

Integrated Antitumor Activities of Cellular Immunotherapy with CIK Lymphocytes and Interferons against KIT/PDGFRA Wild Type GIST
 , , ,
, , ,  ,
,  , , , , ,
, , , , ,  ,
, 
 , and add
Show full author list
, and add
Show full author list
Abstract
1. Introduction
2. Results
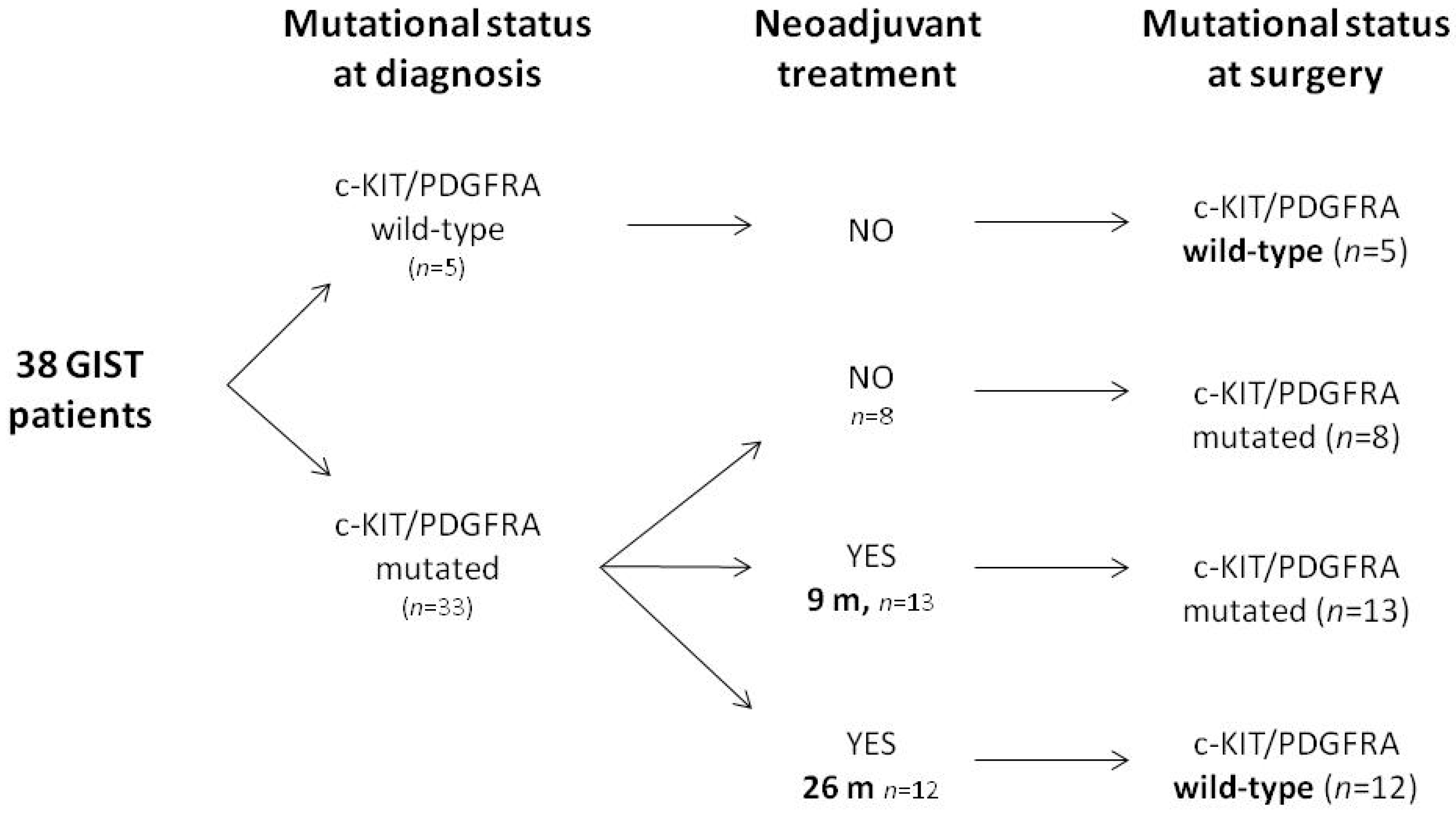
2.1. Generation and Characterization of Primary Cell Lines from Patients with Diagnosis of KIT/PDGFRA WT GIST
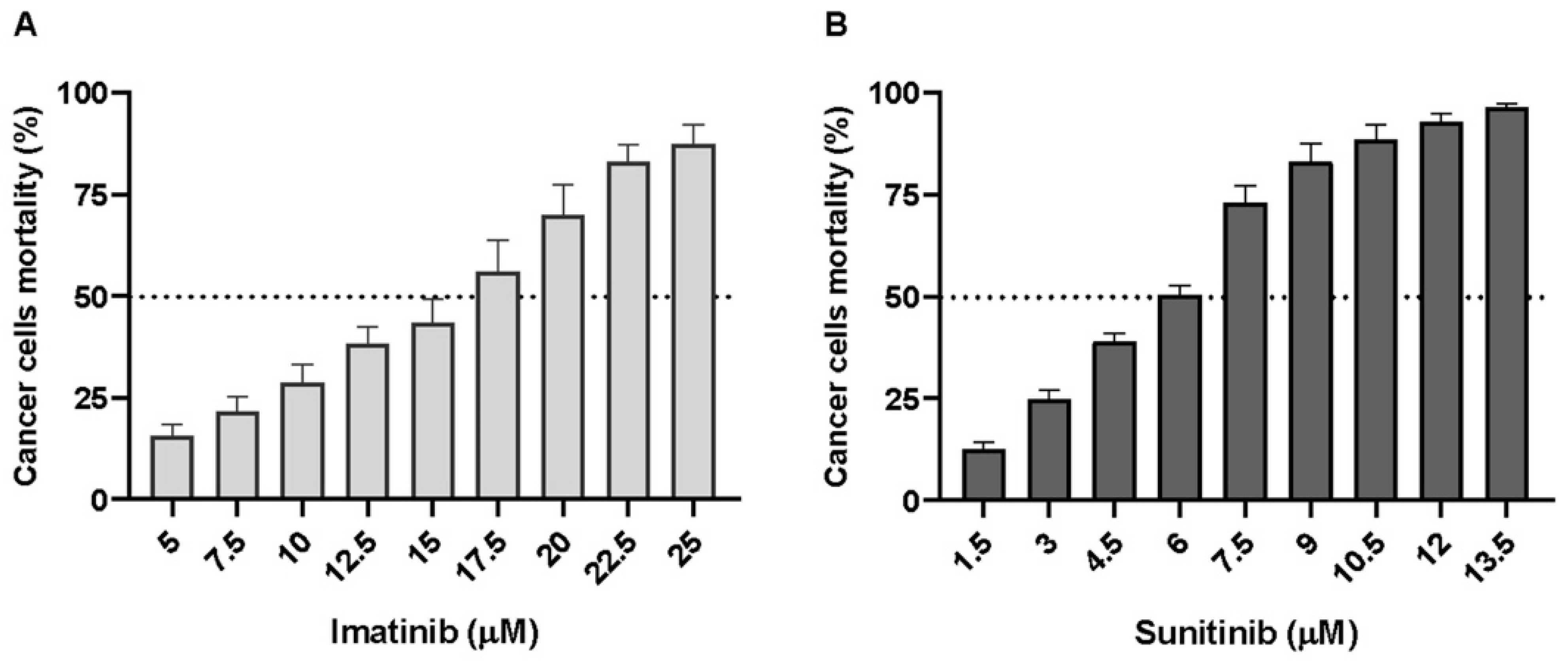
2.2. wtGISTc Are Resistant to Imatinib and Moderately Sensitive to Sunitinib
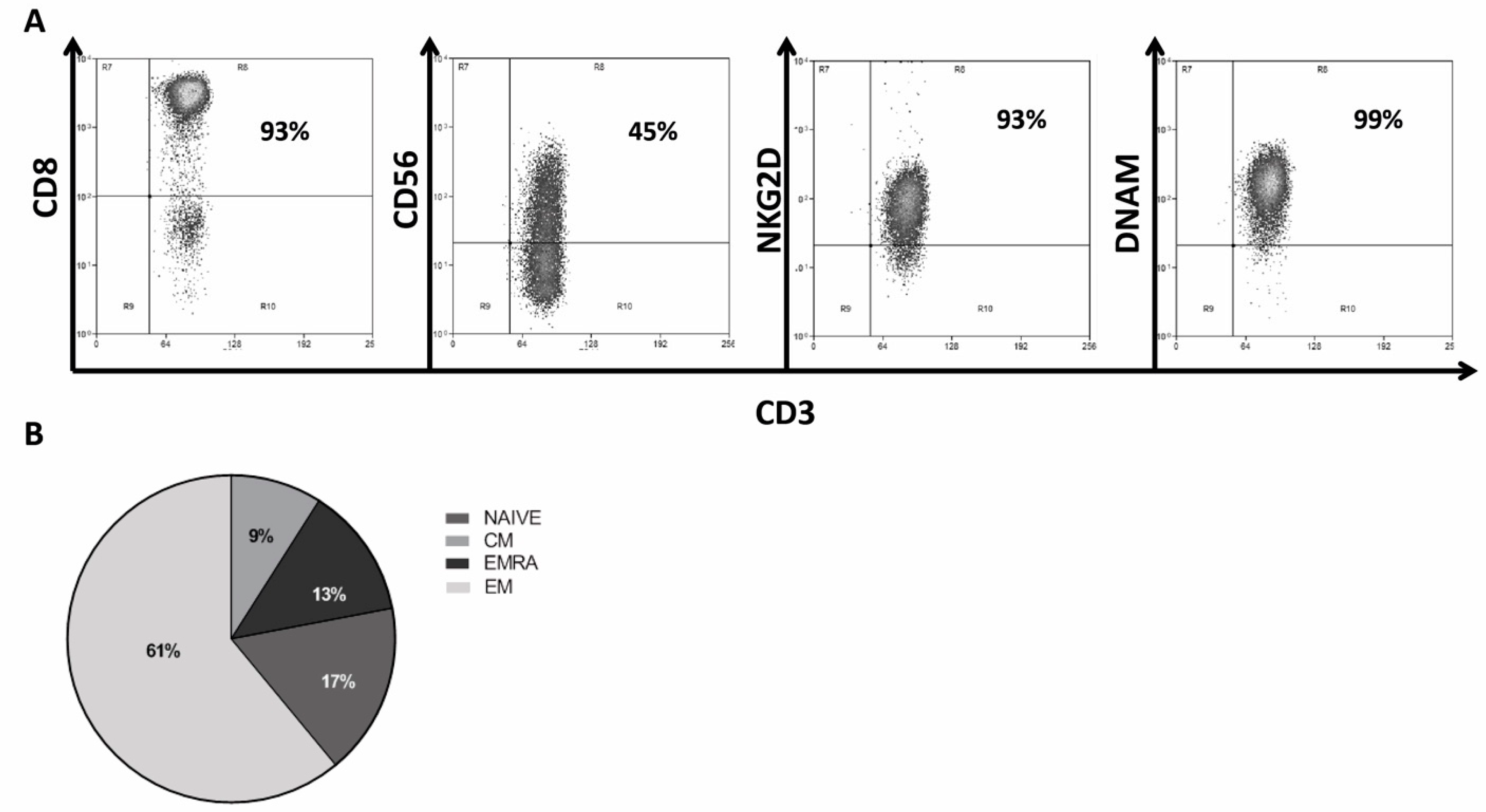
2.3. Generation and Characterization of CIK Lymphocytes from GIST Patients
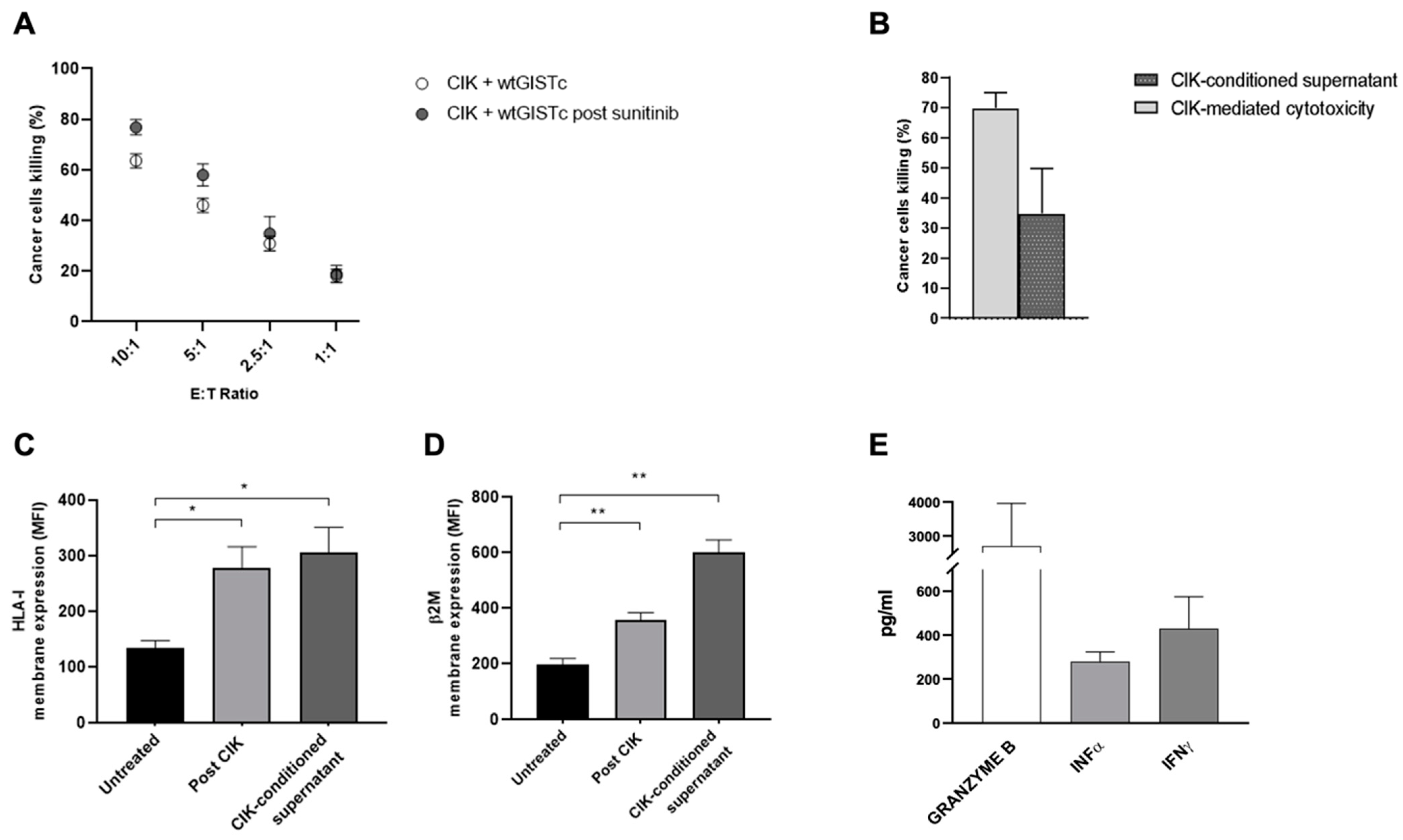
2.4. CIK Are Capable of Direct and Indirect Cancer Cell Killing and Immunomodulation in wtGISTc Resistant to TKI
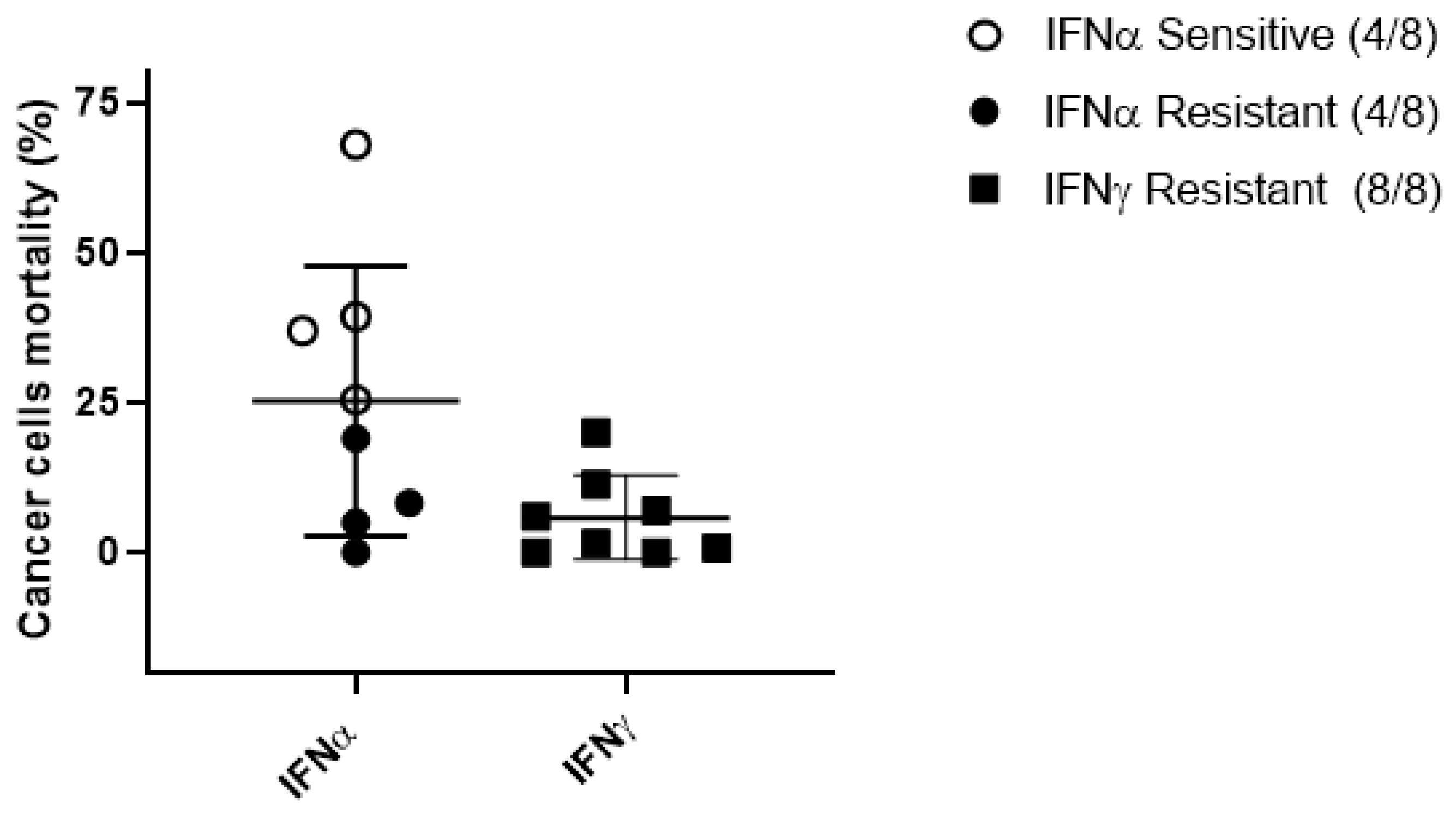
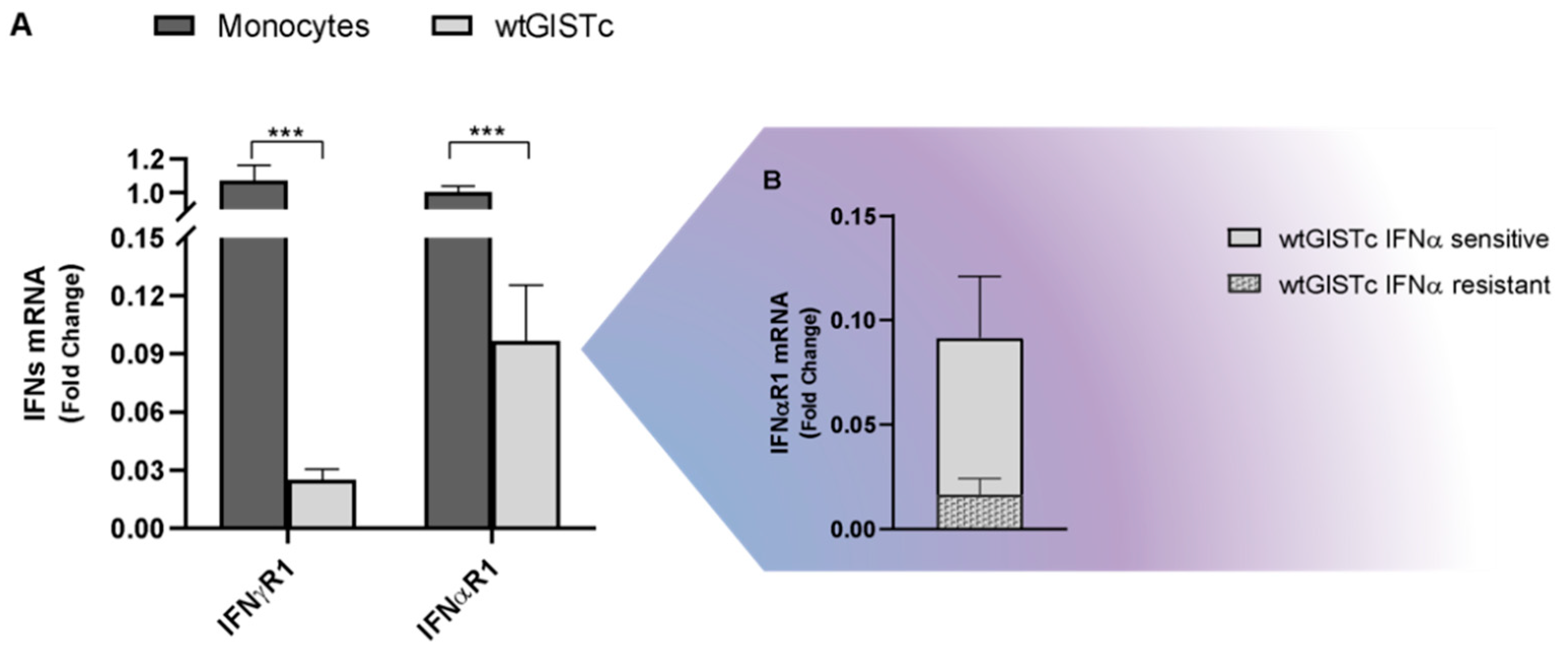
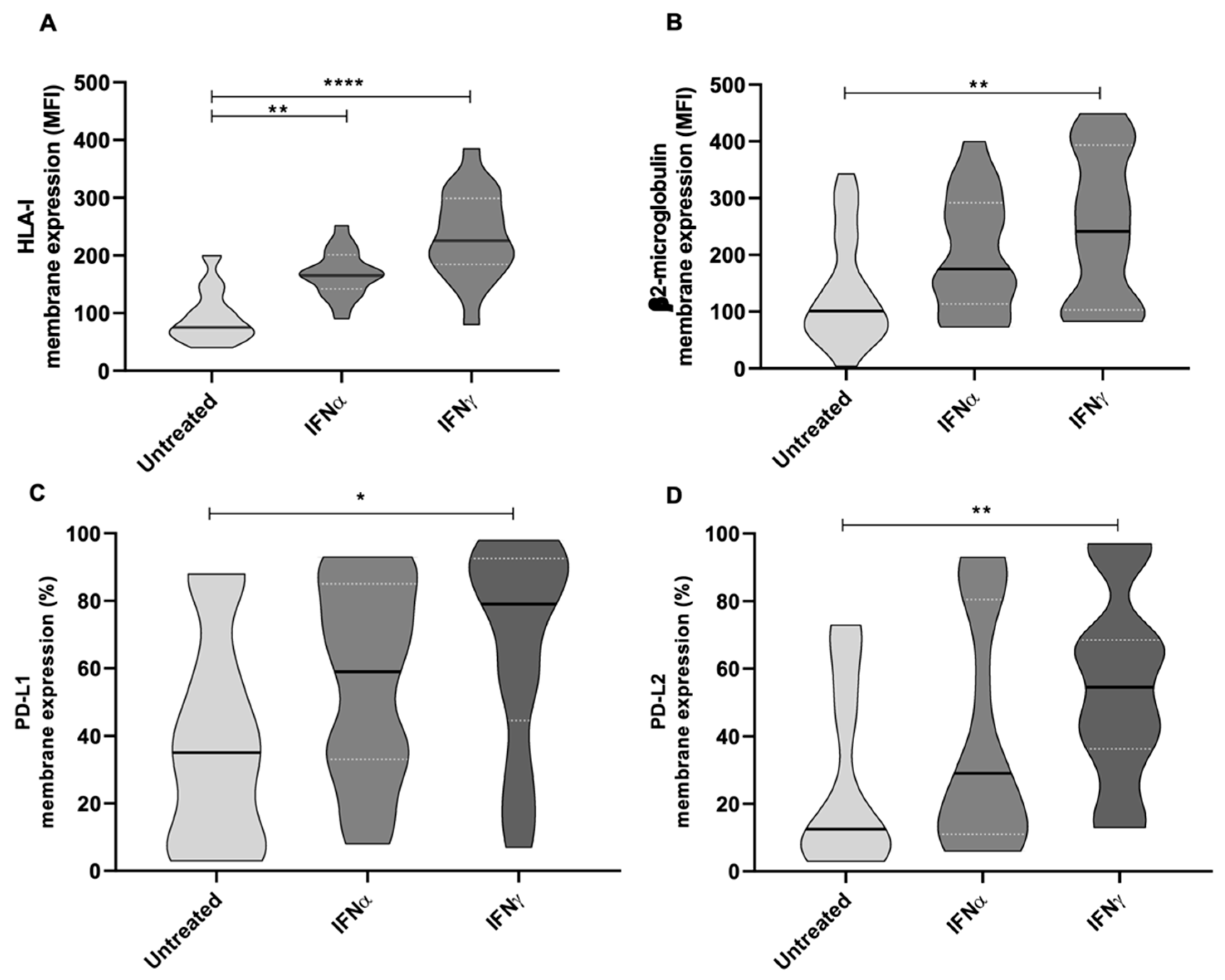
2.5. Antitumor and Immunomodulatory Activity of IFNα and IFNγ in wtGISTc
3. Discussion
4. Materials and Methods
4.1. Experimental Platform and Generation of Wild-Type Primary GIST Cell Lines
4.2. CIK Cells Culture and Ex Vivo Expansion
4.3. Flow Cytometry
4.4. mRNA Analysis
4.5. DNA Sequencing Analysis and Gene Mutation Profile
4.6. In Vitro Drug Sensitivity and Cancer Cell Killing Assays
4.7. In Vitro Soluble Factors Production
