
Gene Expression and DNA Methylation in Human Papillomavirus Positive and Negative Head and Neck Squamous Cell Carcinomas
 ,
,  and
and
Abstract
1. Background
2. Results
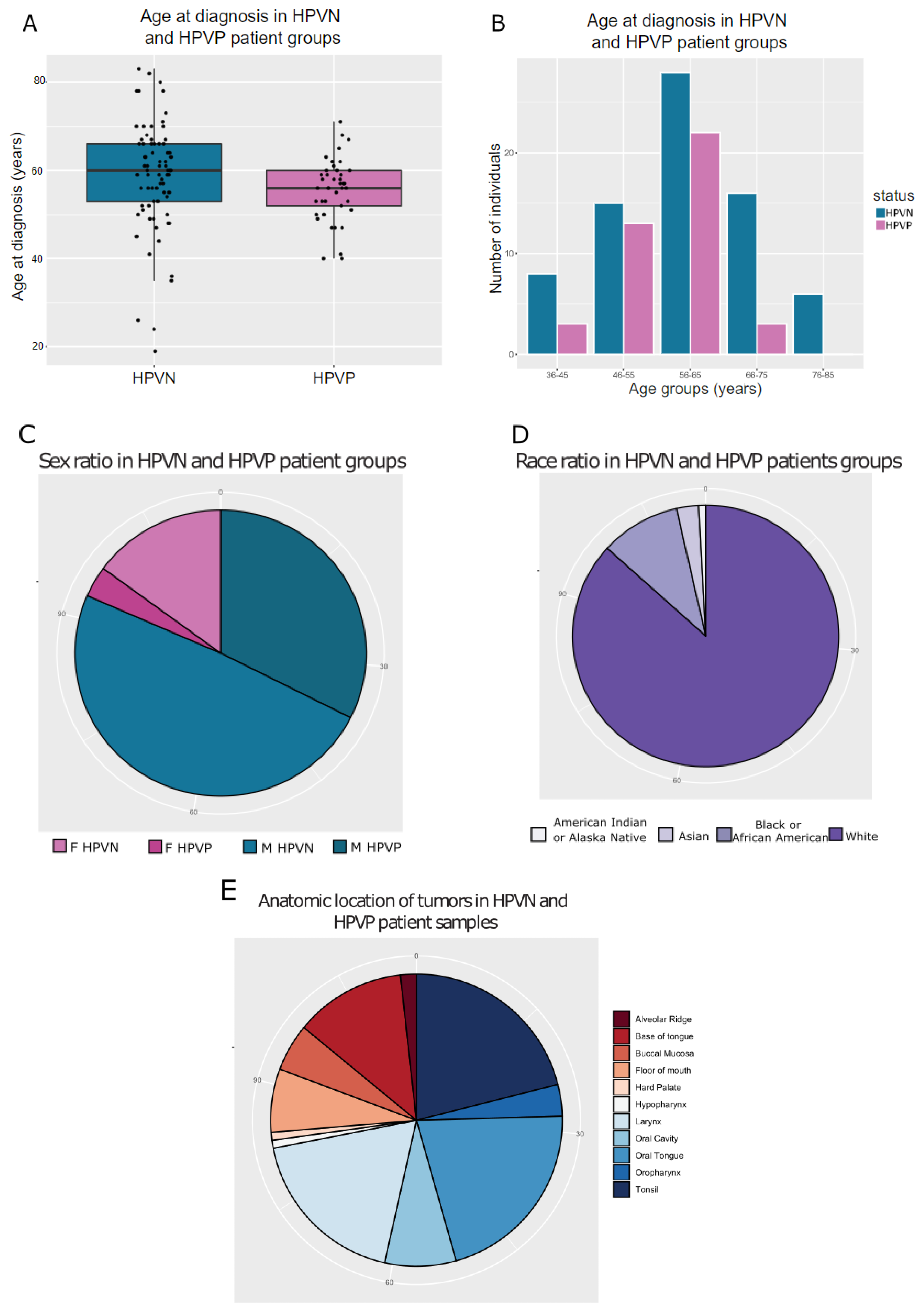
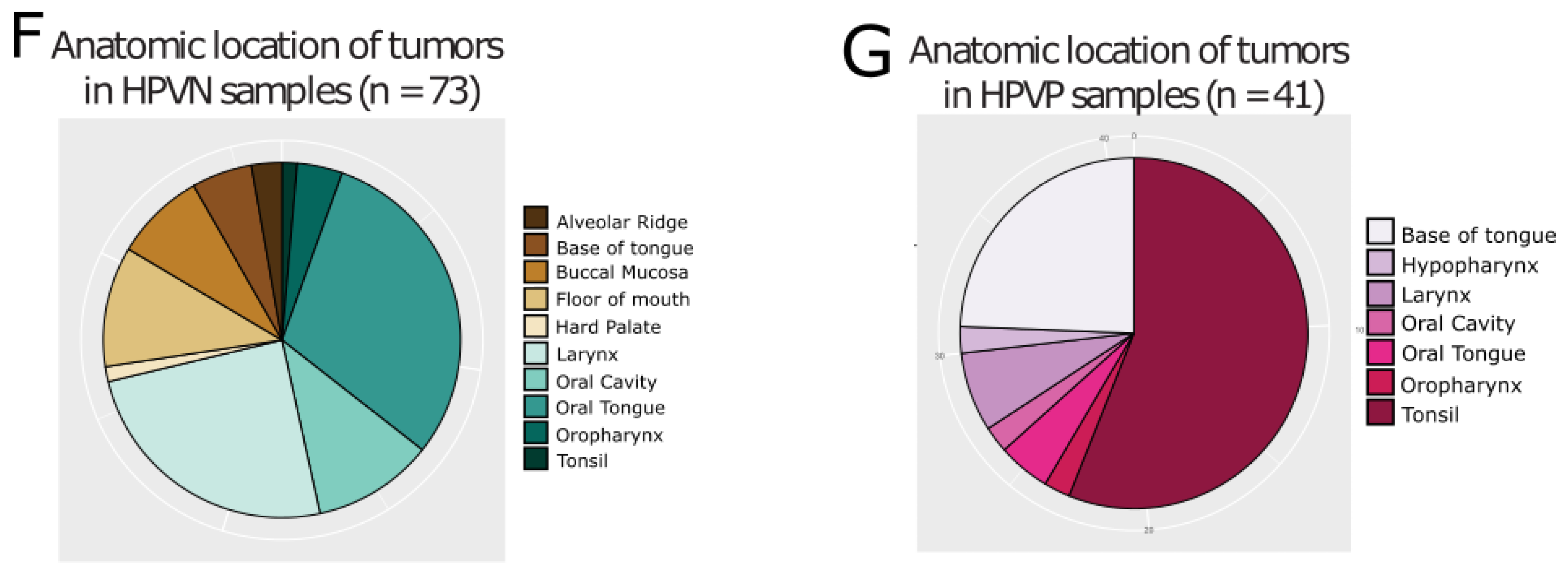
2.1. TCGA HNSCC HPVP and HPVN Patients Have Comparable Clinical History
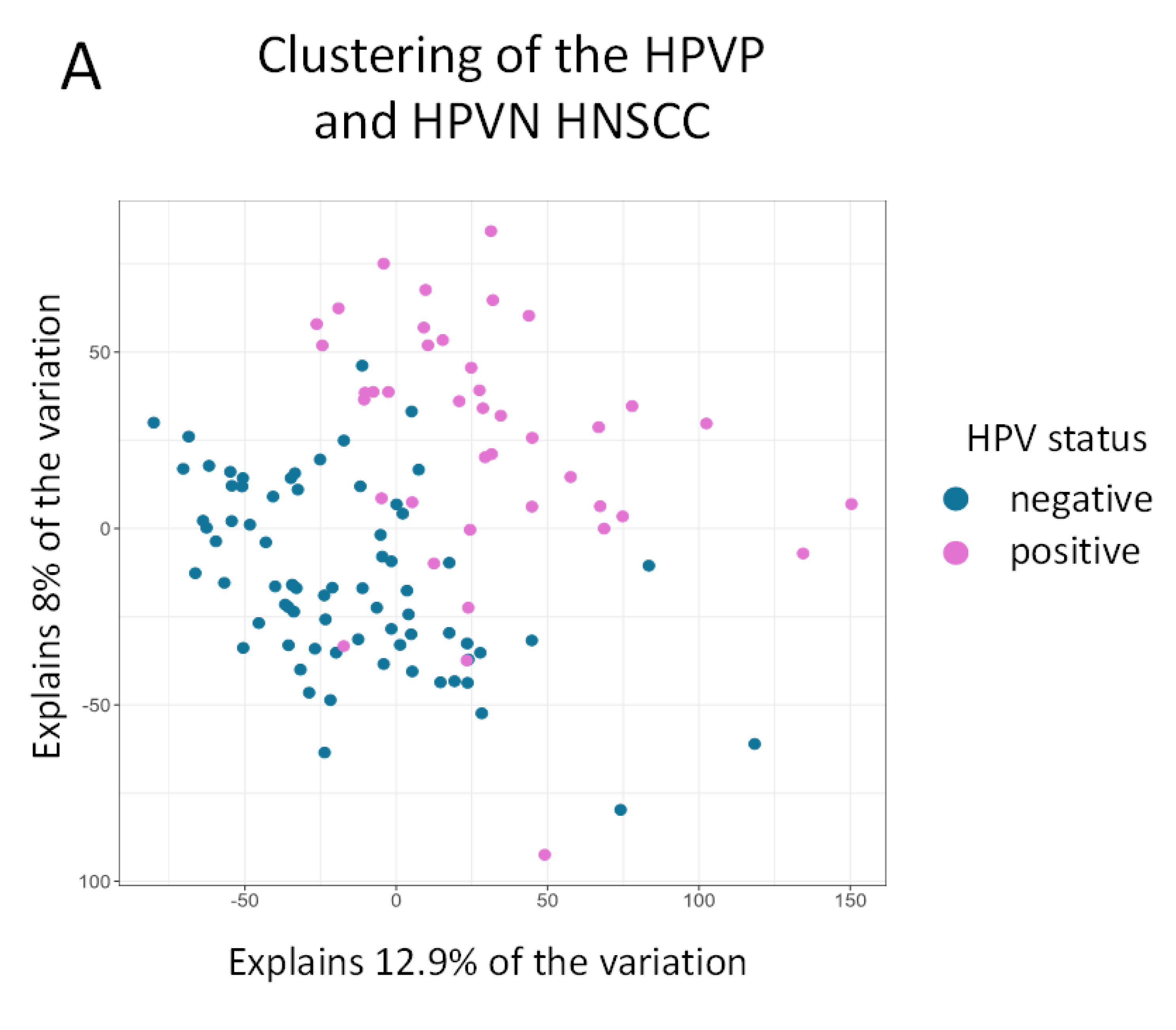
2.2. Clustering of Samples Confirms That HPVN and HPVP Are Two Separate Comparable Groups
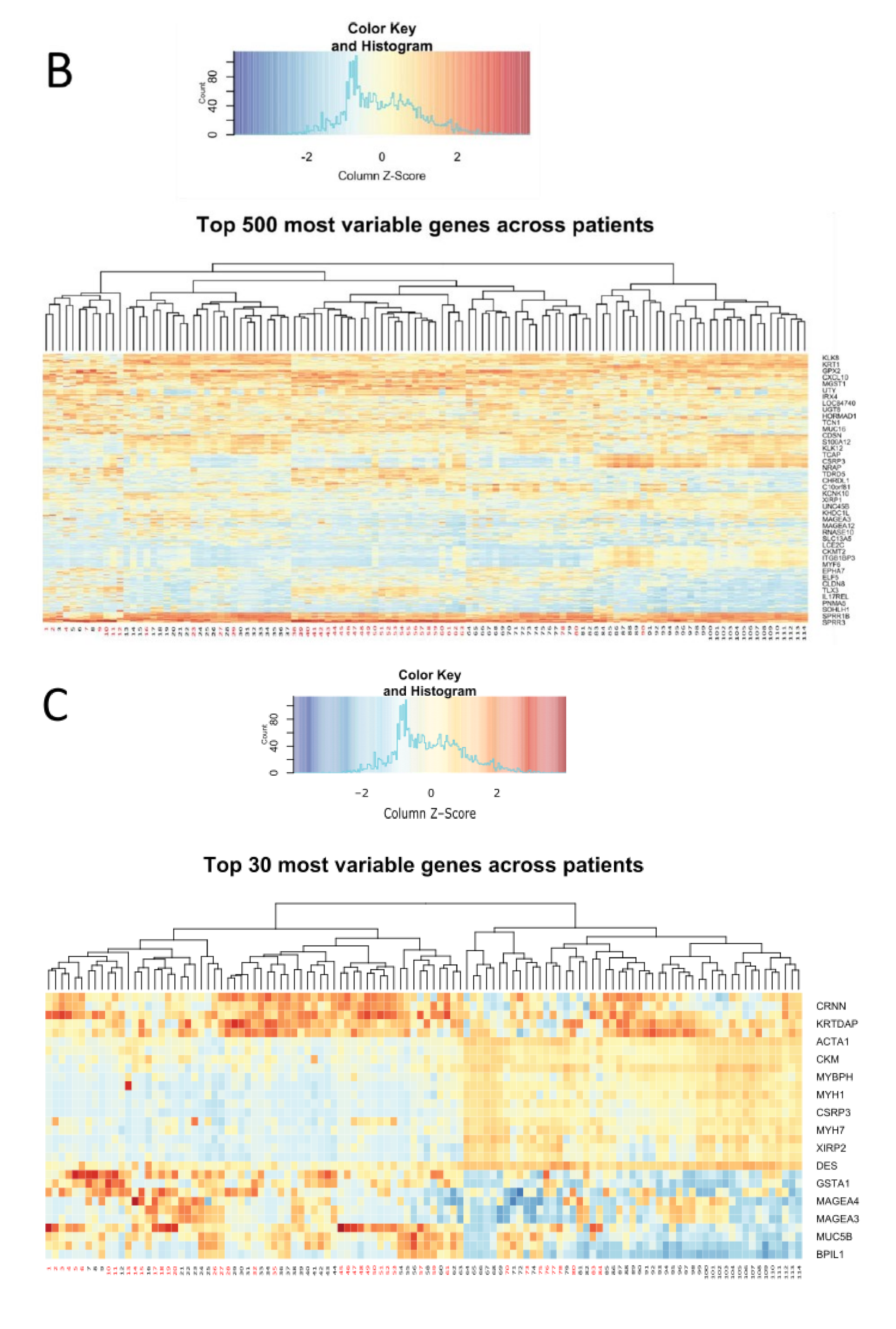
