
Connexin Lateralization Contributes to Male Susceptibility to Atrial Fibrillation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
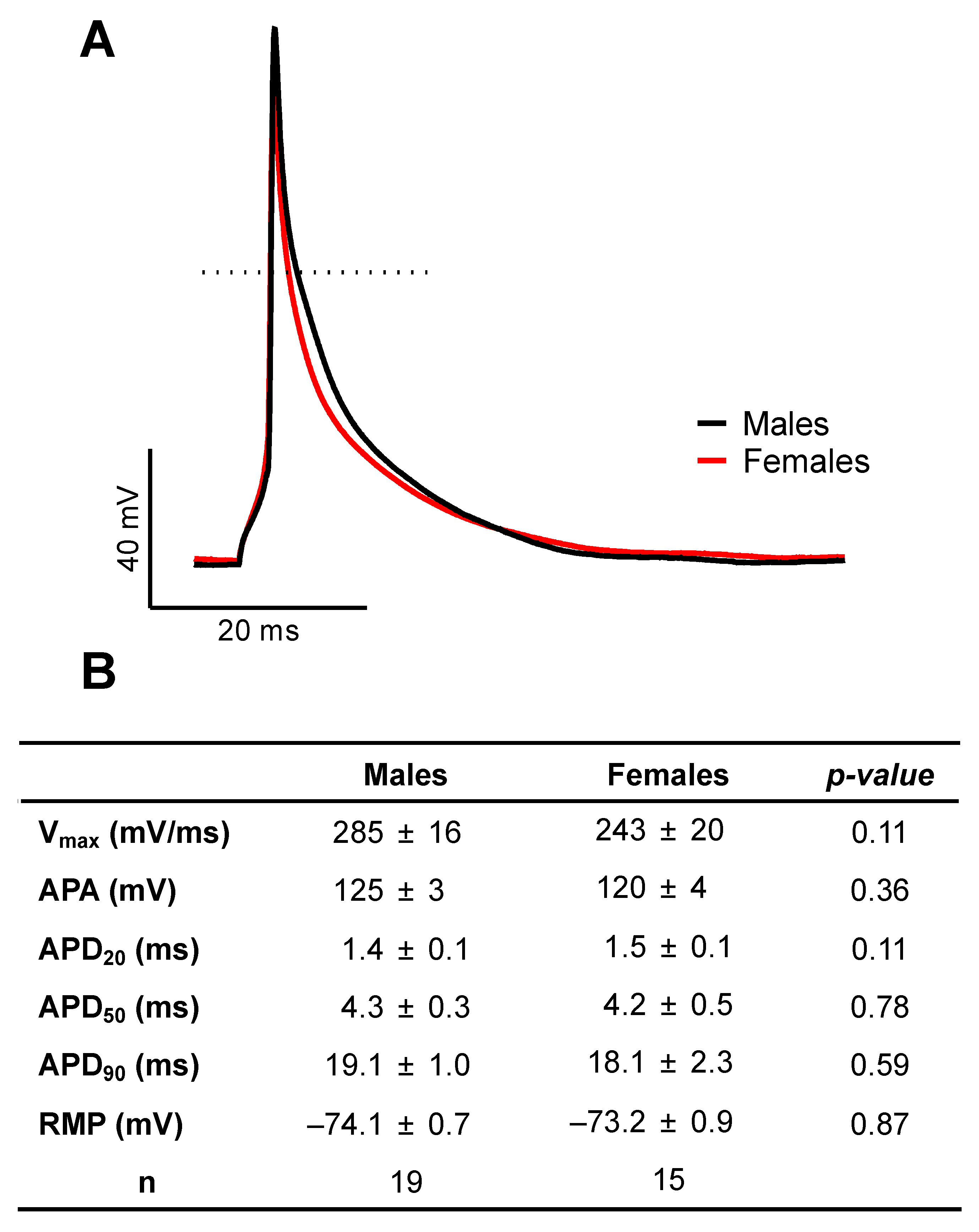
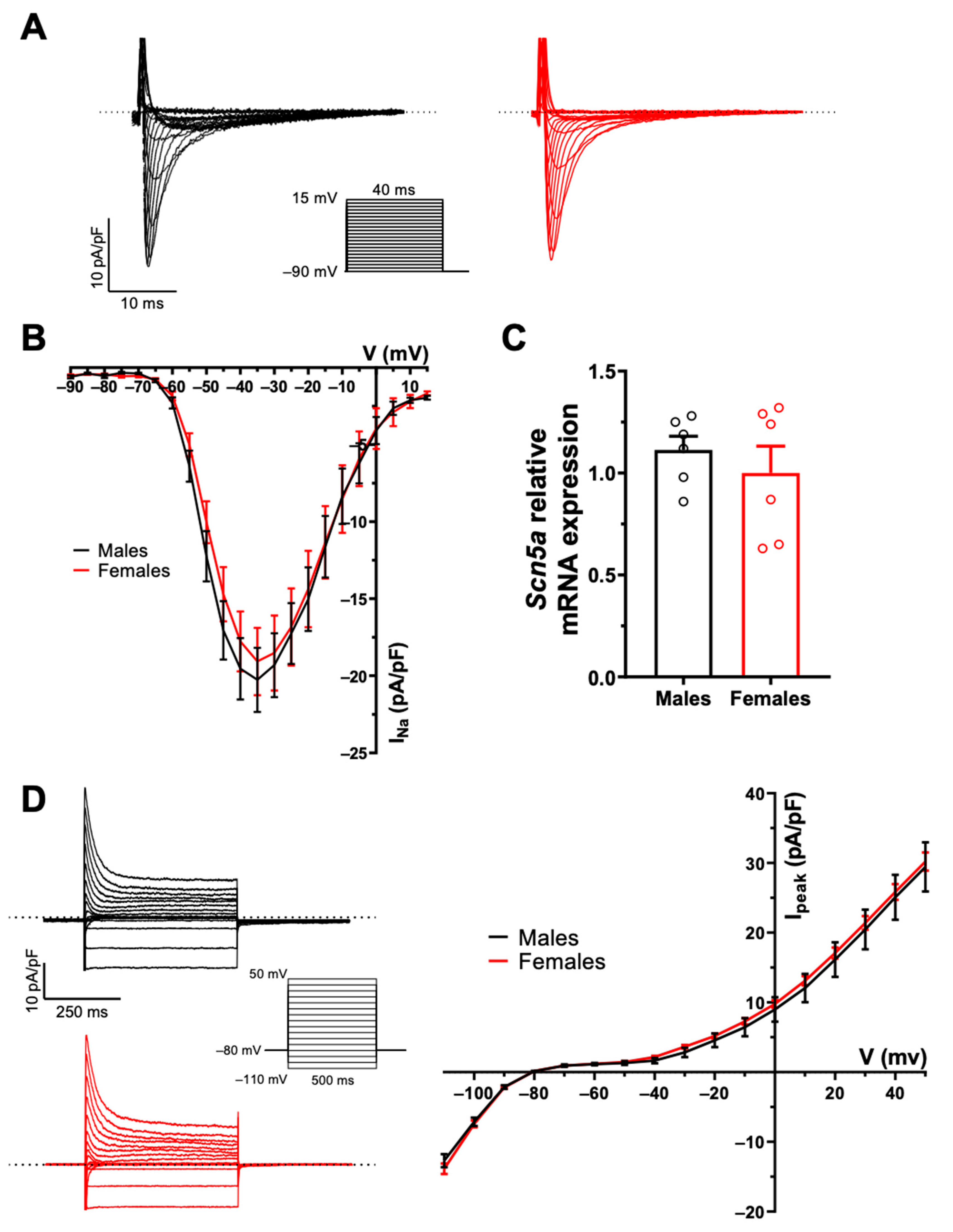
2.1. There Are No Sex Differences in AP Configuration and Ionic Currents
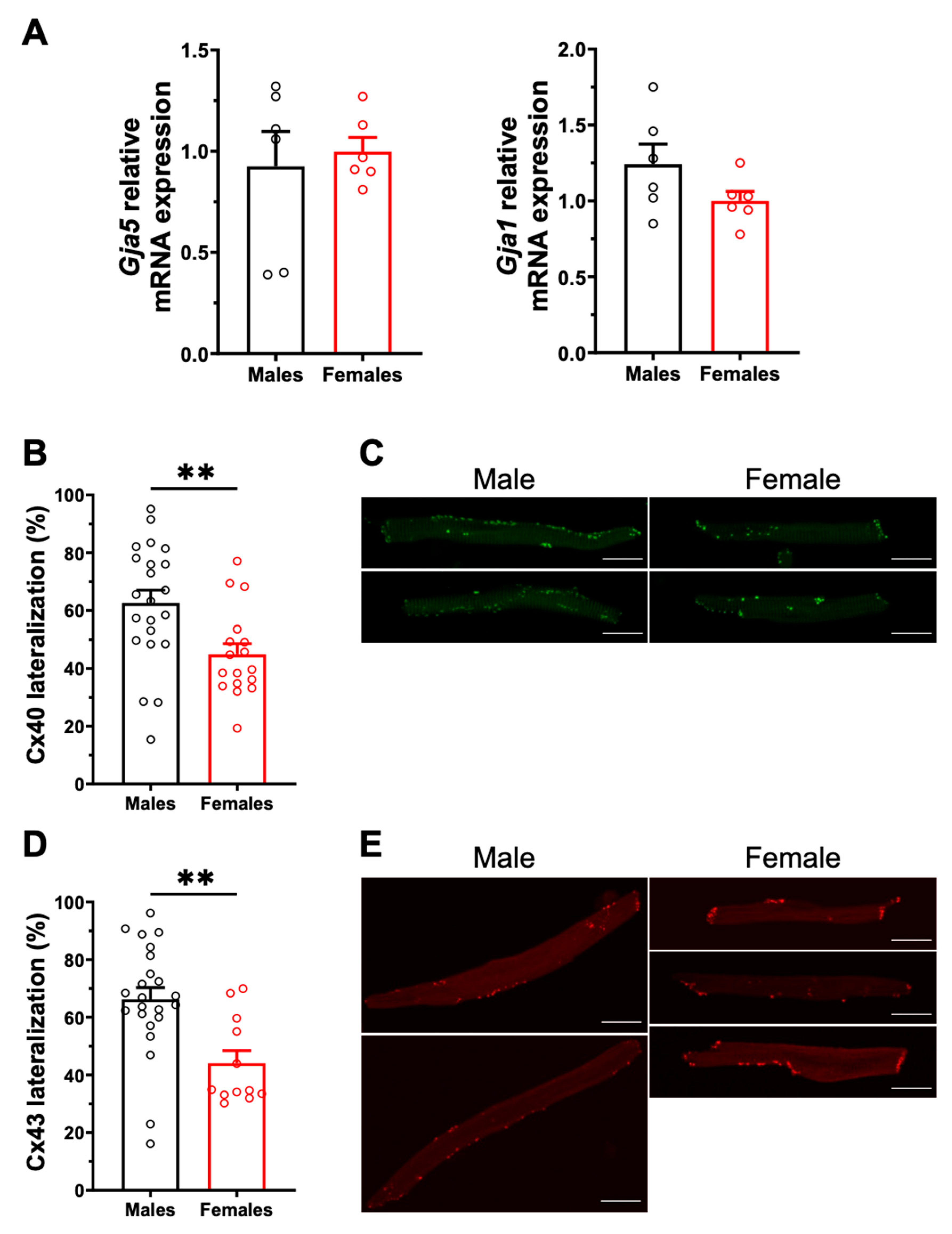
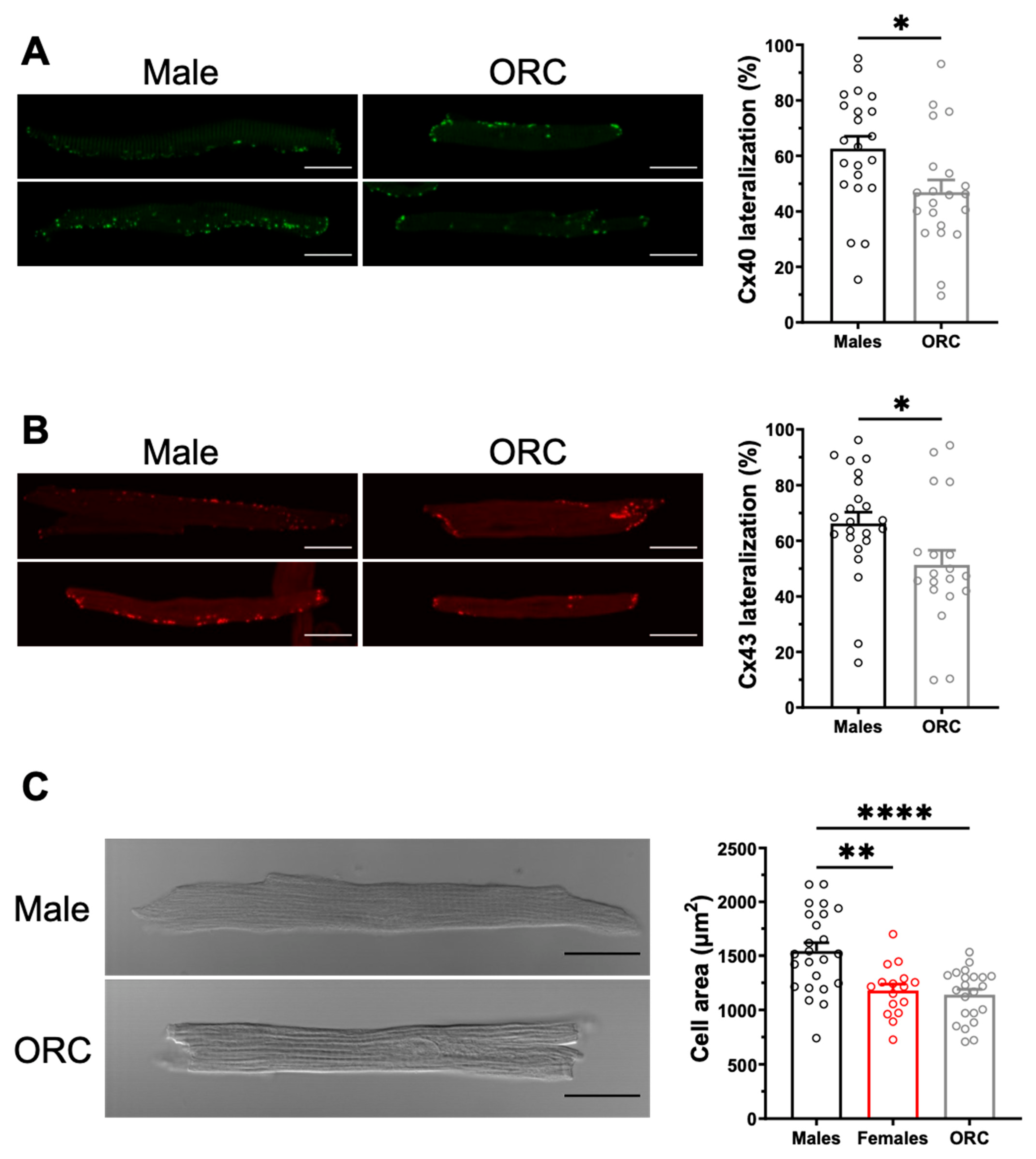
2.2. Sex Differences in Connexin Lateralization
2.3. Potential AF Anatomical Substrates
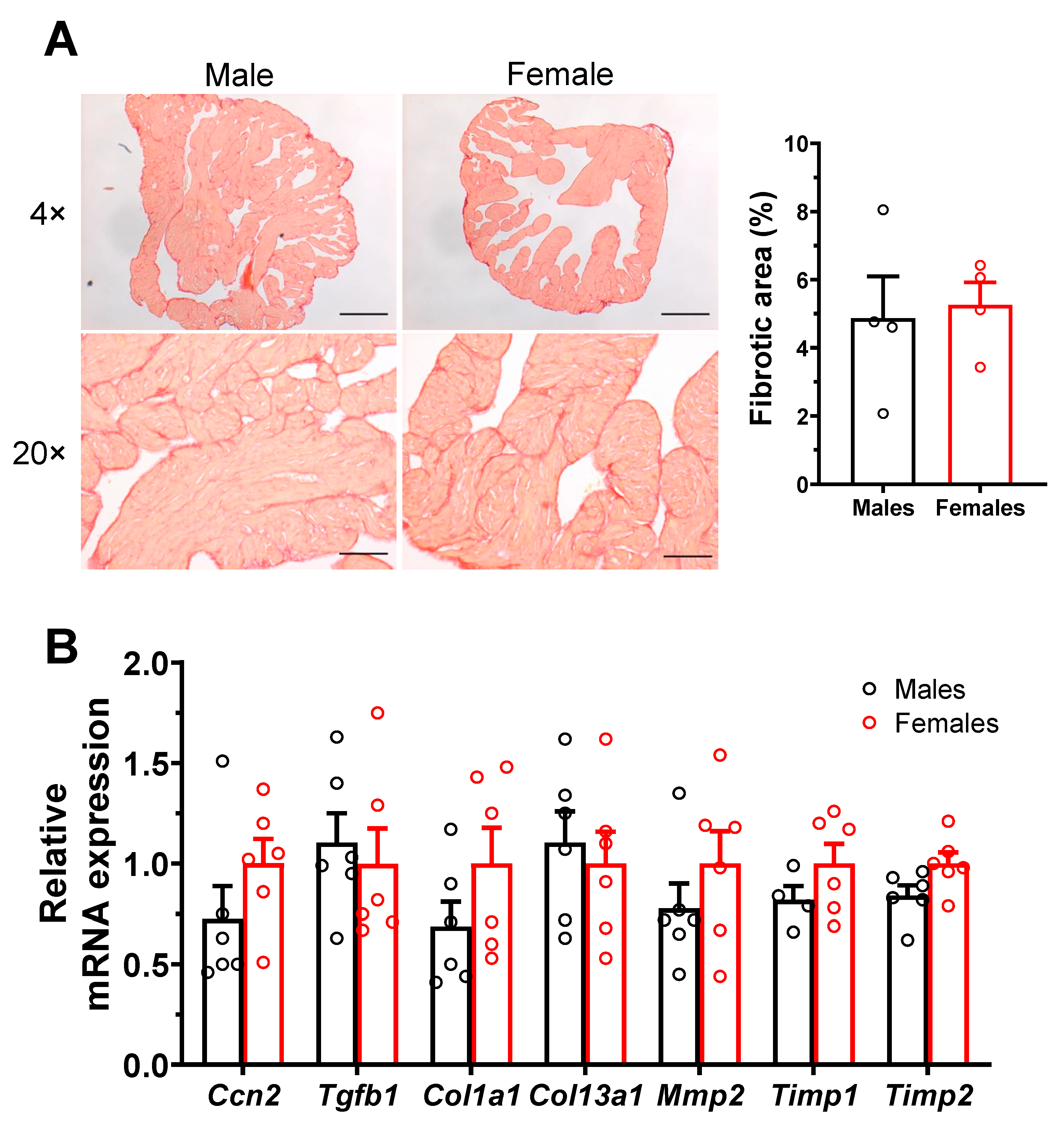
2.3.1. No Sex Differences in Atrial Interstitial Fibrosis and Fibrosis Markers
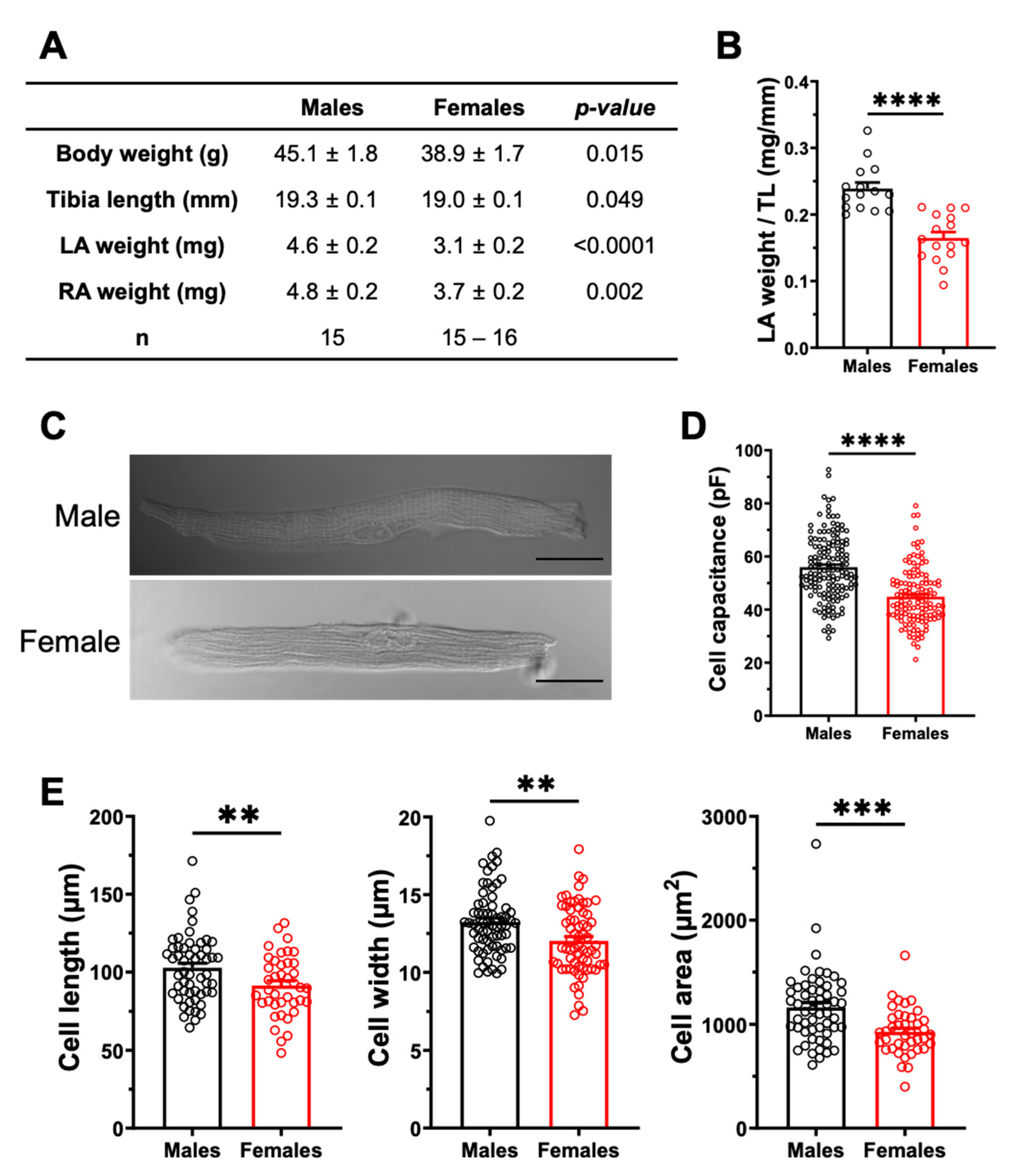
2.3.2. Sex Differences in Atrial Structural Properties
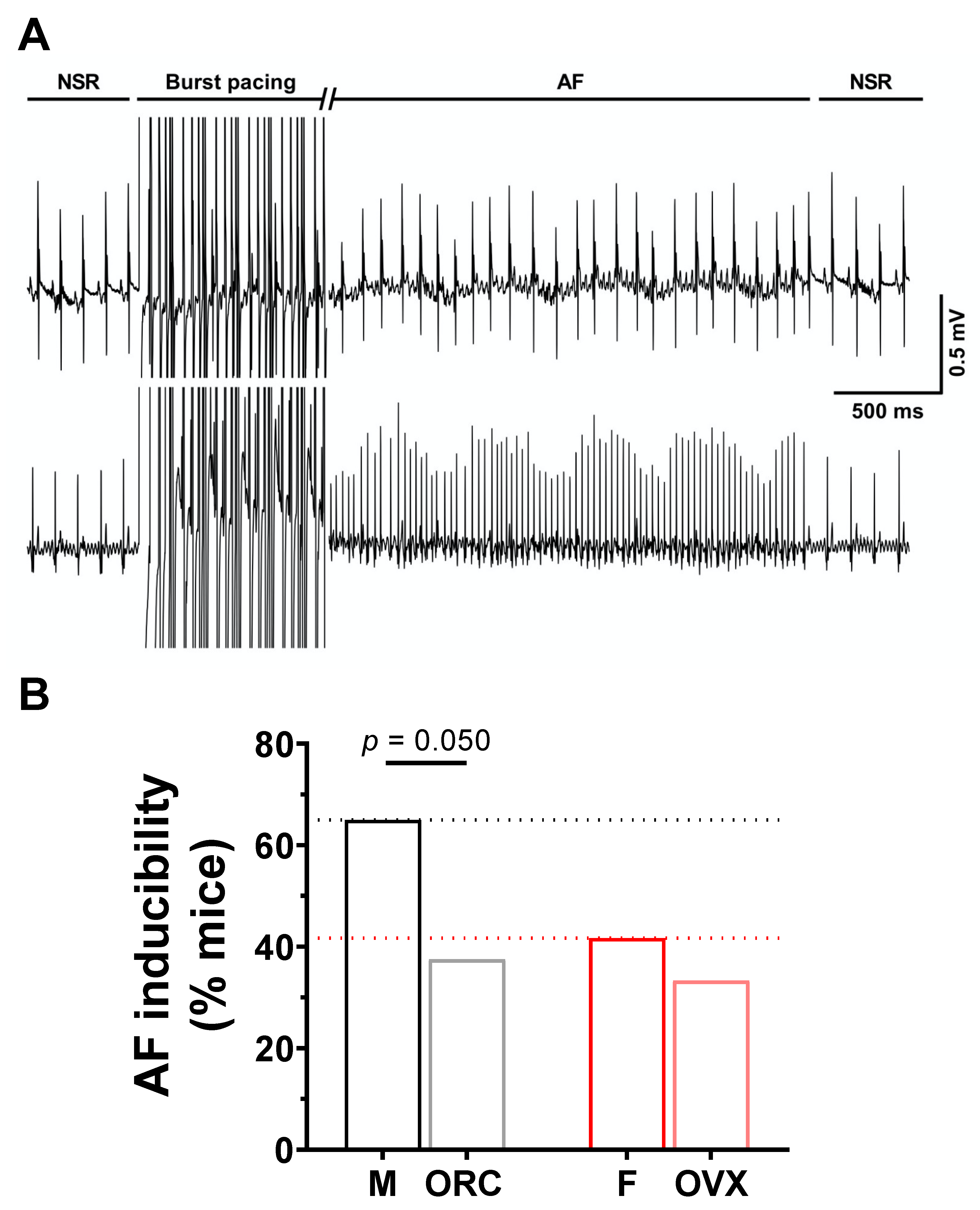
2.4. Influence of Gonadectomy on AF Susceptibility
3. Discussion
4. Methods
4.1. Animals
4.2. Gonadectomies
4.3. Electrophysiological Programmed Stimulations Studies (EPS)
4.4. Isolation of Mouse Atrial Myocytes
4.5. Cellular Electrophysiology
4.5.1. Action Potentials
4.5.2. Potassium Currents
4.5.3. Sodium Currents
4.6. Immunofluorescence
4.7. Picrosirius Red Staining
4.8. Quantitative Polymerase Chain Reaction (qPCR)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Staerk, L.; Sherer, J.A.; Ko, D.; Benjamin, E.J.; Helm, R.H. Atrial Fibrillation: Epidemiology, Pathophysiology, and Clinical Outcomes. Circ. Res. 2017, 120, 1501–1517. [Google Scholar] [CrossRef]
- Nattel, S. New Ideas about Atrial Fibrillation 50 Years On. Nature 2002, 415, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Wolf, P.A.; D’Agostino, R.B.; Silbershatz, H.; Kannel, W.B.; Levy, D. Impact of Atrial Fibrillation on the Risk of Death: The Framingham Heart Study. Circulation 1998, 98, 946–952. [Google Scholar] [CrossRef]
- Ko, D.; Rahman, F.; Schnabel, R.B.; Yin, X.; Benjamin, E.J.; Christophersen, I.E. Atrial Fibrillation in Women: Epidemiology, Pathophysiology, Presentation, and Prognosis. Nat. Rev. Cardiol. 2016, 13, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.-H.; McAnulty, J.H.; Zheng, Z.