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casali, P.G.; Blay, J.Y.; Abecassis, N.; Bajpai, J.; Bauer, S.; Biagini, R.; Bielack, S.; Bonvalot, S.; Boukovinas, I.; Bovee, J.V.M.; et al. Gastrointestinal stromal tumours: ESMO-EURACAN-GENTURIS Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2022, 33, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Van der Graaf, W.T.A.; Tielen, R.; Bonenkamp, J.J.; Lemmens, V.; Verhoeven, R.H.A.; de Wilt, J.H.W. Nationwide trends in the incidence and outcome of patients with gastrointestinal stromal tumour in the imatinib era. Br. J. Surg. 2018, 105, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Søreide, K.; Sandvik, O.M.; Søreide, J.A.; Giljaca, V.; Jureckova, A.; Bulusu, V.R. Global epidemiology of gastrointestinal stromal tumours (GIST): A systematic review of population-based cohort studies. Cancer Epidemiol. 2016, 40, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Demetri, G.D.; von Mehren, M.; Blanke, C.D.; Van den Abbeele, A.D.; Eisenberg, B.; Roberts, P.J.; Heinrich, M.C.; Tuveson, D.A.; Singer, S.; Janicek, M.; et al. Efficacy and safety of imatinib mesylate in advanced gastrointestinal stromal tumors. N. Engl. J. Med. 2002, 347, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Joensuu, H.; Roberts, P.J.; Sarlomo-Rikala, M.; Andersson, L.C.; Tervahartiala, P.; Tuveson, D.; Silberman, S.; Capdeville, R.; Dimitrijevic, S.; Druker, B.; et al. Effect of the tyrosine kinase inhibitor STI571 in a patient with a metastatic gastrointestinal stromal tumor. N. Engl. J. Med. 2001, 344, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Druker, B.J.; Talpaz, M.; Resta, D.J.; Peng, B.; Buchdunger, E.; Ford, J.M.; Lydon, N.B.; Kantarjian, H.; Capdeville, R.; Ohno-Jones, S.; et al. Efficacy and safety of a specific inhibitor of the BCR-ABL tyrosine kinase in chronic myeloid leukemia. N. Engl. J. Med. 2001, 344, 1031–1037. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Martinez-Marín, V.; Serrano, C.; Hindi, N.; López-Guerrero, J.A.; Bisculoa, M.; Ramos-Asensio, R.; Vallejo-Benítez, A.; Marcilla-Plaza, D.; González-Cámpora, R. Gastrointestinal stromal tumors (GISTs): SEAP-SEOM consensus on pathologic and molecular diagnosis. Clin. Transl. Oncol. 2017, 19, 536–545. [Google Scholar] [CrossRef]
- Corless, C.L.; Barnett, C.M.; Heinrich, M.C. Gastrointestinal stromal tumours: Origin and molecular oncology. Nat. Rev. Cancer 2011, 11, 865–878. [Google Scholar] [CrossRef]
- Miettinen, M.; Lasota, J. Histopathology of gastrointestinal stromal tumor. J. Surg. Oncol. 2011, 104, 865–873. [Google Scholar] [CrossRef]
- Rossi, S.; Gasparotto, D.; Miceli, R.; Toffolatti, L.; Gallina, G.; Scaramel, E.; Marzotto, A.; Boscato, E.; Messerini, L.; Bearzi, I.; et al. KIT, PDGFRA, and BRAF mutational spectrum impacts on the natural history of imatinib-naive localized GIST: A population-based study. Am. J. Surg. Pathol. 2015, 39, 922–930. [Google Scholar] [CrossRef]
