
Citicoline Eye Drops Protect Trabecular Meshwork Cells from Oxidative Stress Injury in a 3D In Vitro Glaucoma Model

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
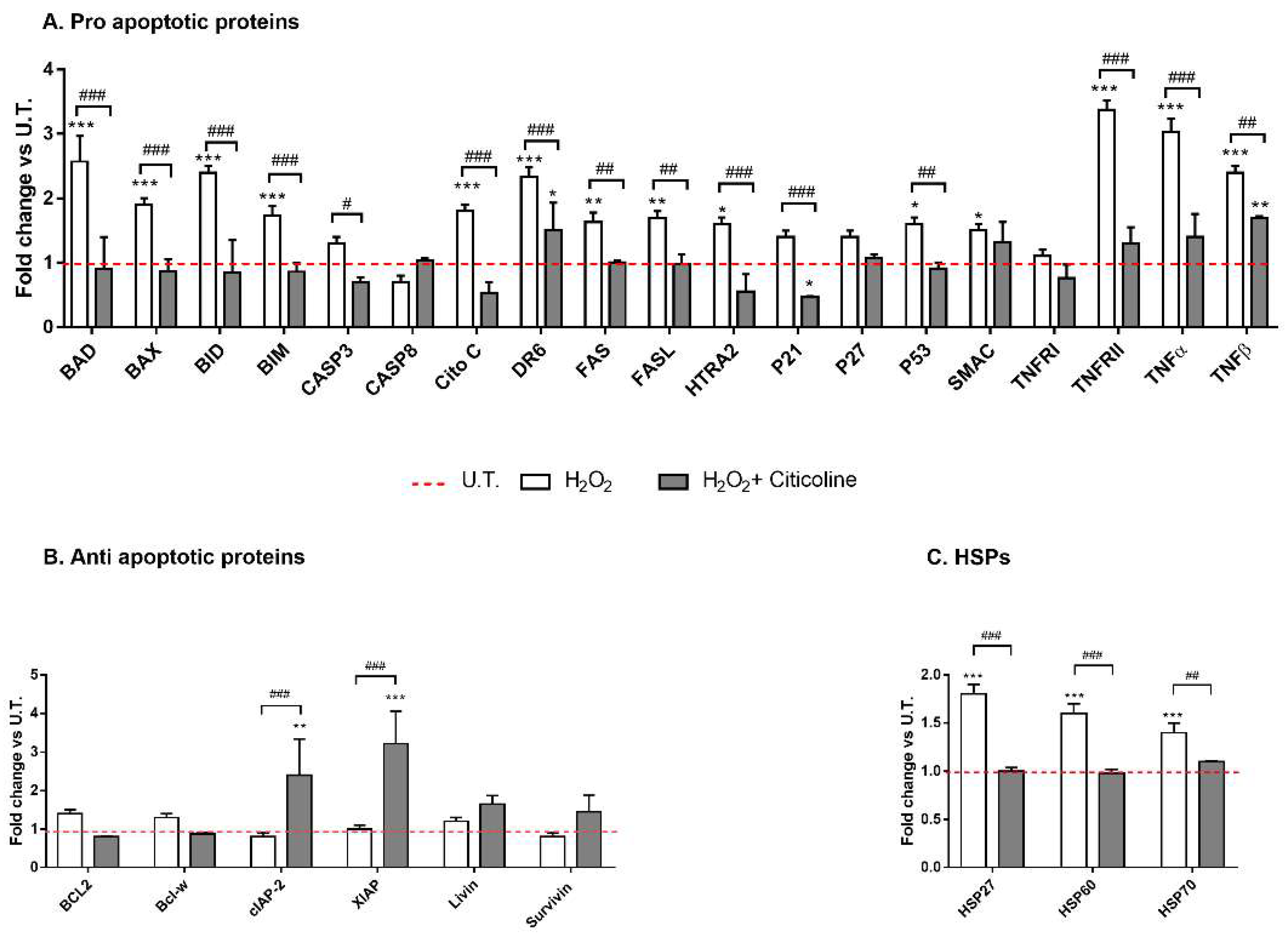
2.1. Citicoline Counteracts the Cytotoxic H2O2-Mediated Effect on HTMC
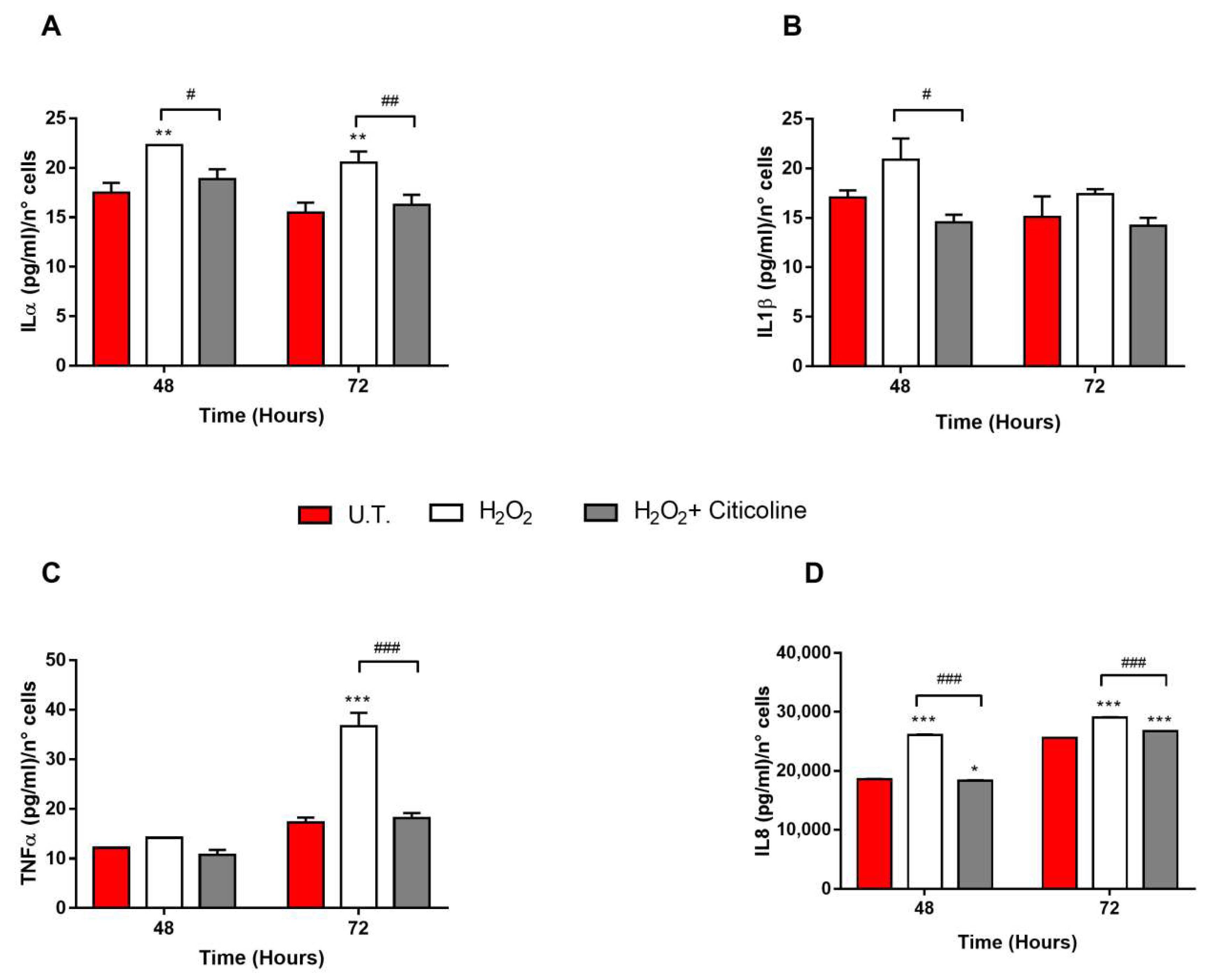
2.2. Citicoline Is Effective in Counteracting H2O2-Induced Cell Senescence and in Reducing the Release of Senescence-Related Pro-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Citicoline Solution
4.3. Experimental Conditions
4.4. DCF Assay
4.5. Viability Assessment
4.6. Apoptosis Array
4.7. Annexin V-FITC and PI Apoptosis Detection Kit
4.8. Mitochondrial Transmembrane Potential Analysis with the JC-1 Fluorochrome
4.9. Senescence-Associated Beta-Galactosidase Activity Analysis
4.10. Pro-Inflammatory Cytokines
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fagone, P.; Jackowski, S. Phosphatidylcholine and the CDP–Choline Cycle. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2013, 1831, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Grieb, P. Neuroprotective Properties of Citicoline: Facts, Doubts and Unresolved Issues. CNS Drugs 2014, 28, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Faiq, M.A.; Wollstein, G.; Schuman, J.S.; Chan, K.C. Cholinergic Nervous System and Glaucoma: From Basic Science to Clinical Applications. Prog. Retin. Eye Res. 2019, 72, 100767. [Google Scholar] [CrossRef]
- Kennedy, E.P.; Weiss, S.B. The Function of Cytidine Coenzymes in the Biosynthesis of Phospholipides. J. Biol. Chem. 1956, 222, 193–214. [Google Scholar] [CrossRef]
- Trovarelli, G.; De Medio, G.E.; Dorman, R.V.; Piccinin, G.L.; Horrocks, L.A.; Porcellati, G. Effect of Cytidine Diphosphate Choline (CDP-Choline) on Ischemia-Induced Alterations of Brain Lipid in the Gerbil. Neurochem. Res. 1981, 6, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Bekdash, R.A. Choline and the Brain: An Epigenetic Perspective. In The Benefits of Natural Products for Neurodegenerative Diseases; Essa, M.M., Akbar, M., Guillemin, G., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, Switzerland, 2016; Volume 12, pp. 381–399. ISBN 978-3-319-28381-4. [Google Scholar]
- Söderberg, M.; Edlund, C.; Kristensson, K.; Dallner, G. Fatty Acid Composition of Brain Phospholipids in Aging and in Alzheimer’s Disease. Lipids 1991, 26, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Medline Clark, W.M.; Warach, S.J.; Pettigrew, L.C. Citicoline. Monograph. Altern. Med. Rev. 2008, 13, 50–57. [Google Scholar]
