
Connecting Aortic Stiffness to Vascular Contraction: Does Sex Matter?


 ,
,  ,
, 
Abstract
1. Introduction
2. Results
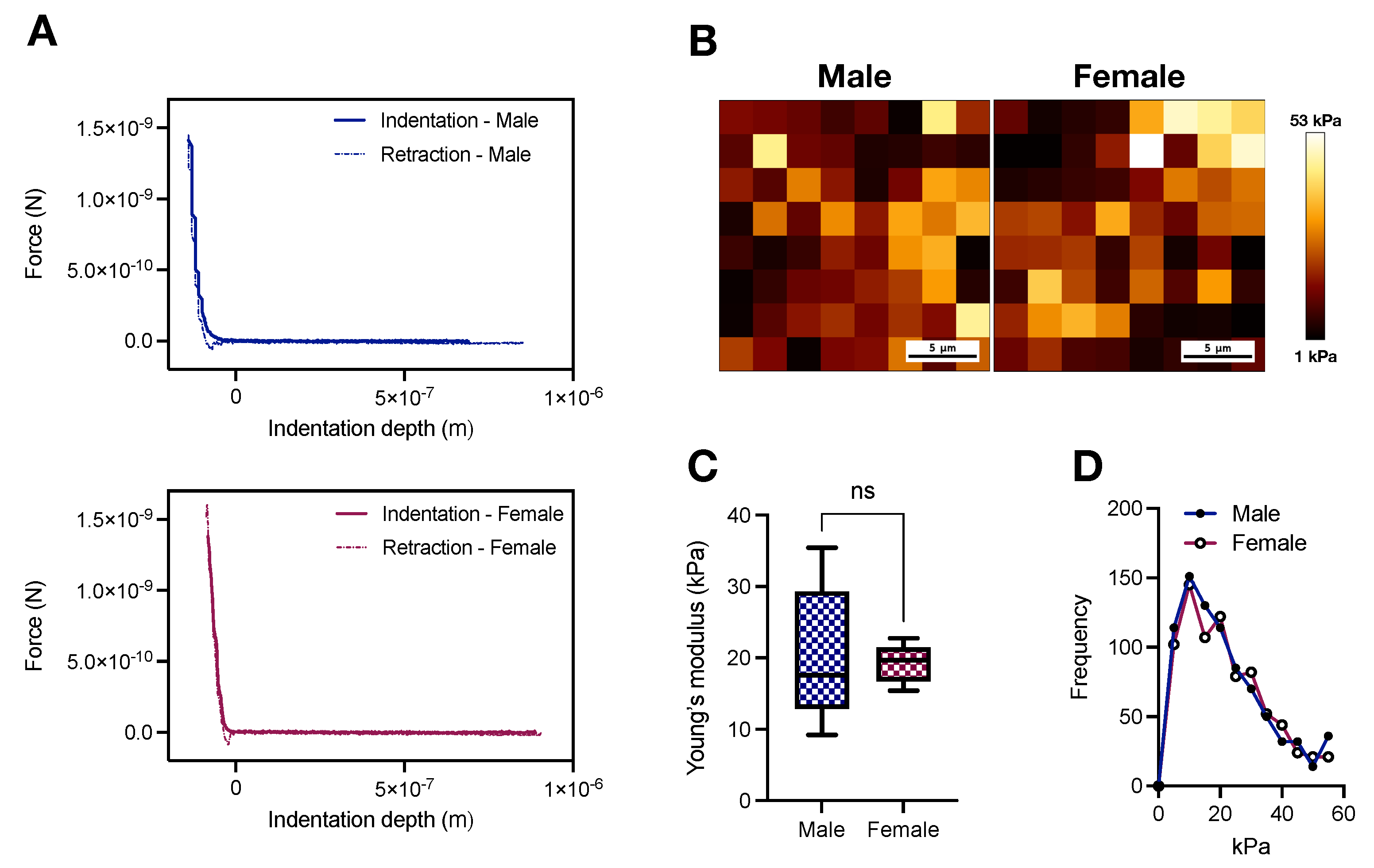
2.1. AFM Stiffness Mapping of the Thoracic Aorta of Young Male and Female Wistar Rats
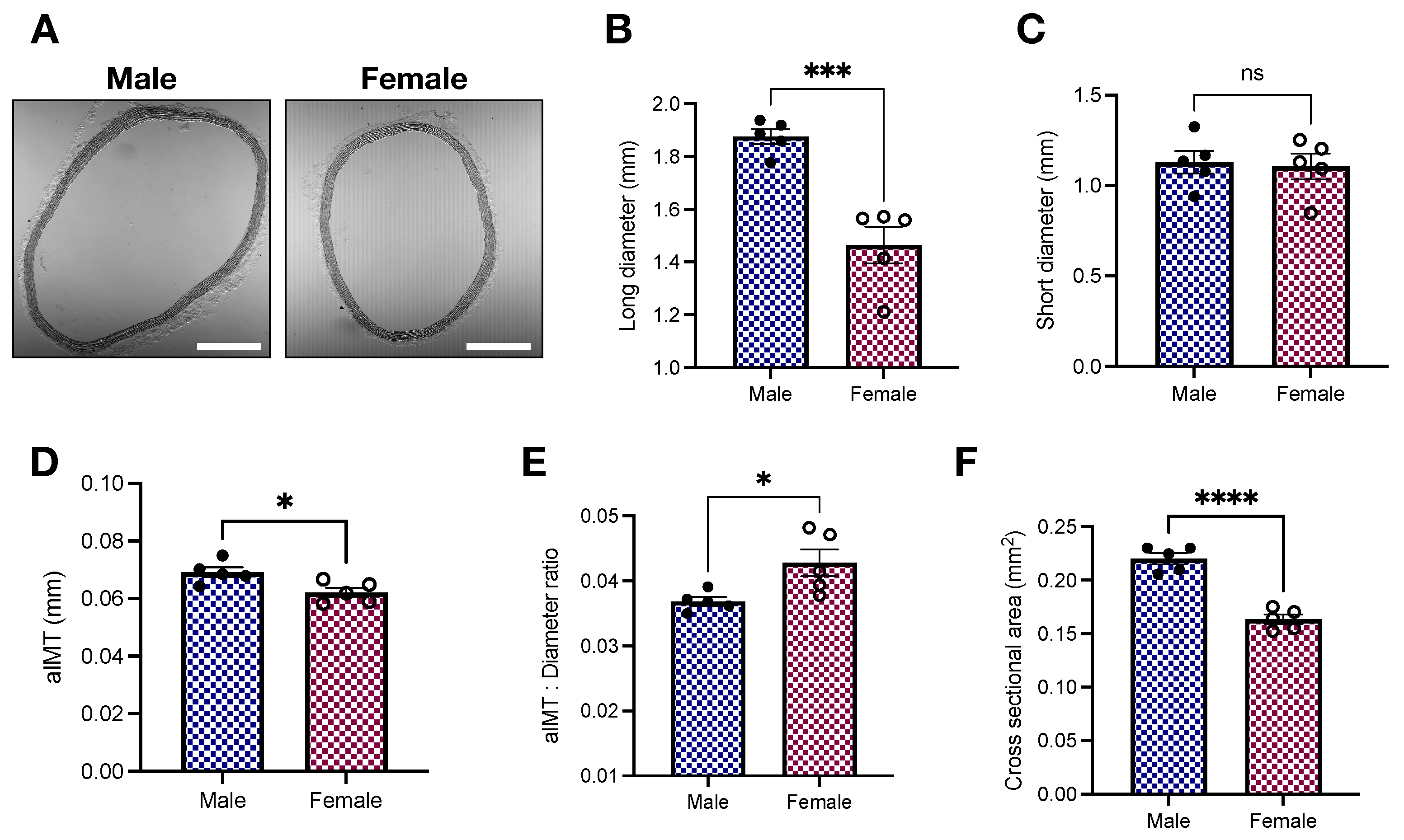
2.2. Histomorphometric Analysis of the Thoracic Aorta of Young Male and Female Wistar Rats
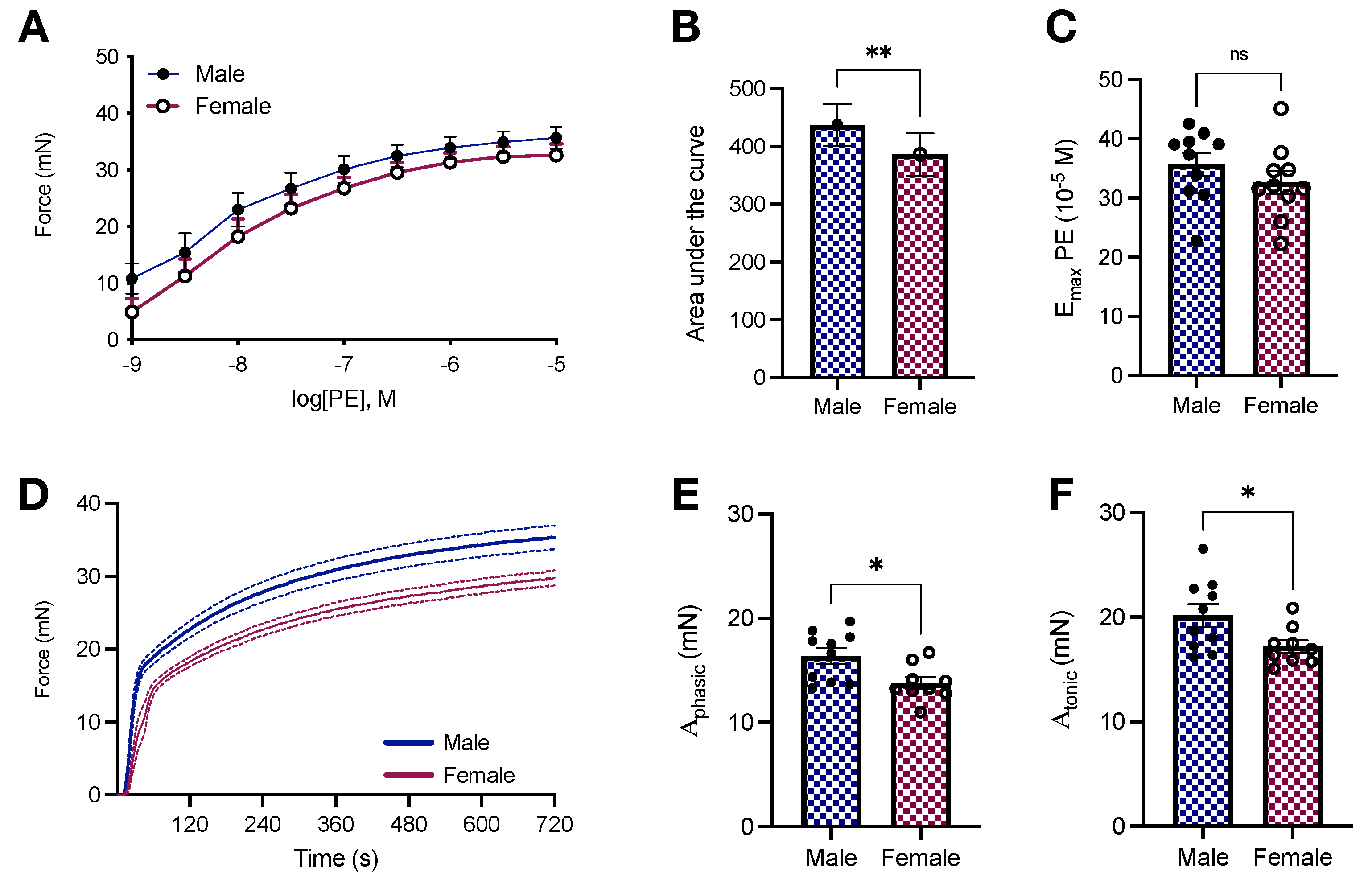
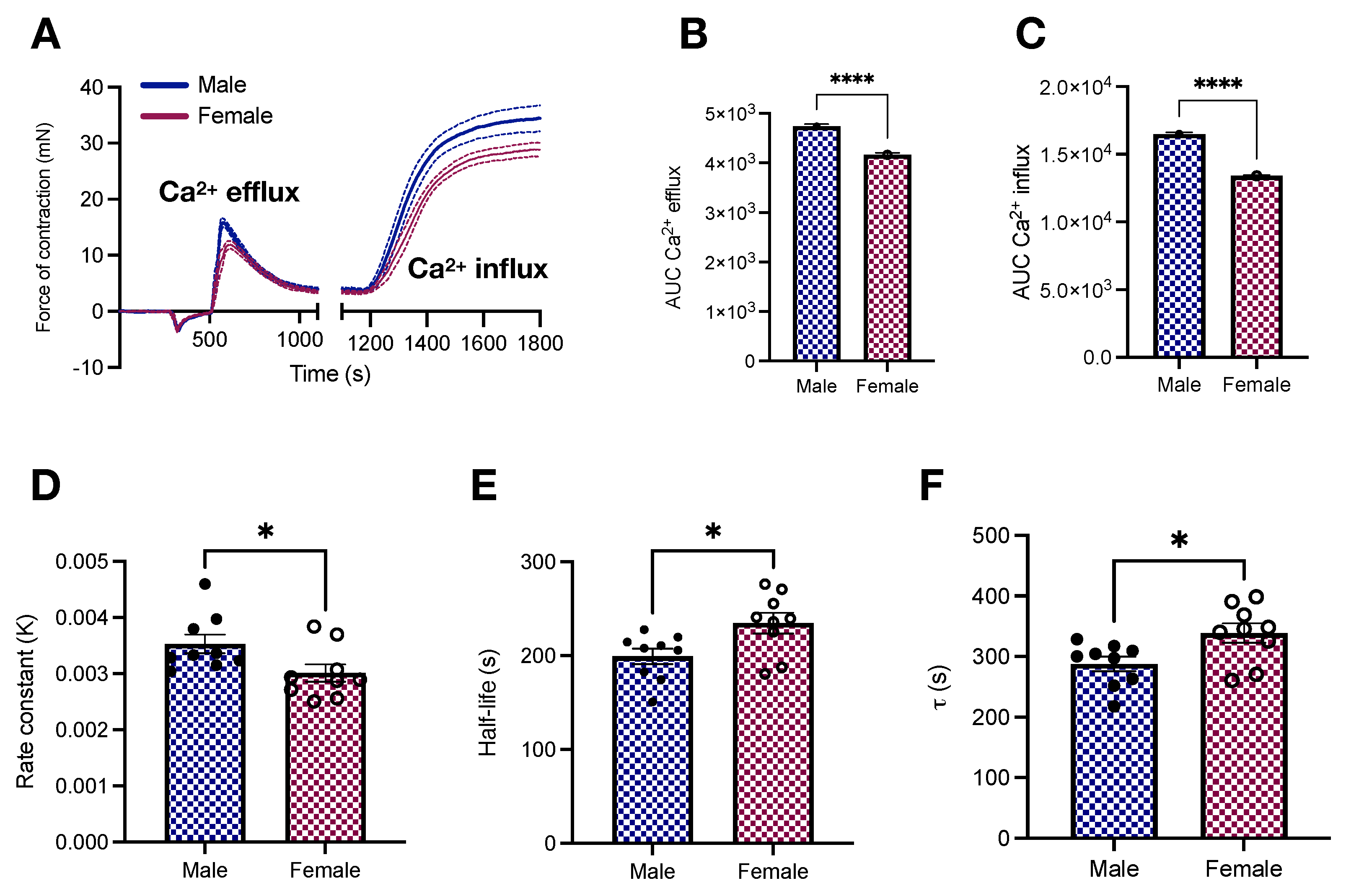
2.3. Vascular Responses to Phenylephrine in Young Male and Female Wistar Rats
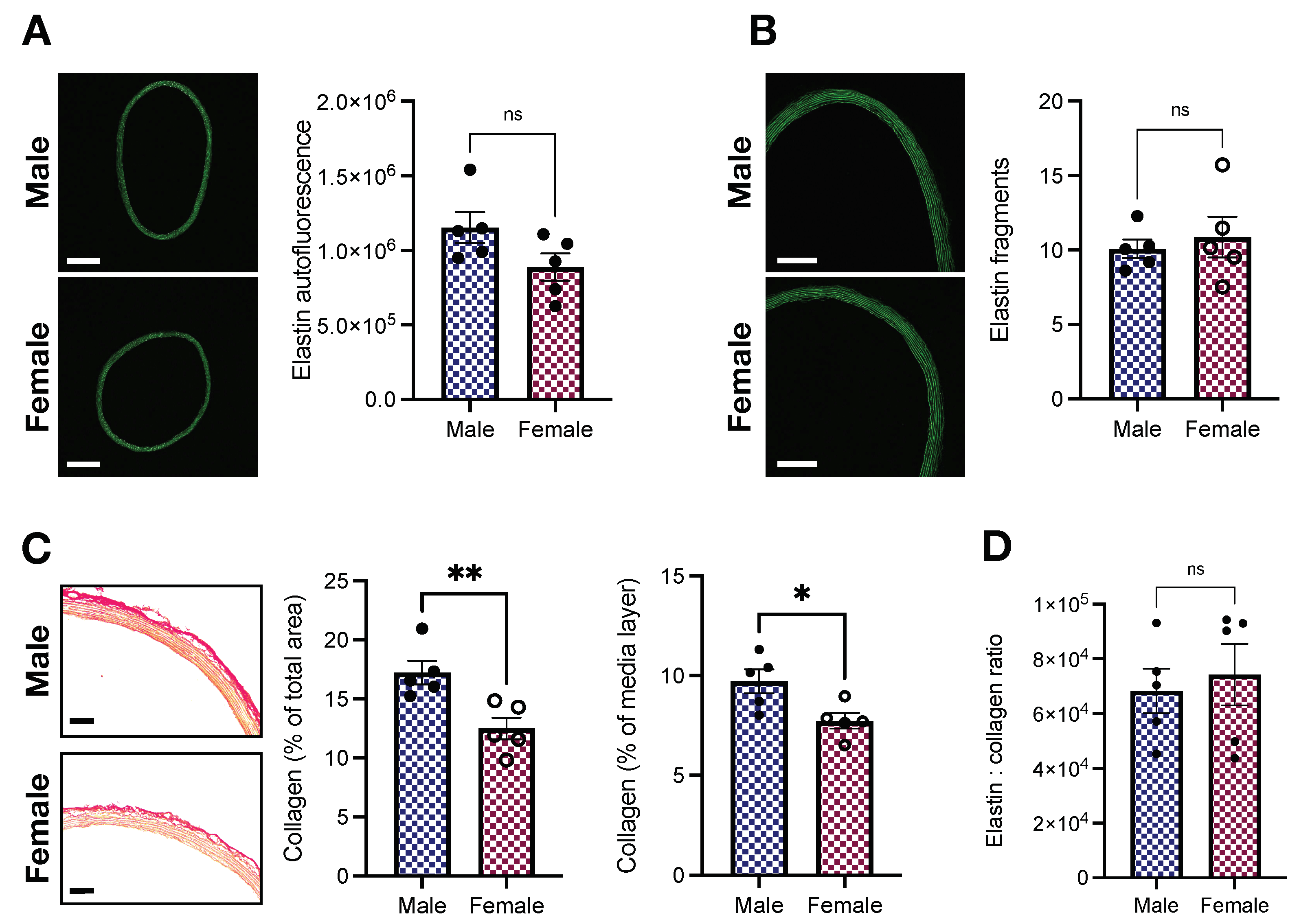
2.4. Elastin and Collagen Levels in the Aorta of Young Male and Female Wistar Rats
3. Discussion
4. Methods and Materials
4.1. Animals
4.2. Atomic Force Microscopy
4.3. Histomorphometric Analysis
4.4. Functional Studies
4.5. Elastin Characterization
4.6. Collagen Detection
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mosca, L.; Barrett-Connor, E.; Kass Wenger, N. Sex/gender differences in cardiovascular disease prevention: What a difference a decade makes. Circulation 2011, 124, 2145–2154. [Google Scholar] [CrossRef] [PubMed]
- Regitz-Zagrosek, V.; Kararigas, G. Mechanistic pathways of sex differences in cardiovascular disease. Physiol. Rev. 2017, 97, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, T. Arterial stiffness and its clinical implications in women. Can. J. Cardiol. 2014, 30, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Ahimastos, A.A.; Formosa, M.; Dart, A.M.; Kingwell, B.A. Gender differences in large artery stiffness pre-and post puberty. J. Clin. Endocrinol. Metab. 2003, 88, 5375–5380. [Google Scholar] [CrossRef]
