
The Dual Role of the β2-Adrenoreceptor in the Modulation of IL-17 and IFN-γ Production by T Cells in Multiple Sclerosis

Abstract
:1. Introduction
2. Results
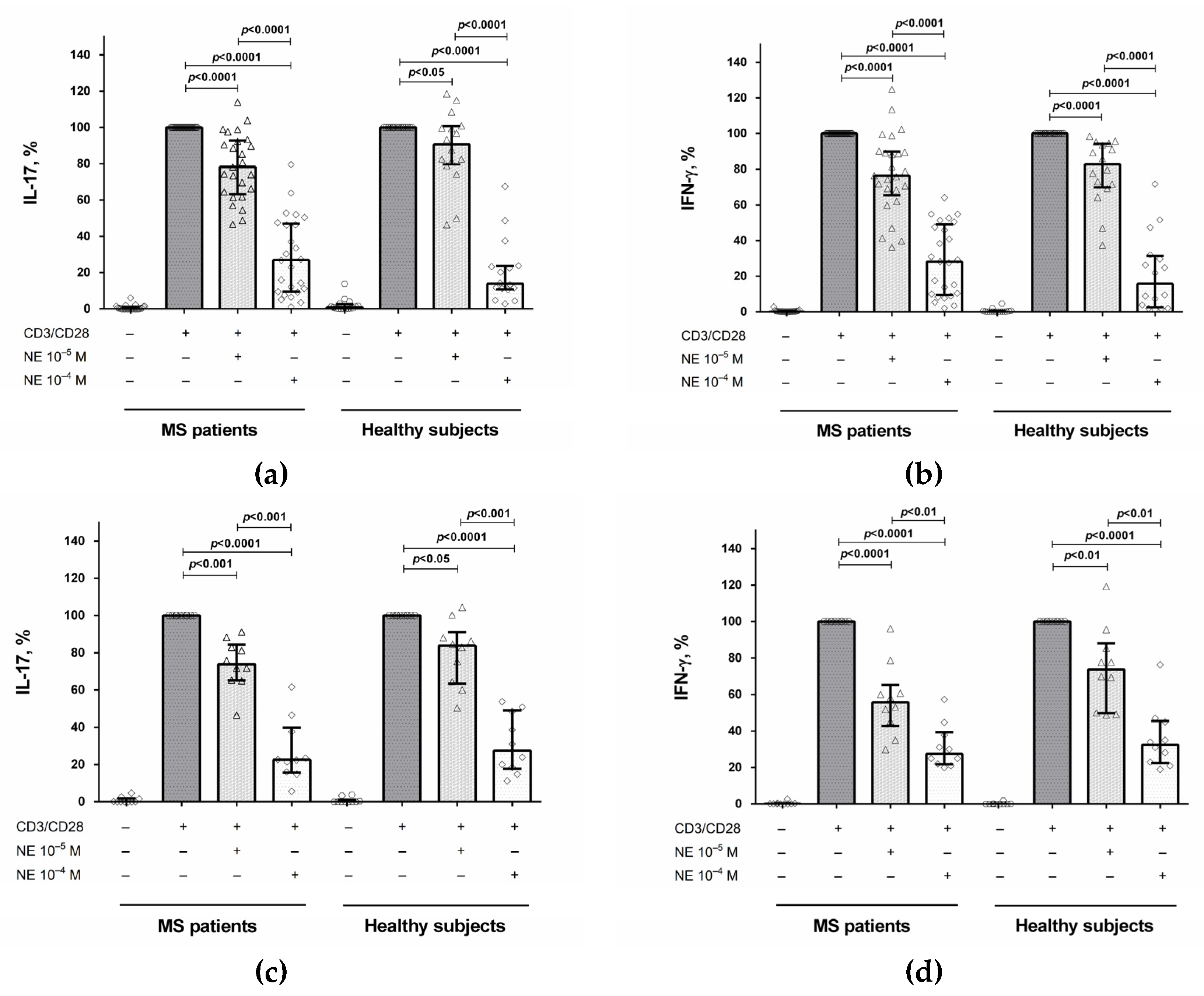
2.1. The Influence of Norepinephrine on IL-17 and IFN-γ Production by CD4+ T Cells and PBMCs
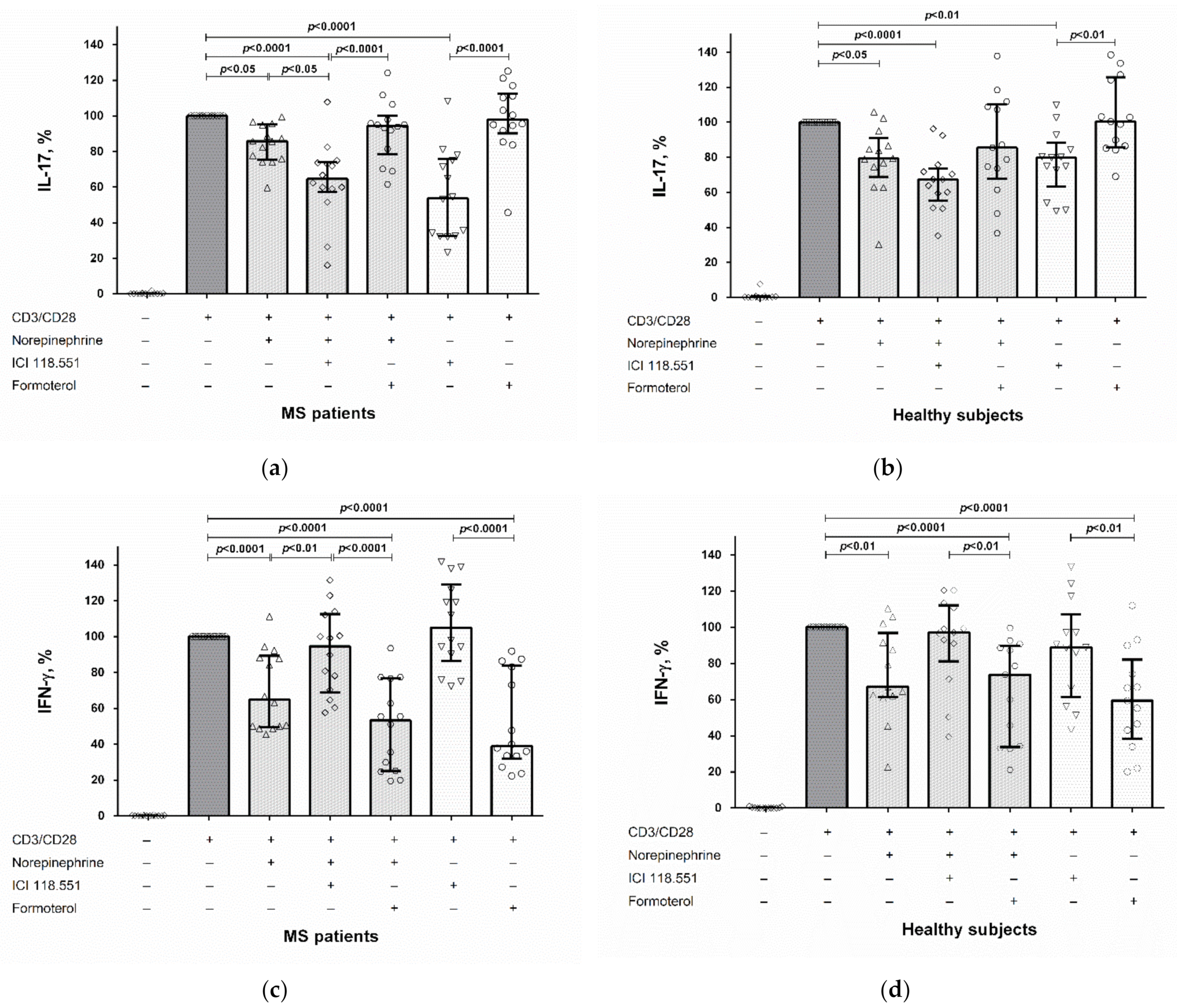
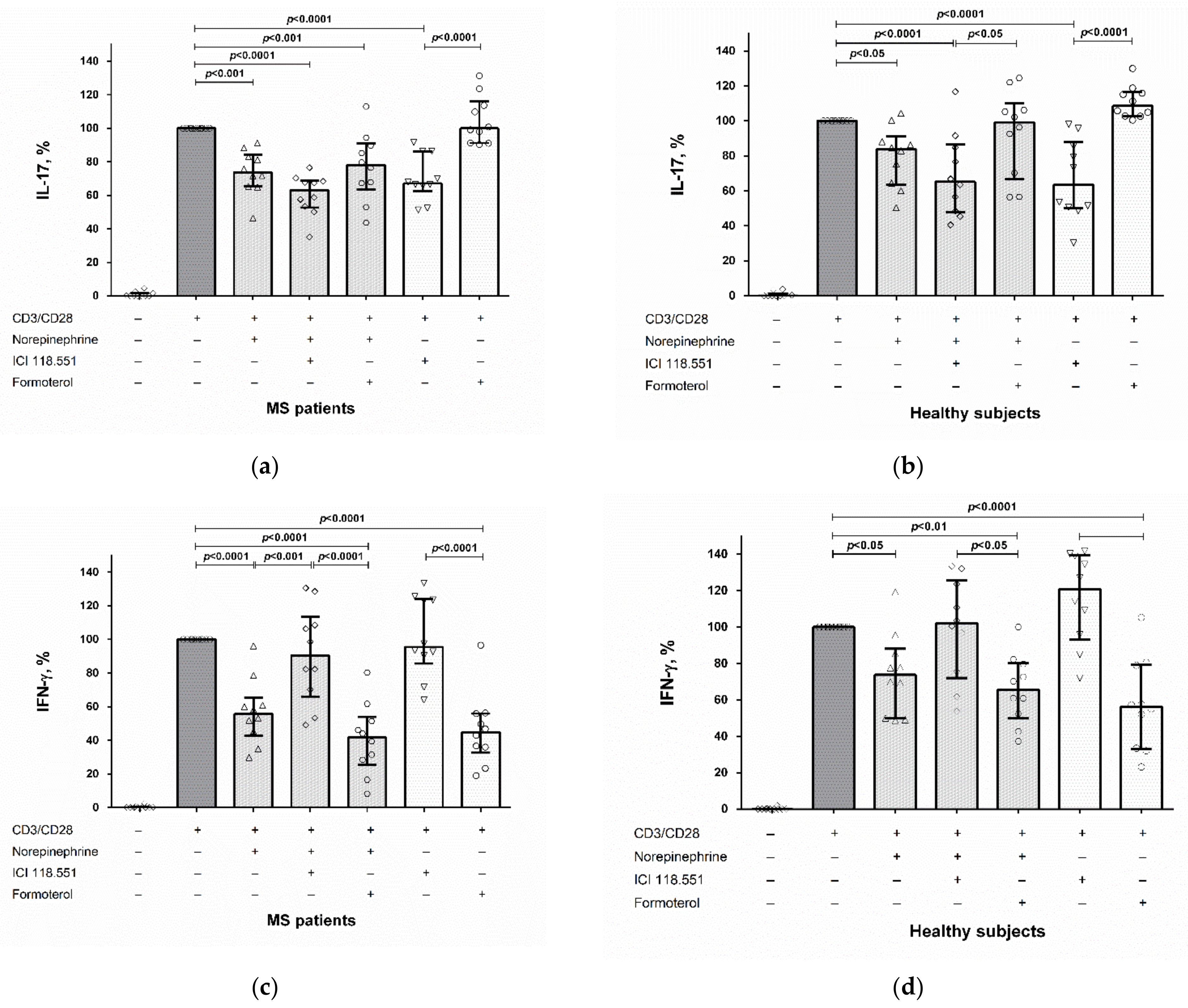
2.2. The Role of the β2-Adrenergic Receptor in the Modulation of IL-17 and IFN-γ Production by CD4+ T Cells and PBMCs
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Epinephrine, Norepinephrine, and 3-Methoxy-4-Hydroxyphenylglycol Evaluation
4.3. Peripheral Blood Mononuclear Cells and CD4+ T Cell Cultures and Stimulation
4.4. Cytokine Evaluation
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carandini, T.; Cercignani, M.; Galimberti, D.; Scarpini, E.; Bozzali, M. The distinct roles of monoamines in multiple sclerosis: A bridge between the immune and nervous systems? Brain Behav. Immun. 2021, 94, 381–391. [Google Scholar] [CrossRef]
- Cosentino, M.; Zaffaroni, M.; Ferrari, M.; Marino, F.; Bombelli, R.; Rasini, E.; Frigo, G.; Ghezzi, A.; Comi, G.; Lecchini, S. Interferon-γ and interferon-β affect endogenous catecholamines in human peripheral blood mononuclear cells: Implications for multiple sclerosis. J. Neuroimmunol. 2005, 162, 112–121. [Google Scholar] [CrossRef]