- Huss, S.; Pasternack, H.; Ihle, M.A.; Merkelbach-Bruse, S.; Heitkötter, B.; Hartmann, W.; Trautmann, M.; Gevensleben, H.; Büttner, R.; Schildhaus, H.U.; et al. Clinicopathological and molecular features of a large cohort of gastrointestinal stromal tumors (GISTs) and review of the literature: BRAF mutations in KIT/PDGFRA wild-type GISTs are rare events. Hum. Pathol. 2017, 62, 206–214. [Google Scholar] [CrossRef]
- Kalfusova, A.; Linke, Z.; Kalinova, M.; Krskova, L.; Hilska, I.; Szabova, J.; Vicha, A.; Kodet, R. Gastrointestinal stromal tumors—Summary of mutational status of the primary/secondary KIT/PDGFRA mutations, BRAF mutations and SDH defects. Pathol. Res. Pract. 2019, 215, 152708. [Google Scholar] [CrossRef]
- Patel, S.R.; Reichardt, P. An updated review of the treatment landscape for advanced gastrointestinal stromal tumors. Cancer 2021, 127, 2187–2195. [Google Scholar] [CrossRef]
- Tan, Y.; Trent, J.C.; Wilky, B.A.; Kerr, D.A.; Rosenberg, A.E. Current status of immunotherapy for gastrointestinal stromal tumor. Cancer Gene Ther. 2017, 24, 130–133. [Google Scholar] [CrossRef]
- Vallilas, C.; Sarantis, P.; Kyriazoglou, A.; Koustas, E.; Theocharis, S.; Papavassiliou, A.G.; Karamouzis, M.V. Gastrointestinal Stromal Tumors (GISTs): Novel Therapeutic Strategies with Immunotherapy and Small Molecules. Int. J. Mol. Sci. 2021, 22, 493. [Google Scholar] [CrossRef]
- Arshad, J.; Costa, P.A.; Barreto-Coelho, P.; Valdes, B.N.; Trent, J.C. Immunotherapy Strategies for Gastrointestinal Stromal Tumor. Cancers 2021, 13, 3525. [Google Scholar] [CrossRef]
