Abstract
The hyperosmolality-gated calcium-permeable channel gene family (OSCA) is one kind of conserved osmosensors, playing a crucial role in maintaining ion and water homeostasis and protecting cellular stability from the damage of hypertonic stress. Although it has been systematically characterized in diverse plants, it is necessary to explore the role of the OSCA family in barley, especially its importance in regulating abiotic stress response. In this study, a total of 13 OSCA genes (HvOSCAs) were identified in barley through an in silico genome search method, which were clustered into 4 clades based on phylogenetic relationships with members in the same clade showing similar protein structures and conserved motif compositions. These HvOSCAs had many cis-regulatory elements related to various abiotic stress, such as MBS and ARE, indicating their potential roles in abiotic stress regulation. Furthermore, their expression patterns were systematically detected under diverse stresses using RNA-seq data and qRT-PCR methods. All of these 13 HvOSCAs were significantly induced by drought, cold, salt and ABA treatment, demonstrating their functions in osmotic regulation. Finally, the genetic variations of the HvOSCAs were investigated using the re-sequencing data, and their nucleotide diversity in wild barley and landrace populations were 0.4966 × 10−3 and 0.391 × 10−3, respectively, indicating that a genetic bottleneck has occurred in the OSCA family during the barley evolution process. This study evaluated the genomic organization, evolutionary relationship and genetic expression of the OSCA family in barley, which not only provides potential candidates for further functional genomic study, but also contributes to genetically improving stress tolerance in barley and other crops.
1. Introduction
As sessile organisms, plants usually have to suffer from the impact of adverse abiotic stresses, such as drought, salinity and extreme high or low temperature [1,2], which not only determine the geographical adaptability of plants, but also seriously limits agricultural productivity worldwide [3]. To cope with these stresses, plants have evolved complicated and interconnected mechanisms to protect them from the impacts of harsh environments in a gradual manner [4]. Generally, plants can perceive and transduce extracellular stress signals into intracellular second messengers, which are able to trigger a serial of signal cascades to regulate the given metabolic and physiological processes through activating or inhibiting the expression of some specific genes, resulting in cellular homeostasis and the reprograming of growth to increase stress resistance or adapt to stressed conditions [5,6].
Calcium ions (Ca2+) mainly serve as crucial secondary messengers, playing a pivotal role in signaling perception and transduction under biotic and abiotic stresses, especially responding to osmotic change. Previous studies have revealed that calcium channels could act as osmosensors across unicellular prokaryotic bacteria to multicellular eukaryotic mammals [7,8,9,10,11]. In plants, several calcium channels have been characterized to find that they not only function to mediate the response to abiotic and biotic stresses, but are also involved in growth regulation and biosynthesis [12,13,14,15,16,17]. Among them, the hyperosmolality-gated calcium-permeable channel family (OSCA) is one of the most important and well-studied calcium channels participating in osmotic adjustment [18,19]. In light of its significance, the OSCA family has been identified in many plant species, including 15 OSCA members found in A. thaliana, 11 in rice [18], 42 in wheat [19], 12 in maize [20,21], 13 in mung bean [22], as well as 35 in cotton [23]. The expression profile of OSCA genes were also systematically investigated under diverse abiotic stresses in different plant species, which demonstrated their differential roles in stress response and tolerance [18,19,20,21,22,23]. Some plant OSCA genes were also functionally validated. In A. thaliana, AtOSCA1.3 was phosphorylated by the immune receptor-associated cytosolic kinase BIK1 by treatment with pathogen-associated molecular patterns (PAMP) fg22, and showed an increase in channel activity, which functioned as a switch to control stomatal closure in the immune signaling pathway [24]. AtOSCA1 was found to act as an osmosensor through a calcium imaging-based genetic screen approach, which was a plasma-membrane located protein with a low hyperosmolality-induced Ca2+ increase (OICI) [25]. In maize, ZmOSCA2.3 and ZmOSCA2.4 were up-regulated under ABA, PEG and NaCl treatments and over-expression of ZmOSCA2.4 could increase the expression of drought tolerance related genes while decreasing the expression of senescence-related genes in A. thaliana, respectively [21]. The expression profile analysis of ZmOSCAs in 15 tissues found that ZmOSCA4.1 was shown to be up-regulated in all tissues and the genetic variations in ZmOSCA4.1 were significantly associated with drought tolerance at the seedling stage through association analysis [20]. Over-expression of OsOSCA1.4 in the A. thaliana osca1 mutant can be complementary with its function in hyperosmolality reactions under salt stress [26]. Over-expression of eight OsOSCAs in A. thaliana demonstrated that these OsOSCA members were mainly involved in osmotic perception and stress adaption, which could regulate stomatal closure, leaf water loss and root growth through mediating osmotic Ca2+ signaling [27]. Additionally, the function of GhOSCA1.1 was validated by a virus-induced gene silenced (VIGS) approach, which controlled the activity of antioxidant enzymes to enhance salt and drought resistance [23].
Barley (Hordeum vulgare L.) is the fourth largest cereal crop all over the world in terms of cultivation area and annual total yield, with great edible, forage and brewing value [28]. Furthermore, barley is also one of the most stress tolerant crops globally, such as salt, low temperature and soil infertility stress, providing the gene pool for genetic improvement against biotic/abiotic stress. However, it is necessary to explore the OSCA gene family further in order to mine the elite candidates regulating abiotic stress tolerance. The objective of the current study was to systematically identify the barley OSCA gene family (HvOSCAs) based on a genome-search approach and its genomic organization, exon-intron structures and phylogenetic relationship were investigated. Furthermore, the expression patterns of them were systematically detected during development stages and under diverse abiotic stresses, and a co-expression network of them was constructed based on RNA-seq data to obtain the stress-related modules. Finally, quantitative real-time PCR (qRT-PCR) analysis was used to validate their expressions under diverse stresses and the stress-related candidates were found. Additionally, the genetic variations of these HvOSCAs were investigated using population re-sequencing data.
2. Results
2.1. Identification of OSCA Gene in Barley
Based on a genome-search method [18,19], a total of 13 putative OSCA genes were identified in the barley genome, which were nominated as HvOSCA1.1 to HvOSCA4.1 according to the orthologs in A. thaliana and rice. As two orthologs of OSCA1.3 and OSCA2.1 were found in barley, they were named HvOSCA1.3_1 and HvOSCA1.3_2, HvOSCA2.1_1 and HvOSCA2.1_2, respectively, indicating that segmental duplication events of OSCA genes had occurred in the barley genome. Sequence characteristics of these HvOSCAs showed that their gene length ranged from 2385 (HvOSCA4.1) to 26,794 (HvOSCA1.1) base-pair (bp), while their coding sequence length ranged from 2133 bp to 2400 bp, encoding 711 to 800 amino acids. The isoelectric point (pI) varied from 6.7 to 9.24, and all of the HvOSCA proteins had a positive GRAVY score, indicating that HvOSCA proteins were hydrophobic proteins. In addition, the subcellular localization prediction found that all of the HvOSCAs were located in the inner-membrane (Table S1), which was consistent with previous studies [18,20,23]. Chromosome distribution analysis found that these 13 HvOSCAs were unevenly distributed on 5 out of 7 barley chromosomes (Figure S1), of which chr1H, chr4H and chr5H contained 3 OSCA members, chr2H and chr3H harbored 2 OSCA members, and chr6H and chr7H did not have any OSCA members (Table S2).