- Stamoula, E.; Siafis, S.; Dardalas, I.; Ainatzoglou, A.; Matsas, A.; Athanasiadis, T.; Sardeli, C.; Stamoulas, K.; Papazisis, G. Antidepressants on Multiple Sclerosis: A Review of In Vitro and In Vivo Models. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Cosentino, M.; Marino, F. Adrenergic and Dopaminergic Modulation of Immunity in Multiple Sclerosis: Teaching Old Drugs New Tricks? J. Neuroimmune Pharmacol. 2013, 8, 163–179. [Google Scholar] [CrossRef]
- Pilipović, I.; Vujnović, I.; Stojić-Vukanić, Z.; Petrović, R.; Kosec, D.; Nacka-Aleksić, M.; Jasnić, N.; Leposavić, G. Noradrenaline modulates CD4+ T cell priming in rat experimental autoimmune encephalomyelitis: A role for the α1-adrenoceptor. Immunol. Res. 2019, 67, 223–240. [Google Scholar] [CrossRef]
- Sanders, V.M.; Straub, R.H. Norepinephrine, the β-Adrenergic Receptor, and Immunity. Brain Behav. Immun. 2002, 16, 290–332. [Google Scholar] [CrossRef]
- Kohm, A.P.; Sanders, V.M. Norepinephrine and beta 2-adrenergic receptor stimulation regulate CD4+ T and B lymphocyte function in vitro and in vivo. Pharmacol. Rev. 2001, 53, 487–525. [Google Scholar]
- Moser, T.; Akgün, K.; Proschmann, U.; Sellner, J.; Ziemssen, T. The role of TH17 cells in multiple sclerosis: Therapeutic implications. Autoimmun. Rev. 2020, 19, 102647. [Google Scholar] [CrossRef]
- Correale, J.; Halfon, M.J.; Jack, D.; Rubstein, A.; Villa, A. Acting centrally or peripherally: A renewed interest in the central nervous system penetration of disease-modifying drugs in multiple sclerosis. Mult. Scler. Relat. Disord. 2021, 56, 103264. [Google Scholar] [CrossRef]
- Boyko, A.; Melnikov, M.; Zhetishev, R.; Pashenkov, M. The Role of Biogenic Amines in the Regulation of Interaction between the Immune and Nervous Systems in Multiple Sclerosis. Neuroimmunomodulation 2016, 23, 217–223. [Google Scholar] [CrossRef]