- Matteucci, A.; Varano, M.; Gaddini, L.; Mallozzi, C.; Villa, M.; Pricci, F.; Malchiodi-Albedi, F. Neuroprotective Effects of Citicoline in in Vitro Models of Retinal Neurodegeneration. Int. J. Mol. Sci. 2014, 15, 6286–6297. [Google Scholar] [CrossRef]
- Ottobelli, L.; Manni, G.L.; Centofanti, M.; Iester, M.; Allevena, F.; Rossetti, L. Citicoline Oral Solution in Glaucoma: Is There a Role in Slowing Disease Progression? Ophthalmologica 2013, 229, 219–226. [Google Scholar] [CrossRef]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Crish, S.D.; Calkins, D.J. Neurodegeneration in Glaucoma: Progression and Calcium-Dependent Intracellular Mechanisms. Neuroscience 2011, 176, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma: A Review. JAMA 2014, 311, 1901. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Tirendi, S.; Bassi, A.M.; Traverso, C.E.; Saccà, S.C. Neuroinflammation in Primary Open-Angle Glaucoma. J. Clin. Med. 2020, 9, 3172. [Google Scholar] [CrossRef]
- Tokuc, E.O.; Yuksel, N.; Kır, H.M.; Acar, E. Evaluation of Serum and Aqueous Humor Klotho Levels in Pseudoexfoliation Syndrome, Pseudoexfoliation and Primary Open-Angle Glaucoma. Int. Ophthalmol. 2021, 41, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Parisi, V.; Oddone, F.; Roberti, G.; Tanga, L.; Carnevale, C.; Ziccardi, L.; Manni, G. Enhancement of Retinal Function and of Neural Conduction Along the Visual Pathway Induced by Treatment with Citicoline Eye Drops in Liposomal Formulation in Open Angle Glaucoma: A Pilot Electrofunctional Study. Adv. Ther. 2019, 36, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, L.; Iester, M.; Tranchina, L.; Ottobelli, L.; Coco, G.; Calcatelli, E.; Ancona, C.; Cirafici, P.; Manni, G. Can Treatment with Citicoline Eyedrops Reduce Progression in Glaucoma? The Results of a Randomized Placebo-Controlled Clinical Trial. J. Glaucoma 2020, 29, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Oddone, F.; Tirendi, S.; Bassi, A.M. Risk Factors for Retinal Ganglion Cell Distress in Glaucoma and Neuroprotective Potential Intervention. Int. J. Mol. Sci. 2021, 22, 7994. [Google Scholar] [CrossRef]