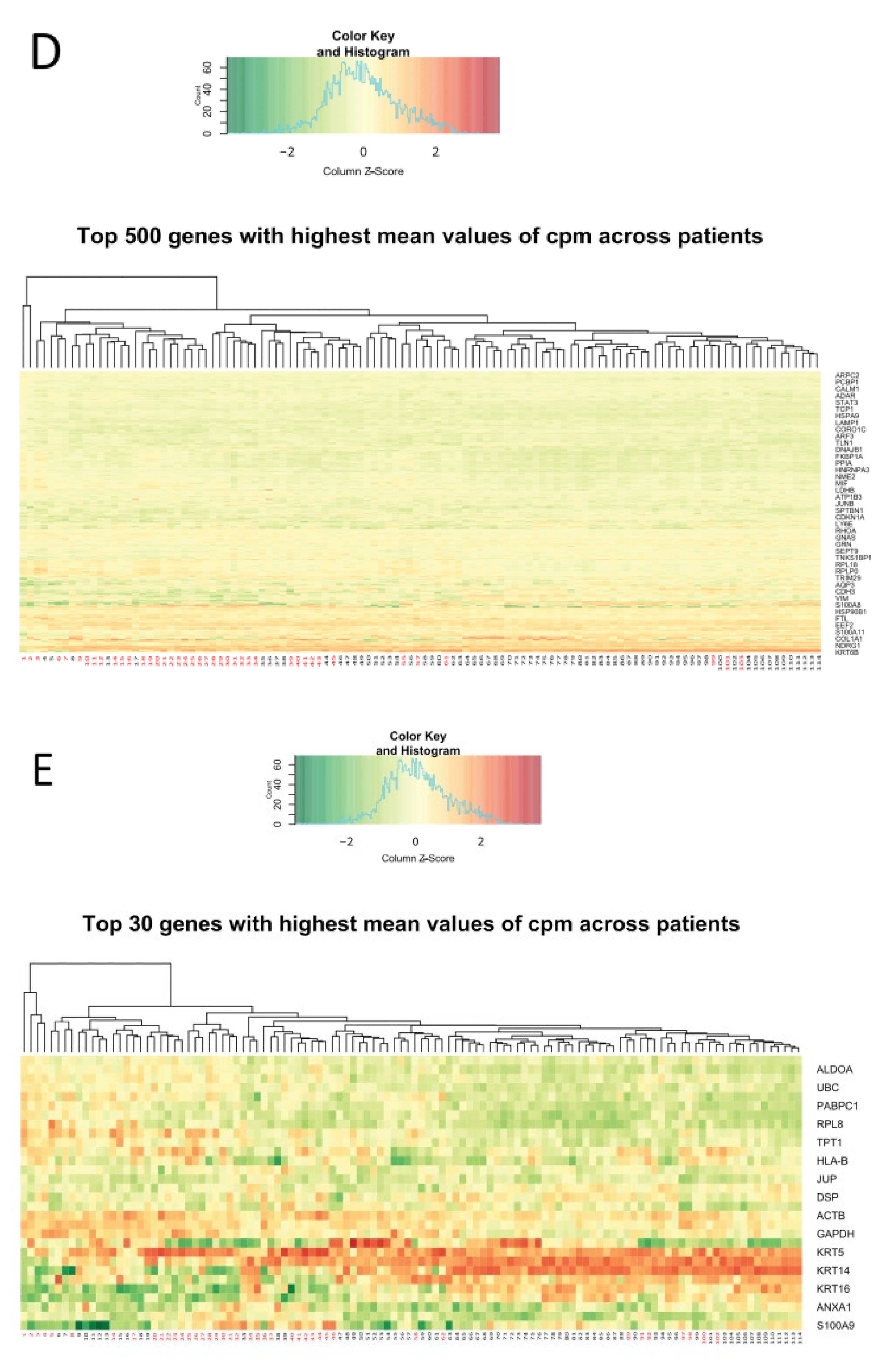
- The pattern of specific groups of genes in a larger scale analysis of the top 500 most variable genes (Figure 2B) and of the top 30 most variable genes (Figure 2C), remained consistent. Genes including kallikreins family genes (serine proteases) remained highly variable between the patients [28,29]. Keratin, a structural component was found as variable, as well as oxidative damage protection proteins (GPX2), cytokines, inflammatory response genes, immune response genes, and cell cycle controlling genes. Figure 2C depicts a gene responsible for stratification of the skin (KRTDAP), that was highly variably expressed, as well as an epithelial immune response and differentiation gene (CRNN). At a larger scale, genes from two groups of patients seemed to cluster mostly separately, significantly resembling the clustering observed in PCA (Figure 2A), with an intermediate overlapping cluster of samples (Figure 2B).