-J.; et al. Worldwide Epidemiology of Atrial Fibrillation: A Global Burden of Disease 2010 Study. Circulation 2014, 129, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Odening, K.E.; Deiß, S.; Dilling-Boer, D.; Didenko, M.; Eriksson, U.; Nedios, S.; Ng, F.S.; Roca Luque, I.; Sanchez Borque, P.; Vernooy, K.; et al. Mechanisms of Sex Differences in Atrial Fibrillation: Role of Hormones and Differences in Electrophysiology, Structure, Function, and Remodelling. EP Eur. 2019, 21, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Tadros, R.; Ton, A.-T.; Fiset, C.; Nattel, S. Sex Differences in Cardiac Electrophysiology and Clinical Arrhythmias: Epidemiology, Therapeutics, and Mechanisms. Can. J. Cardiol. 2014, 30, 783–792. [Google Scholar] [CrossRef]
- Westerman, S.; Wenger, N. Gender Differences in Atrial Fibrillation: A Review of Epidemiology, Management, and Outcomes. CCR 2019, 15, 136–144. [Google Scholar] [CrossRef]
- Linde, C.; Bongiorni, M.G.; Birgersdotter-Green, U.; Curtis, A.B.; Deisenhofer, I.; Furokawa, T.; Gillis, A.M.; Haugaa, K.H.; Lip, G.Y.H.; Van Gelder, I.; et al. Sex Differences in Cardiac Arrhythmia: A Consensus Document of the European Heart Rhythm Association, Endorsed by the Heart Rhythm Society and Asia Pacific Heart Rhythm Society. EP Eur. 2018, 20, 1565–1565ao. [Google Scholar] [CrossRef]
- Friberg, L.; Bergfeldt, L. Atrial Fibrillation Prevalence Revisited. J. Intern. Med. 2013, 274, 461–468. [Google Scholar] [CrossRef]
- Akoum, N.; Mahnkopf, C.; Kholmovski, E.G.; Brachmann, J.; Marrouche, N.F. Age and Sex Differences in Atrial Fibrosis among Patients with Atrial Fibrillation. EP Eur. 2018, 20, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Michelena, H.I.; Powell, B.D.; Brady, P.A.; Friedman, P.A.; Ezekowitz, M.D. Gender in Atrial Fibrillation: Ten Years Later. Gend. Med. 2010, 7, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Dagres, N.; Nieuwlaat, R.; Vardas, P.E.; Andresen, D.; Lévy, S.; Cobbe, S.; Kremastinos, D.T.; Breithardt, G.; Cokkinos, D.V.; Crijns, H.J.G.M. Gender-Related Differences in Presentation, Treatment, and Outcome of Patients With Atrial Fibrillation in Europe. J. Am. Coll. Cardiol. 2007, 49, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Ehdaie, A.; Cingolani, E.; Shehata, M.; Wang, X.; Curtis, A.B.; Chugh, S.S. Sex Differences in Cardiac Arrhythmias: Clinical and Research Implications. Circ. Arrhythm. Electrophysiol. 2018, 11, e005680. [Google Scholar] [CrossRef]
