
The Hydrophobic Residues in Amino Terminal Domains of Cx46 and Cx50 Are Important for Their Gap Junction Channel Ion Permeation and Gating


Abstract
1. Introduction
2. Results
2.1. Homotypic Cx46 and Cx50 Variants Form Functional GJ Channels
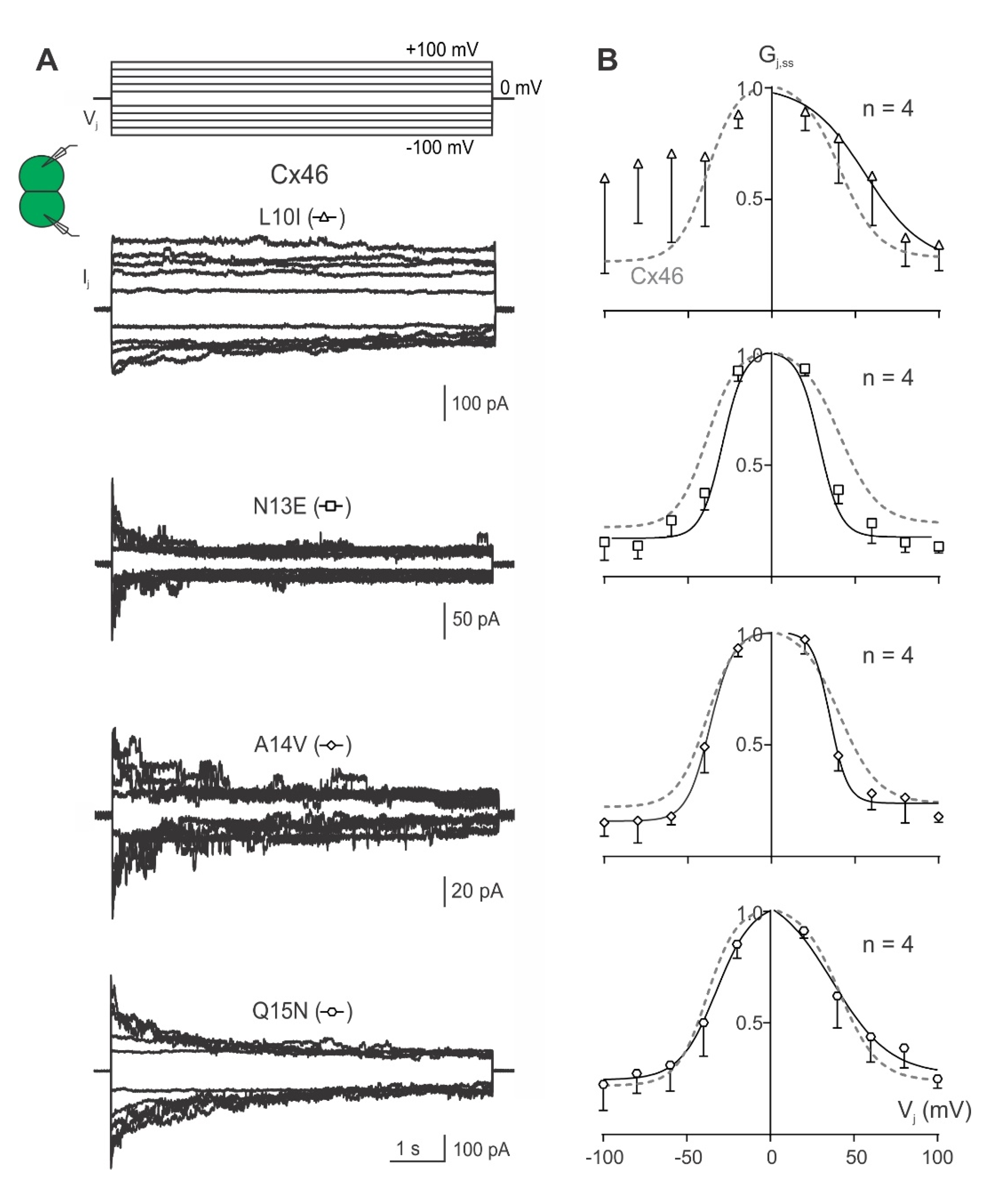
2.2. Vj-Gating Properties of the Homotypic GJs Formed by Cx46 or Cx50 NT Variants
2.3. Unitary Channel Conductance of Homotypic GJs of Cx46 and Cx50 NT Variants
2.4. Homology Modeling of Selected Cx46 and Cx50 Variants
3. Discussion
3.1. Hydrophobic Residues in the NT Domain of Cx46 and Cx50
3.2. Hydrophilic Residues in the NT Domain of Cx46 and Cx50
4. Materials and Methods
4.1. Plasmid Construction
4.2. Cell Culture and Transient Transfections
4.3. Electrophysiological Recordings
4.4. Transjunctional Voltage Dependent Gating
4.5. Single Channel Analysis
4.6. Homology Structure Modeling
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goodenough, D.A.; Goliger, J.A.; Paul, D.L. Connexins, connexons, and intercellular communication. Annu. Rev. Biochem. 1996, 65, 475–502. [Google Scholar] [CrossRef]
- Mese, G.; Richard, G.; White, T.W. Gap junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2516–2524. [Google Scholar] [CrossRef]
- Goldberg, G.S.; Valiunas, V.; Brink, P.R. Selective permeability of gap junction channels. Biochim. Biophys. Acta 2004, 1662, 96–101. [Google Scholar] [CrossRef]
- Gerido, D.A.; White, T.W. Connexin disorders of the ear, skin, and lens. Biochim. Biophys. Acta 2004, 1662, 159–170. [Google Scholar]
- Orellana, J.A.; Avendaño, B.C.; Montero, T.D. Role of connexins and pannexins in ischemic stroke. Curr. Med. Chem. 2014, 21, 2165–2182. [Google Scholar] [CrossRef]
- Sinyuk, M.; Mulkearns-Hubert, E.E.; Reizes, O.; Lathia, J. Cancer Connectors: Connexins, Gap Junctions, and Communication. Front. Oncol. 2018, 8, 646. [Google Scholar]
- Aasen, T.; Mesnil, M.; Naus, C.C.; Lampe, P.D.; Laird, D.W. Gap junctions and cancer: Communicating for 50 years. Nat. Rev. Cancer 2017, 17, 74. [Google Scholar] [CrossRef]
- Söhl, G.; Maxeiner, S.; Willecke, K. Expression and functions of neuronal gap junctions. Nat. Rev. Neurosci. 2005, 6, 191–200. [Google Scholar] [CrossRef]
- Goodenough, D.A.; Paul, D.L. Gap junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef]
- Abascal, F.; Zardoya, R. Evolutionary analyses of gap junction protein families. Biochim. Biophys. Acta 2013, 1828, 4–14. [Google Scholar] [CrossRef]