2.2. Phylogenetic Relationship, Conserved Motifs and Gene Structure Analysis
To explore the phylogenetic relationships of the OSCA family, a neighbor-joining (NJ) phylogenetic tree was constructed using the full-length protein sequences of 13 HvOSCAs, 42 TaOSCAs, 15 AtOSCAs, 11 OsOSCAs and 11 ZmOSCAs (Table S3). Results showed that these OSCA proteins were classified into four clades based on phylogenetic relationship (Figure 1), which was consistent with previous studies [18,19]. Further, thirteen HvOSCAs were distributed asymmetrically in each clade, with five, six, one and one members belonging to clade 1 to clade 4, respectively. In general, the HvOSCAs displayed closer phylogenetic relationships with those of wheat and rice compared to that of A. thaliana.
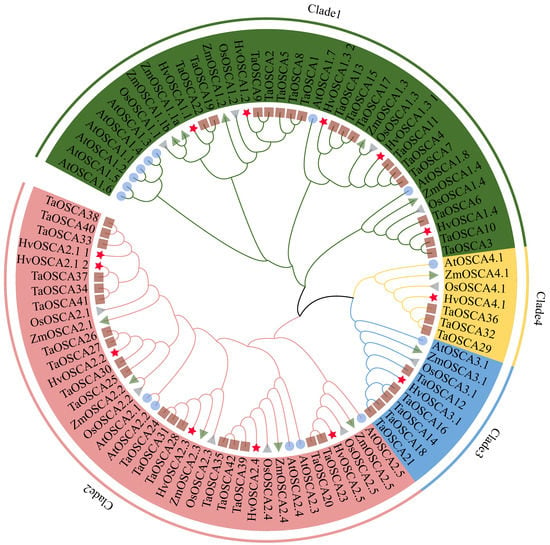
Figure 1.
Phylogenetic analysis of the OSCA genes in Hordeum vulgare (Hv), Arabidopsis thaliana (At), Triticum aestivum (Ta), Oryza satival (Os) and Zea mays (Zm) by the neighbor-joining method. The OSCAs were clustered into four clades; each member of the OSCAs were annotated by (★ for Hv), (● for At), (■ for Ta), (◄ for Os) and (► for Zm), respectively.
Then, the conserved motifs and exon-introns of the HvOSCAs were analyzed (Figure 2). It was indicated that all of the 13 HvOSCAs possessed three conserved function domains, RSN1_TM (PF13967), 7TM (PF02714) and PHM7_cyt (PF14703) (Figure 2B), which were the specific components of OSCA proteins [29,30], indicating the accuracy of the prediction of HvOSCAs. Furthermore, a total of 10 conserved motifs were also found in HvOSCAs (Figure 2C,E, Table S4). The majority of HvOSCAs displayed the relatively consistent motif compositions with 8 to 10 conserved motifs, of which HvOSCA2.1_1, HvOSCA2.1_2, HvOSCA2.2 and HvOSCA2.3 contained 9 motifs except motif-6, while HvOSCA1.1 and HvOSCA3.1 contained 9 motifs except motif-1. Additionally, HvOSCA4.1 had only one conserved motif. These different organizations of conserved motifs might correlate to the different function of these HvOSCAs.

Figure 2.
Motif composition, conserved domain and gene structure analysis of HvOSCAs. (A) Phylogenetic relationship of HvOSCAs, members of different clades were marked in different background colors. (B) Functional motif compositions of these 13 HvOSCAs. Ten motifs were represented by rectangle box in different colors. (C) The function conserved domain of PFAM02714, PFAM13967 and PFAM14703 organization in HvOSCAs, represented by dark blue, dark turquoise boxes and purplish blue, respectively. (D) Intron-exon organizations of HvOSCAs. The introns and exons are represented by the broken line and gray boxes, respectively. (E) The protein sequence of the 10 identified motifs.
Exon-intron structure is an evolutionary force to decide the functional diversification of the members in gene family [31]. It was found that the intron characteristics of HvOSCAs were highly variable with the number ranging from 0 to 11. Similar to its ortholog OsOSCA4.1 and AtOSCA4.1, HvOSCA4.1 had no introns reported, while HvOSCA1.1 had 11 introns. Additionally, HvOSCA1.1 also had the longest intron length, which might due to a retro-transposon-like element insertion. Overall, the members in the same clade based on phylogenetic relationship shared similar motif compositions and exon-intron structure, suggesting that they might have more similar biological functions.
2.3. Cis-Element Analysis of HvOSCAs
A total of 37 kinds of cis-elements were predicted in the upstream 2000 bp from the transcription start sites of HvOSCAs (Figure 3, Table S5), which were widely involved in growth biological process, hormone responsiveness, light responsiveness, metabolic regulation, as well as stress response. Among them, the elements associated with response to biotic or abiotic stresses were identified in almost all HvOSCAs, such as the drought responsive element (MBS) found in eleven HvOSCAs, anaerobic induction elements (ARE) in ten HvOSCAs, anoxic specific induction elements (GC-motif) in five HvOSCAs, as well as a wound responsive element (WUN-motif) in one HvOSCA and a defense-responsive element (AT-rich element) in one HvOSCA. Furthermore, nine types of hormone-responsive regulatory elements were also found, including auxin-responsive elements (AuxRR-core and TGA-element) in four HvOSCAs, gibberellin-responsive elements (P-box and GARE-motif) in eight HvOSCAs, salicylic acid-responsive elements (TCA-element) in four HvOSCAs, MeJA-responsive elements (CGTCA-motif and TGACG-motif) in ten HvOSCAs and ABA-responsive elements (ABRE) in eleven HvOSCAs. Additionally, there were meristem expression-related elements (CAT-box) in five HvOSCAs, circadian control-related elements (circadian) in two HvOSCAs and a cell cycle regulation-related element (SA-like) in one HvOSCA. The results provided some clues about the putative roles that HvOSCAs play in barley growth, development and stress response.
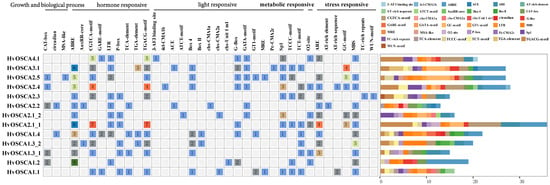
Figure 3.
Cis-elements found in the promoter region of HvOSCAs. (Left): The number and function classification of cis-acting element in each HvOSCA genes. (Right): Distribution of 37 identified cis-acting elements in each HvOSCA; elements are represented by the boxes in different colors.
2.4. Expression Profiles of HvOSCAs in Tissues and under Stress Conditions Based on RNA-seq Data
To obtain some clues about the potential function of HvOSCAs, spatio-temporal expression profiles of them were investigated based on previous open access available RNA-seq data of 16 different tissues at different growth stages (Figure 4A, Tables S6 and S7). Current study results showed that HvOSCA1.1 and HvOSCA3.1 were highly expressed in almost all tested tissues across diverse developmental stages, indicating that these OSCAs played important roles in regulating the growth and development of barley. HvOSCA4.1 was found to be expressed in all tissues/stages but with a relative lower level. At the same time, six HvOSCAs displayed no expression in these tissues/stages, including HvOSCA1.2, HvOSCA1.3_1, HvOSCA1.3_2, HvOSCA3.1, HvOSCA2.3 and HvOSCA2.5, suggesting that they might not function on barley growth and development. HvOSCA1.4 showed high expression in CAR5, RAC and LOO, but no expression in other stages/tissues. HvOSCA2.2 showed relatively higher expression in CAR15, LEM, NOD, RAC and ROC while lower expression in other tissues/stages. Furthermore, some tissue-specific HvOSCAs were also identified. It was interesting that the segmental duplication genes HvOSCA2.1_1 and HvOSCA2.1_2 showed diverging expression patterns in these tissues: HvOSCA2.1_2 was highly expressed in EPI, ETI, LEA and NOD, while HvOSCA2.1_1 only expressed in LEM and LOD, despite having highly conserved sequence organization. This divergent expression pattern might be due to their different cis-element compositions, and further study on their expression differentiation could provide helpful information reharding the evolution and function of the duplicated genes.
