Free Amino Acids, NH4+, NO3− and NUE
The free amino acids were extracted by sulfosalicylic acid according to previous methods [81], and the content of free amino acids was determined by the automatic amino acid analyzer (S-433D, Sykam GmbH, Eresing, Germany) following the operation instruction. The ammonium and nitrate content was determined by the modified ninhydrin colorimetric criteria [82] and Patterson and colleagues’ method [83], respectively. Nitrogen use efficiency was calculated according to previously reported equation: NUtE = biomass DW/N supply, NUpE = total nitrogen content/N supply [84].
4.5. Analysis of Transcript Levels of Genes Involved in N Metabolism
Quantitative RT-PCR was employed to analyze the expression levels of nitrate reductase (NR), nitrite reductase (NiR), asparagine synthetase (AS), asparaginase (ASPG), glutamine synthase (GS), glutamate synthase (GOGAT), cell wall apoplastic invertase (CWINV1), and vacuolar invertase (VI2) genes in the leaves, stem, and roots of poplar. Total RNA was extracted by using RNAprep Pure Polysaccharide Polyphenol Plant Total RNA Extraction Kit (TIANGEN, Beijing, China). The purity and concentration of the extracted RNA were checked by the Microvolume Spectrophotometer (Colibri LB 915, Bad Wildbad, Germany) and then verified through agarose gel electrophoresis. Reverse transcription kit (PrimeScriptTM RT reagent Kit with gDNA Eraser, Takara, Japan) was used to synthesize the first strand of cDNA. Gene-specific primers (Supplement Table S1) were designed by Primer3 (http://primer3.ut.ee/, accessed on 21 February 2022). Quantification of gene amplification was performed on the real-time fluorescent quantitative PCR instrument (Applied Biosystems StepOneTM, Beijing, China) with the fluorescent agent AG SYBR Green Pro Taq HS premixed qPCR kit (Accurate Biology, Changsha, China).
4.6. Statistical Analysis of Data
Analysis of variance (ANOVA) was performed for all data using IBM SPSS Statistics 21 statistical software. The standard errors were based on the pooled error term from the ANOVA table. Differences were considered statistically significant when the p-value of the ANOVA F-test was less than 0.05.
5. Conclusions
Asn is a multipurpose amino acid participating in various biological processes, including plant development and adaptation to environmental stresses. Our study found that the application of exogenous Asn significantly increased the biomass of poplar ‘Nanlin895’ compared to the N-free control by improving the growth status of N starvation. To the best of our knowledge, this is the first research of exogenous application of Asn as a sole N source conducted on poplars. It reveals that Asn, when supplied at an appropriate rate, is conducive to the growth fitness of poplar. Although there was no match for the growth promotion effect of Asn as against the N+ control, it was, however, clear that relative to the N-free plants, application of Asn dramatically alleviated poplar ‘Nanlin895’ seedlings from N deficiency stress. In consistency with previous findings, it can be concluded that the concentrations and forms of N influence poplar growth and performance, including biomass partitioning and root morphology such as root length and root biomass [85].
Our research provides essential information on Asn for effective biomass allocation in poplars. It is reasonable to infer that (1) poplars are competent at using Asn as a sole N source to support their growth, as indicated by the growth parameters and photosynthesis activities of poplar ‘Nanlin895’ plantlets fed with Asn; (2) the feeding of Asn at a proper concentration leads to root morphology alteration, as a result of increasing/decreasing nutrient and possibly water absorption; (3) the exogenous application of Asn induces the transcript levels of key genes and activity of enzymes involved in N and C metabolism pathway, promotes ammonium assimilation, mitigates N metabolic disturbance resulting from N stress, and advances N absorption and utilization in both roots and the above-ground tissues; (4) Asn might play a critical role in regulating protein homeostasis, and in turn, plant growth and stress response.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232113126/s1.