- Bukauskas, F.F.; Verselis, V.K. Gap junction channel gating. Biochim. Biophys. Acta 2004, 1662, 42–60. [Google Scholar] [CrossRef]
- González, D.; Gómez-Hernández, J.M.; Barrio, L. C, Molecular basis of voltage dependence of connexin channels: An integrative appraisal. Prog. Biophys. Mol. Biol. 2007, 94, 66–106. [Google Scholar] [CrossRef]
- Veenstra, R.D.; Wang, H.Z.; Beyer, E.C.; Ramanan, S.V.; Brink, P.R. Connexin37 forms high conductance gap junction channels with subconductance state activity and selective dye and ionic permeabilities. Biophys. J. 1994, 66, 1915–1928. [Google Scholar] [CrossRef]
- Moreno, A.P.; Chanson, M.; Elenes, S.; Anumonwo, J.; Scerri, I.; Gu, H.; Taffet, S.M.; Delmar, M. Role of the carboxyl terminal of connexin43 in transjunctional fast voltage gating. Circ. Res. 2002, 90, 450–457. [Google Scholar] [CrossRef]
- Pellegrini-Calace, M.; Maiwald, T.; Thornton, J.M. PoreWalker: A novel tool for the identification and characterization of channels in transmembrane proteins from their three-dimensional structure. PLoS Comput. Biol. 2009, 5, e1000440. [Google Scholar] [CrossRef]
- Teubner, B.; Degen, J.; Söhl, G.; Güldenagel, M.; Bukauskas, F.F.; Trexler, E.B.; Verselis, V.K.; De Zeeuw, C.I.; Lee, C.G.; Kozak, C.A.; et al. Functional expression of the murine connexin 36 gene coding for a neuron-specific gap junctional protein. J. Membr. Biol. 2000, 176, 249–262. [Google Scholar] [CrossRef]
- Purnick, P.E.; Oh, S.; Abrams, C.K.; Verselis, V.K.; Bargiello, T.A. Reversal of the gating polarity of gap junctions by negative charge substitutions in the N-terminus of connexin 32. Biophys. J. 2000, 79, 2403–2415. [Google Scholar] [CrossRef]
- Myers, J.B.; Haddad, B.G.; O’Neill, S.E.; Chorev, D.S.; Yoshioka, C.C.; Robinson, C.V.; Zuckerman, D.M.; Reichow, S.L. Structure of native lens connexin 46/50 intercellular channels by cryo-EM. Nature 2018, 564, 372–377. [Google Scholar] [CrossRef]
- Xin, L.; Bai, D. Functional roles of the amino terminal domain in determining biophysical properties of Cx50 gap junction channels. Front. Physiol. 2013, 4, 373. [Google Scholar] [CrossRef]
- Tong, J.-J.; Ebihara, L. Structural Determinants for the Differences in Voltage Gating of Chicken Cx56 and Cx45.6 Gap-Junctional Hemichannels. Biophys. J. 2006, 91, 2142–2154. [Google Scholar] [CrossRef]
- Peracchia, C.; Peracchia, L.L. Inversion of both gating polarity and CO2 sensitivity of voltage gating with D3N mutation of Cx50. Am. J. Physiol. Cell Physiol. 2005, 288, C1381–C1389. [Google Scholar] [CrossRef] [PubMed]