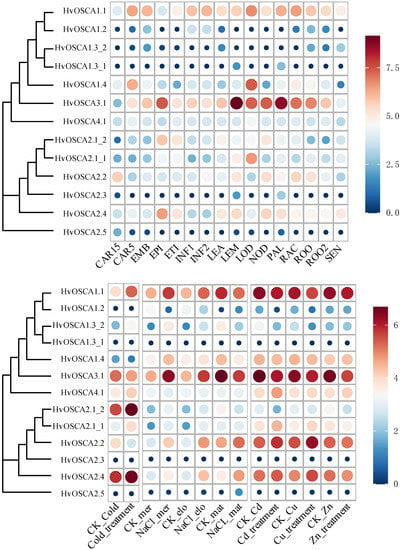
Figure 4.
Expression profiles of HvOSCAs in different tissues and under diverse stresses. (A) The spatio-temporal expression profile of HvOSCAs in different tissues at different developmental stages. FPKM values were normalized by log2(FPKM+1) transformation to display the heatmap color scores. CAR15: bracts removed grains at 15DAP; CAR5: bracts removed grains at 5DAP (CAR5); embryos dissected from 4-day-old germinating grains (EMB); epidermis at 4 weeks old (EPI); etiolated from 10-day-old seedling (ETI); young inflorescences with 5 mm (INF1); young inflorescences with 1–1.5 cm (INF2); shoot with the size of 10 cm from the seedlings (LEA); lemma at 6 weeks after anthesis (LEM); lodicule at 6 weeks after anthesis (LOD); developing tillers at the six-leaf stage (PAL); 6-week-old palea (NOD); rachis at 5 weeks after anthesis (RAC); root from 4-week-old seedlings (ROO2); roots from the seedlings at the 10 cm shoot stage (ROO). (B) Expression profile of HvOSCAs under cold, NaCl and heavy metal ion (Cd, Cu and Zn) conditions. Cold stress treatment: CK of cold (CK_cold), cold (4 °C) treatment (Cold_treatment), salt treatment of barley root (dissected and sampled the barley according to the organizational structure of root for meristem zone, elongation zone and mature zone). CK of meristem zone (CK_mer), meristem zone under salt treatment (NaCl_mer), CK of elongation zone (CK_elo), elongation zone under salt treatment (NaCl_elo), CK of maturation zone (CK_mat) and maturation zone under salt treatment (NaCl_mat). Heavy metal ion (Cd, Cu and Zn) stress. Control of cadmium ion stress (CK_Cd), cadmium ion treatment (Cd_treatment), control of copper ion treatment (CK_Cu), copper ion treatment (Cu_treatment), control of zinc ion stress (CK_Zn) and zipper ion treatment (Zn_treament).
As hyperosmotic stress sensors, OSCAs play a vital role in regulating abiotic stress signaling. Thus, the expression profiles of HvOSCAs were further investigated under cold, salinity and heavy metal ion abiotic stresses using the previous open access RNA-seq data (Figure 4B, Tables S6 and S8). Under cold stress treatment, HvOSCA1.2, HvOSCA1.3_1, HvOSCA2.3 and HvOSCA2.5 did not have any expression, while HvOSCA4.1 highlighted no differential expression between control and cold treatments. Compared to CK, only HvOSCA1.1, HvOSCA2.1_1, HvOSCA2.1_2, HvOSCA1.3_2 and HvOSCA2.4 showed up-regulated expression under cold stress treatment (Figure 4A), which could be considered as the cold-responsive candidates. Then, the expression patterns of HvOSCA genes were investigated in the meristem, elongation and mature zones of roots under salinity stress treatment, and most of them showed differential expressions (Figure 4B, Table S8). Compared with CK (mock), a total of eight, eight and four HvOSCA genes were significantly up-regulated in the meristem, elongation and maturation zones of roots, respectively. Especially, HvOSCA2.2 and HvOSCA2.4 exhibited an increasing expression in all three zones relative to the control. Under metal ion stresses, HvOSCA2.1_1, HvOSCA2.2 and HvOSCA2.4 were up-regulated under copper (Cu)- and cadmium (Cd)-stressed conditions, while down-regulated under zinc (Zn) treatment. By contrast, HvOSCA1.1, HvOSCA1.2, HvOSCA1.3_2 and HvOSCA4.1 were down-regulated under Cu and Cd treatments but up-regulated under zinc treatment. HvOSCA1.1 and HvOSCA1.2 were down-regulated in all of the three heavy metal ion treatments. The metal ion-responsive HvOSCAs provided potential targets for further functional genomics study to reveal their roles in regulating osmotic adjustment and ion toxicity.
2.5. Co-Expression Network Analysis of HvOSCAs Involved in Abiotic Stress Response
To get more information about the function and molecular modules of HvOSCAs that are involved in the response to abiotic stresses, the WGCNA method was employed to analyze the co-expression network based on the RNA-seq data under diverse stresses, and the HvOSCAs associated modules were identified. The present study results showed that a total of 27,987 genes displayed significant expressions, and these were used for constructing co-expression networks. A total of 47 co-expression modules with the gene number ranging from 34 to 7981 were obtained (Figures S2 and S3, Table S9). Then, the modules that 13 HvOSCAs were involved were further identified. Results found that 12 out 13 HvOSCAs were inv olved in 7 co-expression modules, including cyan, brown, blue, turquoise, green, pink and red modules. It is interesting that these 12 HvOSCAs had the conserved RSN1_7TM functional domain that is the most prominent characteristic of calcium-permeable stress-gated channel proteins, while the remaining HvOSCA2.3 without the RSN1_7TM domain was not found in these constructed modules. Furthermore, a correlation analysis of these seven modules with the stress treatments was performed (Figure S4). Interestingly, one, one, four, one, two, one and one HvOSCAs were found in the cyan, brown, blue, turquoise, green, pink and red, respectively. The red module was significantly correlated with cadmium treatment and the turquoise module was correlated with cadmium treatment and copper treatment, respectively. The green module was correlated with NaCl treatment of the mature zone of the root, and also correlated with copper treatment. The brown module was significantly correlated with NaCl treatment of the elongation zone. The cyan and pink modules were both correlated with NaCl treatment of the mature zone. The results suggest the function of HvOSCAs in the heavy metal ion and salt response.
Then, the current study predicted the miRNA that interacted with HvOSCAs and was involved in the stress response. Through prediction, eight HvOSCA genes were found to be targeted by thirteen miRNAs, including miR156, miR6195 and so on. Among them, HvOSCA1.3_1, HvOSCA1.1 and HvOSCA1.1 could be bound by three miRNAs, respectively. Meanwhile, Hvu-miR6206 could target both HvOSCA1.3_1 and HvOSCA1.2, and Hvu-miR6195 could target both HvOSCA1.2 and HvOSCA2.4. Furthermore, with integration of the miRNA–HvOSCA relationship and the co-regulation modules of HvOSCAs, the sophisticated regulatory network that HvOSCAs use as the hub and is mediated by miRNAs was constructed, and a total of 16 miRNA-HvOSCA interactions were obtained (Figure 5, Table S10). The miRNA-mediated networks contributed to better understand the roles of HvOSCAs in regulating stress response and resistance in barley, which paved a way to modulate HvOSCAs expression to induce some physiological changes and then enhance stress resistance through a post-transcriptional approach.