Author Contributions
Conceptualization, T.S. and M.H.; formal analysis, S.W.; investigation, S.W., L.W., J.F., Y.S. and X.L.; writing—original draft preparation, S.W. and M.H.; writing—review and editing, M.H. and T.S.; supervision, M.H. and T.S.; project administration, M.H. and T.S.; funding acquisition, M.H. and T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (NSFC), grant number 31870589; 31700525.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the NSFC for funding this work and the Co-Innovation Center for Sustainable Forestry in Southern China and PAPD for the instrument use.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tegeder, M.; Masclaux-Daubresse, C. Source and Sink Mechanisms of Nitrogen Transport and Use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.-L.; Li, Q.; Zeng, X.-P.; Liu, Y.; Li, Y.-R. Fate of Nitrogen in Agriculture and Environment: Agronomic, Eco-Physiological and Molecular Approaches to Improve Nitrogen Use Efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef]
- DesRochers, A.; van den Driessche, R.; Thomas, B.R. The Interaction between Nitrogen Source, Soil PH, and Drought in the Growth and Physiology of Three Poplar Clones. Can. J. Bot. 2007, 85, 1046–1057. [Google Scholar] [CrossRef]
- Jämtgård, S.; Näsholm, T.; Huss-danell, K. Characteristics of Amino Acid Uptake in Barley. Plant Soil 2008, 302, 221–231. [Google Scholar] [CrossRef]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of Organic Nitrogen by Plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef]
- Franklin, O.; Cambui, C.A.; Gruffman, L.; Palmroth, S.; Oren, R.; Näsholm, T. The Carbon Bonus of Organic Nitrogen Enhances Nitrogen Use Efficiency of Plants. Plant. Cell Environ. 2017, 40, 25–35. [Google Scholar] [CrossRef]
- Schmidt, S.; Näsholm, T.; Rentsch, D. Organic Nitrogen. New Phytol. 2014, 203, 29–31. [Google Scholar] [CrossRef]
- Jones, D.L.; Shannon, D.; Junvee-Fortune, T.; Farrar, J.F. Plant Capture of Free Amino Acids Is Maximized under High Soil Amino Acid Concentrations. Soil Biol. Biochem. 2005, 37, 179–181. [Google Scholar] [CrossRef]
- Owen, A.; Jones, D. Competition for Amino Acids between Wheat Roots and Rhizosphere Microorganisms and the Role of Amino Acids in Plant N Acquisition. Soil Biol. Biochem. 2001, 33, 651–657. [Google Scholar] [CrossRef]
- Gioseffi, E.; De Neergaard, A.; Schjoerring, J.K. Interactions between Uptake of Amino Acids and Inorganic Nitrogen in Wheat Plants. Biogeosciences 2012, 9, 1509–1518. [Google Scholar] [CrossRef]
- Ge, T.; Song, S.; Roberts, P.; Jones, D.L.; Huang, D.; Iwasaki, K. Amino Acids as a Nitrogen Source for Tomato Seedlings: The Use of Dual-Labeled (13C, 15N) Glycine to Test for Direct Uptake by Tomato Seedlings. Environ. Exp. Bot. 2009, 66, 357–361. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Lonhienne, T.G.A.; Rentsch, D.; Robinson, N.; Christie, M.; Webb, R.I.; Gamage, H.K.; Carroll, B.J.; Schenk, P.M.; Schmidt, S. Plants Can Use Protein as a Nitrogen Source without Assistance from Other Organisms. Proc. Natl. Acad. Sci. USA 2008, 105, 4524–4529. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, G.; Jaskani, M.J.; Sajjad, Y.; Akram, A. Effect of Antioxidants, Amino Acids and Plant Growth Regulators on in Vitro Propagation of Rosa Centifolia. Iran. J. Biotechnol. 2016, 14, 51–55. [Google Scholar] [CrossRef]
- Jones, D.L.; Kielland, K. Soil Amino Acid Turnover Dominates the Nitrogen flux in Permafrost-Dominated Taiga Forest Soils. Soil Biol. Biochem. 2002, 34, 209–219. [Google Scholar] [CrossRef]
- Dolev, N.; Katz, Z.; Ludmer, Z.; Ullmann, A.; Brauner, N.; Goikhman, R. Natural Amino Acids as Potential Chelators for Soil Remediation. Environ. Res. 2020, 183, 109140. [Google Scholar] [CrossRef] [PubMed]
