

Development and Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus

 , , ,
, , ,
Abstract
1. Introduction
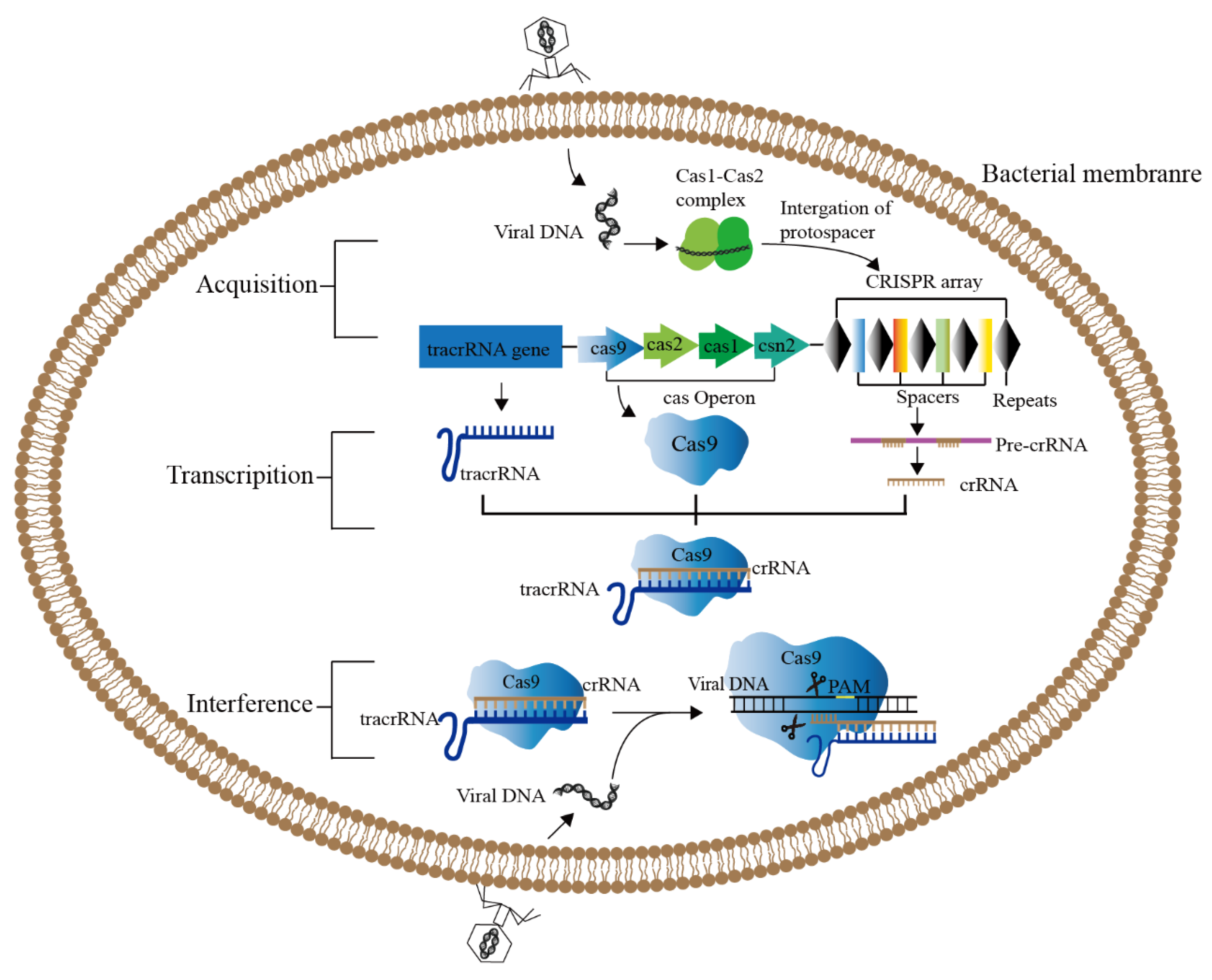
2. CRISPR/Cas9 Genome Editing System
2.1. Components of CRISPR/Cas9 Genome Editing System
2.2. Mechanism of CRISPR/Cas9 Genome Editing System
3. Development of CRISPR/Cas9-Based Genome Editing in Lactobacillus
3.1. Modifications of Cas9 for Genome Editing
3.2. Guide RNA Expression
3.3. CRISPR/Cas9-Assisted ssDNA Recombineering
3.4. CRISPR/Cas9-Assisted dsDNA Recombineering
3.5. RecE/T-Assisted CRISPR/Cas9 System
4. Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus
5. CRISPR/Cas9-Based Transcriptional Regulation in Lactobacillus
6. Future Perspectives
6.1. Improvement of Immune Regulation Traits
6.2. Increased Resistance to Environmental Stress
6.3. Development of Efficient Mucosal Vaccines
7. Conclusions and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Oberg, T.S.; McMahon, D.J.; Culumber, M.D.; McAuliffe, O.; Oberg, C.J. Invited review: Review of taxonomic changes in dairy-related lactobacilli. J. Dairy Sci. 2022, 105, 2750–2770. [Google Scholar] [CrossRef] [PubMed]
- Leuschner, R.G.K.; Robinson, T.P.; Hugas, M.; Cocconcelli, P.S.; Richard-Forget, F.; Klein, G.; Licht, T.R.; Nguyen-The, C.; Querol, A.; Richardson, M.; et al. Qualified presumption of safety (QPS): A generic risk assessment approach for biological agents notified to the European Food Safety Authority (EFSA). Trends Food Sci. Technol. 2010, 21, 425–435. [Google Scholar] [CrossRef]
- Rocchetti, M.T.; Russo, P.; Spano, G.; De Santis, L.; Iarusso, I.; De Simone, N.; Brahimi, S.; Fiocco, D.; Capozzi, V. Exploring the Probiotic Potential of Dairy Industrial-Relevant Lactobacilli. Appl. Sci. 2022, 12, 4989. [Google Scholar] [CrossRef]
- Gangaiah, D.; Mane, S.P.; Tawari, N.R.; Lakshmanan, N.; Ryan, V.; Volland, A.; Susanti, D.; Patel, M.; Abouzeid, A.; Helmes, E.B.; et al. In silico, in vitro and in vivo safety evaluation of Limosilactobacillus reuteri strains ATCC PTA-126787 & ATCC PTA-126788 for potential probiotic applications. PLoS ONE 2022, 17, e0262663. [Google Scholar] [CrossRef]
- Spacova, I.; Ahannach, S.; Breynaert, A.; Erreygers, I.; Wittouck, S.; Bron, P.A.; Van Beeck, W.; Eilers, T.; Alloul, A.; Blansaer, N.; et al. Spontaneous Riboflavin-Overproducing Limosilactobacillus reuteri for Biofortification of Fermented Foods. Front. Nutr. 2022, 9, 916607. [Google Scholar] [CrossRef]
- Luan, X.; Feng, M.; Sun, J. Effect of Lactobacillus plantarum on antioxidant activity in fermented sausage. Food Res. Int. 2021, 144, 110351. [Google Scholar] [CrossRef]
- Yang, Y.; Lian, Y.; Yin, S.; Suo, H.; Zeng, F.; Wang, H.; Song, J.; Zhang, Y. Inhibition of Lactobacillus fermentum SHY10 on the white membrane production of soaked pickled radish. Food Sci. Nutr. 2022, 10, 2236–2244. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, S.; Hao, G.; Yu, H.; Tian, H.; Zhao, G. Role of lactic acid bacteria on the yogurt flavour: A review. Int. J. Food Prop. 2017, 20, S316–S330. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Liu, F.; Jin, Z.; Xia, X. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. Nutr. 2022, 11, 1–15. [Google Scholar] [CrossRef]