- Reilley, M.J.; Bailey, A.; Subbiah, V.; Janku, F.; Naing, A.; Falchook, G.; Karp, D.; Piha-Paul, S.; Tsimberidou, A.; Fu, S.; et al. Phase I clinical trial of combination imatinib and ipilimumab in patients with advanced malignancies. J. Immunother. Cancer 2017, 5, 35. [Google Scholar] [CrossRef]
- D’Angelo, S.P.; Shoushtari, A.N.; Keohan, M.L.; Dickson, M.A.; Gounder, M.M.; Chi, P.; Loo, J.K.; Gaffney, L.; Schneider, L.; Patel, Z.; et al. Combined KIT and CTLA-4 Blockade in Patients with Refractory GIST and Other Advanced Sarcomas: A Phase Ib Study of Dasatinib plus Ipilimumab. Clin. Cancer Res. 2017, 23, 2972–2980. [Google Scholar] [CrossRef]
- Liu, M.; Etherington, M.S.; Hanna, A.; Medina, B.D.; Vitiello, G.A.; Bowler, T.G.; Param, N.J.; Levin, L.; Rossi, F.; DeMatteo, R.P. Oncogenic KIT Modulates Type I IFN-Mediated Antitumor Immunity in GIST. Cancer Immunol. Res. 2021, 9, 542–553. [Google Scholar] [CrossRef]
- Cai, L.; Michelakos, T.; Yamada, T.; Fan, S.; Wang, X.; Schwab, J.H.; Ferrone, C.R.; Ferrone, S. Defective HLA class I antigen processing machinery in cancer. Cancer Immunol. Immunother. 2018, 67, 999–1009. [Google Scholar] [CrossRef]
- Tieniber, A.D.; Hanna, A.N.; Medina, B.D.; Vitiello, G.A.; Etherington, M.S.; Liu, M.; Do, K.J.; Rossi, F.; DeMatteo, R.P. Tyrosine kinase inhibition alters intratumoral CD8+ T-cell subtype composition and activity. Cancer Immunol. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.S.; Hecht, J.R.; Rosen, L.; Wainberg, Z.A.; Wang, X.; Douek, M.; Hagopian, A.; Andes, R.; Sauer, L.; Brackert, S.R.; et al. A Randomized Phase II Study of Nivolumab Monotherapy or Nivolumab Combined with Ipilimumab in Patients with Advanced Gastrointestinal Stromal Tumors. Clin. Cancer Res. 2022, 28, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Leuci, V.; Donini, C.; Grignani, G.; Rotolo, R.; Mesiano, G.; Fiorino, E.; Gammaitoni, L.; D’Ambrosio, L.; Merlini, A.; Landoni, E.; et al. CSPG4-Specific CAR.CIK Lymphocytes as a Novel Therapy for the Treatment of Multiple Soft-Tissue Sarcoma Histotypes. Clin. Cancer Res. 2020, 26, 6321–6334. [Google Scholar] [CrossRef] [PubMed]
- Mesiano, G.; Grignani, G.; Fiorino, E.; Leuci, V.; Rotolo, R.; D’Ambrosio, L.; Salfi, C.; Gammaitoni, L.; Giraudo, L.; Pisacane, A.; et al. Cytokine Induced Killer cells are effective against sarcoma cancer stem cells spared by chemotherapy and target therapy. Oncoimmunology 2018, 7, e1465161. [Google Scholar] [CrossRef]
- Sangiolo, D.; Mesiano, G.; Gammaitoni, L.; Aglietta, M.; Grignani, G. Activity of cytokine-induced killer cells against bone and soft tissue sarcoma. Oncoimmunology 2014, 3, e28269. [Google Scholar] [CrossRef][Green Version]
- Sangiolo, D.; Mesiano, G.; Gammaitoni, L.; Leuci, V.; Todorovic, M.; Giraudo, L.; Cammarata, C.; Dell’Aglio, C.; D’Ambrosio, L.; Pisacane, A.; et al. Cytokine-induced killer cells eradicate bone and soft-tissue sarcomas. Cancer Res. 2014, 74, 119–129. [Google Scholar] [CrossRef]