- Crish, S.D.; Sappington, R.M.; Inman, D.M.; Horner, P.J.; Calkins, D.J. Distal Axonopathy with Structural Persistence in Glaucomatous Neurodegeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 5196–5201. [Google Scholar] [CrossRef]
- Buffault, J.; Labbé, A.; Hamard, P.; Brignole-Baudouin, F.; Baudouin, C. The Trabecular Meshwork: Structure, Function and Clinical Implications. A Review of the Literature. J. Fr. d’Ophtalmol. 2020, 43, e217–e230. [Google Scholar] [CrossRef]
- Saccà, S.C.; Gandolfi, S.; Bagnis, A.; Manni, G.; Damonte, G.; Traverso, C.E.; Izzotti, A. From DNA Damage to Functional Changes of the Trabecular Meshwork in Aging and Glaucoma. Ageing Res. Rev. 2016, 29, 26–41. [Google Scholar] [CrossRef]
- Izzotti, A.; Longobardi, M.; Cartiglia, C.; Saccà, S.C. Proteome Alterations in Primary Open Angle Glaucoma Aqueous Humor. J. Proteome Res. 2010, 9, 4831–4838. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chintala, S.K.; Fini, M.E.; Schuman, J.S. Activation of a Tissue-Specific Stress Response in the Aqueous Outflow Pathway of the Eye Defines the Glaucoma Disease Phenotype. Nat. Med. 2001, 7, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Tirendi, S.; Scarfì, S.; Passalacqua, M.; Oddone, F.; Traverso, C.E.; Rizzato, I.; Bassi, A.M.; Saccà, S.C. 2D- and 3D-Cultures of Human Trabecular Meshwork Cells: A Preliminary Assessment of an in Vitro Model for Glaucoma Study. PLoS ONE 2019, 14, e0221942. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.C.; Tirendi, S.; Scarfì, S.; Passalacqua, M.; Oddone, F.; Traverso, C.E.; Vernazza, S.; Bassi, A.M. An Advanced in Vitro Model to Assess Glaucoma Onset. ALTEX Altern. Anim. Exp. 2020, 37, 265–274. [Google Scholar] [CrossRef]
- Singh, P.K.; Kasetti, R.B.; Zode, G.S.; Goyal, A.; Juzych, M.S.; Kumar, A. Zika Virus Infects Trabecular Meshwork and Causes Trabeculitis and Glaucomatous Pathology in Mouse Eyes. mSphere 2019, 4, e00173-19. [Google Scholar] [CrossRef]
- Rieck, J. The Pathogenesis of Glaucoma in the Interplay with the Immune System. Invest. Ophthalmol. Vis. Sci. 2013, 54, 2393. [Google Scholar] [CrossRef]
- Poyomtip, T. Roles of Toll-Like Receptor 4 for Cellular Pathogenesis in Primary Open-Angle Glaucoma: A Potential Therapeutic Strategy. J. Microbiol. Immunol. Infect. 2019, 52, 201–206. [Google Scholar] [CrossRef]
- Buffault, J.; Brignole-Baudouin, F.; Reboussin, É.; Kessal, K.; Labbé, A.; Mélik Parsadaniantz, S.; Baudouin, C. The Dual Effect of Rho-Kinase Inhibition on Trabecular Meshwork Cells Cytoskeleton and Extracellular Matrix in an In Vitro Model of Glaucoma. J. Clin. Med. 2022, 11, 1001. [Google Scholar] [CrossRef]