Figure 5.
miRNA¬–mRNA interaction network of HvOSCAs involving responses to abiotic stresses. The HvOSCAs are represented by the red diamonds, the predicted miRNAs by yellow ovals and the other interacting genes by blue dots.
2.6. Validation of the Expression of HvOSCAs by qRT-PCR Assays
As OSCA genes function as hyperosmotic stress sensors, they play a vital part in abiotic stress response, hormone regulation and signaling transduction processes [25]. To identify the salt-responsive candidates, the expression profiles of all 13 HvOSCAs were systematically validated under drought, salt, cold and ABA treatments by qRT-PCR analysis (Table S11). Under 100 µM ABA treatment, HvOSCA2.1_1, HvOSCA2.1_2, HvOSCA2.4 and HvOSCA3.1 showed up-regulated expression; in particular, the expression level of HvOSCA2.1_1 was significantly high. HvOSCA4.1 had gradually increased expression with the extension of treatment, and reached the peak at 24 h (h) after treatment. Moreover, the expressions of HvOSCA2.1_2, HvOSCA2.4 and HvOSCA3.1 were significantly up-regulated in the first 6 h, but decreased significantly as the treatment time continued, and finally were significantly lower compared to CK. Contrary to these results, the expressions of HvOSCA1.1, HvOSCA1.4 and HvOSCA2.2 displayed a continued decreasing trend, and reached the lowest levels at 24 h.
Under cold treatment, HvOSCA1.1, HvOSCA2.1_1, HvOSCA2.1_2, HvOSCA2.2, HvOSCA3.1 and HvOSCA4.1 were found to exhibit a remarkably higher expression level in the first 6 h and then decreased at 12 h after treatment. The expression level of HvOSCA1.3_2 was gradually increased with the duration of treatment, and reached the peak at 12 h, and then decreased at 24 h. The expression of HvOSCA1.4 showed no significant difference in the first 6 h of treatment, and then decreased significantly at 12 h, and finally increased sharply at 24 h. These results were consistent with the trends found by RNA-seq data and the differential expressed genes provided the candidates associated with the cold response.
Under 20% PEG-6000 treatment, these HvOSCAs showed completely diverse expression patterns. The expressions of HvOSCA1.1 and HvOSCA1.4 showed consistence with that of CK at 6 h of drought treatment, and increased rapidly to reach the highest level at 12 h. Then, HvOSCA1.1 maintained a high expression at 24 h of treatment, while HvOSCA1.4 decreased remarkably at 24 h. HvOSCA1.3_2 displayed the highest expression level at 6 h, and then decreased gradually during the drought treatment. At the same time, HvOSCA2.1_1, HvOSCA2.2 and HvOSCA4.1 showed similar expression patterns that were significantly down-regulated at 6 h and 12 h of treatment, and then up-regulated at 24 h. HvOSCA2.1_2 seemed to function as a negative regulator of the drought response and displayed significantly down-regulated expression at 12 h. Finally, the expression levels of HvOSCA2.4 and HvOSCA3.1 increased at 6 h, then decreased sharply at 12 h, but increased significantly to the highest level at 24 h. The different expression patterns of these HvOSCAs suggest their divergent regulatory roles in drought response and tolerance.
Under salt stress treatment, these HvOSCAs also showed obviously different expression patterns. HvOSCA2.1_2 displayed the up-regulated expression all the time with a high expression level, while HvOSCA2.2 and HvOSCA4.1 showed down-regulated expressions at all time courses, suggesting that these might function in differential stages of salt stress. HvOSCA3.1 and HvOSCA2.4 were up-regulated at both 6 h and 12 h, while down-regulated at 24 h. The expression levels of HvOSCA1.1 and HvOSCA1.4 was the highest at 6 h, then decreased sharply at 12 h, but increased a little at 24 h. HvOSCA1.3_2 showed no differential expression with CK at 6 h, but was up-regulated at 12 h and then down-regulated at 24 h. The expression of HvOSCA2.1_1 decreased rapidly after salt treatment, and reached the lowest expression at 6 h, then increased steadily to the highest level at 24 h. The results provided some clues on their divergent function on salt response and tolerance. Interestingly, the expressions of HvOSCA1.2, HvOSCA1.3_1, HvOSCA2.3 and HvOSCA3.1 had extremely low or seldom expression under all stress conditions, which was also demonstrated by RNA-seq analysis, indicating they might not function in the stress response (Figure 6, Table S12).
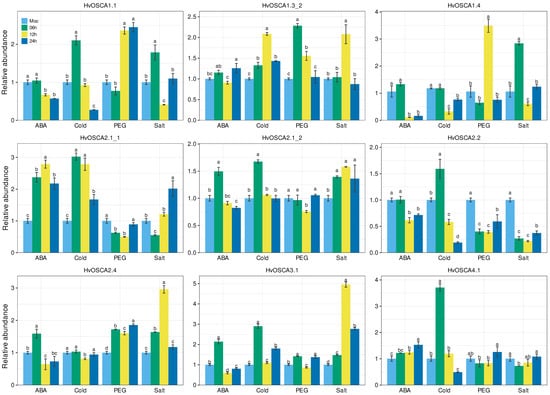
Figure 6.
Validation of the expression signature of nine HvOSCA genes under ABA (100 µM ABA), cold (4 °C), drought (20% PEG-6000) and salt (NaCl) treatment through the qRT-PCR method. The treatments were conducted with 100 µM ABA solution for ABA, 4 °C for cold, 20% PEG 6000 solution for drought, and 150 mM NaCl solution for salt stresses, respectively. Data represent the average of three technical replications and the error bar represents the standard error of mean (SEM).
2.7. Genetic Variations and Haplotype Analysis of HvOSCA
The genetic variations of these HvOSCA genes were investigated based on the resequencing data of wild barley and cultivated barley populations, including 85 wild and 135 landrace accessions. A total of 2077 SNP loci were found in the 13 HvOSCAs and then were used to calculate the population divergence (Fst) and nucleotide diversity (π) values. The current study results found that the Fst value of them ranged from 0 to 0.808825. According to a previous study, the Fst value of a gene exceeds the threshold line (Fst = 0.456) was considered as an artificially selected gene. Among these 13 HvOSCAs, HvOSCA1.3_1, HvOSCA2.2 and HvOSCA1.1, as well as HvOSCA2.2, were under artificial selection during barley domestication (Figure 7A, Table S13).
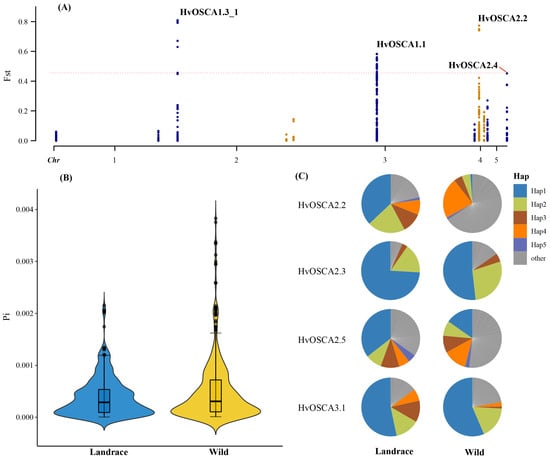
Figure 7.
Genetic variations and haplotype analysis of HvOSCAs based on resequencing data. (A) Genetic divergence and artificial selection analysis of HvOSCAs; (B) nucleotide diversity of HvOSCA genes in landrace and wild barley populations; (C) haplotype frequency analysis of HvOSCAs in landrace and wild barley populations.
