
Biophysical Characterization of Adeno-Associated Virus Vectors Using Ion-Exchange Chromatography Coupled to Light Scattering Detectors



Abstract
1. Introduction
2. Results and Discussion
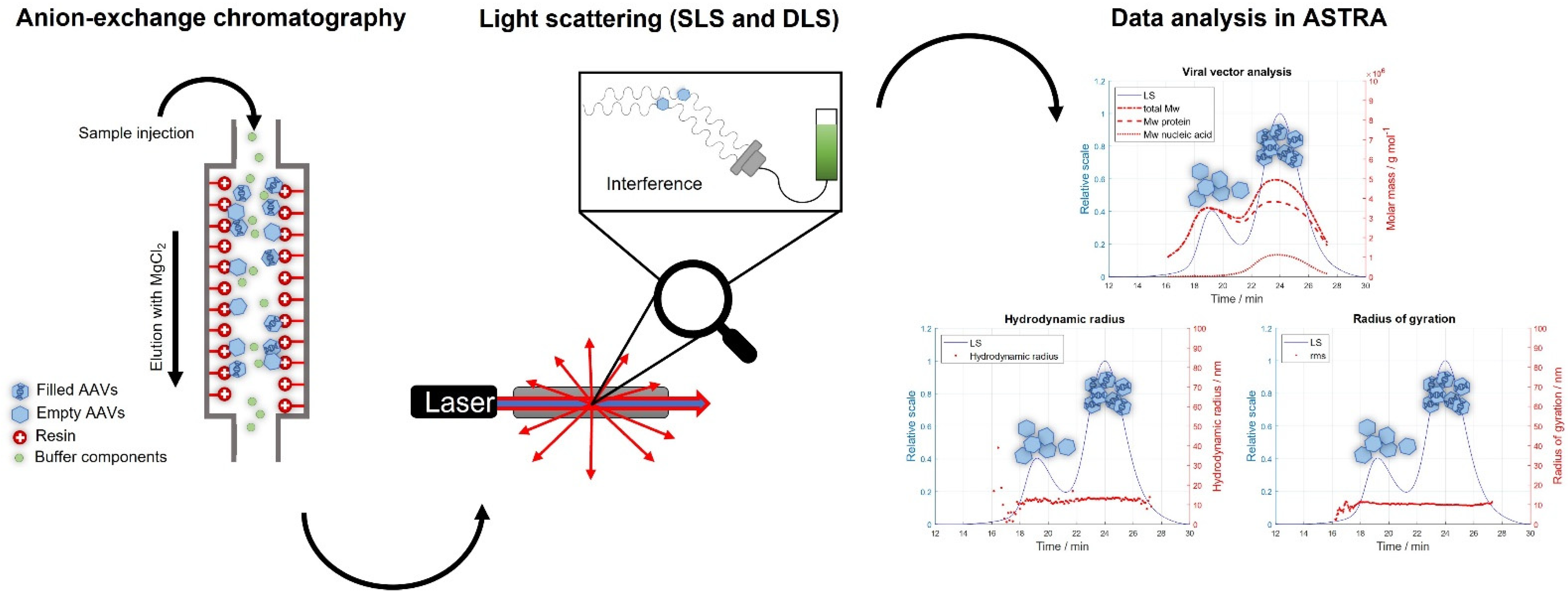
2.1. IEC-MALS Development
2.2. Determination of the UV Extinction Coefficients
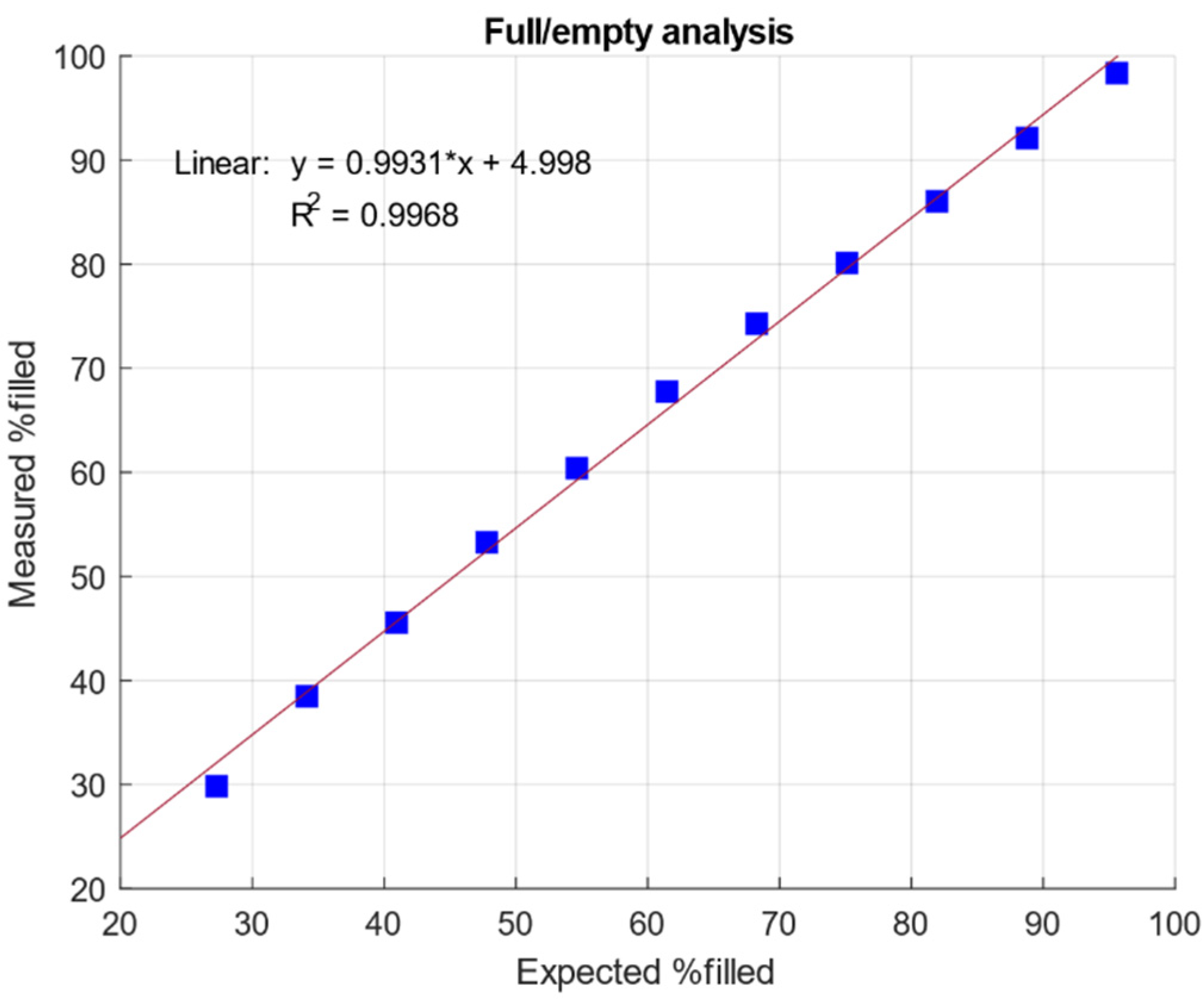
2.3. Comparison of %Filled AAV Capsids to Orthogonal Methods
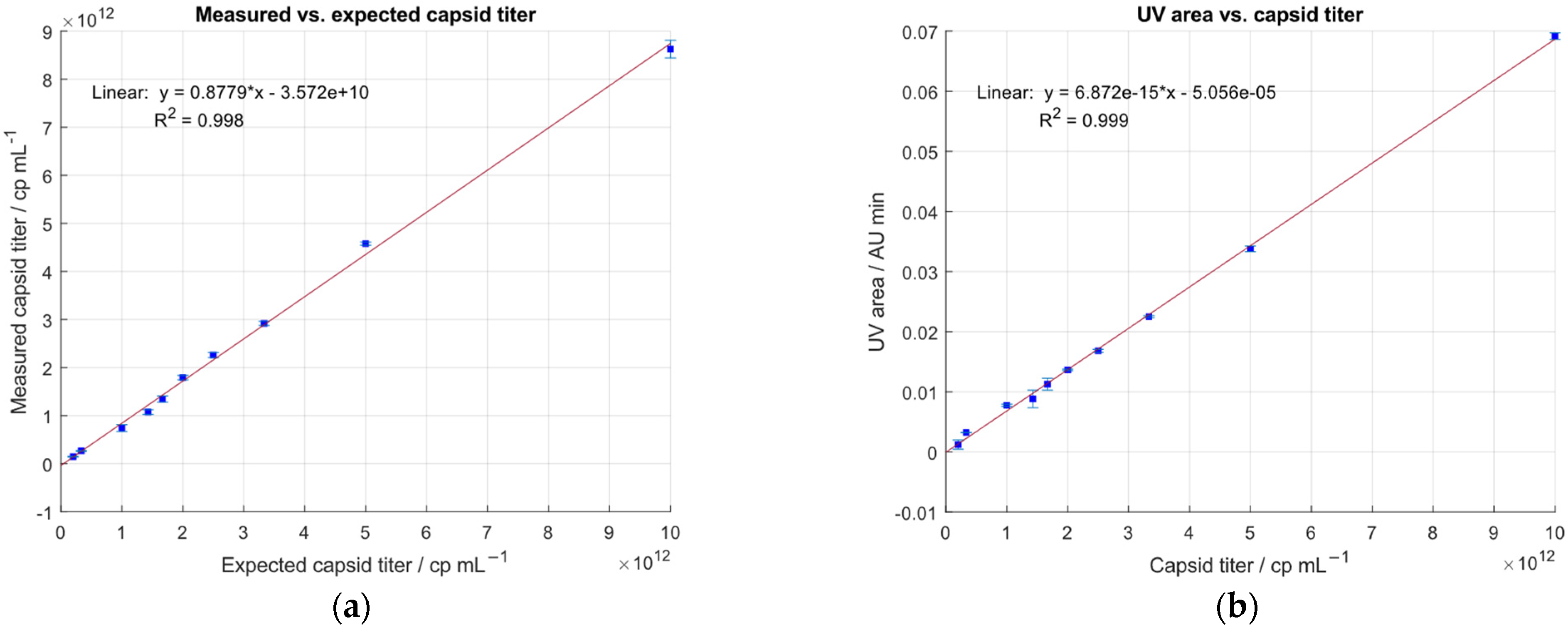
2.4. Linearity of the IEC-MALS Method
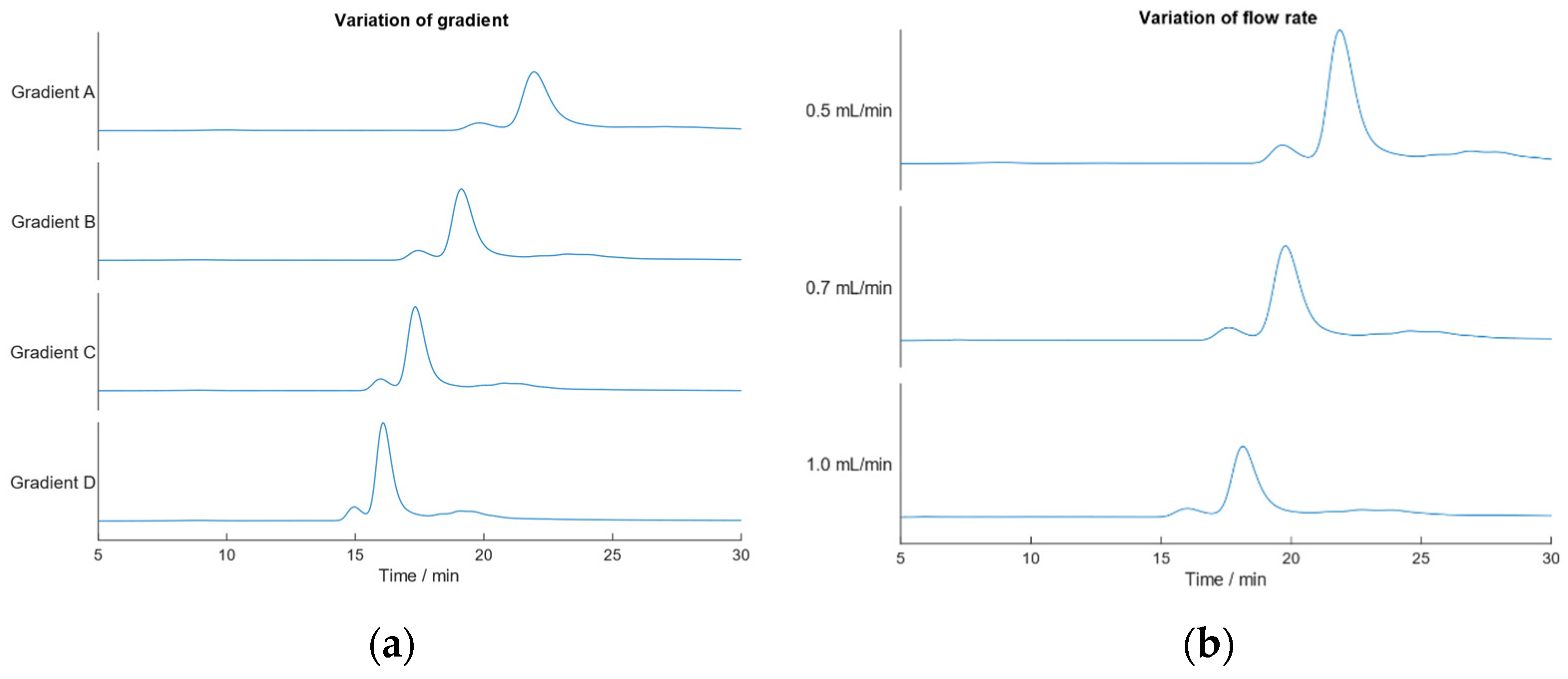
2.5. Robustness of the IEC-MALS Method