- Waduthanthri, K.D.; He, Y.; Montemagno, C.; Cetinel, S. An Injectable Peptide Hydrogel for Reconstruction of the Human Trabecular Meshwork. Acta Biomater. 2019, 100, 244–254. [Google Scholar] [CrossRef]
- Torrejon, K.Y.; Papke, E.L.; Halman, J.R.; Stolwijk, J.; Dautriche, C.N.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. Bioengineered Glaucomatous 3D Human Trabecular Meshwork as an in Vitro Disease Model: Steroid-Induced Glaucomatous 3D HTM Model. Biotechnol. Bioeng. 2016, 113, 1357–1368. [Google Scholar] [CrossRef]
- Aires, I.D.; Ambrósio, A.F.; Santiago, A.R. Modeling Human Glaucoma: Lessons from the in Vitro Models. Ophthalmic Res. 2017, 57, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Bouchemi, M.; Roubeix, C.; Kessal, K.; Riancho, L.; Raveu, A.-L.; Soualmia, H.; Baudouin, C.; Brignole-Baudouin, F. Effect of Benzalkonium Chloride on Trabecular Meshwork Cells in a New in Vitro 3D Trabecular Meshwork Model for Glaucoma. Toxicol. In Vitro 2017, 41, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Haderspeck, J.C.; Chuchuy, J.; Kustermann, S.; Liebau, S.; Loskill, P. Organ-on-a-Chip Technologies That Can Transform Ophthalmic Drug Discovery and Disease Modeling. Expert Opin. Drug Discov. 2019, 14, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Vernazza, S.; Tirendi, S.; Passalacqua, M.; Piacente, F.; Scarfì, S.; Oddone, F.; Bassi, A.M. An Innovative In Vitro Open-Angle Glaucoma Model (IVOM) Shows Changes Induced by Increased Ocular Pressure and Oxidative Stress. Int. J. Mol. Sci. 2021, 22, 12129. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the AlamarBlue Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb.prot095489. [Google Scholar] [CrossRef]
- Ok, C.Y.; Park, S.; Jang, H.-O.; Takata, T.; Lee, O.-H.; Bae, M.-K.; Bae, S.-K. FK866 Protects Human Dental Pulp Cells against Oxidative Stress-Induced Cellular Senescence. Antioxidants 2021, 10, 271. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Giraldi, J.P.; Virno, M.; Covelli, G.; Grechi, G.; De Gregorio, F. Therapeutic Value of Citicoline in the Treatment of Glaucoma (Computerized and Automated Perimetric Investigation). Int. Ophthalmol. 1989, 13, 109–112. [Google Scholar] [CrossRef]