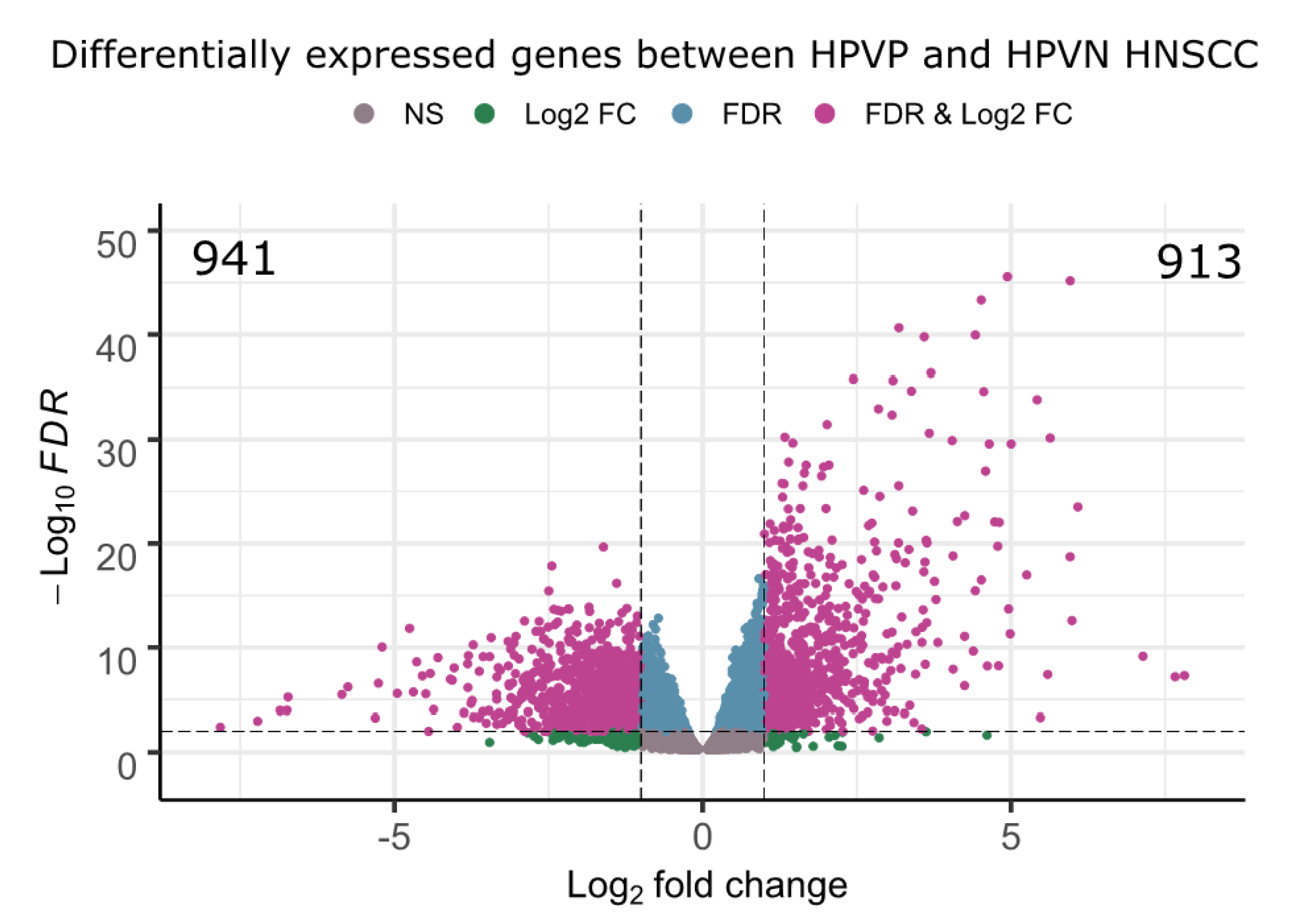
2.3. Transcriptome Analysis Identified 1854 Differentially Expressed Genes among HPVN and HPVP HNSCC Groups
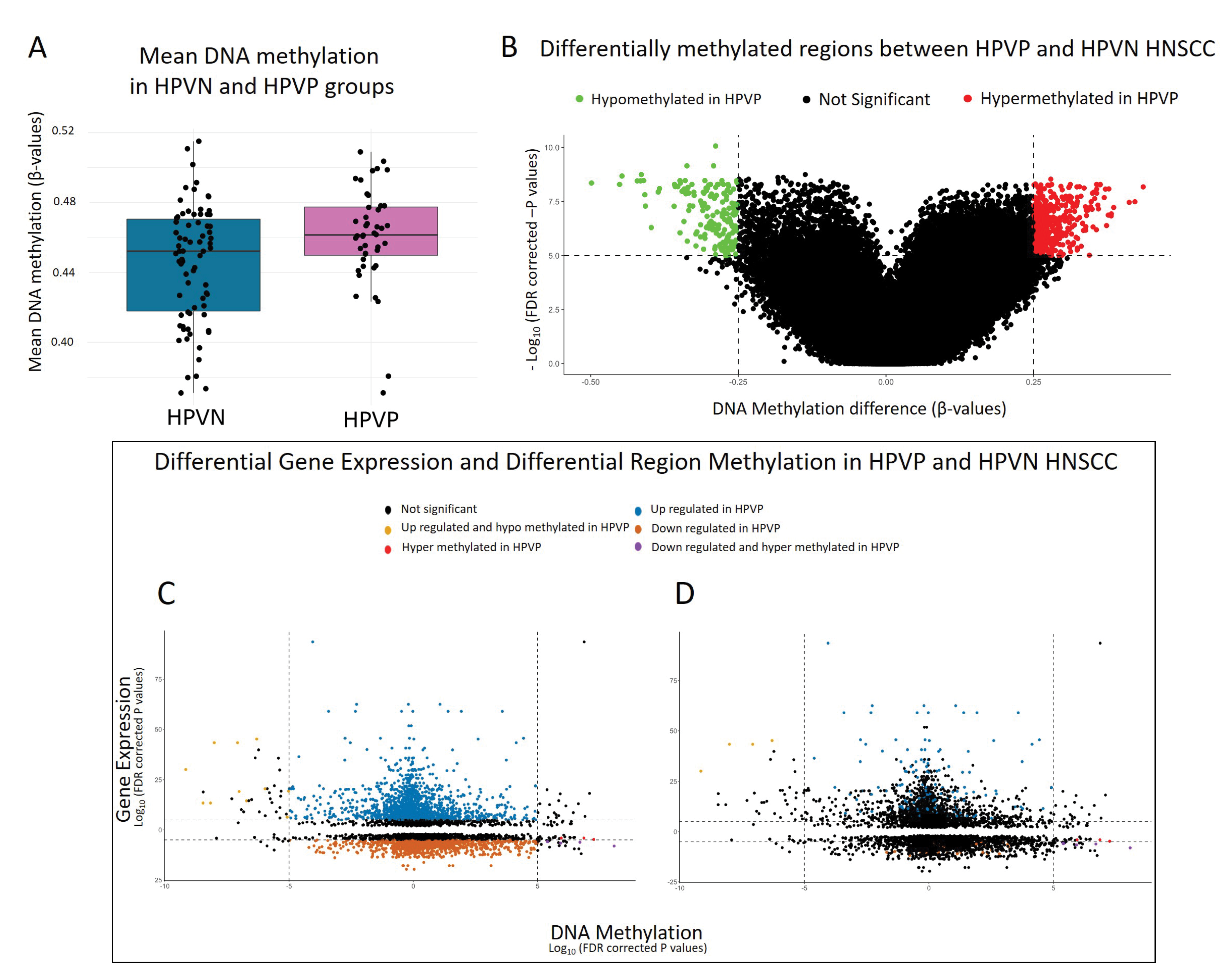
2.4. DNA Methylome Analysis Showed HPVP and HPVN HNSCC Methylation Levels Were Comparable
2.5. Starburst: An Analysis That Bridges Differentially Expressed and Methylated Genes Revealed Similar Patterns to DNA Methylome Analysis and Potential Biomarker Gene for HPVP HNSCC
3. Discussion
4. Methods
4.1. Study Design, Patient Samples, and Analysis Workflow
4.2. Data Preprocessing to Normalize Data
4.3. Data Clustering Analyses
4.4. Transcriptome Analysis: Differential Gene Expression Analysis (DGE) and Pathway Analysis
4.5. DNA Methylome Analysis: Differential Methylation Analysis (DMA)
4.6. Starburst Analysis: Integrative Analysis of DEG and Differentially Methylated Regions (DMR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HPV | Human Papillomavirus |
| HNSCC | head and neck squamous cell carcinoma |
| pRb | Retinoblastoma protein |
| PCNA | proliferating cell nuclear antigen |
| TNFRSF14 | Tumor Necrosis Factor (TNF) Receptor Superfamily Member 14 |
| TRAF1 | TNF Receptor Associated Factor 1 |
| TRAF2 | TNF Receptor Associated Factor 2 |
| BCL2 | B-cell lymphoma 2 apoptosis regulator |
| BIRC3 | Baculoviral Inhibitor of Apoptosis (IAP) Repeat Containing 3 |
| SYCP2 | Synaptonemal Complex Protein 2 |
| TAF7L | TATA-Box Binding Protein Associated Factor 7-Like |
| E6 | HPV early gene 6 |
| E7 | HPV early gene 7 |
| TP53/p53 | tumor suppressor gene/protein p53 |
| HPVN | HPV negative HNSCC |
| HPVP | HPV positive HNSCC |
| p16 | cyclin-dependent kinase inhibitor |
| CCND1 | cyclin D1 |
| TCGA | The Cancer Genome Atlas |
| GDC | NCI Genomic Data Commons |
| PCA | Principal Component Analysis |
| DGE | differential gene expression |
| DMA | differential methylation analysis |
| DMR | differential methylated regions |
| GPX2 | Glutathione peroxidase 2 |
| KRTDAP | Keratinocyte Differentiation Associated Protein |
| CRNN | Cornulin |
| PCA | Principal component analysis |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| ECM | extracellular matrix |
| PANTHER | protein annotation through evolutionary relationship |
| THPA | The Human Protein Atlas |
| TLR | Toll-like receptor |
| NLR | Nod-like receptor |
| NF-kβ | nuclear factor kappa-light-chain-enhancer of activated B cells |
| ZFR2 | Zinc Finger RNA Binding Protein 2 |
| INHBA | Inhibin Subunit Beta A |
References
- Global Cancer Observatory. Available online: https://gco.iarc.fr/ (accessed on 30 August 2022).