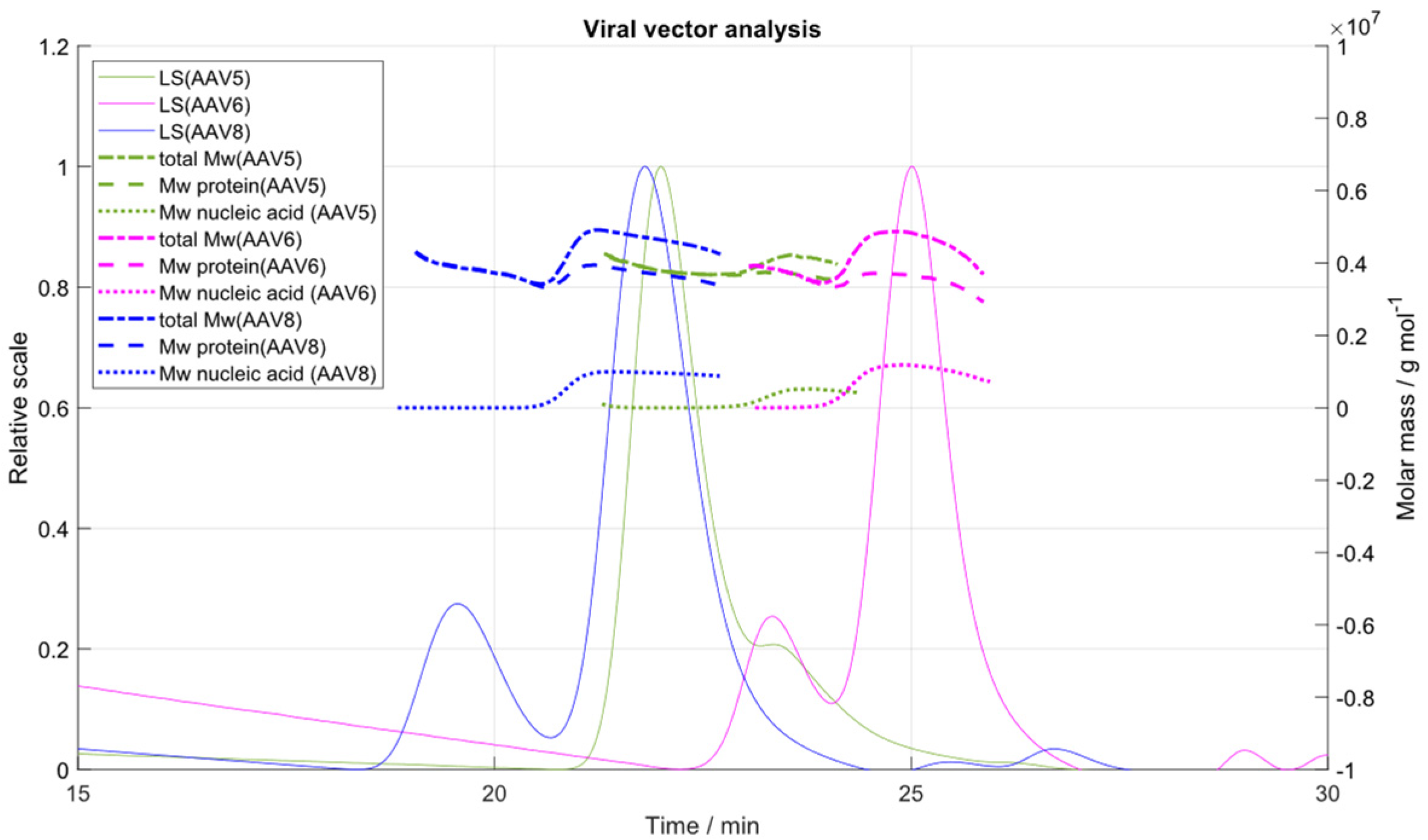
2.6. Application of the IEC-MALS Method to Other Serotpyes
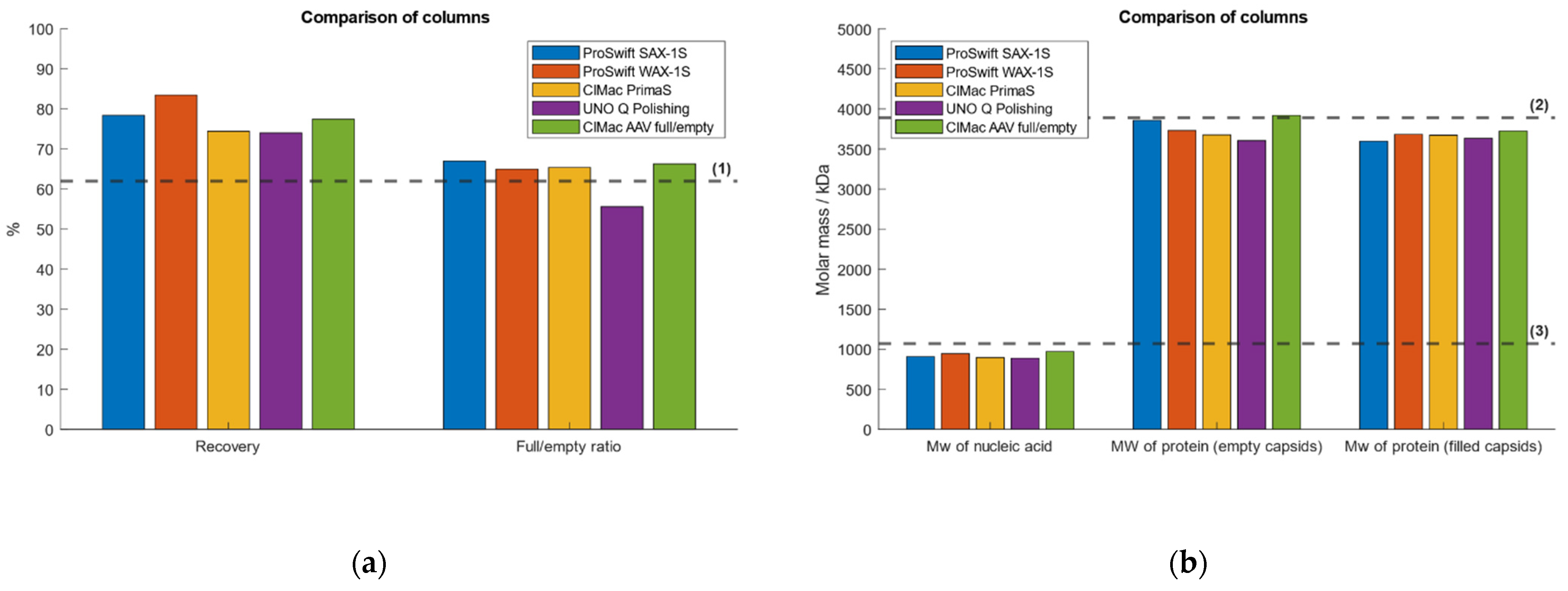
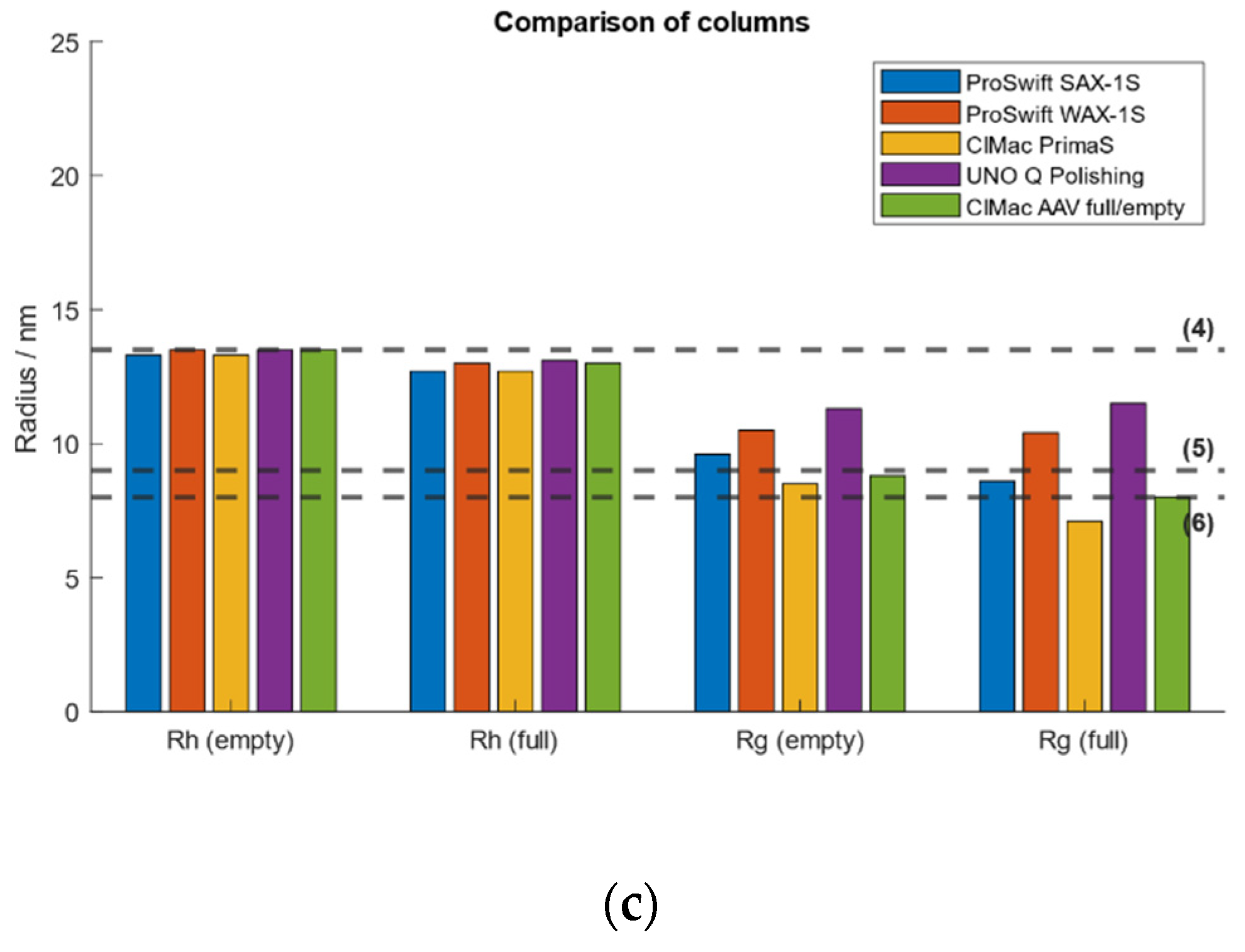
2.7. Test of Different Weak and Strong AEX Columns
2.8. Salt Gradient vs. pH Gradient
3. Materials and Methods
3.1. Samples and Reagents
3.2. Instrument Configuration
3.3. Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Chandler, R.J.; Sands, M.; Venditti, C.P. rAAV integration and genotoxicity: Insights from animal models. Hum. Gene Ther. 2017, 28, 4. [Google Scholar] [CrossRef]