- Coutinho, T.; Borlaug, B.A.; Pellikka, P.A.; Turner, S.T.; Kullo, I.J. Sex differences in arterial stiffness and ventricular-arterial interactions. J. Am. Coll. Cardiol. 2013, 61, 96–103. [Google Scholar] [CrossRef]
- Lu, Y.; Pechlaner, R.; Cai, J.; Yuan, H.; Huang, Z.; Yang, G.; Wang, J.; Chen, Z.; Kiechl, S.; Xu, Q. Trajectories of age-related arterial stiffness in Chinese men and women. J. Am. Coll. Cardiol. 2020, 75, 870–880. [Google Scholar] [CrossRef]
- Bonarjee, V.V. Arterial stiffness: A prognostic marker in coronary heart disease. Available methods and clinical application. Front. Cardiovasc. Med. 2018, 5, 64. [Google Scholar] [CrossRef]
- Mitchell, G.F.; Hwang, S.J.; Vasan, R.S.; Larson, M.G.; Pencina, M.J.; Hamburg, N.M.; Vita, J.A.; Levy, D.; Benjamin, E.J. Arterial stiffness and cardiovascular events: The Framingham Heart Study. Circulation 2010, 121, 505–511. [Google Scholar] [CrossRef]
- Willum Hansen, T.; Staessen, J.A.; Torp-Pedersen, C.; Rasmussen, S.; Thijs, L.; Ibsen, H.; Jeppesen, J. Prognostic value of aortic pulse wave velocity as index of arterial stiffness in the general population. Circulation 2006, 113, 664–670. [Google Scholar] [CrossRef]
- Zieman, S.J.; Melenovsky, V.; Kass, D.A. Mechanisms, pathophysiology, and therapy of arterial stiffness. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 932–943. [Google Scholar] [CrossRef]