- Worldwide Cancer Data|World Cancer Research Fund International. Available online: https://www.wcrf.org/cancer-trends/worldwide-cancer-data/ (accessed on 30 August 2022).
- Jethwa, A.R.; Khariwala, S.S. Tobacco-related carcinogenesis in head and neck cancer. Cancer Metastasis Rev. 2017, 36, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Maier, H.; Dietz, A.; Gewelke, U.; Heller, W.D.; Weidauer, H. Tobacco and alcohol and the risk of head and neck cancer. Clin. Investig. 1992, 70, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Kreimer, A.R. Human Papillomavirus Types in Head and Neck Squamous Cell Carcinomas Worldwide: A Systematic Review. Cancer Epidemiol. Biomark. Prev. 2005, 14, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Ragin, C.C.R.; Modugno, F.; Gollin, S.M. The epidemiology and risk factors of head and neck cancer: A focus on human papillomavirus. J. Dent. Res. 2007, 86, 104–114. [Google Scholar] [CrossRef]
- Sano, D.; Oridate, N. The molecular mechanism of human papillomavirus-induced carcinogenesis in head and neck squamous cell carcinoma. Int. J. Clin. Oncol. 2016, 21, 819–826. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hisamatsu, K.; Suzui, N.; Hara, A.; Tomita, H.; Miyazaki, T. A Review of HPV-Related Head and Neck Cancer. J. Clin. Med. 2018, 7, 241. [Google Scholar] [CrossRef]
- Bajos, N.; Bozon, M.; Beltzer, N.; Laborde, C.; Andro, A.; Ferrand, M.; Goulet, V.; Laporte, A.; Le Van, C.; Leridon, H.; et al. Changes in sexual behaviours: From secular trends to public health policies. AIDS Lond. Engl. 2010, 24, 1185–1191. [Google Scholar] [CrossRef]
- Zheng, Z.-M.; Baker, C.C. Papillomavirus genome structure, expression, and post-transcriptional regulation. Front. Biosci. J. Virtual Libr. 2006, 11, 2286–2302. [Google Scholar] [CrossRef]
- Münger, K.; Werness, B.A.; Dyson, N.; Phelps, W.C.; Harlow, E.; Howley, P.M. Complex formation of human papillomavirus E7 proteins with the retinoblastoma tumor suppressor gene product. EMBO J. 1989, 8, 4099–4105. [Google Scholar] [CrossRef]
- Mantovani, F.; Banks, L. The human papillomavirus E6 protein and its contribution to malignant progression. Oncogene 2001, 20, 7874–7887. [Google Scholar] [CrossRef]
- Rumfield, C.S.; Roller, N.; Pellom, S.T.; Schlom, J.; Jochems, C. Therapeutic Vaccines for HPV-Associated Malignancies. ImmunoTargets Ther. 2020, 9, 167–200. [Google Scholar] [CrossRef] [PubMed]
- Sever, R.; Brugge, J.S. Signal transduction in cancer. Cold Spring Harb. Perspect. Med. 2015, 5, a006098. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H.; Paludan, S.R. Molecular Pathways in Virus-Induced Cytokine Production. Microbiol. Mol. Biol. Rev. 2001, 65, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Kotwal, G.J.; Hatch, S.; Marshall, W.L. Viral infection: An evolving insight into the signal transduction pathways responsible for the innate immune response. Adv. Virol. 2012, 2012, 131457. [Google Scholar] [CrossRef]
- Seiwert, T.Y.; Zuo, Z.; Keck, M.K.; Khattri, A.; Pedamallu, C.S.; Stricker, T.; Brown, C.; Pugh, T.J.; Stojanov, P.; Cho, J.; et al. Integrative and comparative genomic analysis of HPV-positive and HPV-negative head and neck squamous cell carcinomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2015, 21, 632–641. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [CrossRef]
- Esposti, D.D.; Sklias, A.; Lima, S.C.; Beghelli-de la Forest Divonne, S.; Cahais, V.; Fernandez-Jimenez, N.; Cros, M.-P.; Ecsedi, S.; Cuenin, C.; Bouaoun, L.; et al. Unique DNA methylation signature in HPV-positive head and neck squamous cell carcinomas. Genome Med. 2017, 9, 33. [Google Scholar] [CrossRef]
- Lim, Y.; Wan, Y.; Vagenas, D.; Ovchinnikov, D.A.; Perry, C.F.L.; Davis, M.J.; Punyadeera, C. Salivary DNA methylation panel to diagnose HPV-positive and HPV-negative head and neck cancers. BMC Cancer 2016, 16, 749. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Furlan, C.; Lupato, V.; Polesel, J.; Fratta, E. Novel insights into epigenetic drivers of oropharyngeal squamous cell carcinoma: Role of HPV and lifestyle factors. Clin. Epigenetics 2017, 9, 124. [Google Scholar] [CrossRef]
- Khanal, S.; Shumway, B.S.; Zahin, M.; Redman, R.A.; Strickley, J.D.; Trainor, P.J.; Rai, S.N.; Ghim, S.-J.; Jenson, A.B.; Joh, J. Viral DNA integration and methylation of human papillomavirus type 16 in high-grade oral epithelial dysplasia and head and neck squamous cell carcinoma. Oncotarget 2018, 9, 30419–30433. [Google Scholar] [CrossRef]
- Koneva, L.A.; Zhang, Y.; Virani, S.; Hall, P.B.; McHugh, J.B.; Chepeha, D.B.; Wolf, G.T.; Carey, T.E.; Rozek, L.S.; Sartor, M.A. HPV Integration in HNSCC Correlates with Survival Outcomes, Immune Response Signatures, and Candidate Drivers. Mol. Cancer Res. MCR 2018, 16, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Liu, J.; Zhang, L.; Dong, S.; Zhang, J.; Liu, Y.; Zhou, H.; Dong, W. Identification of Potential Biomarkers and Survival Analysis for Head and Neck Squamous Cell Carcinoma Using Bioinformatics Strategy: A Study Based on TCGA and GEO Datasets. BioMed Res. Int. 2019, 2019, e7376034. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Yang, X.; Si, H.; Saleh, A.D.; Xiao, W.; Coupar, J.; Gollin, S.M.; Ferris, R.L.; Issaeva, N.; Yarbrough, W.G.; et al. Genomic and Transcriptomic Characterization Links Cell Lines with Aggressive Head and Neck Cancers. Cell Rep. 2018, 25, 1332–1345.e5. [Google Scholar] [CrossRef] [PubMed]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 2016, 375, 1109–1112. [Google Scholar] [CrossRef]
- Lee, H.; Palm, J.; Grimes, S.M.; Ji, H.P. The Cancer Genome Atlas Clinical Explorer: A web and mobile interface for identifying clinical–genomic driver associations. Genome Med. 2015, 7, 112. [Google Scholar] [CrossRef]
- Diamandis, E.P.; Yousef, G.M. Human tissue kallikreins: A family of new cancer biomarkers. Clin. Chem. 2002, 48, 1198–1205. [Google Scholar] [CrossRef]
- Fuhrman-Luck, R.A.; Loessner, D.; Clements, J.A. Kallikrein-Related Peptidases in Prostate Cancer: From Molecular Function to Clinical Application. EJIFCC 2014, 25, 269–281. [Google Scholar]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef]