- Aziz, G.; Zaidi, A.; Tariq, M. Compositional Quality and Possible Gastrointestinal Performance of Marketed Probiotic Supplements. Probiot. Antimicrob. Proteins 2022, 14, 288–312. [Google Scholar] [CrossRef]
- Randhawa, S.; Sengar, S. The evolution and history of gene editing technologies. In Advances in Crispr/Cas and Related Technologies; Ghosh, D., Ed.; Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2021; Volume 178, pp. 1–62. [Google Scholar]
- Kozovska, Z.; Rajcaniova, S.; Munteanu, P.; Dzacovska, S.; Demkova, L. CRISPR: History and perspectives to the future. Biomed. Pharmacother. 2021, 141, 111917. [Google Scholar] [CrossRef]
- Yin, H.; Kauffman, K.J.; Anderson, D.G. Delivery technologies for genome editing. Nat. Rev. Drug Discov. 2017, 16, 387–399. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. Innovation Talens: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the Immune System of Bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef]
- Jansen, R.; Embden, J.D.A.V.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR-Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J. A Tale of Two Moieties: Rapidly Evolving CRISPR/Cas-Based Genome Editing. Trends Biochem. Sci. 2020, 45, 874–888. [Google Scholar] [CrossRef] [PubMed]
- Akram, F.; Sahreen, S.; Aamir, F.; ul Haq, I.; Malik, K.; Imtiaz, M.; Naseem, W.; Nasir, N.; Waheed, H.M. An Insight into Modern Targeted Genome-Editing Technologies with a Special Focus on CRISPR/Cas9 and its Applications. Mol. Biotechnol. 2022, 26, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Zhang, F.; Koonin, E.V. SnapShot: Class 1 CRISPR-Cas Systems. Cell 2017, 168, 946. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Zhang, F.; Koonin, E.V. SnapShot: Class 2 CRISPR-Cas Systems. Cell 2017, 168, 328. [Google Scholar] [CrossRef]
- Tsai, Y.-S.; Chen, Y.-P.; Lin, S.-W.; Chen, Y.-L.; Chen, C.-C.; Huang, G.-J. Lactobacillus rhamnosus GKLC1 ameliorates cisplatin-induced chronic nephrotoxicity by inhibiting cell inflammation and apoptosis. Biomed. Pharmacother. 2022, 147, 112701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Zhang, L.; Chen, Q.; Tan, F.; Zhao, X. Effect of Lactobacillus fermentum HFY03 on the Antifatigue and Antioxidation Ability of Running Exhausted Mice. Oxidative Med. Cell. Longev. 2021, 2021, 8013681. [Google Scholar] [CrossRef]
- Jam, S.A.M.; Talebi, M.; Alipour, B.; Khosroushahi, A.Y. The therapeutic effect of potentially probiotic Lactobacillus paracasei on dimethylhydrazine induced colorectal cancer in rats. Food Biosci. 2021, 41, 101097. [Google Scholar] [CrossRef]