- Mitchell, G.F.; Guo, C.Y.; Benjamin, E.J.; Larson, M.G.; Keyes, M.J.; Vita, J.A.; Vasan, R.S.; Levy, D. Cross-sectional correlates of increased aortic stiffness in the community: The Framingham Heart Study. Circulation 2007, 115, 2628–2636. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.D.; Bulpitt, C.J.; Pinto, E.S.; Rajkumar, C. The aging of elastic and muscular arteries: A comparison of diabetic and nondiabetic subjects. Diabetes Care 2003, 26, 2133–2138. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, E.; Shoji, T.; Shinohara, K.; Inaba, M.; Okuno, Y.; Miki, T.; Koyama, H.; Emoto, M.; Nishizawa, Y. Preferential stiffening of central over peripheral arteries in type 2 diabetes. Diabetes 2003, 52, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Sehgel, N.L.; Sun, Z.; Hong, Z.; Hunter, W.C.; Hill, M.A.; Vatner, D.E.; Vatner, S.F.; Meininger, G.A. Augmented vascular smooth muscle cell stiffness and adhesion when hypertension is superimposed on aging. Hypertension 2015, 65, 370–377. [Google Scholar] [CrossRef]
- Kim, H.L.; Ahn, D.W.; Kim, S.H.; Lee, D.S.; Yoon, S.H.; Zo, J.H.; Kim, M.A.; Jeong, J.B. Association between body fat parameters and arterial stiffness. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- DuPont, J.J.; Kenney, R.M.; Patel, A.R.; Jaffe, I.Z. Sex differences in mechanisms of arterial stiffness. Br. J. Pharmacol. 2019, 176, 4208–4225. [Google Scholar] [CrossRef]
- Butlin, M.; Tan, I.; Spronck, B.; Avolio, A.P. Measuring arterial stiffness in animal experimental studies. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1068–1077. [Google Scholar] [CrossRef]