- Musa, H.; Fenn, E.; Crye, M.; Gemel, J.; Beyer, E.C.; Veenstra, R.D. Amino terminal glutamate residues confer spermine sensitivity and affect voltage gating and channel conductance of rat connexin40 gap junctions. J. Physiol. 2004, 557 Pt 3, 863–878. [Google Scholar] [CrossRef] [PubMed]
- Verselis, V.K.; Ginter, C.S.; Bargiello, T.A. Opposite voltage gating polarities of two closely related connexins. Nature 1994, 368, 348–351. [Google Scholar] [CrossRef]
- Xin, L.; Gong, X.Q.; Bai, D. The role of amino terminus of mouse Cx50 in determining transjunctional voltage-dependent gating and unitary conductance. Biophys. J. 2010, 99, 2077–2086. [Google Scholar] [CrossRef]
- Xin, L.; Nakagawa, S.; Tsukihara, T.; Bai, D. Aspartic Acid residue d3 critically determines cx50 gap junction channel transjunctional voltage-dependent gating and unitary conductance. Biophys. J. 2012, 102, 1022–1031. [Google Scholar] [CrossRef]
- Kronengold, J.; Srinivas, M.; Verselis, V.K. The N-terminal half of the connexin protein contains the core elements of the pore and voltage gates. J. Membr. Biol. 2012, 245, 453–463. [Google Scholar] [CrossRef]
- Srinivas, M.; Rozental, R.; Kojima, T.; Dermietzel, R.; Mehler, M.; Condorelli, D.F.; Kessler, J.A.; Spray, D.C. Functional properties of channels formed by the neuronal gap junction protein connexin36. J. Neurosci. 1999, 19, 9848–9855. [Google Scholar] [CrossRef]
- Tong, X.; Aoyama, H.; Tsukihara, T.; Bai, D. Charge at the 46th residue of connexin 50 is crucial for the gap-junctional unitary conductance and transjunctional voltage-dependent gating. J. Physiol. 2014, 592, 5187–5202. [Google Scholar] [CrossRef]
- Maeda, S.; Nakagawa, S.; Suga, M.; Yamashita, E.; Oshima, A.; Fujiyoshi, Y.; Tsukihara, T. Structure of the connexin 26 gap junction channel at 3.5 A resolution. Nature 2009, 458, 597–602. [Google Scholar] [CrossRef]
- Maeda, S.; Tsukihara, T. Structure of the gap junction channel and its implications for its biological functions. Cell. Mol. Life Sci. CMLS 2011, 68, 1115–1129. [Google Scholar]
- Kwon, T.; Harris, A.L.; Rossi, A.; Bargiello, T.A. Molecular dynamics simulations of the Cx26 hemichannel: Evaluation of structural models with Brownian dynamics. J. Gen. Physiol. 