- Vujnović, I.; Pilipović, I.; Jasnić, N.; Petrović, R.; Blagojević, V.; Arsenović-Ranin, N.; Stojić-Vukanić, Z.; Djordjević, J.; Leposavić, G. Noradrenaline through β-adrenoceptor contributes to sexual dimorphism in primary CD4+ T-cell response in DA rat EAE model? Cell Immunol. 2019, 336, 48–57. [Google Scholar] [CrossRef]
- Padro, C.J.; Sanders, V.M. Neuroendocrine regulation of inflammation. Semin. Immunol. 2014, 26, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Wahle, M.; Hanefeld, G.; Brunn, S.; Straub, R.H.; Wagner, U.; Krause, A.; Häntzschel, H.; Baerwald, C.G.O. Failure of catecholamines to shift T-cell cytokine responses toward a Th2 profile in patients with rheumatoid arthritis. Arthritis Res. 2006, 8, R138. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Rui, X.-X.; Shi, H.; Qiu, Y.-H.; Peng, Y.-P. Norepinephrine Inhibits Th17 Cells via β2-Adrenergic Receptor (β2-AR) Signaling in a Mouse Model of Rheumatoid Arthritis. Med. Sci. Monit. 2018, 24, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Borger, P.; Hoekstra, Y.; Esselink, M.T.; Postma, D.S.; Zaagsma, J.; Vellenga, E.; Kauffman, H.F. β-Adrenoceptor-mediated Inhibition of IFN-γ, IL-3, and GM-CSF mRNA Accumulation in Activated Human T Lymphocytes Is Solely Mediated by the β2-Adrenoceptor Subtype. Am. J. Respir. Cell Mol. Biol. 1998, 19, 400–407. [Google Scholar] [CrossRef]