- Parisi, V.; Centofanti, M.; Ziccardi, L.; Tanga, L.; Michelessi, M.; Roberti, G.; Manni, G. Treatment with Citicoline Eye Drops Enhances Retinal Function and Neural Conduction along the Visual Pathways in Open Angle Glaucoma. Graefes Arch. Clin. Exp. Ophthalmol. 2015, 253, 1327–1340. [Google Scholar] [CrossRef]
- Roberti, G.; Tanga, L.; Parisi, V.; Sampalmieri, M.; Centofanti, M.; Manni, G. A Preliminary Study of the Neuroprotective Role of Citicoline Eye Drops in Glaucomatous Optic Neuropathy. Indian J. Ophthalmol. 2014, 62, 549. [Google Scholar] [CrossRef]
- Izzotti, A.; Saccà, S.C.; Longobardi, M.; Cartiglia, C. Sensitivity of Ocular Anterior Chamber Tissues to Oxidative Damage and Its Relevance to the Pathogenesis of Glaucoma. Invest. Ophthalmol. Vis. Sci. 2009, 50, 5251. [Google Scholar] [CrossRef] [PubMed]
- Saccà, S.C.; Vernazza, S.; Iorio, E.L.; Tirendi, S.; Bassi, A.M.; Gandolfi, S.; Izzotti, A. Molecular Changes in Glaucomatous Trabecular Meshwork. Correlations with Retinal Ganglion Cell Death and Novel Strategies for Neuroprotection. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 256, pp. 151–188. ISBN 978-0-12-821106-9. [Google Scholar]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxidative Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [PubMed]
- Baudouin, C.; Kolko, M.; Melik-Parsadaniantz, S.; Messmer, E.M. Inflammation in Glaucoma: From the Back to the Front of the Eye, and Beyond. Prog. Retin. Eye Res. 2021, 83, 100916. [Google Scholar] [CrossRef] [PubMed]
- Grieb, P.; Rejdak, R. Pharmacodynamics of Citicoline Relevant to the Treatment of Glaucoma. J. Neurosci. Res. 2002, 67, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Ucciferri, N.; Sbrana, T.; Ahluwalia, A. Allometric Scaling and Cell Ratios in Multi-Organ in Vitro Models of Human Metabolism. Front. Bioeng. Biotechnol. 2014, 2, 74. [Google Scholar] [CrossRef]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Solomon, F.D.P. 3D Cell Culture Systems: Advantages and Applications. J. Cell. Physiol 2015, 230, 16–26. [Google Scholar] [CrossRef]
- Tandon, N.; Marolt, D.; Cimetta, E.; Vunjak-Novakovic, G. Bioreactor Engineering of Stem Cell Environments. Biotechnol. Adv. 2013, 31, 1020–1031. [Google Scholar] [CrossRef]
- Saccà, S.C.; Izzotti, A.; Vernazza, S.; Tirendi, S.; Scarfì, S.; Gandolfi, S.; Bassi, A.M. Can Polyphenols in Eye Drops Be Useful for Trabecular Protection from Oxidative Damage? J. Clin. Med. 2020, 9, 3584. [Google Scholar] [CrossRef]