- Maestro, S.; Weber, N.D.; Zabaleta, N.; Aldabe, R.; Gonzalez-Aseguinolaza, G. Novel vectors and approaches for gene therapy in liver diseases. JHEP Rep. 2021, 3, 100300. [Google Scholar] [CrossRef] [PubMed]
- Aponte-Ubillus, J.; Barajas, D.; Peltier, J.; Bardliving, C.; Shamlou, P.; Gold, D. Molecular design for recombinant adeno-associated virus (rAAV) vector production. Appl. Microbiol. Biotechnol. 2018, 102, 1045–1054. [Google Scholar] [CrossRef]
- Wang, C.; Mulagapati, S.H.R.; Chen, Z.; Du, J.; Zhao, X.; Xi, G.; Chen, L.; Linke, T.; Gao, C.; Schmelzer, A.E.; et al. Developing an Anion Exchange Chromatography Assay for Determining Empty and Full Capsid Contents in AAV6.2. Mol. Ther.—Methods Clin. Dev. 2019, 15, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Oyama, H.; Ishii, K.; Maruno, T.; Torisu, T.; Uchiyama, S. Characterization of Adeno-Associated Virus Capsid Proteins with Two Types of VP3-Related Components by Capillary Gel Electrophoresis and Mass Spectrometry. Hum. Gene Ther. 2021, 32, 21–22. [Google Scholar] [CrossRef]
- Wang, D.; Tai, P.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef]
- Wright, F.J. Quality Control Testing, Characterization and Critical Quality Attributes of AAV Vectors Used for Human Gene Therapy. Biotechnol. J. 2020, 16, 2000022. [Google Scholar] [CrossRef]
- Khatwani, S.L.; Pavlova, A.; Pirot, Z. Anion-exchange HPLC assay for separation and quantification of empty and full capsids in multiple adeno-associated virus serotypes. Mol. Ther.—Methods Clin. Dev. 2021, 21, 548–558. [Google Scholar] [CrossRef]
- EU/3/12/1091: Orphan Designation for the Treatment of Beta Thalassaemia Intermedia and Major. Available online: https://www.ema.europa.eu/en/medicines/human/orphan-designations/eu3121091 (accessed on 17 October 2022).