Furthermore, the genetic diversity of HvOSCAs in the wild barley population was 0.4966 × 10−3, while that of landrace population was 0.391 × 10−3 (Figure 7B, Table S14). Compared to wild barley, the cultivated barley had relatively lower genetic diversity, suggesting that genetic bottleneck had occurred on the barley OSCA family during the barley evolution process. Further, the current study observed that the haplotype frequencies of the HvOSCAs were investigated based on the SNP variations. Results found only four genes showed different haplotype frequencies between wild and cultivated barley, and all of them displayed more haplotypes in wild barley than that of cultivated barley, which also demonstrated the genetic bottleneck effect on the OSCA family when wild barley domesticated to a landrace (Figure 7C, Table S15).
2.8. Subcellular Localization of HvOSCA1.1, HvOSCA 2.4, HvOSCA3.1 and HvOSCA 4.1
In order to validate the subcellular localization of HvOSCA proteins, four HvOSCAs genes, including HvOSCA1.1, HvOSCA2.4, HvOSCA3.1 and HvOSCA4.1 were cloned from the barley cv. Morex to construct their corresponding transient vectors, and then separately injected into tobacco leaf cells to track the localization of the HvOSCA proteins through laser confocal microscopy. As shown in Figure 8, all four proteins were expressed as HvOSCA-GFP fusion protein in transformed tobacco leaf cells, and they were mainly localized in the inner-membrane.
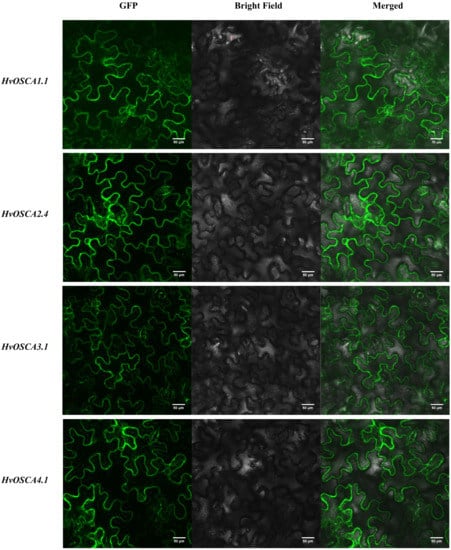
Figure 8.
Subcellular localization of four recombinant CaMV35S: HvOSCA-GFP HvOSCA protein fusions transiently expressed in tobacco leaf cells.
3. Discussion
Plants inevitably undergo changing environments including adverse abiotic environments, such as a drought, soil salinity and extreme temperatures, throughout their life cycle, and they have evolved interconnected regulatory pathways in response to the exogenous and endogenous osmotic changes [3]. Barley ranked as the fourth most important cereal crop worldwide [32], which is widely distributed from the vicinity of Dead Sea in the Middle East where H. vulgare originated, to the high altitude areas of the Qinghai-Tibet plateau [33]. Barley has a wide range of adaptability and can withstand harsh environments [34]. Extensive studies have demonstrated that barely is one of the most stress tolerant staple crops all over the world with the excellent salt tolerance, drought and low temperature resistance, as well as adaptation to soil infertility, providing a gene pool for genetic improvement and breeding of stress tolerant crops. Therefore, mining elite genes underlying stress tolerance in barley will provide a useful gene resource for stress-oriented genetic improvement and breeding. The OSCA family acts as an osmosensor, playing a crucial role in the sensing of osmotic stresses and regulating osmotic adjustment in plants [25,30]. In view of its importance, the OSCA family has been systematically identified in several species, including A. thaliana, rice, maize and wheat, but not so far in barely. In this study, 13 HvOSCA genes were identified at genome level in barley. Based on phylogenetic analysis, these HvOSCAs could be clustered into four clades, and the members clustered in the same clade possessed the detected similar conserved motif composition, exon-intron structures and cis-element organizations, suggesting that they had similar function, which is consistent with previous studies [20]. Domains of the late exocytosis TM putative phosphate transporter and calcium-dependent channel of OSCA genes were found to be extremely conserved in higher plants [25]. All HvOSCA genes identified herein contained these three functional domains, further supporting the high conservation of the OSCA family and also demonstrating the accuracy of our prediction.
The expression pattern is the external embodiment of gene function, and gene expression is largely regulated by the cis-elements in the promoter region [35]. Cis-regulatory elements play an essential role in a gene’s spatio-temporal expression, and then in regulating plant growth, development, morphogenesis, senescence, apoptosis, as well as in coordination and adaptation to the environment [36]. The current study found that HvOSCAs harbored various cis-elements related to growth, hormone responsiveness and also stress responsiveness. Combined with the expression profiles of HvOSCAs from RNA-seq and qRT-PCR analysis, the current study found that the cis-elements could regulate the expressional specificity of some HvOSCAs. For example, HvOSCA1.3_2, harboring five MBS cis-elements that are regulatory elements serving as a MYB binding site involved in drought inducibility, showed a quick response under PEG-induced drought treatment. HvOSCA1.2_2 showed rapid up-regulation expression under ABA treatment and maintained at a high expression level, which contained seven CGTCA-motif elements that involved in the MeJA responsiveness. The current study further reconstructed the co-expression network by the WGCNA method based on public RNA-seq data, and further investigated the correlations between seven modules and simulated abiotic stress treatment including cold, NaCl and heavy metal ion (cadmium, copper and zipper) treatment. Results showed that seven modules were significantly associated with stress treatment. All modules harbored the HvOSCA genes as the hub factors. Furthermore, prediction results of miRNA regulating OSCA genes in seven modules showed that thirteen microRNAs were found to target eight HvOSCA genes involved in the stress-related modules, including Hvu-miR5049f, Hvu-miR6195, Hvu-miR6196 and Hvu-miR156a. Previous studies have been reported that Hvu-miR5049f could respond to drought stress in Tibetan wild barley [37], Hvu-miR6195 and Hvu-miR6196 were detected in rice root under salt treatment [38], and the expression of Hvu-miR156a was modulated by barley leaf stripe (BLS) in Tibetan barley [39]. We postulated that these four miRNAs might play a regulatory role in the response to stress by controlling the expression of OSCA genes in barley.
In previous studies, different OSCA members mainly functioned as stress responsive proteins in dehydration stress or acted as hyperosmolality sensors, and also played the role of a switch to control the influx of Ca2+ in the pathway of immune signal transduction [20,25,26,27,40]. The present study found that HvOSCA2.1_1 was significantly induced by ABA and cold, but was inhibited by drought and salt treatment during the initial 12 h, while there was little change in expression of HvOSCA4.1 under ABA, PEG, salt and cold (except 6 h) treatment compared with control. HvOSCA2.2 was significantly inhibited by drought, salt and ABA, but markedly induced by cold in the initial 6 h. Among the cis-elements of HvOSCA genes, the number of hormone-responsive and stress-responsive elements were more than that of growth and metabolic-responsive elements, signifying that HvOSCAs mainly functioned in stress response and signal transduction.
Population genomic research has provided a crucial approach for the investigation of genetic divergence, domestication and evolution, as well as the genetic basis for the trait in plant populations [41,42,43,44].
The current study evaluated the genetic variations of the HvOSCA family in wild barley and landrace populations. Fst values among different populations indicated that artificial selection had occurred in barley OSCA genes at different degrees between members of four identified HvOSCAs . The results showed that wild barley possessed higher nucleotide diversity and alternative haplotypes compared to landrace populations, which suggested that genetic bottleneck had occurred within this family during barley domestication. These results suggested that barley domestication occurred from the perspectives of the OSCA family, and are promising enrich the genetic diversity for barley breeding against abiotic stress resistance.