- Tuttle, T.; Darios, E.; Watts, S.W.; Roccabianca, S. Aortic stiffness is lower when PVAT is included: A novel ex vivo mechanics study. Am. J. Physiol.-Heart Circ. Physiol. 2022, 322, H1003–H1013. [Google Scholar] [CrossRef]
- Bae, Y.H.; Liu, S.L.; Byfield, F.J.; Janmey, P.A.; Assoian, R.K. Measuring the stiffness of ex vivo mouse aortas using atomic force microscopy. JoVE 2016, 1, e54630. [Google Scholar] [CrossRef]
- Zhu, Y. Atomic force microscopy reveals the role of vascular smooth muscle cell elasticity in hypertension. Recent Dev. At. Force Microsc. Raman Spectrosc. Mater. Charact. 2020, 61. [Google Scholar] [CrossRef]
- McCallinhart, P.E.; Cho, Y.; Sun, Z.; Ghadiali, S.; Meininger, G.A.; Trask, A.J. Reduced stiffness and augmented traction force in type 2 diabetic coronary microvascular smooth muscle. Am. J. Physiol.-Heart Circ. Physiol. 2020, 318, H1410–H1419. [Google Scholar] [CrossRef] [PubMed]
- Targosz-Korecka, M.; Jaglarz, M.; Malek-Zietek, K.E.; Gregorius, A.; Zakrzewska, A.; Sitek, B.; Rajfur, Z.; Chlopicki, S.; Szymonski, M. AFM-based detection of glycocalyx degradation and endothelial stiffening in the db/db mouse model of diabetes. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhu, Y.; Sun, Z.; Trzeciakowski, J.P.; Gansner, M.; Depre, C.; Resuello, R.R.; Natividad, F.F.; Hunter, W.C.; Genin, G.M.; et al. Vascular smooth muscle cell stiffness as a mechanism for increased aortic stiffness with aging. Circ. Res. 2010, 107, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Leloup, A.J.; Van Hove, C.E.; De Moudt, S.; De Meyer, G.R.; De Keulenaer, G.W.; Fransen, P. Vascular smooth muscle cell contraction and relaxation in the isolated aorta: A critical regulator of large artery compliance. Physiol. Rep. 2019, 7, e13934. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, R.; Mihai, C.; Kniss, D.; Ghadiali, S.N. Finite element analysis of traction force microscopy: Influence of cell mechanics, adhesion, and morphology. J. Biomech. Eng. 2013, 135, 071009. [Google Scholar] [CrossRef]
- Camasão, D.; Mantovani, D. The mechanical characterization of blood vessels and their substitutes in the continuous quest for physiological-relevant performances. A critical review. Mater. Today Bio 2021, 10, 100106. [Google Scholar] [CrossRef]
- Semmler, L.; Weberruß, H.; Baumgartner, L.; Pirzer, R.; Oberhoffer-Fritz, R. Vascular diameter and intima-media thickness to diameter ratio values of the carotid artery in 642 healthy children. Eur. J. Pediatr. 2021, 180, 851–860. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; Priviero, F.; Webb, R.C.; Nunes, K.P. Impaired HSP70 expression in the aorta of female rats: A novel insight into sex-specific differences in vascular function. Front. Physiol. 2021, 12, 666696. [Google Scholar] [CrossRef]
- Giachini, F.R.; Lima, V.V.; Filgueira, F.P.; Dorrance, A.M.; Carvalho, M.H.C.; Fortes, Z.B.; Webb, R.C.; Tostes, R.C. STIM1/Orai1 contributes to sex differences in vascular responses to calcium in spontaneously hypertensive rats. Clin. Sci. 2012, 122, 215–226. [Google Scholar] [CrossRef]