- Carvajal Gonczi, C.M.C.; Tabatabaei Shafiei, M.T.; East, A.; Martire, E.; I Maurice-Ventouris, M.H.; Darlington, P.J. Reciprocal modulation of helper Th1 and Th17 cells by the β2-adrenergic receptor agonist drug terbutaline. FEBS J. 2017, 284, 3018–3028. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, A.; Robinson, N.; Palma, G.; Maurea, N.; Desiderio, V.; Botti, G. Can Beta-2-Adrenergic Pathway Be a New Target to Combat SARS-CoV-2 Hyperinflammatory Syndrome?—Lessons Learned From Cancer. Front. Immunol. 2020, 11, 2615. [Google Scholar] [CrossRef]
- Lorton, D.; Bellinger, D.L. Molecular Mechanisms Underlying β-Adrenergic Receptor-Mediated Cross-Talk between Sympathetic Neurons and Immune Cells. Int. J. Mol. Sci. 2015, 16, 5635–5665. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Tai, Y.; Hu, S.; Zhang, M.; Wang, R.; Zhou, W.; Tao, J.; Han, Y.; Wang, Q.; Wei, W. Bidirectional Role of β2-Adrenergic Receptor in Autoimmune Diseases. Front. Pharmacol. 2018, 9, 1313. [Google Scholar] [CrossRef]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An Inventory for Measuring Depression. Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A Brief Screening Tool for Mild Cognitive Impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Sviridova, A.; Rogovskii, V.; Kudrin, V.; Pashenkov, M.; Boyko, A.; Melnikov, M. The role of 5-HT2B-receptors in fluoxetine-mediated modulation of Th17- and Th1-cells in multiple sclerosis. J. Neuroimmunol. 2021, 356, 577608. [Google Scholar] [CrossRef]
- Melnikov, M.; Sharanova, S.; Sviridova, A.; Rogovskii, V.; Murugina, N.; Nikolaeva, A.; Dagil, Y.; Murugin, V.; Ospelnikova, T.; Boyko, A.; et al. The influence of glatiramer acetate on Th17-immune response in multiple sclerosis. PLoS ONE 2020, 15, e0240305. [Google Scholar] [CrossRef]