- Gasparotto, D.; Sbaraglia, M.; Rossi, S.; Baldazzi, D.; Brenca, M.; Mondello, A.; Nardi, F.; Racanelli, D.; Cacciatore, M.; Paolo Dei Tos, A.; et al. Tumor genotype, location, and malignant potential shape the immunogenicity of primary untreated gastrointestinal stromal tumors. JCI Insight 2020, 5, e142560. [Google Scholar] [CrossRef]
- Vitiello, G.A.; Bowler, T.G.; Liu, M.; Medina, B.D.; Zhang, J.Q.; Param, N.J.; Loo, J.K.; Goldfeder, R.L.; Chibon, F.; Rossi, F.; et al. Differential immune profiles distinguish the mutational subtypes of gastrointestinal stromal tumor. J. Clin. Investig. 2019, 129, 1863–1877. [Google Scholar] [CrossRef]
- Chen, L.L.; Gouw, L.; Sabripour, M.; Hwu, W.J.; Benjamin, R.S. Combining targeted therapy with immunotherapy (interferon-α): Rational, efficacy in gastrointestinal stromal tumor model and implications in other malignancies. Oncoimmunology 2012, 1, 773–776. [Google Scholar] [CrossRef][Green Version]
- Chen, L.L.; Chen, X.; Choi, H.; Sang, H.; Chen, L.C.; Zhang, H.; Gouw, L.; Andtbacka, R.H.; Chan, B.K.; Rodesch, C.K.; et al. Exploiting antitumor immunity to overcome relapse and improve remission duration. Cancer Immunol. Immunother. 2012, 61, 1113–1124. [Google Scholar] [CrossRef]
- Narumi, K.; Udagawa, T.; Kondoh, A.; Kobayashi, A.; Hara, H.; Ikarashi, Y.; Ohnami, S.; Takeshita, F.; Ochiya, T.; Okada, T.; et al. In vivo delivery of interferon-α gene enhances tumor immunity and suppresses immunotolerance in reconstituted lymphopenic hosts. Gene Ther. 2012, 19, 34–48. [Google Scholar] [CrossRef]