- Sun, N.; Ni, X.; Wang, H.; Xin, J.; Zhao, Y.; Pan, K.; Jing, B.; Zeng, D. Probiotic Lactobacillus johnsonii BS15 Prevents Memory Dysfunction Induced by Chronic High-Fluorine Intake through Modulating Intestinal Environment and Improving Gut Development. Probiotics Antimicrob. Proteins 2020, 12, 1420–1438. [Google Scholar] [CrossRef]
- Dai, M.; Li, Y.; Xu, L.; Wu, D.; Zhou, Q.; Li, P.; Gu, Q. A Novel Bacteriocin From Lactobacillus Pentosus ZFM94 and Its Antibacterial Mode of Action. Front. Nutr. 2021, 8, 710862. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Z.; Tian, S.; Fu, H.; Leng, X.; Chen, L. Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides. Animals 2021, 11, 2564. [Google Scholar] [CrossRef]
- Damodharan, K.; Palaniyandi, S.A.; Yang, S.H.; Suh, J.W. Functional Probiotic Characterization and In Vivo Cholesterol-Lowering Activity of Lactobacillus helveticus Isolated from Fermented Cow Milk. J. Microbiol. Biotechnol. 2016, 26, 1675–1686. [Google Scholar] [CrossRef]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef]
- Panda, G.; Ray, A. Decrypting the mechanistic basis of CRISPR/Cas9 protein. Prog. Biophys. Mol. Biol. 2022, 172, 60–76. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR–Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef]
- Nishimasu, H.; Ran, F.A.; Hsu, P.D.; Konermann, S.; Shehata, S.I.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal Structure of Cas9 in Complex with Guide RNA and Target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569. [Google Scholar] [CrossRef]
- Hu, J.H.; Miller, S.M.; Geurts, M.H.; Tang, W.; Chen, L.; Sun, N.; Zeina, C.M.; Gao, X.; Rees, H.A.; Lin, Z.; et al. Evolved Cas9 variants with broad PAM compatibility and high DNA specificity. Nature 2018, 556, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, W.; Xin, C.; Yin, J.; Shang, Y.; Ai, C.; Li, J.; Meng, F.-L.; Hu, J. Global detection of DNA repair outcomes induced by CRISPR-Cas9. Nucleic Acids Res. 2021, 49, 8732–8742. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Greene, E.C. DNA Repair Pathway Choices in CRISPR-Cas9-Mediated Genome Editing. Trends Genet. 2021, 37, 639–656. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Feng, Y.-L.; Xiao, J.-J.; Liu, Q.; Sun, X.-N.; Xiang, J.-F.; Kong, N.; Liu, S.-C.; Chen, G.-Q.; Wang, Y.; et al. Harnessing accurate non-homologous end joining for efficient precise deletion in CRISPR/Cas9-mediated genome editing. Genome Biol. 2018, 19, 1–20. [Google Scholar] [CrossRef]
- Bodai, Z.; Bishop, A.L.; Gantz, V.M.; Komor, A.C. Targeting double-strand break indel byproducts with secondary guide RNAs improves Cas9 HDR-mediated genome editing efficiencies. Nat. Commun. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Song, X.; Zhang, X.-Y.; Xiong, Z.-Q.; Liu, X.-X.; Xia, Y.-J.; Wang, S.-J.; Ai, L.-Z. CRISPR-Cas-mediated gene editing in lactic acid bacteria. Mol. Biol. Rep. 2020, 47, 8133–8144. [Google Scholar] [CrossRef]
- Guo, M.; Ren, K.; Zhu, Y.; Tang, Z.; Wang, Y.; Zhang, B.; Huang, Z. Structural insights into a high fidelity variant of SpCas9. Cell Res. 2019, 29, 183–192. [Google Scholar] [CrossRef]