- Wijesurendra, R.S.; Casadei, B. Mechanisms of Atrial Fibrillation. Heart 2019, 105, 1860–1867. [Google Scholar] [CrossRef]
- Nattel, S.; Burstein, B.; Dobrev, D. Atrial Remodeling and Atrial Fibrillation: Mechanisms and Implications. Circ. Arrhythm. Electrophysiol. 2008, 1, 62–73. [Google Scholar] [CrossRef]
- Nattel, S.; Dobrev, D. Electrophysiological and Molecular Mechanisms of Paroxysmal Atrial Fibrillation. Nat. Rev. Cardiol. 2016, 13, 575–590. [Google Scholar] [CrossRef]
- Heijman, J.; Voigt, N.; Nattel, S.; Dobrev, D. Cellular and Molecular Electrophysiology of Atrial Fibrillation Initiation, Maintenance, and Progression. Circ. Res. 2014, 114, 1483–1499. [Google Scholar] [CrossRef]
- Wijffels, M.C.E.F.; Kirchhof, C.J.H.J.; Dorland, R.; Allessie, M.A. Atrial Fibrillation Begets Atrial Fibrillation: A Study in Awake Chronically Instrumented Goats. Circulation 1995, 92, 1954–1968. [Google Scholar] [CrossRef]
- Duffy, H.S.; Wit, A.L. Is There a Role for Remodeled Connexins in AF? No Simple Answers. J. Mol. Cell. Cardiol. 2008, 44, 4–13. [Google Scholar] [CrossRef][Green Version]
- Kato, T.; Iwasaki, Y.; Nattel, S. Connexins and Atrial Fibrillation: Filling in the Gaps. Circulation 2012, 125, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Saffitz, J.E. Connexins, Conduction, and Atrial Fibrillation. N. Engl. J. Med. 2006, 354, 2712–2714. [Google Scholar] [CrossRef] [PubMed]
- Chaldoupi, S.-M.; Loh, P.; Hauer, R.N.W.; de Bakker, J.M.T.; van Rijen, H.V.M. The Role of Connexin40 in Atrial Fibrillation. Cardiovasc. Res. 2009, 84, 15–23. [Google Scholar] [CrossRef]
- Asatryan, B.; Yee, L.; Ben-Haim, Y.; Dobner, S.; Servatius, H.; Roten, L.; Tanner, H.; Crotti, L.; Skinner, J.R.; Remme, C.A.; et al. Sex-Related Differences in Cardiac Channelopathies: Implications for Clinical Practice. Circulation 2021, 143, 739–752. [Google Scholar] [CrossRef]