- Kadowaki, N.; Antonenko, S.; Lau, J.Y.; Liu, Y.J. Natural interferon alpha/beta-producing cells link innate and adaptive immunity. J. Exp. Med. 2000, 192, 219–226. [Google Scholar] [CrossRef]
- Bauer, S.; Hartmann, J.T.; de Wit, M.; Lang, H.; Grabellus, F.; Antoch, G.; Niebel, W.; Erhard, J.; Ebeling, P.; Zeth, M.; et al. Resection of residual disease in patients with metastatic gastrointestinal stromal tumors responding to treatment with imatinib. Int. J. Cancer 2005, 117, 316–325. [Google Scholar] [CrossRef]
- Seifert, A.M.; Zeng, S.; Zhang, J.Q.; Kim, T.S.; Cohen, N.A.; Beckman, M.J.; Medina, B.D.; Maltbaek, J.H.; Loo, J.K.; Crawley, M.H.; et al. PD-1/PD-L1 Blockade Enhances T-cell Activity and Antitumor Efficacy of Imatinib in Gastrointestinal Stromal Tumors. Clin. Cancer Res. 2017, 23, 454–465. [Google Scholar] [CrossRef]
- Zhao, R.; Song, Y.; Wang, Y.; Huang, Y.; Li, Z.; Cui, Y.; Yi, M.; Xia, L.; Zhuang, W.; Wu, X.; et al. PD-1/PD-L1 blockade rescue exhausted CD8+ T cells in gastrointestinal stromal tumours via the PI3K/Akt/mTOR signalling pathway. Cell Prolif. 2019, 52, e12571. [Google Scholar] [CrossRef]
- Bertucci, F.; Finetti, P.; Mamessier, E.; Pantaleo, M.A.; Astolfi, A.; Ostrowski, J.; Birnbaum, D. PDL1 expression is an independent prognostic factor in localized GIST. Oncoimmunology 2015, 4, e1002729. [Google Scholar] [CrossRef]
- Desai, S.A.; Wang, X.; Noronha, E.J.; Zhou, Q.; Rebmann, V.; Grosse-Wilde, H.; Moy, F.J.; Powers, R.; Ferrone, S. