
Impact of a High-Fat Diet on the Metabolomics Profile of 129S6 and C57BL6 Mouse Strains
 ,
,
Abstract
1. Introduction
2. Results
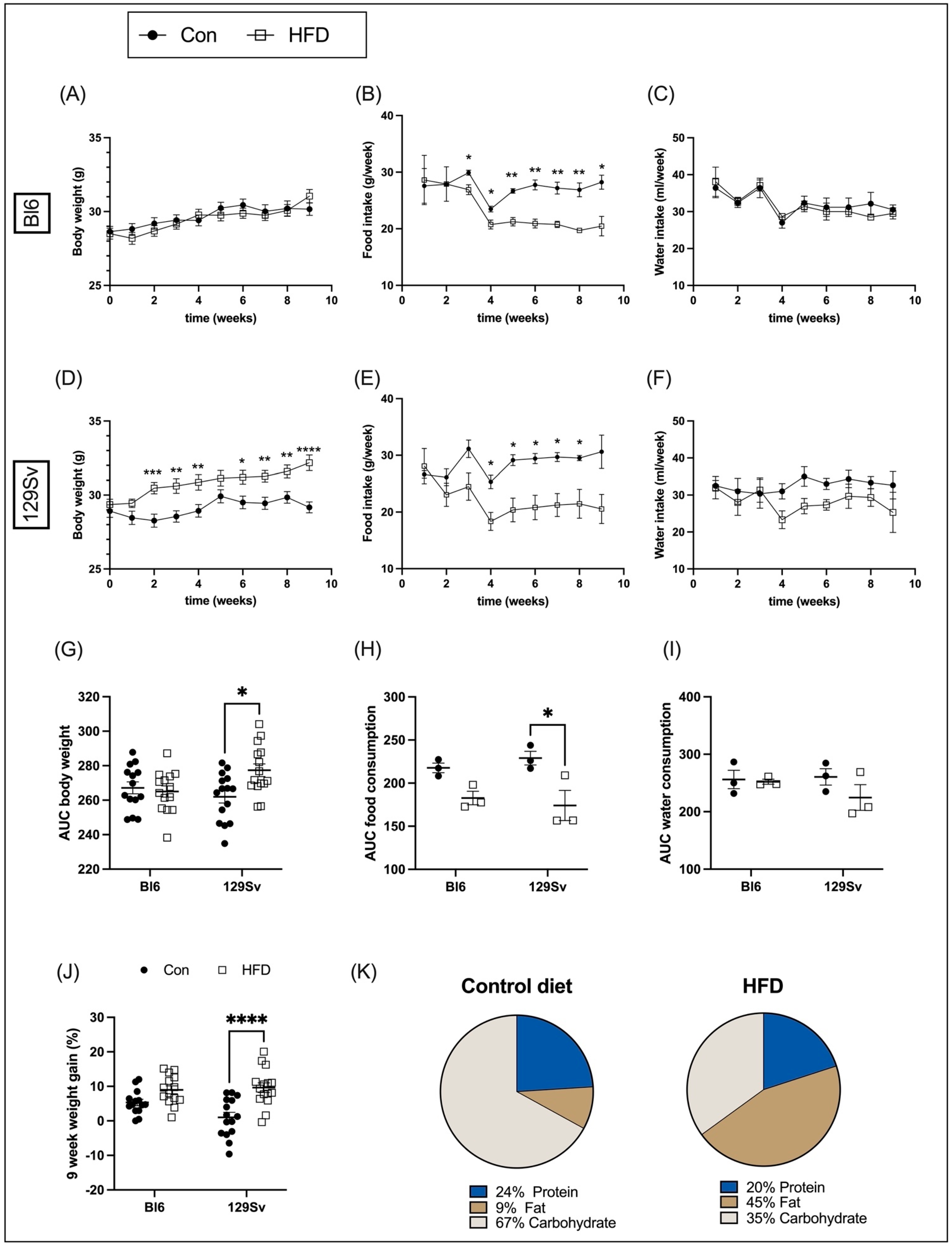
2.1. Body Weight Dynamics, Food, and Water Intake
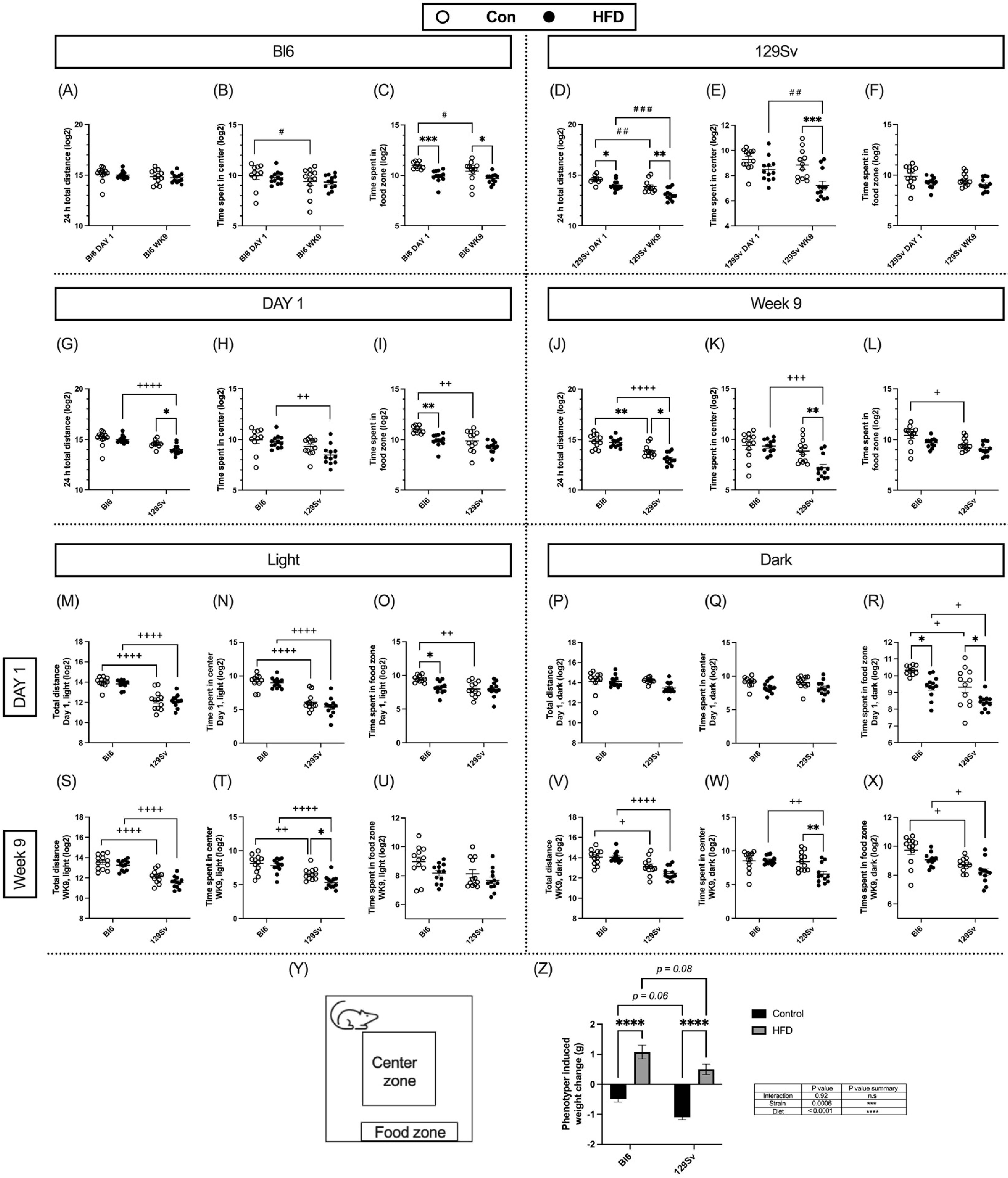
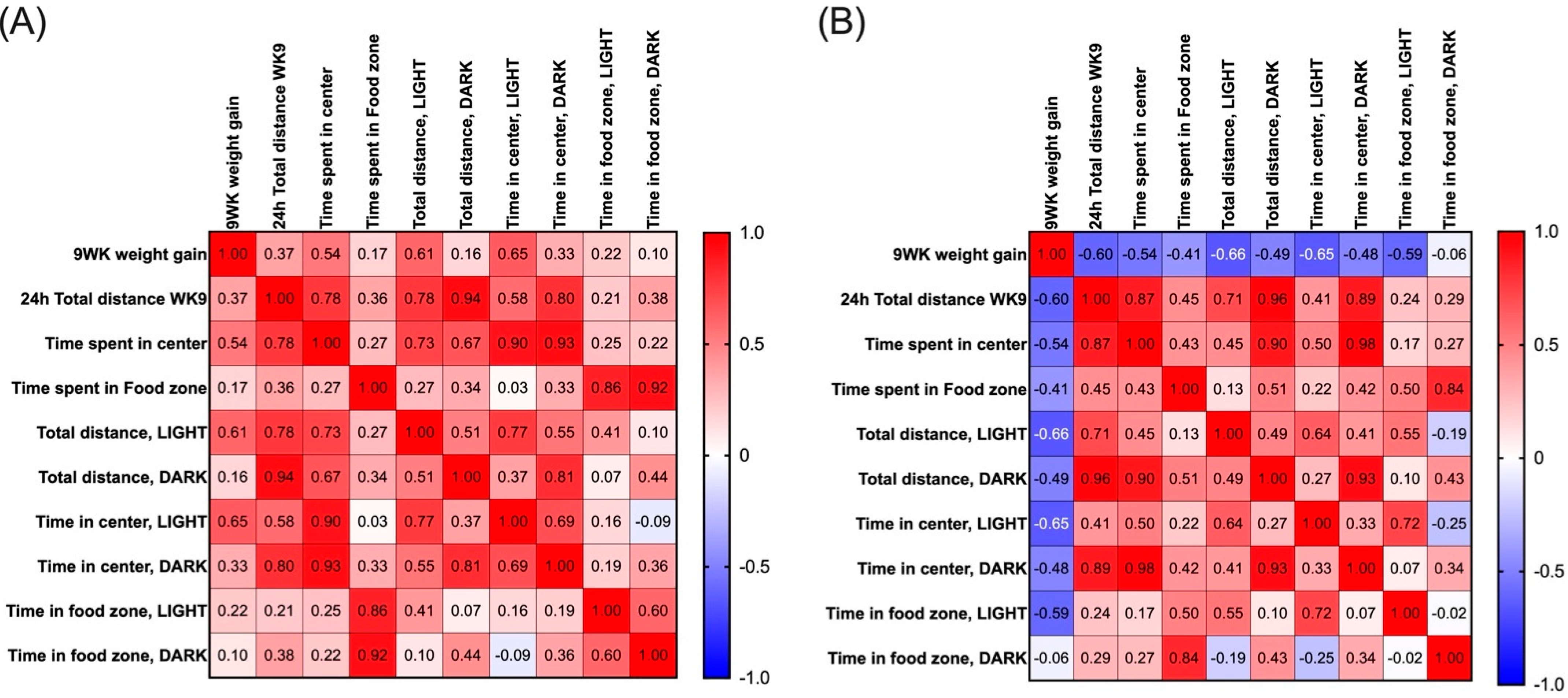
2.2. Impact of HFD on Locomotor Activity
2.3. Metabolic Changes Induced by High-Fat Diet (HFD)
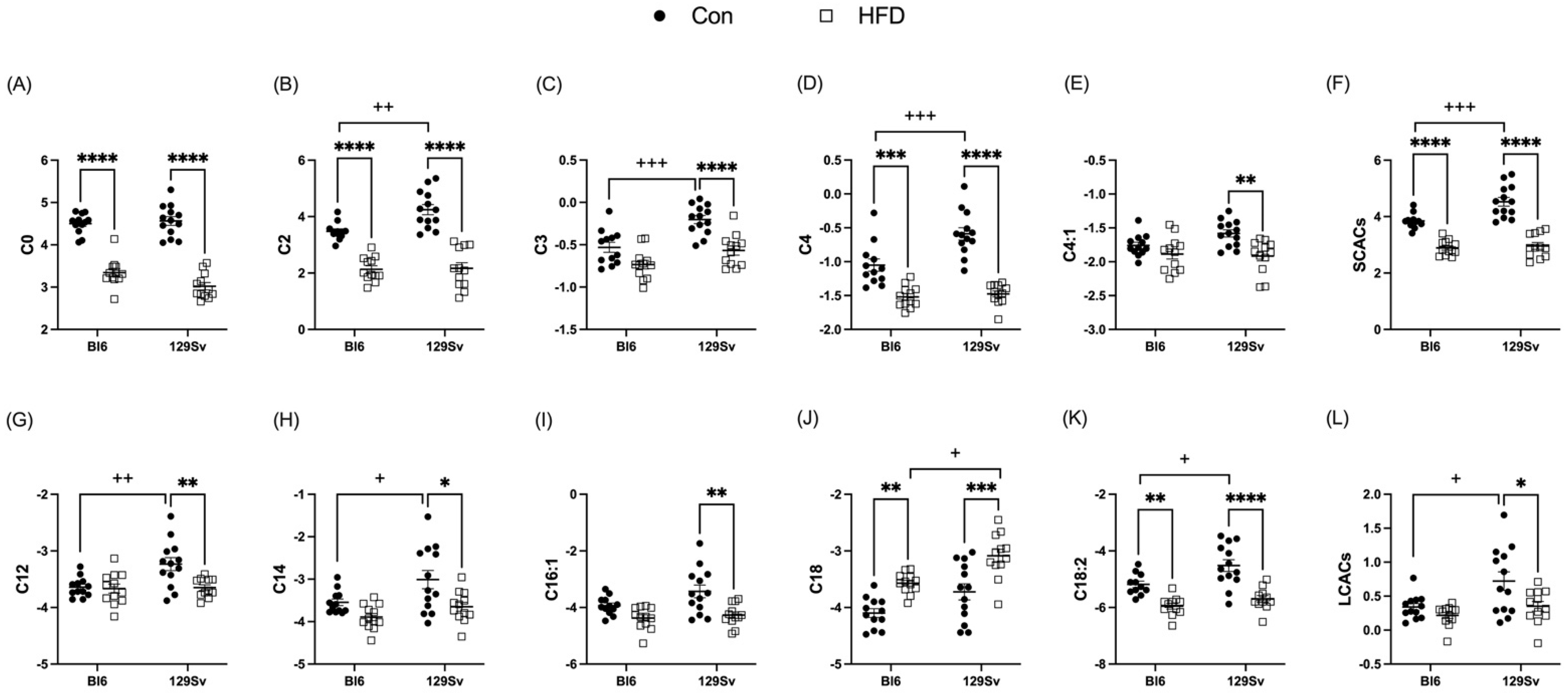
2.3.1. HFD Impact on Acylcarnitine Profile
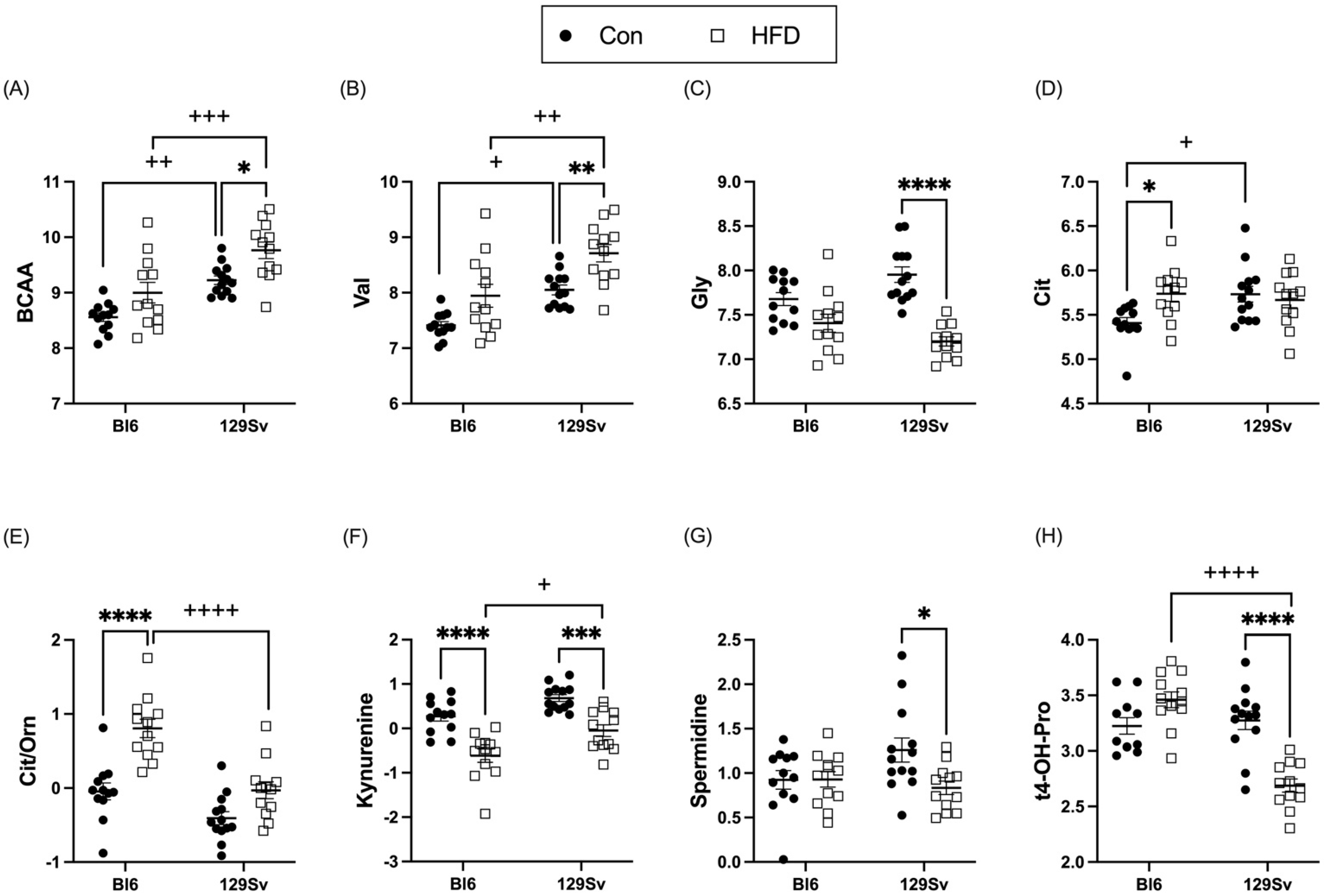
2.3.2. HFD Impact on Amino Acids and Their Derivatives Biogenic Amines
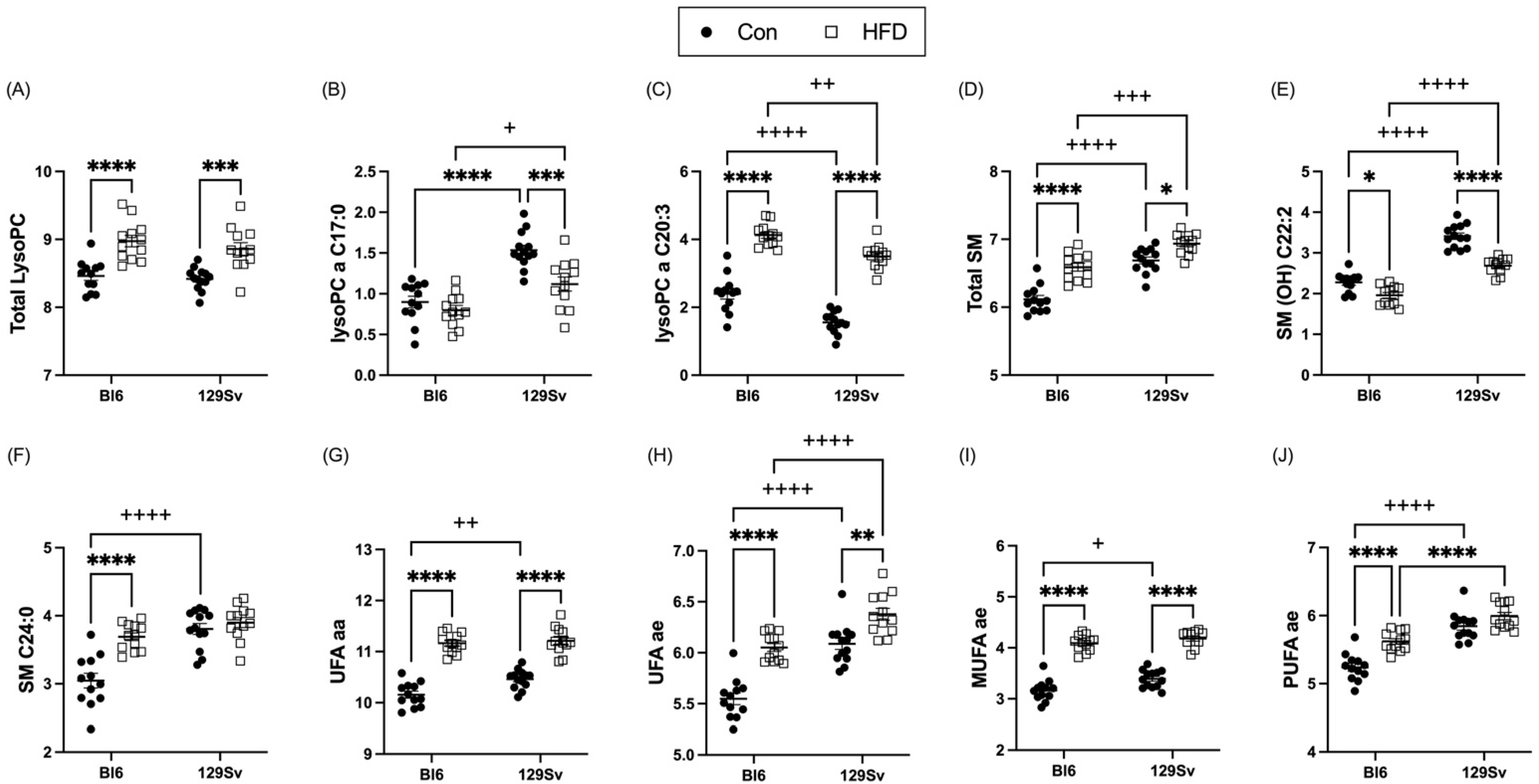
2.3.3. HFD Impact on Lipid Metabolism
2.4. Metabolite Differences Highlighted by GLM Analysis
2.4.1. Metabolic Profile Differences between Bl6 and 129Sv
2.4.2. GLM Analysis of HFD-Induced Alterations in Metabolic Profile of Bl6 Mice
2.4.3. GLM Analysis of HFD-Induced Alterations in Metabolic Profile of 129Sv Mice
3. Discussion
3.1. HFD Causes Weight Gain in 129Sv Mice, but Not in Bl6 Mice
3.2. HFD Reduces Locomotor Activity in 129Sv Mice, While Bl6 Mice Visit Less HFD Food Zone
3.3. Metabolic Profile Differences between Bl6 and 129Sv
3.4. HFD Induces a Greater Number of Alterations in Acylcarnitine Profile of 129Sv Mice
3.5. HFD Induces More Changes in Amino Acids and Their Derivatives Biogenic Amines in 129Sv Mice
3.6. HFD Induces More Changes in Lipid Profile of Bl6 Mice
4. Materials and Methods
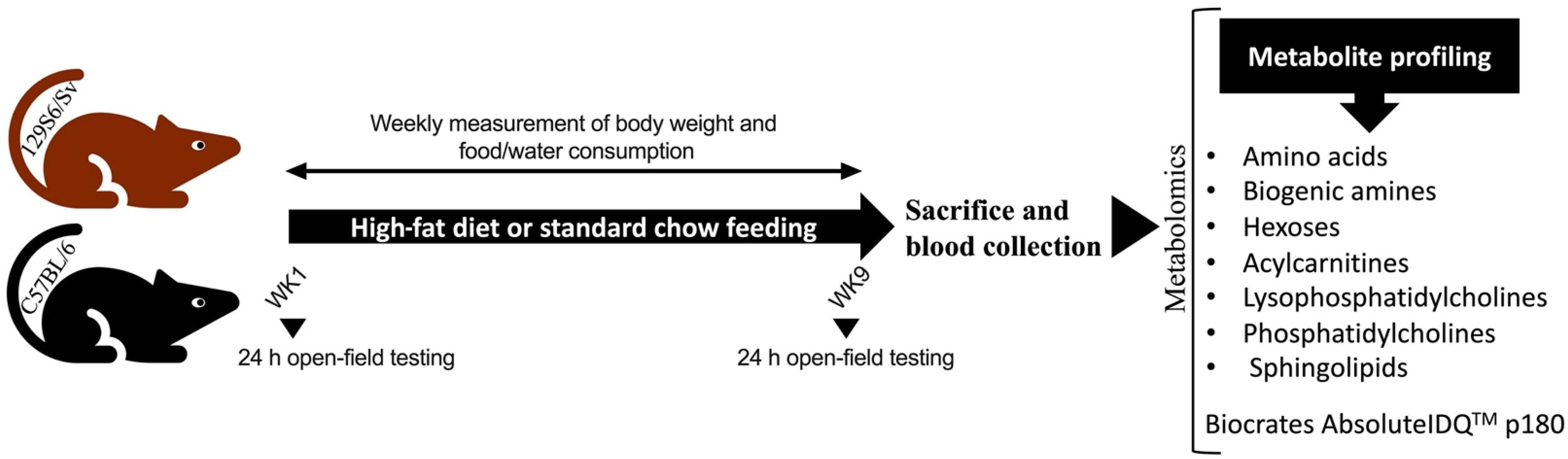
4.1. Mouse Strains and Grouping
4.2. Composition of Diets
4.3. Measurement of Body Weight and Food and Water Intake
4.4. Open-field test
4.5. Sample Collection
4.6. Measurement of Metabolites
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Yoshiki, A.; Moriwaki, K. Mouse Phenome Research: Implications of Genetic Background. ILAR J. 2006, 47, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Narvik, J.; Vanaveski, T.; Innos, J.; Philips, M.-A.; Ottas, A.; Haring, L.; Zilmer, M.; Vasar, E. Metabolic Profile Associated with Distinct Behavioral Coping Strategies of 129Sv and Bl6 Mice in Repeated Motility Test. Sci. Rep. 2018, 8, 3405. [Google Scholar] [CrossRef] [PubMed]
- Abramov, U.; Puussaar, T.; Raud, S.; Kurrikoff, K.; Vasar, E. Behavioural Differences between C57BL/6 and 129S6/SvEv Strains Are Reinforced by Environmental Enrichment. Neurosci. Lett. 2008, 443, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Heinla, I.; Leidmaa, E.; Visnapuu, T.; Philips, M.-A.; Vasar, E. Enrichment and Individual Housing Reinforce the Differences in Aggressiveness and Amphetamine Response in 129S6/SvEv and C57BL/6 Strains. Behav. Brain Res. 2014, 267, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Piirsalu, M.; Taalberg, E.; Lilleväli, K.; Tian, L.; Zilmer, M.; Vasar, E. Treatment with Lipopolysaccharide Induces Distinct Changes in Metabolite Profile and Body Weight in 129Sv and Bl6 Mouse Strains. Front. Pharmacol. 2020, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Leandro, J.; Violante, S.; Argmann, C.A.; Hagen, J.; Dodatko, T.; Bender, A.; Zhang, W.; Williams, E.G.; Bachmann, A.M.; Auwerx, J.; et al. Mild Inborn Errors of Metabolism in Commonly Used Inbred Mouse Strains. Mol. Genet. Metab. 2019, 126, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Zhang, J.; Li, S.; Edwards, J.L. Amine Metabolomics of Hyperglycemic Endothelial Cells Using Capillary LC-MS with Isobaric Tagging. J. Proteome Res. 2011, 10, 5242–5250. [Google Scholar] [CrossRef]
- Zeitoun-Ghandour, S.; Leszczyszyn, O.I.; Blindauer, C.A.; Geier, F.M.; Bundy, J.G.; Stürzenbaum, S.R.C. Elegans Metallothioneins: Response to and Defence against ROS Toxicity. Mol. Biosyst. 2011, 7, 2397–2406. [Google Scholar] [CrossRef]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic Acid Is a Biomarker for Diabetes Risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef]
- Heyes, M.P.; Saito, K.; Crowley, J.S.; Davis, L.E.; Demitrack, M.A.; Der, M.; Dilling, L.A.; Elia, J.; Kruesi, M.J.P.; Lackner, A.; et al. Quinolinic Acid and Kynurenine Pathway Metabolism In Inflammatory And Non-Inflammatory Neurological Disease. Brain 1992, 115, 1249–1273. [Google Scholar] [CrossRef]
- Wu, H.Q.; Ungerstedt, U.; Schwarcz, R. L-α-Aminoadipic Acid as a Regulator of Kynurenic Acid Production in the Hippocampus: A Microdialysis Study in Freely Moving Rats. Eur. J. Pharmacol. 1995, 281, 55–61. [Google Scholar] [CrossRef]
- Tuboly, G.; Tar, L.; Bohar, Z.; Safrany-Fark, A.; Petrovszki, Z.; Kekesi, G.; Vecsei, L.; Pardutz, A.; Horvath, G. The Inimitable Kynurenic Acid: The Roles of Different Ionotropic Receptors in the Action of Kynurenic Acid at a Spinal Level. Brain Res. Bull. 2015, 112, 52–60. [Google Scholar] [CrossRef]
- Wu, Y.; Williams, E.G.; Dubuis, S.; Mottis, A.; Jovaisaite, V.; Houten, S.M.; Argmann, C.A.; Faridi, P.; Wolski, W.; Kutalik, Z.; et al. Multilayered Genetic and Omics Dissection of Mitochondrial Activity in a Mouse Reference Population. Cell 2014, 158, 1415–1430. [Google Scholar] [CrossRef] [PubMed]
- Klebanov, G.I.; YuO, T.; Babenkova, I.V.; Lyubitsky, O.B.; OYu, R.; Boldyrev, A.A.; YuA, V. Effect of Carnosine and Its Components on Free-Radical Reactions. Membr. Cell Biol. 1998, 12, 89–99. [Google Scholar] [PubMed]