- Ozcan, C.; Curtis, A.B. Sex Differences in Arrhythmias. In Cardiac Electrophysiology: From Cell to Bedside; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1051–1059. ISBN 978-1-4557-2856-5. [Google Scholar]
- Trépanier-Boulay, V.; St-Michel, C.; Tremblay, A.; Fiset, C. Gender-Based Differences in Cardiac Repolarization in Mouse Ventricle. Circ. Res. 2001, 89, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Brouillette, J.; Trépanier-Boulay, V.; Fiset, C. Effect of Androgen Deficiency on Mouse Ventricular Repolarization. J. Physiol. 2003, 546, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Brouillette, J.; Rivard, K.; Lizotte, E.; Fiset, C. Sex and Strain Differences in Adult Mouse Cardiac Repolarization: Importance of Androgens. Cardiovasc. Res. 2005, 65, 148–157. [Google Scholar] [CrossRef]
- Bazett, H.C. An analysis of the time-relations of electrocardiograms. Ann. Noninv. Electrocard. 1997, 2, 177–194. [Google Scholar] [CrossRef]
- Abi-Gerges, N.; Philp, K.; Pollard, C.; Wakefield, I.; Hammond, T.G.; Valentin, J.-P. Sex Differences in Ventricular Repolarization: From Cardiac Electrophysiology to Torsades de Pointes. Fundam. Clin. Pharmacol. 2004, 18, 139–151. [Google Scholar] [CrossRef]
- Nerbonne, J.M.; Kass, R.S. Molecular Physiology of Cardiac Repolarization. Physiol. Rev. 2005, 85, 1205–1253. [Google Scholar] [CrossRef]
- Andrade, J.G.; Deyell, M.W.; Lee, A.Y.K.; Macle, L. Sex Differences in Atrial Fibrillation. Can. J. Cardiol. 2018, 34, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Taneja, T.; Windhagen Mahnert, B.; Passman, R.; Goldberger, J.; Kadish, A. Effects of Sex and Age on Electrocardiographic and Cardiac Electrophysiological Properties in Adults. Pacing Clin. Electrophysiol. 2001, 24, 16–21. [Google Scholar] [CrossRef]