Generally, the subcellular localization helps to decipher its cellular function. Previous studies have reported that OSCA proteins were localized in the cell membrane in other plants [18,45]. In the current study, both the in silico prediction and the subcellular localization results indicated that the HvOSCA family members are localized in the inner-membrane. The results in the current study testified the previous conclusions and provided useful information for further functional studies of HvOSCAs.
4. Materials and Methods
4.1. Identification of OSCA Genes in Barley
For the identification of the OSCA gene family in barley, the protein sequences of the barley genome were retrieved from the Ensembl plant database (http://plants.ensembl.org/Hordeum_vulgare/Info/Index, accessed on 10 January 2022). The Hidden Markov Model (HMM) profile of homeodomain (HD) (DUF221) (renamed from DUF221 to RSN1_7TM (PF02714) was used as a query to search against the local protein database using the HMM3.0 tool with the E-value ≤ 1 × 10−10 as the threshold. Furthermore, 15 AtOSCAs and 11 OsOSCAs protein sequences were used to perform a BLASTP search against the local protein database with the threshold of E-value < 1 × 10−5. Subsequently, the results of the HMMER and BLASTP searches were integrated together, and the redundants were manually removed to obtain the putative barley OSCA proteins. These data were submitted to the Conserved Domain Database (CDD) (https://www.ncbi.nlm.nih.gov/cdd, accessed on 10 January 2022), the Simple Modular Architecture Research Tool (SMART) (http://smart.embl-heidelberg.de/, accessed on 5 February 2022) and PFAM (https://pfam.xfam.org, accessed on 1 March 2022) to further confirm their OSCA conserved domain; those with a complete OSCA domain remained as candidates. Finally, the candidates were submitted to the ExPASy (http://web.expasy.org/protparam/, accessed on 1 March 2022) database to investigate the physicochemical properties such as molecular weight (Mw), isoelectric point (pI), instability index (II), aliphatic index (AI) and grand average of hydrophobicity (GRAVY). The subcellular localization of them was predicted by the online CELLO tool (v2.5) (http://cello.life.nctu.edu.tw/, accessed on 1 March 2022).
4.2. Phylogenetic Relationships, Genic Structure, Conserved Motif and Cis-Element Analysis
The protein sequences of the identified barley and previously reported AtOSCAs, ZmOSCAs, OsOSCAs and TaOSCAs were employed to construct the phylogenetic tree. Multiple sequence alignment was performed using Phylip software [46] and the MEGA-X tool was used for a phylogenetic tree constructed by the neighbor-joining (NJ) method with the bootstrap of 1000 replications. The chromosome location and exon-intron structures of these HvOSCA genes were retrieved according to the genome annotation files (http://plants. ensembl.org/Hordeum_vulgare/Info/Index, accessed on 10 January 2022) and then were visualized using MapGene2Chromosome v2.0 (http://mg2c.iask.in/mg2c_v2.0/, accessed on 1 March 2022) and Gene Structure Display Server (GSDS2.0) (http://gsds.cbi.pku.edu.cn/, accessed on 1 March 2022), respectively. The conserved protein motifs were predicted based on online MEME tools (http://alternate.meme-suite.org/, accessed on 1 March 2022) with the following parameters: the maximum number of motifs was set to 10, any number of repetitions was allowed, and the optimum width ranged from 6 to 250.
4.3. Cis-Element Analysis
The 2.0 kb genomic sequences upstream of the transcription start site of HvOSCAs were extracted from the barley reference sequence by in-home Perl script and submitted to the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 12 March 2022) to predict the supposed cis-acting transcriptional regulatory elements.
4.4. Expression Profile Analysis of HvOSCAs Based on RNA-seq Data
A total of 142 RNA-seq data from the samples of different tissues at different development stages (including embryos, bracts, epidermis, young inflorescences, lemma, lodicule, shoot, rachis, developing tillers, palea, roots and leaves) and under diverse biotic and abiotic stresses (including drought, salinity, cold and ABA treatment) were downloaded from the NCBI Sequence Reading Archive (SRA) database (Table S6). The HISAT2 (v2.1.0) [47] and StringTie (v1.3.5) pipeline [48] were used to calculate the value of fragments per kilobase per million (FPKM). The expression profiles of HvOSCAs were retrieved and visualized with heat maps by using the pheatmap package in R software.
4.5. Co-Expression Network Analysis of HvOSCAs under Stressed Conditions
The R package WGCNA (V1.71, R 4.2) [49] was employed to conduct co-expression analysis based on the FPKM values matrix from various stresses with the default parameters except soft power = 8, min_module_size = 30, ME_miss_thread = 0.2. The FPKM values matrix of cold, NaCl and heavy metal ion (cadmium, copper and zipper) treatment were used for correlation analysis, and visualization was done by the ggcor package (V0.9.8.1). Thirteen HvOSCAs protein sequences were used as queries to predict the miRNA binding sites by using published miRNAs of barley in the psRNATarget database (https://www.zhaolab.org/psRNATarget/analysis?function=2, accessed on 18 May 2022) with the default parameters. The software Cytoscape v3.9.1 [50] was employed to visualize the regulatory network with miRNA of HvOSCAs.
4.6. Plant Materials Preparation, Stress Treatment and qRT-PCR Analysis
The seeds of barley cv. Morex, which is stored by the Wheat Genomics Lab in Northwest A&F University (Central Shaanxi Province, China; 34 26′ N latitude, 108 07′ E longitude, altitude: 458 m), were sterilized by 5% (w/v) sodium hypochlorite (NaClO) (Xinyicheng, Shaanxi, China) for 5 min, and then rinsed using distilled water for 3 min, and finally incubated on wet filter paper in a petri dish at 25 °C for 5 days [51]. This experiment was conducted from 10 to 30 March 2022 in NWSUAF. When the seedlings had grown to 4–5 cm, they were transplanted to hydroponic colonization plates, and put in culture medium (aqueous solution with 1/2 Hogeland nutrient) to reach the trifoliate leaf stage in a growth chamber under 20 °C /15 °C day/night, 16 h light/8 h dark cycle, and 50% relative humidity. Hydraulically cultured trifoliate leaf stage seedlings were exposed to culture medium, adding 150 mM NaCl, 20% PEG, 4 °C, or 100 µM ABA, and the roots and leaves were sampled at 0, 6 h, 12 h and 24 h as the stressed treatments [51,52]. The seedlings grown in the culture medium without any treatment were sampled simultaneously at the corresponding time point as the control. All the samples were collected from at least three replicates, frozen by liquid nitrogen immediately and then stored at −80 °C. Total RNA was extracted by Plant RNA Kit reagent (Omega BioTek, Norcross, GA, USA), then synthesis cDNA by 5× All-in-one RT MasterMix (ABM,Vancouver, Canada) following the manufacturer’s protocols. The HvACTIN (HORVU.MOREX.r2.5HG0378970) was used as the reference gene. The relative expression of HvOSCAs was quantified using the TB-Green ®Premix Ex Taq™ II kit (Takara, Dalian, China) and ABI7500 Real-Time PCR Systems (Applied Biosystems, Foster City, CA, USA). The qRT-PCR thermal cycling condition as follows: 95 °C for 30 s, followed by 40 cycles of 15 s at 95 °C, and 30 s at 60 °C. The 2−ΔΔCT quantification method was used for calculating the relative expression level [52,53]. Three parallel technical replicates were set for each sample. The primers used in the current study are listed in Table S11.