- Huai, C.; Li, G.; Yao, R.; Zhang, Y.; Cao, M.; Kong, L.; Jia, C.; Yuan, H.; Chen, H.; Lu, D.; et al. Structural insights into DNA cleavage activation of CRISPR-Cas9 system. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, S.; Kim, Y.; Park, J.; Min, S.; Choi, J.W.; Huang, T.P.; Yoon, S.; Liu, D.R.; Kim, H.H. High-throughput analysis of the activities of xCas9, SpCas9-NG and SpCas9 at matched and mismatched target sequences in human cells. Nat. Biomed. Eng. 2020, 4, 111–124. [Google Scholar] [CrossRef]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [Google Scholar] [CrossRef]
- Ding, X.; Seebeck, T.; Feng, Y.; Jiang, Y.; Davis, G.D.; Chen, F. Improving CRISPR-Cas9 Genome Editing Efficiency by Fusion with Chromatin-Modulating Peptides. Cris. J. 2019, 2, 51–63. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Janssen, J.M.; Le Bouteiller, M.; Frock, R.L.; Goncalves, M.A.F.V. Precise and broad scope genome editing based on high-specificity Cas9 nickases. Nucleic Acids Res. 2021, 49, 1173–1198. [Google Scholar] [CrossRef]
- Gopalappa, R.; Suresh, B.; Ramakrishna, S.; Kim, H. Paired D10A Cas9 nickases are sometimes more efficient than individual nucleases for gene disruption. Nucleic Acids Res. 2018, 46, e71. [Google Scholar] [CrossRef]
- Cho, S.W.; Kim, S.; Kim, Y.; Kweon, J.; Kim, H.S.; Bae, S.; Kim, J.-S. Analysis of off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 2014, 24, 132–141. [Google Scholar] [CrossRef]
- Goh, Y.J.; Barrangou, R. Portable CRISPR-Cas9(N) System for Flexible Genome Engineering in Lactobacillus acidophilus, Lactobacillus gasseri, and Lactobacillus paracasei. Appl. Environ. Microbiol. 2021, 87, e02669-20. [Google Scholar] [CrossRef]
- Huston, N.C.; Tycko, J.; Tillotson, E.L.; Wilson, C.J.; Myer, V.E.; Jayaram, H.; Steinberg, B.E. Identification of Guide-Intrinsic Determinants of Cas9 Specificity. Cris. J. 2019, 2, 172–185. [Google Scholar] [CrossRef]
- Doench, J.G.; Hartenian, E.; Graham, D.B.; Tothova, Z.; Hegde, M.; Smith, I.; Sullender, M.; Ebert, B.L.; Xavier, R.J.; Root, D.E. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat. Biotechnol. 2014, 32, 1262–1267. [Google Scholar] [CrossRef]
- Manghwar, H.; Li, B.; Ding, X.; Hussain, A.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas Systems in Genome Editing: Methodologies and Tools for sgRNA Design, Off-Target Evaluation, and Strategies to Mitigate Off-Target Effects. Adv. Sci. 2020, 7, 1902312. [Google Scholar] [CrossRef]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef]
- Ding, W.; Zhang, Y.; Shi, S. Development and Application of CRISPR/Cas in Microbial Biotechnology. Front. Bioeng. Biotechnol. 2020, 8, 711. [Google Scholar] [CrossRef]
- Huang, H.; Song, X.; Yang, S. Development of a RecE/T-Assisted CRISPR-Cas9 Toolbox for Lactobacillus. Biotechnol. J. 2019, 14, 1800690. [Google Scholar] [CrossRef]