- Keller, K.E.; Bhattacharya, S.K.; Borrás, T.; Brunner, T.M.; Chansangpetch, S.; Clark, A.F.; Dismuke, W.M.; Du, Y.; Elliott, M.H.; Ethier, C.R.; et al. Consensus Recommendations for Trabecular Meshwork Cell Isolation, Characterization and Culture. Exp. Eye Res. 2018, 171, 164–173. [Google Scholar] [CrossRef]
- Vernazza, S. HTMCs 3D Culture Method Using Corning Matrigel Matrix V1. PLoS ONE 2019, 14, e0221942. [Google Scholar]
- Wang, H.; Joseph, J.A. Quantifying Cellular Oxidative Stress by Dichlorofluorescein Assay Using Microplate Reader11Mention of a Trade Name, Proprietary Product, or Specific Equipment Does Not Constitute a Guarantee by the United States Department of Agriculture and Does Not Imply Its Approval to the Exclusion of Other Products That May Be Suitable. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A Biomarker That Identifies Senescent Human Cells in Culture and in Aging Skin In Vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vernazza, S.; Passalacqua, M.; Tirendi, S.; Marengo, B.; Domenicotti, C.; Sbardella, D.; Oddone, F.; Bassi, A.M. Citicoline Eye Drops Protect Trabecular Meshwork Cells from Oxidative Stress Injury in a 3D In Vitro Glaucoma Model. Int. J. Mol. Sci. 2022, 23, 11375. https://doi.org/10.3390/ijms231911375
Vernazza S, Passalacqua M, Tirendi S, Marengo B, Domenicotti C, Sbardella D, Oddone F, Bassi AM. Citicoline Eye Drops Protect Trabecular Meshwork Cells from Oxidative Stress Injury in a 3D In Vitro Glaucoma Model. International Journal of Molecular Sciences. 2022; 23(19):11375. https://doi.org/10.3390/ijms231911375
Chicago/Turabian StyleVernazza, Stefania, Mario Passalacqua, Sara Tirendi, Barbara Marengo, Cinzia Domenicotti, Diego Sbardella, Francesco Oddone, and Anna Maria Bassi. 2022. "Citicoline Eye Drops Protect Trabecular Meshwork Cells from Oxidative Stress Injury in a 3D In Vitro Glaucoma Model" International Journal of Molecular Sciences 23, no. 19: 11375. https://doi.org/10.3390/ijms231911375
APA StyleVernazza, S., Passalacqua, M., Tirendi, S., Marengo, B., Domenicotti, C., Sbardella, D., Oddone, F., & Bassi, A. M. (2022). Citicoline Eye Drops Protect Trabecular Meshwork Cells from Oxidative Stress Injury in a 3D In Vitro Glaucoma Model. International Journal of Molecular Sciences, 23(19), 11375. https://doi.org/10.3390/ijms231911375