2011, 138, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Rossi, A.R.; Harris, A.L. Computational Studies of Molecular Permeation through Connexin26 Channels. Biophys. J. 2016, 110, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Villanelo, F.; Escalona, Y.; Pareja-Barrueto, C.; Garate, J.A.; Skerrett, I.M.; Perez-Acle, T. Accessing gap-junction channel structure-function relationships through molecular modeling and simulations. BMC Cell Biol. 2017, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Bennett, B.C.; Purdy, M.D.; Baker, K.A.; Acharya, C.; McIntire, W.E.; Stevens, R.C.; Zhang, Q.; Harris, A.L.; Abagyan, R.; Yeager, M. An electrostatic mechanism for Ca(2+)-mediated regulation of gap junction channels. Nat. Commun. 2016, 7, 8770. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.K.; Jagielnicki, M.; McIntire, W.E.; Purdy, M.D.; Dharmarajan, V.; Griffin, P.R.; Yeager, M. A Steric "Ball-and-Chain" Mechanism for pH-Mediated Regulation of Gap Junction Channels. Cell Rep. 2020, 31, 107482. [Google Scholar] [CrossRef]
- Flores, J.A.; Haddad, B.G.; Dolan, K.A.; Myers, J.B.; Yoshioka, C.C.; Copperman, J.; Zuckerman, D.M.; Reichow, S.L. Connexin-46/50 in a dynamic lipid environment resolved by CryoEM at 1.9 Å. Nat. Commun. 2020, 11, 4331. [Google Scholar] [CrossRef]
- Bai, D. Structural analysis of key gap junction domains—Lessons from genome data and disease-linked mutants. Semin. Cell Dev. Biol. 2016, 50, 74–82. [Google Scholar] [CrossRef]
- Yue, B.; Haddad, B.G.; Khan, U.; Chen, H.; Atalla, M.; Zhang, Z.; Zuckerman, D.M.; Reichow, S.L.; Bai, D. Connexin 46 and connexin 50 gap junction channel properties are shaped by structural and dynamic features of their N-terminal domains. J. Physiol. 2021, 599, 3313–3335. [Google Scholar] [CrossRef]
- Hansen, L.; Yao, W.; Eiberg, H.; Funding, M.; Riise, R.; Kjaer, K.W.; Hejtmancik, J.F.; Rosenberg, T. The congenital “ant-egg” cataract phenotype is caused by a missense mutation in connexin46. Mol. Vis. 2006, 12, 1033–1039. [Google Scholar]
- Tong, J.J.; Sohn, B.C.; Lam, A.; Walters, D.E.; Vertel, B.M.; Ebihara, L. Properties of two cataract-associated mutations located in the NH2 terminus of connexin 46. Am. J. Physiol. Cell Physiol. 2013, 304, C823–C832. [Google Scholar] [CrossRef][Green Version]
- Javadiyan, S.; Lucas, S.E.M.; Wangmo, D.; Ngy, M.; Edussuriya, K.; Craig, J.E.; Rudkin, A.; Casson, R.; Selva, D.; Sharma, S.; et al. Identification of novel mutations causing pediatric cataract in Bhutan, Cambodia, and Sri Lanka. Mol. Genet. Genom. Med. 2018, 6, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.S.; Bennett, T.M.; Culican, S.M.; Shiels, A. Exome sequencing identifies novel and recurrent mutations in GJA8 and CRYGD associated with inherited cataract. Hum. Genom. 