- Song, C.W.; Lee, J.; Lee, S.Y. Genome engineering and gene expression control for bacterial strain development. Biotechnol. J. 2015, 10, 56–68. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, W.; Zhou, H.; Tu, Q.; Tang, Y.-J.; Stewart, A.F.; Zhang, Y.; Bian, X. Improved dsDNA recombineering enables versatile multiplex genome engineering of kilobase-scale sequences in diverse bacteria. Nucleic Acids Res. 2022, 50, e15. [Google Scholar] [CrossRef]
- Oh, J.-H.; van Pijkeren, J.-P. CRISPR-Cas9-assisted recombineering in Lactobacillus reuteri. Nucleic Acids Res. 2014, 42, e131. [Google Scholar] [CrossRef]
- Leenay, R.T.; Vento, J.M.; Shah, M.; Martino, M.E.; Leulier, F.; Beisel, C.L. Genome Editing with CRISPR-Cas9 in Lactobacillus plantarum Revealed That Editing Outcomes Can Vary Across Strains and Between Methods. Biotechnol. J. 2019, 14, 1700583. [Google Scholar] [CrossRef]
- Zhou, D.; Jiang, Z.N.; Pang, Q.X.; Zhu, Y.; Wang, Q.; Qi, Q.S. CRISPR/Cas9-Assisted Seamless Genome Editing in Lactobacillus plantarum and Its Application in N-Acetylglucosamine Production. Appl. Environ. Microbiol. 2019, 85, e01367-19. [Google Scholar] [CrossRef]
- Vento, J.M.; Beisel, C.L. Genome Editing with Cas9 in Lactobacilli. Methods Mol. Biol. 2022, 2479, 245–261. [Google Scholar] [CrossRef]
- Fu, J.; Bian, X.; Hu, S.; Wang, H.; Huang, F.; Seibert, P.M.; Plaza, A.; Xia, L.; Mueller, R.; Stewart, A.F.; et al. Full-length RecE enhances linear-linear homologous recombination and facilitates direct cloning for bioprospecting. Nat. Biotechnol. 2012, 30, 440–446. [Google Scholar] [CrossRef]
- Yang, P.; Wang, J.; Qi, Q. Prophage recombinases-mediated genome engineering in Lactobacillus plantarum. Microb. Cell Factories 2015, 14, 1–11. [Google Scholar] [CrossRef]
- Yin, J.; Zhu, H.; Xia, L.; Ding, X.; Hoffmann, T.; Hoffmann, M.; Bian, X.; Mueller, R.; Fu, J.; Stewart, A.F.; et al. A new recombineering system for Photorhabdus and Xenorhabdus. Nucleic Acids Res. 2015, 43, e36. [Google Scholar] [CrossRef]
- Silva, L.A.; Pereira Lopes Neto, J.H.; Cardarelli, H.R. Exopolysaccharides produced by Lactobacillus plantarum: Technological properties, biological activity, and potential application in the food industry. Ann. Microbiol. 2019, 69, 321–328. [Google Scholar] [CrossRef]
- Mohd Yusof, H.; Mohamad, R.; Zaidan, U.H.; Rahman, N.A.A. Sustainable microbial cell nanofactory for zinc oxide nanoparticles production by zinc-tolerant probiotic Lactobacillus plantarum strain TA4. Microb. Cell Factories 2020, 19, 1–7. [Google Scholar] [CrossRef]
- Huang, Y.; You, C.; Liu, Z. Cloning of D-lactate dehydrogenase genes of Lactobacillus delbrueckii subsp bulgaricus and their roles in D-lactic acid production. 3 Biotech 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Tian, X.W.; Liu, X.H.; Zhang, Y.F.; Chen, Y.; Hang, H.F.; Chu, J.; Zhuang, Y.P. Metabolic engineering coupled with adaptive evolution strategies for the efficient production of high-quality L-lactic acid by Lactobacillus paracasei. Bioresour. Technol. 2021, 323, 124549. [Google Scholar] [CrossRef]