- Jeurink, P.V.; Vissers, Y.M.; Rappard, B.; Savelkoul, H.F. T cell responses in fresh and cryopreserved peripheral blood mononuclear cells: Kinetics of cell viability, cellular subsets, proliferation, and cytokine production. Cryobiology 2008, 57, 91–103. [Google Scholar] [CrossRef]
- Case, A.J.; Roessner, C.T.; Tian, J.; Zimmerman, M.C. Mitochondrial Superoxide Signaling Contributes to Norepinephrine-Mediated T-Lymphocyte Cytokine Profiles. PLoS ONE 2016, 11, e0164609. [Google Scholar] [CrossRef]
- Hashimoto, T.; Kitamura, N.; Kobayashi, N.; Suko, M.; Kaminuma, O.; Mori, A. Effect of Formoterol on Allergen-Induced Cytokine Synthesis by Atopic Asthmatics. Int. Arch. Allergy Immunol. 2007, 143 (Suppl. 1), 106–109. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Factor | Stimulation | MS Patients, n = 25 | Healthy Subjects, n = 16 |
|---|---|---|---|
| IL-17, pg/mL | None | 0 (0; 3) | 4 (2; 8) |
| Anti-CD3/CD28 | 503 (237; 1098) | 632 (251; 874) | |
| IFN-γ, pg/mL | None | 7 (3; 24) | 2 (0; 26) |
| Anti-CD3/CD28 | 5598 (3987; 7491) | 4991 (2803; 5846) |
| Factor | Stimulation | MS Patients, n = 15 | Healthy Subjects, n = 15 |
|---|---|---|---|
| IL-17, pg/mL | None | 0 (0; 2) | 0 (0; 1) |
| Anti-CD3/CD28 | 325 (171; 498) | 301 (184; 495) | |
| IFN-γ, pg/mL | None | 7 (4; 23) | 7 (3; 9) |
| Anti-CD3/CD28 | 2790 (2232; 5232) | 2409 (1213; 5439) |
| Factor | MS Patients, n = 25 | Healthy Subjects, n = 16 |
|---|---|---|
| Epinephrine, pmol/mL | 36.97 (21.3; 78.16) | 38.22 (29.9; 64.02) |
| Norepinephrine, pmol/mL | 1772 (1317; 2262) | 1733 (1153; 2564) |