2014, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liang, Y.; Zhou, Y.; Zeng, H.; Jia, S.; Shi, J. A Missense Mutation in GJA8 Encoding Connexin 50 in a Chinese Pedigree with Autosomal Dominant Congenital Cataract. Tohoku J. Exp. Med. 2018, 244, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Wang, J.; Li, T.; Chan, R.; Atalla, M.; Chen, R.C.; Khazaneh, M.T.; An, R.J.; Stathopulos, P.B. Differential Domain Distribution of gnomAD- and Disease-Linked Connexin Missense Variants. Int. J. Mol. Sci. 2021, 22, 7832. [Google Scholar] [CrossRef] [PubMed]
- Tejada, M.G.; Sudhakar, S.; Kim, N.K.; Aoyama, H.; Shilton, B.H.; Bai, D. Variants with increased negative electrostatic potential in the Cx50 gap junction pore increased unitary channel conductance and magnesium modulation. Biochem. J. 2018, 475, 3315–3330. [Google Scholar] [CrossRef]
- Tong, J.J.; Khan, U.; Haddad, B.G.; Minogue, P.J.; Beyer, E.C.; Berthoud, V.M.; Reichow, S.L.; Ebihara, L. Molecular mechanisms underlying enhanced hemichannel function of a cataract-associated Cx50 mutant. Biophys. J. 2021, 120, 5644–5656. [Google Scholar] [CrossRef]
- Noureldin, M.; Chen, H.; Bai, D. Functional Characterization of Novel Atrial Fibrillation-Linked GJA5 (Cx40) Mutants. Int. J. Mol. Sci. 2018, 19, 977. [Google Scholar] [CrossRef]
- Jassim, A.; Aoyama, H.; Ye, W.G.; Chen, H.; Bai, D. Engineered Cx40 variants increased docking and function of heterotypic Cx40/Cx43 gap junction channels. J. Mol. Cell. Cardiol. 2016, 90, 11–20. [Google Scholar] [CrossRef]
- Bai, D.; Cameron, J.A. Patch clamp analysis of gap junction channel properties. In Gap Junction Channels and Hemichannels; CRC Press: Boca Raton, FL, USA, 2016; pp. 93–114. [Google Scholar]
- Moreno, A.P.; Rook, M.B.; Fishman, G.I.; Spray, D.C. Gap junction channels: Distinct voltage-sensitive and -insensitive conductance states. Biophys. J. 1994, 67, 113–119. [Google Scholar] [CrossRef]
- Wilders, R.; Jongsma, H.J. Limitations of the dual voltage clamp method in assaying conductance and kinetics of gap junction channels. Biophys. J. 1992, 63, 942–953. [Google Scholar] [CrossRef]
- Bukauskas, F.F.; Weingart, R. Voltage-dependent gating of single gap junction channels in an insect cell line. Biophys. J. 1994, 67, 613–625. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Protein Structure Modeling with MODELLER. Methods Mol. Biol. 2021, 2199, 239–255. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GJs | Vj Polarity | Gmin | V0 (mV) | A |