- Wang, G.Q.; Yu, H.I.; Feng, X.; Tang, H.Y.; Xiong, Z.Q.; Xia, Y.J.; Ai, L.Z.; Song, X. Specific bile salt hydrolase genes in Lactobacillus plantarum AR113 and relationship with bile salt resistance. Lwt-Food Sci. Technol. 2021, 145, 111208. [Google Scholar] [CrossRef]
- Song, X.; Huang, H.; Xiong, Z.; Ai, L.; Yang, S. CRISPR-Cas9(D10A) Nickase-Assisted Genome Editing in Lactobacillus casei. Appl. Environ. Microbiol. 2017, 83, e01259-17. [Google Scholar] [CrossRef]
- Li, F.; Mei, Z.; Ju, N.; Sui, L.; Fan, X.; Wang, Z.; Li, J.; Jiang, Y.; Cui, W.; Shan, Z.; et al. Evaluation of the immunogenicity of auxotrophic Lactobacillus with CRISPR-Cas9D10A system-mediated chromosomal editing to express porcine rotavirus capsid protein VP4. Virulence 2022, 13, 1315–1330. [Google Scholar] [CrossRef]
- Stout, E.A.; Sanozky-Dawes, R.; Goh, Y.J.; Crawley, A.B.; Klaenhammer, T.R.; Barrangou, R. Deletion-based escape of CRISPR-Cas9 targeting in Lactobacillus gasseri. Microbiol. -Sgm 2018, 164, 1098–1111. [Google Scholar] [CrossRef]
- Myrbraten, I.S.; Wiull, K.; Salehian, Z.; Havarstein, L.S.; Straume, D.; Mathiesen, G.; Kjos, M. CRISPR Interference for Rapid Knockdown of Essential Cell Cycle Genes in Lactobacillus plantarum. Msphere 2019, 4, e00007-19. [Google Scholar] [CrossRef]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef]
- Thakore, P.I.; D’Ippolito, A.M.; Song, L.; Safi, A.; Shivakumar, N.K.; Kabadi, A.M.; Reddy, T.E.; Crawford, G.E.; Gersbach, C.A. Highly specific epigenome editing by CRISPR-Cas9 repressors for silencing of distal regulatory elements. Nat. Methods 2015, 12, 1143–1149. [Google Scholar] [CrossRef]
- Pulecio, J.; Verma, N.; Mejia-Ramirez, E.; Huangfu, D.; Raya, A. CRISPR/Cas9-Based Engineering of the Epigenome. Cell Stem Cell 2017, 21, 431–447. [Google Scholar] [CrossRef]
- Xu, X.; Oi, L.S. A CRISPR-dCas Toolbox for Genetic Engineering and Synthetic Biology. J. Mol. Biol. 2019, 431, 34–47. [Google Scholar] [CrossRef]
- Wang, H.; La Russa, M.; Qi, L.S. CRISPR/Cas9 in Genome Editing and Beyond. Annu. Rev. Biochem. 2016, 85, 227–264. [Google Scholar] [CrossRef]
- Gilbert, L.A.; Horlbeck, M.A.; Adamson, B.; Villalta, J.E.; Chen, Y.; Whitehead, E.H.; Guimaraes, C.; Panning, B.; Ploegh, H.L.; Bassik, M.C.; et al. Genome-Scale CRISPR-Mediated Control of Gene Repression and Activation. Cell 2014, 159, 647–661. [Google Scholar] [CrossRef]
- Saifaldeen, M.; Al-Ansari, D.E.; Ramotar, D.; Aouida, M. CRISPR FokI Dead Cas9 System: Principles and Applications in Genome Engineering. Cells 2020, 9, 2518. [Google Scholar] [CrossRef]
- Zheng, Y.; Su, T.; Qi, Q. Microbial CRISPRi and CRISPRa Systems for Metabolic Engineering. Biotechnol. Bioprocess Eng. 2019, 24, 579–591. [Google Scholar] [CrossRef]
- Thomas, C.M.; Hong, T.; van Pijkeren, J.P.; Hemarajata, P.; Trinh, D.V.; Hu, W.; Britton, R.A.; Kalkum, M.; Versalovic, J. Histamine Derived from Probiotic Lactobacillus reuteri Suppresses TNF via Modulation of PKA and ERK Signaling. PLoS ONE 2012, 7, e31951. [Google Scholar] [CrossRef]