- Loria, A.S.; Brinson, K.N.; Fox, B.M.; Sullivan, J.C. Sex-specific alterations in NOS regulation of vascular function in aorta and mesenteric arteries from spontaneously hypertensive rats compared to Wistar Kyoto rats. Physiol. Rep. 2014, 2, e12125. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Díez, J. Arterial stiffness and extracellular matrix. Atheroscler. Large Arter. Cardiovasc. Risk 2007, 44, 76–95. [Google Scholar] [CrossRef]
- Ogola, B.O.; Zimmerman, M.A.; Clark, G.L.; Abshire, C.M.; Gentry, K.M.; Miller, K.S.; Lindsey, S.H. New insights into arterial stiffening: Does sex matter? Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H1073–H1087. [Google Scholar] [CrossRef]
- Sehgel, N.L.; Zhu, Y.; Sun, Z.; Trzeciakowski, J.P.; Hong, Z.; Hunter, W.C.; Vatner, D.E.; Meininger, G.A.; Vatner, S.F. Increased vascular smooth muscle cell stiffness: A novel mechanism for aortic stiffness in hypertension. Am. J. Physiol.-Heart Circ. Physiol. 2013, 305, H1281–H1287. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Chao, H.; Tang, B.; Avolio, A.P.; Schlaich, M.P.; Nolde, J.M.; Adji, A.; Carnagarin, R. Female gender is associated with higher susceptibility of weight induced arterial stiffening and rise in blood pressure. J. Clin. Med. 2021, 10, 3479. [Google Scholar] [CrossRef] [PubMed]
- Nunes, K.P.; Webb, R.C. New insights into RhoA/Rho-kinase signaling: A key regulator of vascular contraction. Small GTPases 2021, 12, 458–469. [Google Scholar] [CrossRef]
- Hayashi, T.; Fukuto, J.M.; Ignarro, L.J.; Chaudhuri, G. Basal release of nitric oxide from aortic rings is greater in female rabbits than in male rabbits: Implications for atherosclerosis. Proc. Natl. Acad. Sci. USA 1992, 89, 11259–11263. [Google Scholar] [CrossRef]