- FDA Approves First Cell-Based Gene Therapy to Treat Adult and Pediatric Patients with Beta-Thalassemia Who Require Regular Blood Transfusions. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-cell-based-gene-therapy-treat-adult-and-pediatric-patients-beta-thalassemia-who (accessed on 17 October 2022).
- Wileys, J. Gene Therapy Clinical Trials Worldwide. Available online: https://a873679.fmphost.com/fmi/webd/GTCT (accessed on 14 March 2022).
- Nam, H.-J.; Lane, M.D.; Padron, E.; Gurda, B.; McKenna, R.; Kohlbrenner, E.; Aslanidi, G.; Byrne, B.; Muzyczka, N.; Zolotukhin, S.; et al. Structure of Adeno-Associated Virus Serotype 8, a Gene Therapy Vector. J. Virol. 2007, 81, 12260–12271. [Google Scholar] [CrossRef]
- Wright, F.J. Product-Related Impurities in Clinical-Grade Recombinant AAV Vectors: Characterization and Risk Assessment. Biomedicines 2014, 2, 80–97. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and Dynamics of Adeno-Associated Virus Serotype 1 VP1-Unique N-Terminal Domain and Its Role in Capsid Trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, G.; Shalaev, E.; Karami, T.; Cunningham, J.; Slater, N.; Rivers, H. Pharmaceutical Development of AAV-Based Gene Therapy Products for the Eye. Pharm. Res. 2019, 36, 29. [Google Scholar] [CrossRef]
- Gimpel, A.L.; Katsikis, G.; Sha, S.; Maloney, A.J.; Hong, M.S.; Nguyen, T.N.T.; Wolfrum, J.; Springs, S.L.; Sinskey, A.J.; Manalis, S.R.; et al. Analytical methods for process and product characterization of recombinant adeno-associated virus-based gene therapies. Mol. Ther.—Methods Clin. Dev. 2021, 20, 740–754. [Google Scholar] [CrossRef]
- McIntosh, N.L.; Berguig, G.Y.; Karim, O.A.; Cortesio, C.L.; De Angelis, R.; Khan, A.A.; Gold, D.; Maga, J.A.; Bhat, V.S. Comprehensive characterization and quantification of adeno associated vectors by size exclusion chromatography and multi angle light scattering. Sci. Rep. 2021, 11, 3012. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Chen, W.-C.; Argento, C.; Clarner, P.; Bhatt, V.; Dickerson, R.; Bou-Assaf, G.; Bakhshayeshi, M.; Lu, X.; Bergelson, S.; et al. Analytical Strategies for Quantification of Adeno-Associated Virus Empty Capsids to Support Process Development. Hum. Gene Ther. Methods 2019, 30, 144–152. [Google Scholar] [CrossRef]
- Ye, H. Simultaneous determination of protein aggregation, degradation, and absolute molecular weight by size exclusion chromatography-multiangle laser light scattering. Anal. Biochem. 2006, 356, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Amartely, H.; Avraham, O.; Friedler, A.; Livnah, O.; Lebendiker, M. Coupling Multi Angle Light Scattering to Ion Exchange chromatography (IEX-MALS) for protein characterization. Sci. Rep. 2018, 8, 6907. [Google Scholar] [CrossRef]
- Wyatt, P. Light scattering and the absolute characterization of macromolecules. Anal. Chim. Acta 1993, 272, 1–40. [Google Scholar] [CrossRef]
- Sahin, E.; Roberts, C.J. Size-exclusion chromatography with multi-angle light scattering for elucidating protein aggregation mechanisms. Methods Mol. Biol. 2012, 899, 403–423. [Google Scholar] [CrossRef]
- Weihong, Q.; Mingxi, W.; Yaqing, W.; Ruian, X. Scalable Downstream Strategies for Purification of Recombinant Adeno- Associated Virus Vectors in Light of the Properties. Curr. Pharm. Biotechnol. 2015, 16, 684–695. [Google Scholar] [CrossRef]