- Bae, O.-N.; Majid, A. Role of Histidine/Histamine in Carnosine-Induced Neuroprotection during Ischemic Brain Damage. Brain Res. 2013, 1527, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.T.; Jeong, E.A.; Shin, H.J.; Lee, Y.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Roh, G.S. Resveratrol Attenuates Obesity-Associated Peripheral and Central Inflammation and Improves Memory Deficit in Mice Fed a High-Fat Diet. Diabetes 2012, 61, 1444–1454. [Google Scholar] [CrossRef]
- Miller, A.A.; Spencer, S.J. Obesity and Neuroinflammation: A Pathway to Cognitive Impairment. Brain Behav. Immun. 2014, 42, 10–21. [Google Scholar] [CrossRef]
- Boitard, C.; Cavaroc, A.; Sauvant, J.; Aubert, A.; Castanon, N.; Layé, S.; Ferreira, G. Impairment of Hippocampal-Dependent Memory Induced by Juvenile High-Fat Diet Intake Is Associated with Enhanced Hippocampal Inflammation in Rats. Brain Behav. Immun. 2014, 40, 9–17. [Google Scholar] [CrossRef]
- Kaare, M.; Mikheim, K.; Lilleväli, K.; Kilk, K.; Jagomäe, T.; Leidmaa, E.; Piirsalu, M.; Porosk, R.; Singh, K.; Reimets, R.; et al. High-Fat Diet Induces Pre-Diabetes and Distinct Sex-Specific Metabolic Alterations in Negr1-Deficient Mice. Biomedicines 2021, 9, 1148. [Google Scholar] [CrossRef]
- Võikar, V.; Kõks, S.; Vasar, E.; Rauvala, H. Strain and Gender Differences in the Behavior of Mouse Lines Commonly Used in Transgenic Studies. Physiol. Behav. 2001, 72, 271–281. [Google Scholar] [CrossRef]
- Koeberl, D.D.; Young, S.P.; Gregersen, N.; Vockley, J.; Smith, W.E.; Benjamin, D.K.; An, Y.; Weavil, S.D.; Chaing, S.H.; Bali, D.; et al. Rare Disorders of Metabolism with Elevated Butyryl- and Isobutyryl-Carnitine Detected by Tandem Mass Spectrometry Newborn Screening. Pediatr. Res. 2003, 54, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and Pathophysiology of Carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.; Biasetti, M.; Messina, S.; Dominy, J. The Cytoprotective Role of Taurine in Exercise-Induced Muscle Injury. Amino Acids 2002, 22, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Rajanikant, G.; Zemke, D.; Senut, M.-C.; Frenkel, M.B.; Chen, A.F.; Gupta, R.; Majid, A. Carnosine Is Neuroprotective Against Permanent Focal Cerebral Ischemia in Mice. Stroke 2007, 38, 3023–3031. [Google Scholar] [CrossRef]
- Guiotto, A.; Calderan, A.; Ruzza, P.; Borin, G. Carnosine and Carnosine-Related Antioxidants: A Review. Curr. Med. Chem. 2005, 12, 2293–2315. [Google Scholar] [CrossRef]
- Mong, M.; Chao, C.; Yin, M. Histidine and Carnosine Alleviated Hepatic Steatosis in Mice Consumed High Saturated Fat Diet. Eur. J. Pharmacol. 2011, 653, 82–88. [Google Scholar] [CrossRef]
- Xu, W.-Y.; Shen, Y.; Zhu, H.; Gao, J.; Zhang, C.; Tang, L.; Lu, S.-Y.; Shen, C.-L.; Zhang, H.-X.; Li, Z.; et al. 2-Aminoadipic Acid Protects against Obesity and Diabetes. J. Endocrinol. 2019, 243, 111–123. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature That Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef]
- Bonen, A.; Chabowski, A.; Luiken, J.J.F.P.; Glatz, J.F.C. Is Membrane Transport of FFA Mediated by Lipid, Protein, or Both? Mechanisms and Regulation of Protein-Mediated Cellular Fatty Acid Uptake: Molecular, Biochemical, and Physiological Evidence. Physiology 2007, 22, 15–29. [Google Scholar] [CrossRef]
- Wanders, R.J.A.; Komen, J.; Kemp, S. Fatty Acid Omega-Oxidation as a Rescue Pathway for Fatty Acid Oxidation Disorders in Humans. FEBS J. 2011, 278, 182–194. [Google Scholar] [CrossRef]
- Liu, M.; Huang, Y.; Zhang, H.; Aitken, D.; Nevitt, M.C.; Rockel, J.S.; Pelletier, J.-P.; Lewis, C.E.; Torner, J.; Rampersaud, Y.R.; et al. Restricting Branched-Chain Amino Acids within a High-Fat Diet Prevents Obesity. Metabolites 2022, 12, 334. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferré, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martínez-González, M.A.; Salas-Salvadó, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-Analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Takashina, C.; Tsujino, I.; Watanabe, T.; Sakaue, S.; Ikeda, D.; Yamada, A.; Sato, T.; Ohira, H.; Otsuka, Y.; Oyama-Manabe, N.; et al. Associations among the Plasma Amino Acid Profile, Obesity, and Glucose Metabolism in Japanese Adults with Normal Glucose Tolerance. Nutr. Metab. (Lond.) 2016, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Favennec, M.; Hennart, B.; Caiazzo, R.; Leloire, A.; Yengo, L.; Verbanck, M.; Arredouani, A.; Marre, M.; Pigeyre, M.; Bessede, A.; et al. The Kynurenine Pathway Is Activated in Human Obesity and Shifted toward Kynurenine Monooxygenase Activation. Obes. (Silver Spring) 2015, 23, 2066–2074. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kobayashi, T.; Kamata, K. Role of Lysophosphatidylcholine (LPC) in Atherosclerosis. Curr. Med. Chem. 2007, 14, 3209–3220. [Google Scholar] [CrossRef]
- Miao, H.; Chen, H.; Pei, S.; Bai, X.; Vaziri, N.D.; Zhao, Y.-Y. Plasma Lipidomics Reveal Profound Perturbation of Glycerophospholipids, Fatty Acids, and Sphingolipids in Diet-Induced Hyperlipidemia. Chem. Biol. Interact. 2015, 228, 79–87. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Diet | HFD | ||||||
|---|---|---|---|---|---|---|---|
| ß (95% CI) | t-Value | p-Value | ß (95% Cl) | t-Value | p-Value | ||
| Total distance; light | 0.76 (0.46, 1.05) | 5.29 | 0.00003 | Total distance | 0.81 (0.48, 1.13) | 5.26 | 0.0001 |
| Ile | −0.77 (−1.06, −0.49) | −5.60 | 0.00002 | Total distance; light | 0.83 (0.52, 1.14) | 5.67 | 0.00005 |
| Leu | −0.76 (−1.06, −0.47) | −5.37 | 0.00003 | Center time | 0.72 (0.34, 1.10) | 4.06 | 0.001 |
| Val | −0.75 (−1.05, −0.44) | −5.14 | 0.00004 | Center time; light | 0.83 (0.52, 1.14) | 5.78 | 0.00004 |
| Carnosine | 0.87 (0.64, 1.09) | 7.92 | <0.00001 | alpha-AAA | 0.85 (0.56, 1.14) | 6.24 | 0.00002 |
| lysoPC a C16:1 | 0.81 (0.55, 1.08) | 6.42 | <0.00001 | Carnosine | 0.81 (0.49, 1.13) | 5.52 | 0.00007 |
| lysoPC a C17:0 | −0.83 (−1.08, −0.57) | −6.72 | <0.00001 | Putrescine | 0.71 (0.32, 1.10) | 3.88 | 0.001 |
| lysoPC a C24:0 | −0.77 (−1.06, −0.48) | −5.58 | 0.00002 | t4-OH-Pro | 0.79 (0.46, 1.13) | 5.07 | 0.0001 |
| PC aa C30:2 | −0.77 (−1.06, −0.48) | −5.54 | 0.00002 | lysoPC a C16:1 | 0.71 (0.32, 1.10) | 3.89 | 0.001 |
| PC aa C36:0 | −0.73 (−1.04, −0.42) | −4.94 | 0.00007 | lysoPC a C20:3 | 0.71 (0.32, 1.10) | 3.86 | 0.002 |
| PC aa C36:2 | −0.85 (−1.09, −0.60) | −7.26 | <0.00001 | PC aa C32:1 | 0.70 (0.31, 1.10) | 3.85 | 0.002 |
| PC aa C40:2 | −0.74 (−1.04, −0.43) | −4.98 | 0.00006 | PC aa C34:3 | 0.81 (0.49, 1.13) | 5.40 | 0.00007 |
| PC aa C40:5 | −0.77 (−1.06, −0.48) | −5.47 | 0.00002 | PC ae C32:1 | −0.77 (−1.12, −0.41) | −4.61 | 0.0003 |
| PC aa C40:6 | −0.74 (−1.05, −0.44) | −5.05 | 0.00005 | PC ae C34:2 | −0.84 (−1.14, −0.54) | −6.04 | 0.00002 |
| PC ae C30:2 | −0.85 (−1.09, −0.61) | −7.42 | <0.00001 | PC ae C36:2 | −0.85 (−1.14, −0.56) | −6.24 | 0.00002 |
| PC ae C32:1 | −0.80 (−1.07, −0.53) | −6.09 | <0.00001 | PC ae C36:5 | −0.75 (−1.11, −0.38) | −4.33 | 0.0006 |
| PC ae C34:0 | −0.81 (−1.08, −0.55) | −6.35 | <0.00001 | PC ae C38:2 | −0.80 (−1.13, −0.48) | −5.25 | 0.0001 |
| PC ae C34:2 | −0.74 (−1.04, −0.43) | −5.03 | 0.00006 | PC ae C38:6 | −0.85 (−1.14, −0.56) | −6.31 | 0.00001 |
| PC ae C34:3 | −0.88 (−1.10, −0.66) | −8.50 | <0.00001 | PC ae C40:6 | −0.82 (−1.13, −0.50) | −5.50 | 0.00006 |
| PC ae C36:1 | −0.80 (−1.07, −0.53) | −6.10 | <0.00001 | SM (OH) C14:1 | −0.87 (−1.14, −0.59) | −6.76 | <0.00001 |
| PC ae C36:2 | −0.92 (−1.10, −0.74) | −10.61 | <0.00001 | SM (OH) C16:1 | −0.85 (−1.14, −0.57) | −6.32 | 0.00001 |
| PC ae C38:2 | −0.92 (−1.10, −0.75) | −11.09 | <0.00001 | SM (OH) C22:1 | −0.81 (−1.13, −0.49) | −5.37 | 0.00008 |
| PC ae C38:3 | −0.83 (−1.08, −0.58) | −6.85 | <0.00001 | SM (OH) C22:2 | −0.85 (−1.14, −0.55) | −6.12 | 0.00002 |
| PC ae C38:4 | −0.87 (−1.09, −0.65) | −8.07 | <0.00001 | SM C16:0 | −0.74 (−1.11, −0.38) | −4.31 | 0.0006 |