|---|---|---|---|---|
| Cx46 | + | 0.24 ± 0.07 1 | 39.1 ± 5.5 | 0.09 ± 0.04 |
| (n = 5) | − | 0.23 ± 0.09 | 38.6 ± 6.8 | 0.11 ± 0.09 |
| L10I | + | 0.22 ± 0.30 | 56.1 ± 17.8 | 0.06 ± 0.06 |
| (n = 4) | − | N/A | N/A | N/A |
| N13E | + | 0.17 ± 0.04 | 33.6 ± 3.6 | 0.16 ± 0.06 |
| (n = 4) | − | 0.17 ± 0.04 | 32.9 ± 1.8 | 0.15 ± 0.06 |
| A14V | + | 0.24 ± 0.04 | 35.1 ± 4.2 | 0.19 ± 0.14 |
| (n = 4) | − | 0.15 ± 0.04 | 37.1 ± 3.0 | 0.14 ± 0.08 |
| Q15N | + | 0.27 ± 0.12 | 37.3 ± 12.4 | 0.06 ± 0.04 |
| (n = 4) | − | 0.25 ± 0.08 | 32.3 ± 7.8 | 0.09 ± 0.06 |
| GJs | Vj Polarity | Gmin | V0 (mV) | A |
|---|---|---|---|---|
| Cx50 | + | 0.20 ± 0.15 1 | 41.5 ± 11.8 | 0.07 ± 0.05 |
| (n = 6) | − | 0.25 ± 0.15 | 43.6 ± 11.8 | 0.07 ± 0.05 |
| I10L | + | 0.22 ± 0.04 | 33.0 ± 5.0 | 0.11 ± 0.04 |
| (n = 4) | − | 0.22 ± 0.06 | 33.7 ± 4.2 | 0.17 ± 0.10 |
| E13N | + | 0.16 ± 0.12 | 33.2 ± 6.9 | 0.1 ± 0.17 |
| (n = 3) | − | 0.29 ± 0.14 | 30.0 ± 11.7 | 0.1 ± 0.14 |
| V14A | + | 0.23 ± 0.08 | 45.4 ± 6.8 | 0.13 ± 0.10 |
| (n = 4) | − | 0.23 ± 0.08 | 37.8 ± 5.8 | 0.14 ± 0.14 |
| N15Q | + | 0.23 ± 0.13 | 37.1 ± 12.5 | 0.08 ± 0.07 |
| (n = 5) | − | 0.18 ± 0.11 | 30.6 ± 11.7 | 0.09 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaradat, R.; Li, X.; Chen, H.; Stathopulos, P.B.; Bai, D. The Hydrophobic Residues in Amino Terminal Domains of Cx46 and Cx50 Are Important for Their Gap Junction Channel Ion Permeation and Gating. Int. J. Mol. Sci. 2022, 23, 11605. https://doi.org/10.3390/ijms231911605
Jaradat R, Li X, Chen H, Stathopulos PB, Bai D. The Hydrophobic Residues in Amino Terminal Domains of Cx46 and Cx50 Are Important for Their Gap Junction Channel Ion Permeation and Gating. International Journal of Molecular Sciences. 2022; 23(19):11605. https://doi.org/10.3390/ijms231911605
Chicago/Turabian StyleJaradat, Roa’a, Xiaole Li, Honghong Chen, Peter B. Stathopulos, and Donglin Bai. 2022. "The Hydrophobic Residues in Amino Terminal Domains of Cx46 and Cx50 Are Important for Their Gap Junction Channel Ion Permeation and Gating" International Journal of Molecular Sciences 23, no. 19: 11605. https://doi.org/10.3390/ijms231911605
APA StyleJaradat, R., Li, X., Chen, H., Stathopulos, P. B., & Bai, D. (2022). The Hydrophobic Residues in Amino Terminal Domains of Cx46 and Cx50 Are Important for Their Gap Junction Channel Ion Permeation and Gating. International Journal of Molecular Sciences, 23(19), 11605. https://doi.org/10.3390/ijms231911605