- Garc, F.; Sim, S.; Navarro-p, V.; Alfosea-sim, M. Scientific Advances in Biostimulation Reported in the 5th Biostimulant World Congress. Horticulturae 2022, 8, 665. [Google Scholar]
- Jiao, Y.; Chen, Y.; Ma, C.; Qin, J.; Nguyen, T.H.N.; Liu, D.; Gan, H.; Ding, S.; Luo, Z.-B. Phenylalanine as a Nitrogen Source Induces Root Growth and Nitrogen-Use Efficiency in Populus × Canescens. Tree Physiol. 2018, 38, 66–82. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative Physiological and Metabolic Analysis Reveals a Complex Mechanism Involved in Drought Tolerance in Chickpea (Cicer arietinum L.) Induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Kang, T.; Wu, H.D.; Lu, B.Y.; Luo, X.J.; Gong, C.M.; Bai, J. Low Concentrations of Glycine Inhibit Photorespiration and Enhance the Net Rate of Photosynthesis in Caragana Korshinskii. Photosynthetica 2018, 56, 512–519. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Zaheer Akbar, M.; Shakoor, M.B.; Mahmood, A.; Ishaque, W.; Hussain, A. Foliar Application of Aspartic Acid Lowers Cadmium Uptake and Cd-Induced Oxidative Stress in Rice under Cd Stress. Environ. Sci. Pollut. Res. 2017, 24, 21938–21947. [Google Scholar] [CrossRef]
- Sadak, M.S.H.; Abdelhamid, M.T.; Schmidhalter, U. Effect of Foliar Application of Amino Acids on Plant Yield and Some Physiological Parameters in Bean Palnts Irrigated with Seawater. Acta Biol. Colomb. 2015, 20, 141–152. [Google Scholar]
- Geshnizjani, N.; Khosh-khui, M. Promoted Growth and Improved Quality of Gerbera Jamesonni L. Flowers Using Exogenous Application of Amino Acids. Int. J. Hortic. Sci. Technol. 2016, 3, 155–166. [Google Scholar] [CrossRef]
- Akladious, S.A.; Abbas, S.M. Alleviation of Sea Water Stress on Tomato Plants by Foliar Application of Aspartic Acid and Glutathione. Bangladesh J. Bot. 2013, 42, 31–43. [Google Scholar] [CrossRef]
- Abd El-samad, H.M.; Shaddad, M.A.K.; Barakat, N. Improvement of Plants Salt Tolerance by Exogenous Application of Amino Acids. J. Med. Plants Res. 2011, 5, 5692–5699. [Google Scholar]
- Kan, C.; Chung, T.; Juo, Y.; Hsieh, M. Glutamine Rapidly Induces the Expression of Key Transcription Factor Genes Involved in Nitrogen and Stress Responses in Rice Roots. BMC Genomics 2015, 16, 731. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Islam, M.M.; Wang, R.; Guo, J.; Luo, H.; Chen, F.; Li, X. Glutamine Application Promotes Nitrogen and Biomass Accumulation in the Shoot of Seedlings of the Maize Hybrid ZD958. Planta 2020, 251, 66. [Google Scholar] [CrossRef]
- Ali, Q.; Athar, H.-R.; Haider, M.Z.; Shahid, S.; Aslam, N.; Shehzad, F.; Naseem, J.; Ashraf, R.; Ali, A.; Hussain, S.M. Role of Amino Acids in Improving Abiotic Stress Tolerance to Plants. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 175–204. ISBN 9780203705315. [Google Scholar]
- Brouquisse, R.; James, F.; Pradet, A.; Raymond, P. Asparagine Metabolism and Nitrogen Distribution during Protein Degradation in Sugar-Starved Maize Root Tips. Planta 1992, 188, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological Functions of Asparagine Synthetase in Plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Mansfeld, J.; Gebauer, S.; Dathe, K.; Ulbrich-Hofmann, R. Secretory Phospholipase A2 from Arabidopsis Thaliana: Insights into the Three-Dimensional Structure and the Amino Acids Involved in Catalysis. Biochemistry 2006, 45, 5687–5694. [Google Scholar] [CrossRef]
- Gaufichon, L.; Rothstein, S.J.; Suzuki, A. Asparagine Metabolic Pathways in Arabidopsis. Plant Cell Physiol. 2016, 57, 675–689. [Google Scholar] [CrossRef]