| PC ae C38:6 | −0.87 (−1.09, −0.65) | −8.14 | <0.00001 | SM C16:1 | −0.83 (−1.14, −0.52) | −5.72 | 0.00004 |
| PC ae C40:2 | −0.84 (−1.09, −0.60) | −7.20 | <0.00001 | SM C20:2 | 0.81 (0.49, 1.13) | 5.42 | 0.00007 |
| PC ae C40:4 | −0.78 (−1.07, −0.50) | −5.77 | 0.00001 | ||||
| PC ae C40:5 | −0.82 (−1.08, −0.56) | −6.63 | <0.00001 | ||||
| PC ae C40:6 | −0.87 (−1.09, −0.65) | −8.16 | <0.00001 | ||||
| PC ae C42:5 | −0.82 (−1.08, −0.56) | −6.56 | <0.00001 | ||||
| SM (OH) C14:1 | −0.90 (−1.10, −0.70) | −9.26 | <0.00001 | ||||
| SM (OH) C16:1 | −0.85 (−1.09, −0.60) | −7.25 | <0.00001 | ||||
| SM (OH) C22:1 | −0.92 (−1.10, −0.74) | −10.55 | <0.00001 | ||||
| SM (OH) C22:2 | −0.92 (−1.10, −0.75) | −11.04 | <0.00001 | ||||
| SM C16:0 | −0.86 (−1.09, −0.63) | −7.80 | <0.00001 | ||||
| SM C16:1 | −0.90 (−1.10, −0.69) | −9.23 | <0.00001 | ||||
| SM C24:0 | −0.81 (−1.07, −0.55) | −6.40 | <0.00001 | ||||
| SM C24:1 | −0.85 (−1.09, −0.62) | −7.52 | <0.00001 | ||||
| Bl6 | 129Sv | ||||||
|---|---|---|---|---|---|---|---|
| ß (ß 95% CI) | t-Value | p-Value | ß (ß 95% CI) | t-Value | p-Value | ||
| C0 | 0.91 (0.72, 1.10) | 9.90 | <0.00001 | 9-week weight gain (%) | −0.60 (−1.02, −0.18) | −3.01 | 0.008 |
| C2 | 0.89 (0.69, 1.10) | 8.94 | <0.00001 | C0 | 0.94 (0.76, 1.12) | 11.14 | <0.00001 |
| C4- | 0.70 (0.36, 1.03) | 4.36 | 0.0003 | C2 | 0.85 (0.57, 1.13) | 6.45 | <0.00001 |
| C18 | −0.77 (−1.07, −0.47) | −5.34 | 0.00003 | C3 | 0.68 (0.30, 1.07) | 3.74 | 0.002 |
| C18:2 | 0.75 (0.43, 1.06) | 5.00 | 0.00007 | C4- | 0.89 (0.65, 1.13) | 7.90 | <0.00001 |
| Kynurenine | 0.73 (0.41, 1.05) | 4.73 | 0.0001 | C14:2 | 0.77 (0.43, 1.11) | 4.77 | 0.0002 |
| lysoPC a C18:0 | −0.78 (−1.07, −0.48) | −5.52 | 0.00002 | C18:2 | 0.78 (0.45, 1.11) | 5.04 | 0.0001 |
| lysoPC a C18:1 | −0.88 (−1.10, −0.66) | −8.40 | <0.00001 | Gly | 0.83 (0.53, 1.12) | 5.85 | 0.00002 |
| lysoPC a C20:3 | −0.89 (−1.10, −0.67) | −8.58 | <0.00001 | Kynurenine | 0.64 (0.23, 1.05) | 3.34 | 0.004 |
| lysoPC a C26:0 | −0.70 (−1.03, −0.36) | −4.34 | 0.0003 | t4-OH-Pro | 0.76 (0.41, 1.10) | 4.63 | 0.0003 |
| PC aa C28:1 | −0.83 (−1.09, −0.57) | −6.71 | <0.00001 | lysoPC a C18:0 | −0.75 (−1.10, −0.40) | −4.58 | 0.0003 |
| PC aa C30:2 | −0.88 (−1.10, −0.66) | −8.43 | <0.00001 | lysoPC a C18:1 | −0.88 (−1.13, −0.63) | −7.49 | <0.00001 |
| PC aa C32:1 | −0.81 (−1.08, −0.53) | −6.14 | <0.00001 | lysoPC a C20:3 | −0.94 (−1.12, −0.75) | −10.60 | <0.00001 |
| PC aa C34:1 | −0.96 (−1.09, −0.82) | −14.97 | <0.00001 | lysoPC a C26:0 | −0.58 (−1.01, −0.15) | −2.88 | 0.01 |
| PC aa C34:2 | −0.76 (−1.07, −0.46) | −5.30 | 0.00003 | lysoPC a C28:1 | −0.82 (−1.12, −0.53) | −5.84 | 0.00003 |
| PC aa C36:1 | −0.97 (−1.08, −0.86) | −17.93 | <0.00001 | PC aa C28:1 | −0.89 (−1.13, −0.65) | −7.87 | <0.00001 |
| PC aa C36:2 | −0.95 (−1.09, −0.82) | −14.31 | <0.00001 | PC aa C30:2 | −0.93 (−1.13, −0.73) | −9.97 | <0.00001 |
| PC aa C36:3 | −0.94 (−1.10, −0.78) | −12.20 | <0.00001 | PC aa C32:3 | −0.80 (−1.12, −0.48) | −5.33 | 0.00007 |
| PC aa C38:0 | −0.71 (−1.04, −0.38) | −4.49 | 0.0002 | PC aa C34:1 | −0.86 (−1.13, −0.60) | −6.85 | <0.00001 |
| PC aa C38:3 | −0.92 (−1.10, −0.75) | −10.87 | <0.00001 | PC aa C36:1 | −0.96 (−1.11, −0.80) | −13.06 | <0.00001 |
| PC aa C38:4 | −0.68 (−1.02, −0.34) | −4.19 | 0.0005 | PC aa C36:2 | −0.94 (−1.12, −0.77) | −11.50 | <0.00001 |
| PC aa C38:5 | −0.67 (−1.01, −0.32) | −4.00 | 0.0007 | PC aa C36:3 | −0.92 (−1.13, −0.71) | −9.43 | <0.00001 |
| PC aa C40:3 | −0.69 (−1.03, −0.36) | −4.30 | 0.0004 | PC aa C38:1 | −0.77 (−1.11, −0.44) | −4.87 | 0.0002 |
| PC ae C30:2 | −0.79 (−1.08, −0.51) | −5.81 | 0.00001 | PC aa C38:3 | −0.92 (−1.13, −0.72) | −9.73 | <0.00001 |
| PC ae C32:1 | −0.76 (−1.06, −0.45) | −5.17 | 0.00005 | PC aa C38:5 | −0.77 (−1.11, −0.44) | −4.87 | 0.0002 |
| PC ae C32:2 | −0.91 (−1.10, −0.72) | −10.08 | <0.00001 | PC aa C40:3 | −0.62 (−1.03, −0.20) | −3.12 | 0.007 |
| PC ae C34:1 | −0.95 (−1.10, −0.80) | −13.15 | <0.00001 | PC aa C40:5 | −0.81 (−1.12, −0.50) | −5.55 | 0.00004 |
| PC ae C34:3 | −0.80 (−1.08, −0.52) | −5.98 | <0.00001 | PC ae C30:2 | 0.84 (0.56, 1.13) | 6.24 | 0.00001 |
| PC ae C36:0 | −0.84 (−1.09, −0.59) | −6.97 | <0.00001 | PC ae C32:1 | −0.72 (−1.09, −0.35) | −4.10 | 0.0008 |
| PC ae C36:1 | −0.96 (−1.09, −0.84) | −16.05 | <0.00001 | PC ae C32:2 | −0.87 (−1.13, −0.62) | −7.18 | <0.00001 |
| PC ae C36:2 | −0.73 (−1.05, −0.41) | −4.79 | 0.0001 | PC ae C34:1 | −0.87 (−1.13, −0.60) | −6.92 | <0.00001 |
| PC ae C36:3 | −0.89 (−1.10, −0.68) | −8.85 | <0.00001 | PC ae C34:3 | −0.73 (−1.09, −0.37) | −4.30 | 0.0006 |
| PC ae C36:4 | −0.71 (−1.04, −0.38) | −4.48 | 0.0002 | PC ae C36:0 | −0.74 (−1.10, −0.39) | −4.42 | 0.0004 |
| PC ae C38:1 | −0.83 (−1.09, −0.57) | −6.58 | <0.00001 | PC ae C36:1 | −0.86 (−1.13, −0.58) | −6.65 | <0.00001 |
| PC ae C38:3 | −0.85 (−1.10, −0.61) | −7.31 | <0.00001 | PC ae C36:3 | −0.75 (−1.10, −0.40) | −4.58 | 0.0003 |
| PC ae C38:4 | −0.67 (−1.01, −0.32) | −4.00 | 0.0007 | PC ae C36:5 | −0.74 (−1.10, −0.38) | −4.39 | 0.0005 |
| PC ae C38:5 | −0.91 (−1.10, −0.71) | −9.74 | <0.00001 | PC ae C38:1 | −0.79 (−1.11, −0.46) | −5.15 | 0.0001 |
| PC ae C40:5 | −0.71 (−1.04, −0.38) | −4.50 | 0.0002 | PC ae C38:2 | 0.91 (0.69, 1.13) | 8.81 | <0.00001 |