- Yogeswara, I.B.A.; Maneerat, S.; Haltrich, D. Glutamate Decarboxylase from Lactic Acid Bacteria-A Key Enzyme in GABA Synthesis. Microorganisms 2020, 8, 1923. [Google Scholar] [CrossRef]
- Gong, L.C.; Ren, C.; Xu, Y. Deciphering the crucial roles of transcriptional regulator GadR on gamma-aminobutyric acid production and acid resistance in Lactobacillus brevis. Microb. Cell Factories 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Gong, L.C.; Ren, C.; Xu, Y. GlnR Negatively Regulates Glutamate-Dependent Acid Resistance in Lactobacillus brevis. Appl. Environ. Microbiol. 2020, 86, e02615-19. [Google Scholar] [CrossRef]
- Perez-Lopez, A.; Behnsen, J.; Nuccio, S.-P.; Raffatellu, M. Mucosal immunity to pathogenic intestinal bacteria. Nat. Rev. Immunol. 2016, 16, 135–148. [Google Scholar] [CrossRef]
- Iwasaki, A. Exploiting Mucosal Immunity for Antiviral Vaccines. Annu. Rev. Immunol. 2016, 34, 575–608. [Google Scholar] [CrossRef]
- LeCureux, J.S.; Dean, G.A. Lactobacillus Mucosal Vaccine Vectors: Immune Responses against Bacterial and Viral Antigens. Msphere 2018, 3, e00061-18. [Google Scholar] [CrossRef]
- Tang, B.; Li, J.; Li, T.; Xie, Y.; Guan, W.; Zhao, Y.; Yang, S.; Liu, M.; Xu, D. Vaccines as a Strategy to Control Trichinellosis. Front. Microbiol. 2022, 13, 857786. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, B.; Huang, H.-B.; Li, J.-Y.; Pan, T.-X.; Tang, Y.; Shi, C.-W.; Chen, H.-L.; Wang, N.; Yang, G.-L.; et al. Immunoprotective effects of invasive Lactobacillus plantarum delivered nucleic acid vaccine coexpressing Trichinella spiralis CPF1 and murine interleukin-4. Vet. Parasitol. 2021, 298, 109556. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, J.; Zhang-Ding, Z.; Xin, C.; Liu, M.; Wang, Y.; Ai, C.; Hu, J. In-depth assessment of the PAM compatibility and editing activities of Cas9 variants. Nucleic Acids Res. 2021, 49, 8785–8795. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Goh, Y.J.; Pan, M.C.; Sanozky-Dawes, R.; Barrangou, R. Genome editing using the endogenous type I CRISPR-Cas system in Lactobacillus crispatus. Proc. Natl. Acad. Sci. USA 2019, 116, 15774–15783. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactobacillus Species | Function | Reference |
|---|---|---|
| Lactobacillus rhamnosus | Inhibiting cell inflammation and apoptosis | [26] |
| L. fermentum | Antifatigue and antioxidation | [27] |
| L. paracasei | Antitumor | [28] |
| L. johnsonii | Modulating intestinal environment and improving gut development | [29] |
| L. pentosus | Inhibit intestinal pathogenic bacteria | [30] |
| L. casei | Increasing antioxidative capacity | [31] |
| L. helveticus | Cholesterol-lowering activity | [32] |
| Species | Tools | Plasmids | DNA Repair System | Gene Editing Type | Efficiency | References |
|---|---|---|---|---|---|---|
| Lactobacillus plantarum | CRISPR/Cas9 system | pLdbsh1, pLdbsh2, pLdbsh3, pLdbsh4 | HDR | gene knock-out | Success | [77] |