| MHPG, pmol/mL | 262.9 (139.7; 392.7) | 214.2 (148.6; 282.5) |
| Factor | MS Patients, n = 25 | Healthy Subjects, n = 16 |
|---|---|---|
| Age, years | 31 (25; 34) | 30 (28; 33) |
| Men/women (% women) | 8/17 (68) | 6/10 (63) |
| Duration of MS, years | 2 (2; 5) | NA |
| EDSS, score | 1.5 (1.5; 2) | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnikov, M.; Rogovskii, V.; Sviridova, A.; Lopatina, A.; Pashenkov, M.; Boyko, A. The Dual Role of the β2-Adrenoreceptor in the Modulation of IL-17 and IFN-γ Production by T Cells in Multiple Sclerosis. Int. J. Mol. Sci. 2022, 23, 668. https://doi.org/10.3390/ijms23020668
Melnikov M, Rogovskii V, Sviridova A, Lopatina A, Pashenkov M, Boyko A. The Dual Role of the β2-Adrenoreceptor in the Modulation of IL-17 and IFN-γ Production by T Cells in Multiple Sclerosis. International Journal of Molecular Sciences. 2022; 23(2):668. https://doi.org/10.3390/ijms23020668
Chicago/Turabian StyleMelnikov, Mikhail, Vladimir Rogovskii, Anastasiya Sviridova, Anna Lopatina, Mikhail Pashenkov, and Alexey Boyko. 2022. "The Dual Role of the β2-Adrenoreceptor in the Modulation of IL-17 and IFN-γ Production by T Cells in Multiple Sclerosis" International Journal of Molecular Sciences 23, no. 2: 668. https://doi.org/10.3390/ijms23020668
APA StyleMelnikov, M., Rogovskii, V., Sviridova, A., Lopatina, A., Pashenkov, M., & Boyko, A. (2022). The Dual Role of the β2-Adrenoreceptor in the Modulation of IL-17 and IFN-γ Production by T Cells in Multiple Sclerosis. International Journal of Molecular Sciences, 23(2), 668. https://doi.org/10.3390/ijms23020668