- Liu, X.-K.; Jahangir, A.; Terzic, A.; Gersh, B.J.; Hammill, S.C.; Shen, W.-K. Age- and Sex-Related Atrial Electrophysiologic and Structural Changes. Am. J. Cardiol. 2004, 94, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Ravens, U. Sex Differences in Cardiac Electrophysiology. Can. J. Physiol. Pharmacol. 2018, 96, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Janse, M.J. Why Does Atrial Fibrillation Occur? Eur. Heart J. 1997, 18, 12–18. [Google Scholar] [CrossRef]
- Guerra, F.; Ciliberti, G.; Capucci, A. Sex Differences in Atrial Fibrillation: The Case of Testosterone. Eur. J. Prev. Cardiol. 2018, 25, 1131–1132. [Google Scholar] [CrossRef]
- Magnani, J.W.; Moser, C.B.; Murabito, J.M.; Sullivan, L.M.; Wang, N.; Ellinor, P.T.; Vasan, R.S.; Benjamin, E.J.; Coviello, A.D. Association of Sex Hormones, Aging, and Atrial Fibrillation in Men: The Framingham Heart Study. Circ: Arrhythmia Electrophysiol. 2014, 7, 307–312. [Google Scholar] [CrossRef]
- Jiangtao, L.; Dongchen, Z.; Shudong, X.; Yunpeng, S.; Lihong, W.; Liangrong, Z.; Jianhua, Z. Reduced Testosterone Levels in Males with Lone Atrial Fibrillation. Clin. Cardiol. 2009, 32, 43–46. [Google Scholar] [CrossRef]
- Rosenberg, M.A.; Shores, M.M.; Matsumoto, A.M.; Bůžková, P.; Lange, L.A.; Kronmal, R.A.; Heckbert, S.R.; Mukamal, K.J. Serum Androgens and Risk of Atrial Fibrillation in Older Men: The Cardiovascular Health Study. Clin. Cardiol. 2018, 41, 830–836. [Google Scholar] [CrossRef]
- Zeller, T.; Schnabel, R.B.; Appelbaum, S.; Ojeda, F.; Berisha, F.; Schulte-Steinberg, B.; Brueckmann, B.-E.; Kuulasmaa, K.; Jousilahti, P.; Blankenberg, S.; et al. Low Testosterone Levels Are Predictive for Incident Atrial Fibrillation and Ischaemic Stroke in Men, but Protective in Women—Results from the FINRISK Study. Eur. J. Prev. Cardiol. 2018, 25, 1133–1139. [Google Scholar] [CrossRef]
- O’Neal, W.T.; Nazarian, S.; Alonso, A.; Heckbert, S.R.; Vaccarino, V.; Soliman, E.Z. Sex Hormones and the Risk of Atrial Fibrillation: The Multi-Ethnic Study of Atherosclerosis (MESA). Endocrine 2017, 58, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Berger, D.; Folsom, A.R.; Schreiner, P.J.; Chen, L.Y.; Michos, E.D.; O’Neal, W.T.; Soliman, E.Z.; Alonso, A. Plasma Total Testosterone and Risk of Incident Atrial Fibrillation: The Atherosclerosis Risk in Communities (ARIC) Study. Maturitas 2019, 125, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.L.; Martinez, C.M.; Gallagher, E.J. Atrial Fibrillation and Anabolic Steroids. J. Emerg. Med. 1999, 17, 851–857. [Google Scholar] [CrossRef]