- Coruzzi, G.M. Primary N-Assimilation into Amino Acids in Arabidopsis. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2003; Volume 2, p. e0010. ISBN 1543-8120. [Google Scholar]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in Plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Pant, A.; Yang, Z. Asparagine: An Achilles Heel of Virus Replication? ACS Infect. Dis. 2020, 6, 2301–2303. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Pavlova, N.N.; Zhang, J. Asparagine, a Critical Limiting Metabolite during Glutamine Starvation. Mol. Cell. Oncol. 2018, 5, e1441633. [Google Scholar] [CrossRef] [PubMed]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine Promotes Cancer Cell Proliferation through Use as an Amino Acid Exchange Factor. Nat. Commun. 2016, 7, 11457. [Google Scholar] [CrossRef] [PubMed]
- Aidar, M.P.M.; Schmidt, S.; Moss, G.; Stewart, G.R.; Joly, C.A. Nitrogen Use Strategies of Neotropical Rainforest Trees in Threatened Atlantic Forest. Plant Cell Environ. 2003, 26, 389–399. [Google Scholar] [CrossRef]
- Lin, J.F.; Wu, S.H. Molecular Events in Senescing Arabidopsis Leaves. Plant J. 2004, 39, 612–628. [Google Scholar] [CrossRef]
- Sulieman, S.; Tran, L.S.P. Asparagine: An Amide of Particular Distinction in the Regulation of Symbiotic Nitrogen Fixation of Legumes. Crit. Rev. Biotechnol. 2013, 33, 309–327. [Google Scholar] [CrossRef]
- Rashmi, D.; Barvkar, V.T.; Nadaf, A.; Mundhe, S.; Kadoo, N.Y. Integrative Omics Analysis in Pandanus Odorifer (Forssk.) Kuntze Reveals the Role of Asparagine Synthetase in Salinity Tolerance. Sci. Rep. 2019, 9, 932. [Google Scholar] [CrossRef]
- Oddy, J.; Raffan, S.; Wilkinson, M.D.; Elmore, J.S.; Halford, N.G. Stress, Nutrients and Genotype: Understanding and Managing Asparagine Accumulation in Wheat Grain. CABI Agric. Biosci. 2020, 1, 10. [Google Scholar] [CrossRef]
- Hwang, I.S.; An, S.H.; Hwang, B.K. Pepper Asparagine Synthetase 1 (CaAS1) Is Required for Plant Nitrogen Assimilation and Defense Responses to Microbial Pathogens. Plant J. 2011, 67, 749–762. [Google Scholar] [CrossRef]
- Kaya, C.; Aydemir, S.; Sonmez, O.; Ashraf, M.; Dikilitas, M. Regulation of Growth and Some Key Physiological Processes in Salt-Stressed Maize (Zea mays L.) Plants by Exogenous Application of Asparagine and Glycerol. Acta Bot. Croat. 2013, 72, 157–168. [Google Scholar] [CrossRef]
- Ivanov, A.; Kameka, A.; Pajak, A.; Bruneau, L.; Beyaert, R.; Hernández-Sebastià, C.; Marsolais, F. Arabidopsis Mutants Lacking Asparaginases Develop Normally but Exhibit Enhanced Root Inhibition by Exogenous Asparagine. Amino Acids 2012, 42, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, V.; Chatzissavvidis, C.; Dimassi-Theriou, K.; Therios, I. Effect of Asparagine, Cysteine, Citrulline, and Glutamine on in Vitro Rooting and Biochemical Constituents in Cherry Rootstocks. Biol. Plant. 2016, 60, 1–12. [Google Scholar] [CrossRef]
- Sarker, K.; Kabir, A.; Sharmin, S.; Nasrin, Z.; Alam, M. Improved Somatic Embryogenesis Using L-Asparagine in Wheat (Triticum aestivum L.). Sjemenarstvo 2007, 24, 187–196. [Google Scholar]
- Haroun, S.A.; Shukry, W.M.; El-Sawy, O. Effect of Asparagine or Glutamine on Growth and Metabolic Changes in Phaseolus Vulgaris under in Vitro Conditions. Biosci. Res. 2010, 7, 1–21. [Google Scholar]
- Jorkesh, A.; Aminifard, M.H. Foliar Application of Asparagine and Casein on Biochemical and Morphological Attributes of Garden Cress (Lepidium sativum L.) under Greenhouse Conditions. Adv. Hortic. Sci. 2019, 33, 227–233. [Google Scholar] [CrossRef]