| L. paracasei | CRISPR/Cas9 system | pNcas-ΔldhL1, pNcas-ΔldhD-ldhL1 | HDR | gene knock-out, gene knock-in | Success | [76] |
| L. casei | CRISPR/Cas9D10A system | pLCNICK | HDR | gene deletion and insertion | 25–62% | [78] |
| L. acidophilus | CRISPR/Cas9D10A system | pLbCas9N (pTRK1204 (rafE)), pTRK1205 (lacS), pTRK1254 (ltaS), pTRK1255 (mCherry) | HDR | gene knock-out | 100% | [57] |
| L. gasseri | CRISPR/Cas9D10A system | pLbCas9N (pTRK1256 (2crr) | HDR | gene knock-out | 100% | [57] |
| L. paracasei | CRISPR/Cas9D10A system | pLbCas9N (pTRK1257 (glgA)) | HDR | gene knock-out | 100% | [57] |
| L. paracasei | CRISPR/Cas9D10A system | pLCNICK | HDR | gene knock-out | Success | [79] |
| L. plantarum | CRISPRi | pSIP-SH-dCas9, pSgRNA | HDR | gene knockdown | Success | [81] |
| L. plantarum | RecE/T-assisted CRISPR/Cas9 system | pHSP02, pHSP04 | HDR | gene deletion, gene replacement/insertion | 80–100% | [63] |
| L. brevis | RecE/T-assisted CRISPR/Cas9 system | pHSB01, pHSB04, pHSB05 | HDR | gene deletion, gene knock-out | 75–100% | [63] |
| L. reuteri | CRISPR/Cas9-assisted ssDNA recombineering | pVPL3004, pVPL3017 | HDR | gene mutation, codon saturation mutagenesis | 90–100% | [66] |
| L. plantarum | CRISPR/Cas9-assisted ssDNA recombineering | pCas9_RSR | HDR | point mutations | 100% | [67] |
| L. plantarum | CRISPR/Cas9-assisted ssDNA recombineering | pCas9_RSR | HDR | silent mutation | Success | [67] |
| L. plantarum | CRISPR/Cas9-assisted ssDNA recombineering | pCas9_RSR | HDR | a complete gene deletion | Success | [67] |
| L. plantarum | CRISPR/Cas9-assisted dsDNA recombineering | pSIP-C9 (nagB), pSIP-C9 (lox), p411-RecT, p411-RecT-Dam | HDR | gene knock-out, insertion, and point mutation | 53.3%, 58.3%, 62.5% | [68] |
| L. plantarum | CRISPR/Cas9-assisted dsDNA recombineering | PCB578, pCB591 | HDR | gene mutation | Success | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Y.; Zhang, C.; Li, T.; Jin, F.-J.; Sung, Y.-J.; Oh, H.-M.; Lee, H.-G.; Jin, L. Development and Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus. Int. J. Mol. Sci. 2022, 23, 12852. https://doi.org/10.3390/ijms232112852
Mu Y, Zhang C, Li T, Jin F-J, Sung Y-J, Oh H-M, Lee H-G, Jin L. Development and Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus. International Journal of Molecular Sciences. 2022; 23(21):12852. https://doi.org/10.3390/ijms232112852
Chicago/Turabian StyleMu, Yulin, Chengxiao Zhang, Taihua Li, Feng-Jie Jin, Yun-Ju Sung, Hee-Mock Oh, Hyung-Gwan Lee, and Long Jin. 2022. "Development and Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus" International Journal of Molecular Sciences 23, no. 21: 12852. https://doi.org/10.3390/ijms232112852
APA StyleMu, Y., Zhang, C., Li, T., Jin, F.-J., Sung, Y.-J., Oh, H.-M., Lee, H.-G., & Jin, L. (2022). Development and Applications of CRISPR/Cas9-Based Genome Editing in Lactobacillus. International Journal of Molecular Sciences, 23(21), 12852. https://doi.org/10.3390/ijms232112852