- Lau, D.H.; Stiles, M.K.; John, B.; Shashidhar; Young, G.D.; Sanders, P. Atrial Fibrillation and Anabolic Steroid Abuse. Int. J. Cardiol. 2007, 117, e86–e87. [Google Scholar] [CrossRef]
- Tsai, W.-C.; Lee, T.-I.; Chen, Y.-C.; Kao, Y.-H.; Lu, Y.-Y.; Lin, Y.-K.; Chen, S.-A.; Chen, Y.-J. Testosterone Replacement Increases Aged Pulmonary Vein and Left Atrium Arrhythmogenesis with Enhanced Adrenergic Activity. Int. J. Cardiol. 2014, 176, 110–118. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Li, P.-H.; Shi, L.; Su, W.-Z.; Li, D.-S.; Xue, G.-L.; Zhao, Y.; Li, C.-Z.; Li, Y.; Zhou, Y.; et al. Valsartan Reduced the Vulnerability to Atrial Fibrillation by Preventing Action Potential Prolongation and Conduction Slowing in Castrated Male Mice: SUN et al. J. Cardiovasc. Electrophysiol. 2018, 29, 1436–1443. [Google Scholar] [CrossRef]
- Tsuneda, T.; Yamashita, T.; Kato, T.; Sekiguchi, A.; Sagara, K.; Sawada, H.; Aizawa, T.; Fu, L.-T.; Fujiki, A.; Inoue, H. Deficiency of Testosterone Associates with the Substrate of Atrial Fibrillation in the Rat Model. J. Cardiovasc. Electrophysiol. 2009, 20, 1055–1060. [Google Scholar] [CrossRef]
- Rossi, R.; Grimaldi, T.; Origliani, G.; Fantini, G.; Coppi, F.; Modena, M.G. Menopause and Cardiovascular Risk. Pathophysiol. Haemost. Thromb. 2002, 32, 325–328. [Google Scholar] [CrossRef]
- Magnani, J.W.; Moser, C.B.; Murabito, J.M.; Nelson, K.P.; Fontes, J.D.; Lubitz, S.A.; Sullivan, L.M.; Ellinor, P.T.; Benjamin, E.J. Age of Natural Menopause and Atrial Fibrillation: The Framingham Heart Study. Am. Heart J. 2012, 163, 729–734. [Google Scholar] [CrossRef]
- Perez, M.V.; Wang, P.J.; Larson, J.C.; Virnig, B.A.; Cochrane, B.; Curb, J.D.; Klein, L.; Manson, J.E.; Martin, L.W.; Robinson, J.; et al. Effects of Postmenopausal Hormone Therapy on Incident Atrial Fibrillation: The Women’s Health Initiative Randomized Controlled Trials. Circ: Arrhythmia Electrophysiol. 2012, 5, 1108–1116. [Google Scholar] [CrossRef]