- Fekete, S.; Beck, A.; Veuthey, J.-L.; Guillarme, D. Ion-exchange chromatography for the characterization of biopharmaceuticals. J. Pharm. Biomed. Anal. 2015, 113, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Tsonev, L.I.; Hirsh, A.G. Theory and applications of a novel ion exchange chromatographic technology using controlled pH gradients for separating proteins on anionic and cationic stationary phases. J. Chromatogr. A 2008, 1200, 166–182. [Google Scholar] [CrossRef]
- Zhao, H.; Brown, P.H.; Schuck, P. On the Distribution of Protein Refractive Index Increments. Biophys. J. 2011, 100, 2309–2317. [Google Scholar] [CrossRef] [PubMed]
- Some, D.; Amartely, H.; Tsadok, A.; Lebendiker, M. Characterization of Proteins by Size-Exclusion Chromatography Coupled to Multi-Angle Light Scattering (SEC-MALS). J. Vis. Exp. JoVE 2019, 8, 6907. [Google Scholar] [CrossRef]
- Larkin, M.; Wyatt, P. Light scattering techniques and their application to formulation and aggregation concerns. In Formulation and Process Development Strategies for Manufacturing of a Biopharmaceutica; Wiley & Sons: New York, NY, USA, 2010; pp. 269–305. [Google Scholar]
- Patel, B.A.; Gospodarek, A.; Larkin, M.; Kenrick, S.A.; Haverick, M.A.; Tugcu, N.; Brower, M.A.; Richardson, D.D. Multi-angle light scattering as a process analytical technology measuring real-time molecular weight for downstream process control. mAbs 2018, 10, 945–950. [Google Scholar] [CrossRef]
- TOSOH Corporation. Light Scattering for Determination of Molecular Weight and Radius of Gyration. [White Paper]. 2022. Available online: https://www.separations.eu.tosohbioscience.com/OpenPDF.aspx?path=~/File%20Library/TBG/Products%20Download/Whitepaper/w20i16a.pdf (accessed on 7 June 2022).
- Gagnon, P.; Goricar, B.; Mencin, N.; Zvanut, T.; Peljhan, S.; Leskovec, M.; Strancar, A. Multiple-Monitor HPLC Assays for Rapid Process Development, In-Process Monitoring, and Validation of AAV Production and Purification. Pharmaceutics 2021, 13, 113. [Google Scholar] [CrossRef]
- Porterfield, J.Z.; Zlotnick, A. A simple and general method for determining the protein and nucleic acid content of viruses by UV absorbance. Virology 2010, 407, 281–288. [Google Scholar] [CrossRef]
- Some, D.; Johann, C. Characterization of Nanopharmaceuticals with Field-Flow Fractionation and Light Scattering (FFF-MALS-DLS). [White Paper]. Available online: https://www.controlledreleasesociety.org/news/characterization-nano-pharmaceuticals-field-flow-fractionation-and-light-scattering-fff-mals (accessed on 7 June 2022).
- Lock, M.; Alvira, M.R.; Wilson, J.M. Analysis of Particle Content of Recombinant Adeno-Associated Virus Serotype 8 Vectors by Ion-Exchange Chromatography. Hum. Gene Ther. Methods 2012, 23, 56–64. [Google Scholar] [CrossRef]
- Rose, J.A.; Maizel, J.V.; Inman, J.K.; Shatkin, A.J. Structural Proteins of Adenovirus-Associated Viruses. J. Virol. 1971, 8, 766–770. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gradient A | Gradient B | Gradient C | Gradient D | Flow Rate 0.5 mL min−1 | Flow Rate 0.7 mL min−1 | Flow Rate 1.0 mL min−1 | |
|---|---|---|---|---|---|---|---|
| Capsid Titer ELISA/cp mL−1 | 1.00 × 1013 | 1.00 × 1013 | 1.00 × 1013 | 1.00 × 1013 | 1.00 × 1013 | 1.00 × 1013 | 1.00 × 1013 |
| Measured Capsid Titer/cp mL−1 | 8.23 × 1012 | 8.50 × 1012 | 8.35 × 1012 | 8.40 × 1012 | 8.21 × 1012 | 8.75 × 1012 | 9.25 × 1012 |
| RSD/% | 1.5 | 0.3 | 0.6 | 0.8 | 4.3 | 0.8 | 0.3 |
| Recovery/% | 82 | 85 | 84 | 84 | 82 | 88 | 92 |
| Expected %full | 62 | 62 | 62 | 62 | 62 | 62 | 62 |
| Measured %full | 66 | 66 | 66 | 67 | 65 | 59 | 59 |
| Difference %full/% | 6 | 6 | 7 | 7 | 5 | 4 | 5 |
| Expected Mw Nucleic Acid/kDa | 1070 | 1070 | 1070 | 1070 | 1070 | 1070 | 1070 |
| Measured Mw Nucleic Acid/kDa | 959 | 949 | 956 | 951 | 936 | 904 | 902 |