- Mi, H.; Dong, Q.; Muruganujan, A.; Gaudet, P.; Lewis, S.; Thomas, P.D. PANTHER version 7: Improved phylogenetic trees, orthologs and collaboration with the Gene Ontology Consortium. Nucleic Acids Res. 2010, 38, D204–D210. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.A.; Addison, C.L. β1 integrin: An emerging player in the modulation of tumorigenesis and response to therapy. Cell Adhes. Migr. 2012, 6, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Husain, N.; Neyaz, A. Human papillomavirus associated head and neck squamous cell carcinoma: Controversies and new concepts. J. Oral Biol. Craniofacial Res. 2017, 7, 198–205. [Google Scholar] [CrossRef]
- Dok, R.; Nuyts, S. HPV Positive Head and Neck Cancers: Molecular Pathogenesis and Evolving Treatment Strategies. Cancers 2016, 8, 41. [Google Scholar] [CrossRef]
- Vigneswaran, N.; Williams, M.D. Epidemiologic trends in head and neck cancer and aids in diagnosis. Oral Maxillofac. Surg. Clin. N. Am. 2014, 26, 123–141. [Google Scholar] [CrossRef]
- Özdemir, B.C.; Dotto, G.-P. Racial Differences in Cancer Susceptibility and Survival: More Than the Color of the Skin? Trends Cancer 2017, 3, 181–197. [Google Scholar] [CrossRef]
- Joshi, P.; Dutta, S.; Chaturvedi, P.; Nair, S. Head and Neck Cancers in Developing Countries. Rambam Maimonides Med. J. 2014, 5, e0009. [Google Scholar] [CrossRef]
- Gourin, C.G.; Podolsky, R.H. Racial disparities in patients with head and neck squamous cell carcinoma. Laryngoscope 2006, 116, 1093–1106. [Google Scholar] [CrossRef]
- Marur, S.; D’Souza, G.; Westra, W.H.; Forastiere, A.A. HPV-associated head and neck cancer: A virus-related cancer epidemic. Lancet Oncol. 2010, 11, 781–789. [Google Scholar] [CrossRef]
- Mahal, B.A.; Catalano, P.J.; Haddad, R.I.; Hanna, G.J.; Kass, J.I.; Schoenfeld, J.D.; Tishler, R.B.; Margalit, D.N. Incidence and Demographic Burden of HPV-Associated Oropharyngeal Head and Neck Cancers in the United States. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1660–1667. [Google Scholar] [CrossRef]
- Offenberg, H.H.; Schalk, J.A.; Meuwissen, R.L.; van Aalderen, M.; Kester, H.A.; Dietrich, A.J.; Heyting, C. SCP2: A major protein component of the axial elements of synaptonemal complexes of the rat. Nucleic Acids Res. 1998, 26, 2572–2579. [Google Scholar] [CrossRef]
- Pannone, G.; Santoro, A.; Papagerakis, S.; Lo Muzio, L.; De Rosa, G.; Bufo, P. The role of human papillomavirus in the pathogenesis of head & neck squamous cell carcinoma: An overview. Infect. Agent. Cancer 2011, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Masterson, L.; Sorgeloos, F.; Winder, D.; Lechner, M.; Marker, A.; Malhotra, S.; Sudhoff, H.; Jani, P.; Goon, P.; Sterling, J. Deregulation of SYCP2 predicts early stage human papillomavirus-positive oropharyngeal carcinoma: A prospective whole transcriptome analysis. Cancer Sci. 2015, 106, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Pyeon, D.; Newton, M.A.; Lambert, P.F.; den Boon, J.A.; Sengupta, S.; Marsit, C.J.; Woodworth, C.D.; Connor, J.P.; Haugen, T.H.; Smith, E.M.; et al. Fundamental differences in cell cycle deregulation in human papillomavirus-positive and human papillomavirus-negative head/neck and cervical cancers. Cancer Res. 2007, 67, 4605–4619. [Google Scholar] [CrossRef]
- Martinez, I.; Wang, J.; Hobson, K.F.; Ferris, R.L.; Khan, S.A. Identification of differentially expressed genes in HPV-positive and HPV-negative oropharyngeal squamous cell carcinomas. Eur. J. Cancer 2007, 43, 415–432. [Google Scholar] [CrossRef]
- Slebos, R.J.C.; Yi, Y.; Ely, K.; Carter, J.; Evjen, A.; Zhang, X.; Shyr, Y.; Murphy, B.M.; Cmelak, A.J.; Burkey, B.B.; et al. Gene expression differences associated with human papillomavirus status in head and neck squamous cell carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 701–709. [Google Scholar] [CrossRef]
- Zhou, H.; Grubisic, I.; Zheng, K.; He, Y.; Wang, P.J.; Kaplan, T.; Tjian, R. Taf7l cooperates with Trf2 to regulate spermiogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 16886–16891. [Google Scholar] [CrossRef]
- Mobasheri, M.B.; Shirkoohi, R.; Modarressi, M.H. Cancer/Testis OIP5 and TAF7L Genes are Up-Regulated in Breast Cancer. Asian Pac. J. Cancer Prev. APJCP 2015, 16, 4623–4628. [Google Scholar] [CrossRef]
- Stoimenov, I.; Helleday, T. PCNA on the crossroad of cancer. Biochem. Soc. Trans. 2009, 37, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Fey, S.J.; Bellatin, J.; Larsen, P.M.; Celis, J.E. Identification of a nuclear polypeptide (“cyclin”) whose relative proportion is sensitive to changes in the rate of cell proliferation and to transformation. Prog. Clin. Biol. Res. 1982, 85, 235–248. [Google Scholar]
- Hoelz, D.J.; Arnold, R.J.; Dobrolecki, L.E.; Abdel-Aziz, W.; Loehrer, A.P.; Novotny, M.V.; Schnaper, L.; Hickey, R.J.; Malkas, L.H. The discovery of labile methyl esters on proliferating cell nuclear antigen by MS/MS. Proteomics 2006, 6, 4808–4816. [Google Scholar] [CrossRef]
- Montgomery, R.I.; Warner, M.S.; Lum, B.J.; Spear, P.G. Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell 1996, 87, 427–436. [Google Scholar] [CrossRef]
- Edilova, M.I.; Abdul-Sater, A.A.; Watts, T.H. TRAF1 Signaling in Human Health and Disease. Front. Immunol. 2018, 9, 2969. [Google Scholar] [CrossRef] [PubMed]
- Marsters, S.A.; Ayres, T.M.; Skubatch, M.; Gray, C.L.; Rothe, M.; Ashkenazi, A. Herpesvirus entry mediator, a member of the tumor necrosis factor receptor (TNFR) family, interacts with members of the TNFR-associated factor family and activates the transcription factors NF-kappaB and AP-1. J. Biol. Chem. 1997, 272, 14029–14032. [Google Scholar] [CrossRef]
- Rothe, M.; Pan, M.G.; Henzel, W.J.; Ayres, T.M.; Goeddel, D.V. The TNFR2-TRAF signaling complex contains two novel proteins related to baculoviral inhibitor of apoptosis proteins. Cell 1995, 83, 1243–1252. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 18 September 2021).
- Evans, M.R.; James, C.D.; Loughran, O.; Nulton, T.J.; Wang, X.; Bristol, M.L.; Windle, B.; Morgan, I.M. An oral keratinocyte life cycle model identifies novel host genome regulation by human papillomavirus 16 relevant to HPV positive head and neck cancer. Oncotarget 2017, 8, 81892–81909. [Google Scholar] [CrossRef]
- An, X.; Hao, Y.; Meneses, P.I. Host cell transcriptome modification upon exogenous HPV16 L2 protein expression. Oncotarget 2017, 8, 90730–90747. [Google Scholar] [CrossRef]
- Certo, M.; Del Gaizo Moore, V.; Nishino, M.; Wei, G.; Korsmeyer, S.; Armstrong, S.A.; Letai, A. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 2006, 9, 351–365. [Google Scholar] [CrossRef]
- Akl, H.; Vervloessem, T.; Kiviluoto, S.; Bittremieux, M.; Parys, J.B.; De Smedt, H.; Bultynck, G. A dual role for the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus endoplasmic reticulum. Biochim. Biophys. Acta 2014, 1843, 2240–2252. [Google Scholar] [CrossRef] [PubMed]
- Letai, A.G. Diagnosing and exploiting cancer’s addiction to blocks in apoptosis. Nat. Rev. Cancer 2008, 8, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, R.; Wilander, E.; Oberg, K. Expression and prognostic significance of Bcl-2 in ovarian tumours. Br. J. Cancer 1995, 72, 1324–1329. [Google Scholar] [CrossRef] [PubMed]
- Lamers, F.; Schild, L.; den Hartog, I.J.M.; Ebus, M.E.; Westerhout, E.M.; Ora, I.; Koster, J.; Versteeg, R.; Caron, H.N.; Molenaar, J.J. Targeted BCL2 inhibition effectively inhibits neuroblastoma tumour growth. Eur. J. Cancer 2012, 48, 3093–3103. [Google Scholar] [CrossRef]
- Zhao, D.; Ding, X.; Peng, J.; Zheng, Y.; Zhang, S. Prognostic significance of bcl-2 and p53 expression in colorectal carcinoma. J. Zhejiang Univ. Sci. B 2005, 6, 1163–1169. [Google Scholar] [CrossRef]
- Pena, J.C.; Thompson, C.B.; Recant, W.; Vokes, E.E.; Rudin, C.M. Bcl-xL and Bcl-2 expression in squamous cell carcinoma of the head and neck. Cancer 1999, 85, 164–170. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; de Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130. [Google Scholar] [CrossRef]
- Munger, K.; Gwin, T.K.; McLaughlin-Drubin, M.E. p16 in HPV-associated cancers. Oncotarget 2013, 4, 1864–1865. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Use R! Springer: New York, NY, USA, 2009; ISBN 978-0-387-98141-3. [Google Scholar]
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes. 2014. [Google Scholar]
- Keitt, T. colorRamps: Builds Color Tables. 2012. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots: Various R Programming Tools for Plotting Data. 2020. [Google Scholar]
- Team, R. RStudio: Integrated Development for R; Team R: Boston, MA, USA, 2018. [Google Scholar]
- R Core Team. European Environment Agency. 2019. Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 18 September 2021).
- Silva, T.C.; Colaprico, A.; Olsen, C.; D’Angelo, F.; Bontempi, G.; Ceccarelli, M.; Noushmehr, H. TCGA Workflow: Analyze cancer genomics and epigenomics data using Bioconductor packages. F1000Research 2016, 5, 1542. [Google Scholar] [CrossRef]
- Mounir, M.; Lucchetta, M.; Silva, T.C.; Olsen, C.; Bontempi, G.; Chen, X.; Noushmehr, H.; Colaprico, A.; Papaleo, E. New functionalities in the TCGAbiolinks package for the study and integration of cancer data from GDC and GTEx. PLoS Comput. Biol. 2019, 15, e1006701. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed]
- Dragulescu, A.; Arendt, C. xlsx: Read, Write, Format Excel 2007 and Excel 97/2000/XP/2003 Files. 2020. Available online: https://github.com/colearendt/xlsx (accessed on 30 August 2022).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed]
- Phipson, B.; Trigos, A.; Ritchie, M.; Su, S.; Doyle, M.; Dashnow, H.; Law, C. RNA-seq Analysis in R; Differential Expression Analysis 2016. Available online: https://combine-australia.github.io/RNAseq-R/06-rnaseq-day1.html (accessed on 30 August 2019).
- Morgan, M.; Obenchain, V.; Hester, J.; Pagès, H. SummarizedExperiment: SummarizedExperiment container, Bioconductor version: Release (3.13). 2021.
- Gentleman, R.; Carey, V.J.; Huber, W.; Hahne, F. Genefilter: Genefilter: Methods for Filtering Genes from High-Throughput Experiments, Bioconductor Version: Release (3.13). 2021.