- Bretler, D.-M.; Hansen, P.R.; Lindhardsen, J.; Ahlehoff, O.; Andersson, C.; Jensen, T.B.; Raunsø, J.; Torp-Pedersen, C.; Gislason, G.H. Hormone Replacement Therapy and Risk of New-Onset Atrial Fibrillation after Myocardial Infarction—A Nationwide Cohort Study. PLoS ONE 2012, 7, e51580. [Google Scholar] [CrossRef]
- Tsai, W.-C.; Haung, Y.-B.; Kuo, H.-F.; Tang, W.-H.; Hsu, P.-C.; Su, H.-M.; Lin, T.-H.; Chu, C.-S.; Jhuo, S.-J.; Lee, K.-T.; et al. Hormone Replacement Therapy and Risk of Atrial Fibrillation in Taiwanese Menopause Women: A Nationwide Cohort Study. Sci. Rep. 2016, 6, 24132. [Google Scholar] [CrossRef]
- Wong, J.A.; Rexrode, K.M.; Sandhu, R.K.; Moorthy, M.V.; Conen, D.; Albert, C.M. Menopausal Age, Postmenopausal Hormone Therapy and Incident Atrial Fibrillation. Heart 2017, 103, 1954–1961. [Google Scholar] [CrossRef]
- Haïssaguerre, M.; Jaïs, P.; Shah, D.C.; Takahashi, A.; Hocini, M.; Quiniou, G.; Garrigue, S.; Le Mouroux, A.; Le Métayer, P.; Clémenty, J. Spontaneous Initiation of Atrial Fibrillation by Ectopic Beats Originating in the Pulmonary Veins. N. Engl. J. Med. 1998, 339, 659–666. [Google Scholar] [CrossRef]
- Lizotte, E.; Grandy, S.A.; Tremblay, A.; Allen, B.G.; Fiset, C. Expression, Distribution and Regulation of Sex Steroid Hormone Receptors in Mouse Heart. Cell. Physiol. Biochem. 2009, 23, 75–86. [Google Scholar] [CrossRef]
- El Gebeily, G.; El Khoury, N.; Mathieu, S.; Brouillette, J.; Fiset, C. Estrogen Regulation of the Transient Outward K+ Current Involves Estrogen Receptor Alpha in Mouse Heart. J. Mol. Cell. Cardiol. 2015, 86, 85–94. [Google Scholar] [CrossRef]
- El Khoury, N.; Ross, J.L.; Long, V.; Thibault, S.; Ethier, N.; Fiset, C. Pregnancy and Oestrogen Regulate Sinoatrial Node Calcium Homeostasis and Accelerate Pacemaking. Cardiovasc. Res. 2018, 114, 1605–1616. [Google Scholar] [CrossRef]
- Demers, J.; Ton, A.; Huynh, F.; Thibault, S.; Ducharme, A.; Paradis, P.; Nemer, M.; Fiset, C. Atrial Electrical Remodeling in Mice With Cardiac-Specific Overexpression of Angiotensin II Type 1 Receptor. JAHA 2022, 11, e023974. [Google Scholar] [CrossRef]