| RSD/% | 0.1 | 0.7 | 0.8 | 0.7 | 1.9 | 0.9 | 0.1 |
| Difference MW Nucleic Acid/% | 10 | 11 | 11 | 11 | 13 | 16 | 16 |
| Measured Mw Protein (Empty)/kDa | 3842 | 3798 | 3809 | 3819 | 3750 | 3652 | 3573 |
| RSD/% | 1.7 | 0.5 | 0.5 | 0.5 | 1.0 | 0.8 | 0.4 |
| Measured Mw Protein (Full)/kDa | 3713 | 3690 | 3694 | 3695 | 3657 | 3659 | 3655 |
| RSD/% | 0.3 | 0.3 | 0.4 | 0.3 | 0.1 | 0.3 | 0.0 |
| UV 260/280 Ratio (Empty) | 0.69 | 0.68 | 0.68 | 0.65 | 0.66 | 0.46 | 0.74 |
| UV 260/280 Ratio (Full) | 1.34 | 1.32 | 1.32 | 1.33 | 1.34 | 1.31 | 1.33 |
| AAV5 | AAV6 | AAV8 | |
|---|---|---|---|
| Capsid Titer ELISA/cp mL−1 | 6.73 × 1013 | 2.12 × 1012 | 1.00 × 1013 |
| Measured Capsid Titer/cp mL−1 | 6.20 × 1013 | 2.28 × 1012 | 7.74 × 1012 |
| RSD/% | 3.3 | 1.4 | 2.1 |
| Recovery/% | 92 | 107 | 77 |
| Expected %full | 10 | 68 | 62 |
| Measured %full | 12 | 66 | 66 |
| Difference %full/% | 15 | 3 | 7 |
| Expected Mw Nucleic Acid/kDa | 760 | 1240 | 1071 |
| Measured Mw Nucleic Acid/kDa | 457 | 1128 | 970 |
| RSD/% | 4.9 | 1.6 | 0.9 |
| Difference MW Nucleic Acid/% | 40 | 9 | 9 |
| Measured Mw Protein (Empty)/kDa | 3759 | 3850 | 3920 |
| RSD/% | 0.8 | 0.9 | 1.3 |
| Measured Mw Protein (Full)/kDa | 3466 | 3627 | 3724 |
| RSD/% | 3.1 | 0.3 | 0.5 |
| rh (Empty)/nm | 12.5 | 8.4 | 13 |
| RSD/% | 0.3 | 13.5 | 2.3 |
| rh (Full)/nm | 9 | 12 | 14 |
| RSD/% | 10.4 | 0.9 | 0.5 |
| rms (Empty)/nm | 9 | 8 | 9 |
| RSD/% | 22.4 | 35.2 | 11.8 |
| rms (Full)/nm | 9 | 8 | 8 |
| RSD/% | 6.8 | 13.1 | 16.4 |
| Polydispersity | 1.011 | 1.040 | 1.012 |
| RSD/% | 1.0 | 0.3 | 0.2 |
| pH Gradient | Salt Gradient | |
|---|---|---|
| Capsid Titer ELISA/cp mL−1 | 1.00 × 1013 | 1.00 × 1013 |
| Measured Capsid Titer/cp mL−1 | 9.11 × 1012 | 8.87 × 1012 |
| RSD/% | 0.8 | 1.5 |
| Recovery/% | 91 | 89 |
| Expected %full | 62 | 62 |
| Measured %full | 58 | 68 |
| Difference %full/% | 6 | 9 |
| Expected Mw Nucleic Acid/kDa | 1070 | 1070 |
| Measured Mw Nucleic Acid/kDa | 1014 | 986 |
| RSD/% | 0.6 | 1.1 |
| Difference MW Nucleic Acid/% | 5 | 9 |
| Measured Mw Protein (Empty)/kDa | 3359 | 3802 |
| RSD/% | 0.7 | 1.1 |
| Measured Mw Protein (Full)/kDa | 3670 | 3818 |
| RSD/% | 0.3 | 0.3 |
| UV 260/280 Ratio (Empty) | 0.77 | 0.83 |
| UV 260/280 Ratio (Full) | 1.26 | 1.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, C.; Innthaler, B.; Lemmerer, M.; Pletzenauer, R.; Birner-Gruenberger, R. Biophysical Characterization of Adeno-Associated Virus Vectors Using Ion-Exchange Chromatography Coupled to Light Scattering Detectors. Int. J. Mol. Sci. 2022, 23, 12715. https://doi.org/10.3390/ijms232112715
Wagner C, Innthaler B, Lemmerer M, Pletzenauer R, Birner-Gruenberger R. Biophysical Characterization of Adeno-Associated Virus Vectors Using Ion-Exchange Chromatography Coupled to Light Scattering Detectors. International Journal of Molecular Sciences. 2022; 23(21):12715. https://doi.org/10.3390/ijms232112715
Chicago/Turabian StyleWagner, Christina, Bernd Innthaler, Martin Lemmerer, Robert Pletzenauer, and Ruth Birner-Gruenberger. 2022. "Biophysical Characterization of Adeno-Associated Virus Vectors Using Ion-Exchange Chromatography Coupled to Light Scattering Detectors" International Journal of Molecular Sciences 23, no. 21: 12715. https://doi.org/10.3390/ijms232112715
APA StyleWagner, C., Innthaler, B., Lemmerer, M., Pletzenauer, R., & Birner-Gruenberger, R. (2022). Biophysical Characterization of Adeno-Associated Virus Vectors Using Ion-Exchange Chromatography Coupled to Light Scattering Detectors. International Journal of Molecular Sciences, 23(21), 12715. https://doi.org/10.3390/ijms232112715