- Delgado, A.; Dong, P.; Sameti, M.; Zimin, V.N.; Lee, J.; Gharaibeh, Y.; Bezerra, H.G.; Wilson, D.; Bashur, C.; Gu, L. Mechanical Characterization of Calcification in Diseased Coronary Artery with Atomic Force Microscope. In Proceedings of the Frontiers in Biomedical Devices; American Society of Mechanical Engineers: New York, NY, USA, 2022; Volume 84815, p. V001T01A004. [Google Scholar] [CrossRef]
- Radmacher, M. Measuring the elastic properties of biological samples with the AFM. IEEE Eng. Med. Biol. Mag. 1997, 16, 47–57. [Google Scholar] [CrossRef]
- de Oliveira, A.A.; Faustino, J.; Webb, R.C.; Nunes, K.P. Blockade of the TLR4–MD2 complex lowers blood pressure and improves vascular function in a murine model of type 1 diabetes. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- de Oliveira, A.A.; Nunes, K.P. An additional physiological role for HSP70: Assistance of vascular reactivity. Life Sci. 2020, 256, 117986. [Google Scholar] [CrossRef]
- de Oliveira, A.A.; Priviero, F.; Tostes, R.C.; Webb, R.C.; Nunes, K.P. Dissecting the interaction between HSP70 and vascular contraction: Role of Ca2+ handling mechanisms. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fransen, P.; Van Hove, C.E.; Leloup, A.J.; Martinet, W.; De Meyer, G.R.; Lemmens, K.; Bult, H.; Schrijvers, D.M. Dissecting out the complex Ca2+-mediated phenylephrine-induced contractions of mouse aortic segments. PLoS ONE 2015, 10, e0121634. [Google Scholar] [CrossRef] [PubMed]