4.7. Genetic Variations and Haplotype Analysis of HvOSCAs
The resequencing data of 220 genotypes of barley (including 135 landraces and 85 wild barley) were obtained by an exome-captured approach from the NCBI SRA database (PRJEB8044/ERP009079), producing a visual component framework (VCF) file [43,52]. The SNPs of OSCA genes subsets were retrieved from the VCF file. The fixation index (Fst) was analyzed by vcftools v0.1.16 with the parameters as window = 100, step = 1, and visualized by R package CMplot. The nucleotide diversity (π) was also calculated by using vcftools v0.1.16, and visualized by R package ggsci. The variant of all HvOSCAs were annotated by snpEff software, the intro-variants were filtered, and the protein-coding variants were used for haplotype analysis by in-home python scripts [54], and visualized by R packages ggsci and scales.
4.8. Subcellular Localization of HvOSCA-GFP Fusion Proteins
One HvOSCA member from each clade based on phylogenetic relationships (HvOSCA1.1, HvOSCA 2.4, HvOSCA3.1 and HvOSCA4.1) was randomly selected for amplification using the primers listed in Table S11. The PCR products were separately recovered and cloned into the multiple cloning sites of NdeI and SacI in the pBI121-GFP vector. Each recombinant vector was separately transformed into Agrobacterium tumefaciens strain GV3101 prepared for agro-infiltration. For transient expression of HvOSCAs in tobacco, the Agrobacterium strains harboring the recombined vector were injected into 4-week-old tobacco leaves. Then, the images of the leaves were captured after 48 h of agro-infiltration by a laser confocal microscope (Olympus, Tokyo, Japan).
5. Conclusions
The current study identified and characterized a new OSCA gene family in barley with 13 HvOSCAs belonging to 4 clades. The exon-intron structures analysis found that they displayed highly variable intron numbers ranging from 0 to 11. Expression profiles and qRT-PCR validation found that these HvOSCAs were significantly induced by drought, cold, salt and phytohormone ABA treatment, demonstrating their function in stress response. Finally, genetic variations and haplotype analysis indicated that artificial selection was conserved on the OSCA family during wild barley domestication to the barley landrace. This study not only paves the way for further elucidating the function of HvOSCAs on stress response and tolerance but also provides a potential gene resource for breeding studies on stress tolerance in barely and other crops.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms232113027/s1.
Author Contributions
Conceptualization, K.S. and W.S.; formal analysis, K.S.; funding acquisition, W.S.; investigation, K.S., W.P., T.S. and Y.C. (Yue Cheng); project administration, W.S.; resources, W.S.; validation, K.S., T.S., W.P., Y.C. (Yingqi Chu) and Y.Y.; visualization, K.S and W.P. and B.M.; writing—original draft, K.S.; writing-review and editing, K.S and W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Programmer of Introduction Talents of Innovative Discipline to Universities (Project 111) from the State Administration of Foreign Experts Affairs, China (Grant No. #B18042), and partially supported by the Key Research and Development Program of Shaanxi Province, China (Grant Nos. CSBAA202202 and CSBAA202203).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All of the datasets supporting the results of this article are included within the article and its additional files.
Acknowledgments
We appreciate Jun’an Yan and Guang Yang for their help in chart drawing and reading through the earlier manuscript and giving constructive advices, and we are grateful to the High-Performance Computing Center of Northwest A&F University for providing computational resources.
Conflicts of Interest
All authors declare no conflict of interest.
References
- Braam, J. In touch: Plant responses to mechanical stimuli. New Phytol. 2005, 165, 373–389. [Google Scholar] [CrossRef]
- Scheres, B.; van der Putten, W.H. The plant perceptron connects environment to development. Nature 2017, 543, 337–345. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Feijó, J.A.; Wudick, M.M. ‘Calcium is life’. J. Exp. Bot. 2018, 69, 4147–4150. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, Y.; Jahan, N.; Chen, G.; Ren, D.; Guo, L. Sensing of Abiotic Stress and Ionic Stress Responses in Plants. Int. J. Mol. Sci. 2018, 19, 3298. [Google Scholar] [CrossRef] [PubMed]
- Sukharev, S.I.; Blount, P.; Martinac, B.; Kung, C. Mechanosensitive channels of Escherichia coli: The MscL gene, protein, and activities. Annu. Rev. Physiol. 1997, 59, 633–657. [Google Scholar] [CrossRef]
- Sachs, F. Mechanical transduction by ion channels: A cautionary tale. World J. Neurol. 2015, 5, 74. [Google Scholar] [CrossRef]
- Maingret, F.; Fosset, M.; Lesage, F.; Lazdunski, M.; Honoré, E. TRAAK Is a Mammalian Neuronal Mechano-gated K+Channel*. J. Biol. Chem. 1999, 274, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Sharif-Naeini, R.; Folgering, J.R.; Bichet, D.; Duprat, F.; Honoré, E. Canonical TRP channels and mechanotransduction: From physiology to disease states. Pflug. Arch. Eur. J. Physiol. 2010, 460, 571–581. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef]
- Frietsch, S.; Wang, Y.-F.; Sladek, C.; Poulsen, L.R.; Romanowsky, S.M.; Schroeder, J.I.; Harper, J.F. A cyclic nucleotide-gated channel is essential for polarized tip growth of pollen. Proc. Natl. Acad. Sci. USA 2007, 104, 14531–14536. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.-F.; Fei, C.-F.; Dong, J.-Y.; Gu, L.-L.; Wang, Y.-F. Arabidopsis CNGC18 is a Ca2+-permeable channel. Mol. Plant 2014, 7, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.-F.; Gu, L.-L.; Wang, H.-Q.; Fei, C.-F.; Fang, X.; Hussain, J.; Sun, S.-J.; Dong, J.-Y.; Liu, H.; Wang, Y.-F. Cyclic nucleotide-gated channel 18 is an essential Ca2+ channel in pollen tube tips for pollen tube guidance to ovules in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 3096–3101. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yin, H.; Gu, J.; Li, L.; Liu, Z.; Jiang, X.; Zhou, H.; Wei, S.; Zhang, S.; Wu, J. Genomic characterization, phylogenetic comparison and differential expression of the cyclic nucleotide-gated channels gene family in pear (Pyrus bretchneideri Rehd.). Genomics 2015, 105, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Weiland, M.; Mancuso, S.; Baluska, F. Signalling via glutamate and GLRs in Arabidopsis thaliana. Funct. Plant Biol. 2015, 43, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Michard, E.; Lima, P.T.; Borges, F.; Silva, A.C.; Portes, M.T.; Carvalho, J.E.; Gilliham, M.; Liu, L.-H.; Obermeyer, G.; Feijó, J.A. Glutamate receptor-like genes form Ca2+ channels in pollen tubes and are regulated by pistil D-serine. Science 2011, 332, 434–437. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yuan, F.; Wen, Z.; Li, Y.; Wang, F.; Zhu, T.; Zhuo, W.; Jin, X.; Wang, Y.; Zhao, H.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed]