- Moigne, M.L.; Guérin, V.; Furet, P.; Billard, V.; Lebrec, A.; Roman, H.; Citerne, S.; Morvan-bertrand, A.; Limami, A.; Vian, A.; et al. Asparagine and Sugars Are Both Required to Sustain Secondary Axis Elongation after Bud Outgrowth in Rosa Hybrida. J. Plant Physiol. 2018, 222, 17–27. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Martin, T.A.; Davis, J.M. Short-Term Physiological and Developmental Responses to Nitrogen Availability in Hybrid Poplar. New Phytol. 2005, 167, 41–52. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.; Masclaux-Daubresse, C.; Wang, N.; Wang, H.; Zheng, B. Morphological and Physiological Responses to Contrasting Nitrogen Regimes in Populus Cathayana Is Linked to Resources Allocation and Carbon/Nitrogen Partition. Environ. Exp. Bot. 2019, 162, 247–255. [Google Scholar] [CrossRef]
- Qu, C.; Hao, B.; Xu, X.; Wang, Y.; Yang, C.; Xu, Z.; Liu, G. Functional Research on Three Presumed Asparagine Synthetase Family Members in Poplar. Genes 2019, 10, 326. [Google Scholar] [CrossRef]
- Han, M.; Xu, M.; Wang, S.; Wu, L.; Sun, S.; Su, T. Effects of Exogenous L-Glutamine as a Sole Nitrogen Source on Physiological Characteristics and Nitrogen Use Efficiency of Poplar. Plant Physiol. Biochem. 2022, 172, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Berthrong, S.T. The Uptake of Amino Acids by Microbes and Trees in Three Cold-Temperate Forests. Ecology 2005, 86, 3345–3353. [Google Scholar] [CrossRef]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen Supply Influences Photosynthesis Establishment along the Sugarcane Leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Lin, H.; Gorbunov, M.Y. What Limits Photosynthetic Energy Conversion Efficiency in Nature? Lessons from the Oceans. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160376. [Google Scholar] [CrossRef]
- Zhu, X.-G.; Long, S.P.; Ort, D.R. What Is the Maximum Efficiency with Which Photosynthesis Can Convert Solar Energy into Biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef]
- Ireland, R.J.; Joy, K.W. Two Routes for Asparagine Metabolism in Pisum sativum L. Planta 1981, 151, 289–292. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Devi, S.; Buckseth, T.; Ali, N.; Singh, R.K.; Zinta, R.; Dua, V.K.; Chakrabarti, S.K. Precision Phenotyping of Contrasting Potato (Solanum tuberosum L.) Varieties in a Novel Aeroponics System for Improving Nitrogen Use Efficiency: In Search of Key Traits and Genes. J. Integr. Agric. 2020, 19, 51–61. [Google Scholar] [CrossRef]
- Pandey, A.; Devi, L.L.; Singh, A.P. Review: Emerging Roles of Brassinosteroid in Nutrient Foraging. Plant Sci. 2020, 296, 110474. [Google Scholar] [CrossRef]
- Wan, H.; Wu, L.; Yang, Y.; Zhou, G.; Ruan, Y.-L. Evolution of Sucrose Metabolism: The Dichotomy of Invertases and Beyond. Trends Plant Sci. 2018, 23, 163–177. [Google Scholar] [CrossRef]
- Xu, H.; Curtis, T.Y.; Powers, S.J.; Raffan, S.; Gao, R.; Huang, J.; Heiner, M.; Gilbert, D.R.; Halford, N.G. Genomic, Biochemical, and Modeling Analyses of Asparagine Synthetases from Wheat. Front. Plant Sci. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Ta, T.C.; Joy, K.W.; Ireland, R. Role of Asparagine in the Photochemistry Nitrogen Metaboliem of Pea Leaves. Plant Physiol. 1985, 78, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Yu, Y.; Shen, L.; Zhang, G.-F.; Karner, C.M. SLC1A5 Provides Glutamine and Asparagine Necessary for Bone Development in Mice. Elife 2021, 10, e71595. [Google Scholar] [CrossRef] [PubMed]
- Forsum, O.; Svennerstam, H.; Ganeteg, U. Capacities and Constraints of Amino Acid Utilization in Arabidopsis. New Phytol. 2008, 179, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Mills, S. Potential of Exogenous L-Amino Acids in Salinity Stress Alleviation during Germination and Early Post-Germinative Seedling Growth of Lactuca sativa L.; University of Canterbury: Christchurch, New Zealand, 2014. [Google Scholar]
- Yoneyama, T.; Suzuki, A. Light-Independent Nitrogen Assimilation in Plant Leaves: Nitrate Incorporation into Glutamine, Glutamate, Aspartate, and Asparagine Traced By15n. Plants 2020, 9, 1303. [Google Scholar] [CrossRef]