- Romier, B.; Dray, C.; Vanalderwiert, L.; Wahart, A.; Hocine, T.; Dortignac, A.; Garbar, C.; Garbar, C.; Boulagnon, C.; Bouland, N.; et al. Apelin expression deficiency in mice contributes to vascular stiffening by extracellular matrix remodeling of the aortic wall. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (g) | Glucose Levels (mg/dl) | Mean Arterial Pressure (mmHg) |
|---|---|---|---|
| Male | 495.4 ± 9.74 | 154.2 ± 18.02 | 87.6 ± 3.93 |
| Female | 228.4 ± 5.32 | 181.4 ± 8.99 | 89.8 ± 2.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, A.A.; Priviero, F.; Delgado, A.; Dong, P.; Mendoza, V.O.; Gu, L.; Webb, R.C.; Nunes, K.P. Connecting Aortic Stiffness to Vascular Contraction: Does Sex Matter? Int. J. Mol. Sci. 2022, 23, 11314. https://doi.org/10.3390/ijms231911314
de Oliveira AA, Priviero F, Delgado A, Dong P, Mendoza VO, Gu L, Webb RC, Nunes KP. Connecting Aortic Stiffness to Vascular Contraction: Does Sex Matter? International Journal of Molecular Sciences. 2022; 23(19):11314. https://doi.org/10.3390/ijms231911314
Chicago/Turabian Stylede Oliveira, Amanda A., Fernanda Priviero, Ana Delgado, Pengfei Dong, Valentina O. Mendoza, Linxia Gu, R. Clinton Webb, and Kenia P. Nunes. 2022. "Connecting Aortic Stiffness to Vascular Contraction: Does Sex Matter?" International Journal of Molecular Sciences 23, no. 19: 11314. https://doi.org/10.3390/ijms231911314
APA Stylede Oliveira, A. A., Priviero, F., Delgado, A., Dong, P., Mendoza, V. O., Gu, L., Webb, R. C., & Nunes, K. P. (2022). Connecting Aortic Stiffness to Vascular Contraction: Does Sex Matter? International Journal of Molecular Sciences, 23(19), 11314. https://doi.org/10.3390/ijms231911314