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Ramos, M.; Schiffer, L.; Re, A.; Azhar, R.; Basunia, A.; Rodriguez, C.; Chan, T.; Chapman, P.; Davis, S.R.; Gomez-Cabrero, D.; et al. Software for the Integration of Multiomics Experiments in Bioconductor. Cancer Res. 2017, 77, e39–e42. [Google Scholar] [CrossRef]
- Blighe, K.; Rana, S.; Turkes, E.; Ostendorf, B.; Grioni, A.; Lewis, M. EnhancedVolcano: Publication-ready volcano plots with enhanced colouring and labeling, Bioconductor Version: Release (3.13). 2021.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Representative KEGG Pathways | Mapped Differentially Expressed Genes in HPVP vs. HPVN |
|---|---|
| ECM-receptor interaction | CD36, ITGA6, ITGA5, ITGB3 |
| Focal adhesion | BCL2, EGF, EGFR, ERBB2, IGF1, VEGFC |
| Viral protein interaction with cytokine and cytokine receptor | IL18, IL18RAP, IL19, LTA, TNFRSF14, IL6 |
| Proteoglycans in cancer | TP53, EGFR, ERBB2, IGF1 |
| Transcriptional misregulation in cancer | TP53, BCL2A1, CCNA1, CDKN2C, CSF2, GADD45G, ID2, IL6, MYCN, MEF2C, TLX3, TRAF1 |
| Human papillomavirus infection | CCNE2, CDK6, E2F1, PDGFRB, EGF, EGFR, TP53 |
| TNF signaling pathway | BIRC3, CCL20, CSF2, IL15, IL6, TRAF1, TRAF2, VEGFC |
| Cell cycle | BIRC3, BCL2, BCL2A1, NGF, TRAF1, TRAF2, PCNA, TP53, GADD45G |
| TGF-beta signaling pathway | AMH, DCN, ID2, IFNG, INHBA, INHBB, LTBP1, NOG, THBS1 |
| Apoptosis | BCL2, BCL2A1, BIRC3, GADD45G, NGF, TP53, TRAF1, TRAF2 |
| Category | Regulation Level | q-Value | Database |
|---|---|---|---|
| Transcription Factors | |||
| NFkB | upregulated | 3.29 × 10−2 | TRANSFAC and JASPAR PWMs |
| SP1 human | downregulated | 2.37 × 10−6 | TRRUST Transcription Factors 2019 |
| Pathways | |||
| Retinoblastoma gene in cancer WP2446 | upregulated | 4.58 × 10−6 | WikiPathways~2019 Human |
| DNA strand elongation Homo Sapiens R-HAS-69190 | upregulated | 2.02 × 10−4 | Reactome~2016 |
| Beta1 integrin cell surface interactions Homo Sapiens | downregulated | 1.25 × 10−15 | NCI-Nature 2016 |
| ITGB1 | downregulated | 5.64 × 10−5 | PPI Hub Proteins |
| Integrin signaling pathway Homo Sapiens | downregulated | 2.02 × 10−6 | PANTHER 2016 |
| Ontologies | |||
| G1/S transition in mitotic cell cycle (GO:0000082) | upregulated | 3.04 × 10−2 | GO Biological Processes 2018 |
| T cell receptor complex (GO:0042101) | upregulated | 7.57 × 10−5 | GO Cellular Component 2018 |
| Collagen binding (GO:0005518) | downregulated | 1.70 × 10−8 | GO Molecular Function 2018 |
| Upregulated Processes | Downregulated Processes |
|---|---|
| Category: Biological Processes | Category: Biological Processes |
| Cellular Process was top hit (GO:0009987) 311/913 genes | Cellular Process was top hit (GO:0009987) 344/941 genes |
| subcategory | subcategory |
| cell cycle (GO:0007049) 44/311 genes | cellular response to stimulus (GO:0051716) 110/344 genes |
| ID | Pathway | Gene Ratio | q-Value |
|---|---|---|---|
| hsa04512 | ECM-receptor interaction | 32/714 | 3.47 × 10−10 |
| hsa04060 | Cytokine-cytokine receptor interaction | 61/714 | 3.38 × 10−8 |
| hsa04640 | Hematopoetic cell lineage | 30/714 | 5.86 × 10−8 |
| hsa04974 | Protein digestion and absorption | 29/714 | 1.19 × 10−7 |
| hsa04510 | Focal adhesion | 44/714 | 6.22 × 10−7 |
| hsa05410 | Hypertrophic cardiomyopathy (HCM) | 25/714 | 8.20 × 10−6 |
| hsa04151 | PI3K-Akt signaling pathway | 61/714 | 1.42 × 10−5 |
| hsa04061 | Viral protein interaction with cytokine and cytokine receptor | 26/714 | 1.47 × 10−5 |
| hsa05150 | Staphylococcus aureus infection | 24/714 | 6.90 × 10−5 |
| Probe ID | Gene Symbol | Adjusted p-Value | Status in HPVP |
|---|---|---|---|
| cg11456145 | CDC42EP5 | 8.40 × 10−11 | Hypomethylated |
| cg07915849 | ABCA17P;ABCA3 | 6.97 × 10−10 | Hypomethylated |
| cg10504436 | DERL3 | 6.97 × 10−10 | Hypomethylated |
| cg12181372 | SYCP2 | 1.77 × 10−9 | Hypomethylated |
| cg22220310 | SDF4;B3GALT6 | 2.08 × 10−9 | Hypomethylated |
| cg07907859 | FAM133A;NAP1L3 | 2.90 × 10−9 | Hypermethylated |
| cg00757182 | ZNF773 | 4.81 × 10−9 | Hypermethylated |
| cg12387713 | MSX2 | 5.08 × 10−9 | Hypermethylated |
| cg13458645 | PITX2 | 5.08 × 10−9 | Hypermethylated |
| cg11876013 | SCHIP1 | 5.29 × 10−9 | Hypermethylated |
| Gene Symbol | Status in HPVP | Gene Name |
|---|---|---|
| TAF7L | Hypomethylated | TATA-Box Binding Protein Associated Factor 7 Like |
| SYCP2 | Hypomethylated | Synaptonemal Complex Protein 2 |
| LOC285954;INHBA | Hypermethylated | Inhibin Subunit Beta A |
| SULF1 | Hypermethylated | Sulfatase 1 |
| CCNA1 | Hypermethylated | Cyclin A1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinić, S.; Rich, A.; Anayannis, N.V.; Cabarcas-Petroski, S.; Schramm, L.; Meneses, P.I. Gene Expression and DNA Methylation in Human Papillomavirus Positive and Negative Head and Neck Squamous Cell Carcinomas. Int. J. Mol. Sci. 2022, 23, 10967. https://doi.org/10.3390/ijms231810967
Hinić S, Rich A, Anayannis NV, Cabarcas-Petroski S, Schramm L, Meneses PI. Gene Expression and DNA Methylation in Human Papillomavirus Positive and Negative Head and Neck Squamous Cell Carcinomas. International Journal of Molecular Sciences. 2022; 23(18):10967. https://doi.org/10.3390/ijms231810967
Chicago/Turabian StyleHinić, Snežana, April Rich, Nicole V. Anayannis, Stephanie Cabarcas-Petroski, Laura Schramm, and Patricio I. Meneses. 2022. "Gene Expression and DNA Methylation in Human Papillomavirus Positive and Negative Head and Neck Squamous Cell Carcinomas" International Journal of Molecular Sciences 23, no. 18: 10967. https://doi.org/10.3390/ijms231810967
APA StyleHinić, S., Rich, A., Anayannis, N. V., Cabarcas-Petroski, S., Schramm, L., & Meneses, P. I. (2022). Gene Expression and DNA Methylation in Human Papillomavirus Positive and Negative Head and Neck Squamous Cell Carcinomas. International Journal of Molecular Sciences, 23(18), 10967. https://doi.org/10.3390/ijms231810967