- Ehlting, B.; Dluzniewska, P.; Dietrich, H.; Selle, A.; Teuber, M.; Hansch, R.; Nehls, U.; Polle, A.; Schnitzler, J.-P.; Rennenberg, H.; et al. Interaction of Nitrogen Nutrition and Salinity in Grey Poplar (Populus tremula × alba). Plant. Cell Environ. 2007, 30, 796–811. [Google Scholar] [CrossRef]
- Teixeira, W.F.; Fagan, E.B.; Soares, L.H.; Soares, J.N.; Reichardt, K.; Neto, D.D. Seed and Foliar Application of Amino Acids Improve Variables of Nitrogen Metabolism and Productivity in Soybean Crop. Front. Plant Sci. 2018, 9, 396. [Google Scholar] [CrossRef]
- Postles, J.; Curtis, T.Y.; Powers, S.J.; Elmore, J.S.; Mottram, D.S.; Halford, N.G. Changes in Free Amino Acid Concentration in Rye Grain in Response to Nitrogen and Sulfur Availability, and Expression Analysis of Genes Involved in Asparagine Metabolism. Front. Plant Sci. 2016, 7, 917. [Google Scholar] [CrossRef]
- Kanamori, T.; Matsumoto, H. Asparagine Biosynthesis by Oryza Sativa Seedlings. Phytochemistry 1974, 13, 1407–1412. [Google Scholar] [CrossRef]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological Implications of Arginine Metabolism in Plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef]
- Bown, A.W.; Shelp, B.J. Plant GABA: Not Just a Metabolite. Trends Plant Sci. 2016, 21, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, J.; Venneti, S.; Cross, J.R.; Takagi, T.; Bhinder, B.; Djaballah, H.; Kanai, M.; Cheng, E.H.; Judkins, A.R.; et al. Asparagine Plays a Critical Role in Regulating Cellular Adaptation to Glutamine Depletion. Mol. Cell 2014, 56, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Batra, S.; Zhang, J. Asparagine: A Metabolite to Be Targeted in Cancers. Metabolites 2021, 11, 402. [Google Scholar] [CrossRef]
- Genix, P.; Bligny, R.; Martin, J.B.; Douce, R. Transient Accumulation of Asparagine in Sycamore Cells after a Long Period of Sucrose Starvation. Plant Physiol. 1990, 94, 717–722. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Kanyuka, K.; Parry, M.A.J.; Powers, S.J.; Halford, N.G. GCN2-Dependent Phosphorylation of Eukaryotic Translation Initiation Factor-2α in Arabidopsis. J. Exp. Bot. 2008, 59, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Disturbed Ammonium Assimilation Is Associated with Growth Inhibition of Roots in Rice Seedlings Caused by NaCl. Plant Growth Regul. 1996, 18, 233–238. [Google Scholar] [CrossRef]
- Gan, H.; Jiao, Y.; Jia, J.; Wang, X.; Li, H.; Shi, W.; Peng, C.; Polle, A.; Luo, Z. Bin Phosphorus and Nitrogen Physiology of Two Contrasting Poplar Genotypes When Exposed to Phosphorus and/or Nitrogen Starvation. Tree Physiol. 2015, 36, 22–38. [Google Scholar] [CrossRef]
- Liao, Y.; Cui, R.; Yuan, T.; Xie, Y.; Gao, Y. Cysteine and Methionine Contribute Differentially to Regulate Alternative Oxidase in Leaves of Poplar (Populus Deltoides x Populus Euramericana ‘Nanlin 895’) Seedlings Exposed to Different Salinity. J. Plant Physiol. 2019, 240, 153017. [Google Scholar] [CrossRef]
- Ying, X. Determination of Trace Ammonia Content in Water by Ninhydrin Colorinetry. Chem. Fertil. Ind. 2011, 38, 27–30. [Google Scholar]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct Signalling Pathways and Transcriptome Response Signatures Differentiate Ammonium- and Nitrate-Supplied Plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can Less Yield More? Is Reducing Nutrient Input into the Environment Compatible with Maintaining Crop Production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Lonhienne, T.G.A.; Trusov, Y.; Young, A.; Rentsch, D.; Näsholm, T.; Schmidt, S.; Paungfoo-Lonhienne, C. Effects of Externally Supplied Protein on Root Morphology and Biomass Allocation in Arabidopsis. Sci. Rep. 2014, 4, 5505. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).