| PC ae C42:2 | −0.81 (−1.08, −0.54) | −6.16 | <0.00001 | PC ae C38:3 | −0.83 (−1.13, −0.53) | −5.95 | 0.00002 |
| SM (OH) C14:1 | −0.83 (−1.09, −0.57) | −6.72 | <0.00001 | PC ae C38:5 | −0.88 (−1.13, −0.63) | −7.40 | <0.00001 |
| SM (OH) C16:1 | −0.71 (−1.04, −0.39) | −4.54 | 0.0002 | PC ae C40:4 | 0.88 (0.62, 1.13) | 7.26 | <0.00001 |
| SM C16:0 | −0,92 (−1.10, −0.74) | −10.49 | <0.00001 | PC ae C42:0 | 0.77 (0.43, 1.11) | 4.84 | 0.0002 |
| SM C16:1 | −0.80 (−1.08, −0.52) | −6.00 | <0.00001 | PC ae C42:2 | −0.76 (−1.10, −0.42) | −4.68 | 0.0003 |
| SM C18:0 | −0,87 (−1.10, −0.64) | −7.81 | <0.00001 | SM (OH) C14:1 | −0.90 (−1.13, −0.67) | −8.24 | <0.00001 |
| SM C18:1 | −0.70 (−1.04, −0.37) | −4.44 | 0.0002 | SM (OH) C16:1 | −0.88 (−1.13, −0.63) | −7.52 | <0.00001 |
| SM C22:3 | −0.66 (−1.01, −0.31) | −3.95 | 0.0008 | SM (OH) C22:2 | 0.89 (0.65, 1.13) | 7.89 | <0.00001 |
| SM C24:0 | −0.75 (−1.06, −0.44) | −5.06 | 0.00006 | SM C16:0 | −0.94 (−1.12, −0.76) | −11.14 | <0.00001 |
| SM C16:1 | −0.76 (−1.10, −0.41) | −4.65 | 0.0003 | ||||
| SM C18:1 | −0.78 (−1.11, −0.44) | −4.94 | 0.0001 | ||||
| Control Diet (3.230 kcal/kg) | High-Fat Diet (4.615 kcal/kg) | |
|---|---|---|
| Fat (%) | 9 kJ | 45 kJ |
| Protein (%) | 24 kJ | 20 kJ |
| Carbohydrate (%) | 67 kJ | 35 kJ |
| Crude Nutrients [%] | ||
| Crude protein (N × 6.25) | 19.0 | 22.0 |
| Crude fat | 3.3 | 23.6 |
| Crude fiber | 4.9 | 5.7 |
| Crude ash | 6.4 | 5.3 |
| Starch | 35.2 | 6.8 |
| Sugar | 5.3 | 21.1 |
| N free extracts | 54.2 | 40.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piirsalu, M.; Taalberg, E.; Jayaram, M.; Lilleväli, K.; Zilmer, M.; Vasar, E. Impact of a High-Fat Diet on the Metabolomics Profile of 129S6 and C57BL6 Mouse Strains. Int. J. Mol. Sci. 2022, 23, 11682. https://doi.org/10.3390/ijms231911682
Piirsalu M, Taalberg E, Jayaram M, Lilleväli K, Zilmer M, Vasar E. Impact of a High-Fat Diet on the Metabolomics Profile of 129S6 and C57BL6 Mouse Strains. International Journal of Molecular Sciences. 2022; 23(19):11682. https://doi.org/10.3390/ijms231911682
Chicago/Turabian StylePiirsalu, Maria, Egon Taalberg, Mohan Jayaram, Kersti Lilleväli, Mihkel Zilmer, and Eero Vasar. 2022. "Impact of a High-Fat Diet on the Metabolomics Profile of 129S6 and C57BL6 Mouse Strains" International Journal of Molecular Sciences 23, no. 19: 11682. https://doi.org/10.3390/ijms231911682
APA StylePiirsalu, M., Taalberg, E., Jayaram, M., Lilleväli, K., Zilmer, M., & Vasar, E. (2022). Impact of a High-Fat Diet on the Metabolomics Profile of 129S6 and C57BL6 Mouse Strains. International Journal of Molecular Sciences, 23(19), 11682. https://doi.org/10.3390/ijms231911682