- Tong, K.; Wu, X.; He, L.; Qiu, S.; Liu, S.; Cai, L.; Rao, S.; Chen, J. Genome-Wide Identification and Expression Profile of OSCA Gene Family Members in Triticum aestivum L. Int. J. Mol. Sci. 2022, 23, 469. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Feng, X.; Du, H.; Wang, H. Genome-wide analysis of maize OSCA family members and their involvement in drought stress. PeerJ 2019, 7, e6765. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Zhang, P.; Lu, X.; Wang, G.; Wang, Z.; Zhang, Q.; Zhang, X.; Wei, X.; Mei, F.; Wei, L.; et al. Systematic Analysis of the Maize OSCA Genes Revealing ZmOSCA Family Members Involved in Osmotic Stress and ZmOSCA2.4 Confers Enhanced Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 351. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, M.; Wu, R.; Chen, X.; Liu, F.; Xing, B. Genome-wide analysis of OSCA gene family members in Vigna radiata and their involvement in the osmotic response. BMC Plant Biol 2021, 21, 408. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, Y.; Yang, F.; Magwanga, R.O.; Cai, X.; Wang, X.; Wang, Y.; Hou, Y.; Wang, K.; Liu, F.; et al. Genome-wide identification of OSCA gene family and their potential function in the regulation of dehydration and salt stress in Gossypium hirsutum. J. Cotton Res. 2019, 2, 1–18. [Google Scholar] [CrossRef]
- Thor, K.; Jiang, S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.; Dindas, J.; Derbyshire, P.; Leitao, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Zhai, Y.; Wen, Z.; Han, Y.; Zhuo, W.; Wang, F.; Xi, C.; Liu, J.; Gao, P.; Zhao, H.; Wang, Y.; et al. Heterogeneous expression of plasma-membrane-localised OsOSCA1.4 complements osmotic sensing based on hyperosmolality and salt stress in Arabidopsis osca1 mutant. Cell Calcium 2020, 91, 102261. [Google Scholar] [CrossRef]
- Zhai, Y.; Wen, Z.; Fang, W.; Wang, Y.; Xi, C.; Liu, J.; Zhao, H.; Wang, Y.; Han, S. Functional analysis of rice OSCA genes overexpressed in the arabidopsis osca1 mutant due to drought and salt stresses. Transgenic Res. 2021, 30, 811–820. [Google Scholar] [CrossRef]
- Kirschner, G.K.; Rosignoli, S.; Guo, L.; Vardanega, I.; Imani, J.; Altmüller, J.; Milner, S.G.; Balzano, R.; Nagel, K.A.; Pflugfelder, D.; et al. ENHANCED GRAVITROPISM 2 encodes a STERILE ALPHA MOTIF—containing protein that controls root growth angle in barley and wheat. Proc. Natl. Acad. Sci. USA 2021, 118, e2101526118. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, B.; Smith, E.N.; Drees, B.; Brem, R.B.; Kruglyak, L.; Bumgarner, R.E.; Schadt, E.E. Integrating large-scale functional genomic data to dissect the complexity of yeast regulatory networks. Nat. Genet. 2008, 40, 854–861. [Google Scholar] [CrossRef]
- Hou, C.; Tian, W.; Kleist, T.; He, K.; Garcia, V.; Bai, F.; Hao, Y.; Luan, S.; Li, L. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.C.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Secur. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- Aldughpassi, A.; Wolever, T.M.S.; Abdel-Aal, E.S.M. Barley Food and Product. In Encyclopedia of Food and Health; Elsevier Ltd.: Alpharetta, GA, USA, 2016; pp. 328–331. [Google Scholar]
- Kumar, A.; Verma, R.P.S.; Singh, A.; Kumar Sharma, H.; Devi, G. “Barley landraces: Ecological heritage for edaphic stress adaptations and sustainable production”. Environ. Sustain. Indic. 2020, 6, 100035. [Google Scholar] [CrossRef]
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-regulatory elements used to control gene expression in plants. Plant Cell Tissue Organ Cult. 2016, 127, 269–287. [Google Scholar] [CrossRef]
- Priest, H.D.; Filichkin, S.A.; Mockler, T.C. cis-Regulatory elements in plant cell signaling. Curr. Opin. Plant Biol. 2009, 12, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.-W.; Zhao, J.; Chen, Q.; Wu, F. Genome-wide characterization of drought stress responsive long non-coding RNAs in Tibetan wild barley. Environ. Exp. Bot. 2019, 164, 124–134. [Google Scholar] [CrossRef]
- Huang, X.; Jiejie, F.; Wang, R.; Zhang, H.; Huang, J. Comparative analysis of microRNAs and their targets in the roots of two cultivars with contrasting salt tolerance in rice (Oryza sativa L.). Plant Growth Regul. 2019, 87, 139–148. [Google Scholar] [CrossRef]
- Yao, X.; Wang, Y.; Yao, Y.; Bai, Y.; Wu, K.; Qiao, Y. Identification microRNAs and target genes in Tibetan hulless barley to BLS infection. Agron. J. 2021, 113, 2273–2292. [Google Scholar] [CrossRef]
- Maity, K.; Heumann, J.M.; McGrath, A.P.; Kopcho, N.J.; Hsu, P.K.; Lee, C.W.; Mapes, J.H.; Garza, D.; Krishnan, S.; Morgan, G.P.; et al. Cryo-EM structure of OSCA1.2 from Oryza sativa elucidates the mechanical basis of potential membrane hyperosmolality gating. Proc. Natl. Acad. Sci. USA 2019, 116, 14309–14318. [Google Scholar] [CrossRef]
- Mascher, M.; Richmond, T.A.; Gerhardt, D.J.; Himmelbach, A.; Clissold, L.; Sampath, D.; Ayling, S.; Steuernagel, B.; Pfeifer, M.; D’Ascenzo, M.; et al. Barley whole exome capture: A tool for genomic research in the genus Hordeum and beyond. Plant J. Cell Mol. Biol. 2013, 76, 494–505. [Google Scholar] [CrossRef]
- Pankin, A.; Altmüller, J.; Becker, C.; von Korff, M. Targeted resequencing reveals genomic signatures of barley domestication. New Phytol. 2018, 218, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.; Mascher, M.; Dawson, I.K.; Kyriakidis, S.; Calixto, C.; Freund, F.; Bayer, M.; Milne, I.; Marshall-Griffiths, T.; Heinen, S.; et al. Exome sequencing of geographically diverse barley landraces and wild relatives gives insights into environmental adaptation. Nat. Genet. 2016, 48, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Guo, Y.; Xu, Q.; Mascher, M.; Guo, G.; Li, S.; Mao, L.; Liu, Q.; Xia, Z.; Zhou, J.; et al. Origin and evolution of qingke barley in Tibet. Nat. Commun. 2018, 9, 5433. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.; Li, F.; Han, Y.; Yao, Z.; Xu, Z.; Chen, X.; Liu, J.; Zhang, Y.; Wang, A. Identification of OSCA gene family in Solanum habrochaites and its function analysis under stress. BMC Genom. 2022, 23, 547. [Google Scholar] [CrossRef] [PubMed]
- Retief, J.D. Phylogenetic Analysis Using PHYLIP. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 1999; pp. 243–258. [Google Scholar]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]
- Li, T.; Pan, W.; Yuan, Y.; Liu, Y.; Li, Y.; Wu, X.; Wang, F.; Cui, L. Identification, Characterization, and Expression Profile Analysis of the mTERF Gene Family and Its Role in the Response to Abiotic Stress in Barley (Hordeum vulgare L.). Front. Plant Sci. 2021, 12, 684619. [Google Scholar] [CrossRef]
- Ai, Q.; Pan, W.; Zeng, Y.; Li, Y.; Cui, L. CCCH Zinc finger genes in Barley: Genome-wide identification, evolution, expression and haplotype analysis. BMC Plant Biol. 2022, 22, 117. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, Y.; Wei, X.; Cui, L.; Nie, X. Genetic Diversity of Transcription Factor Genes in Triticum and Mining for Promising Haplotypes for Beneficial Agronomic Traits. Front. Plant Sci. 2022, 13, 899292. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).