- Mathieu, S.; El Khoury, N.; Rivard, K.; Gélinas, R.; Goyette, P.; Paradis, P.; Nemer, M.; Fiset, C. Reduction in Na+ Current by Angiotensin II Is Mediated by PKCα in Mouse and Human-Induced Pluripotent Stem Cell–Derived Cardiomyocytes. Heart Rhythm. 2016, 13, 1346–1354. [Google Scholar] [CrossRef]
- Mathieu, S.; El Khoury, N.; Rivard, K.; Paradis, P.; Nemer, M.; Fiset, C. Angiotensin II Overstimulation Leads to an Increased Susceptibility to Dilated Cardiomyopathy and Higher Mortality in Female Mice. Sci. Rep. 2018, 8, 952. [Google Scholar] [CrossRef]
- Brouillette, J.; Clark, R.B.; Giles, W.R.; Fiset, C. Functional Properties of K + Currents in Adult Mouse Ventricular Myocytes: Action Potential Repolarization in Murine Cardiac Myocytes. J. Physiol. 2004, 559, 777–798. [Google Scholar] [CrossRef] [PubMed]
- Grandy, S.A.; Brouillette, J.; Fiset, C. Reduction of Ventricular Sodium Current in a Mouse Model of HIV. J. Cardiovasc. Electrophysiol. 2010, 21, 916–922. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thibault, S.; Ton, A.-T.; Huynh, F.; Fiset, C. Connexin Lateralization Contributes to Male Susceptibility to Atrial Fibrillation. Int. J. Mol. Sci. 2022, 23, 10696. https://doi.org/10.3390/ijms231810696
Thibault S, Ton A-T, Huynh F, Fiset C. Connexin Lateralization Contributes to Male Susceptibility to Atrial Fibrillation. International Journal of Molecular Sciences. 2022; 23(18):10696. https://doi.org/10.3390/ijms231810696
Chicago/Turabian StyleThibault, Simon, Anh-Tuan Ton, François Huynh, and Céline Fiset. 2022. "Connexin Lateralization Contributes to Male Susceptibility to Atrial Fibrillation" International Journal of Molecular Sciences 23, no. 18: 10696. https://doi.org/10.3390/ijms231810696
APA StyleThibault, S., Ton, A.-T., Huynh, F., & Fiset, C. (2022). Connexin Lateralization Contributes to Male Susceptibility to Atrial Fibrillation. International Journal of Molecular Sciences, 23(18), 10696. https://doi.org/10.3390/ijms231810696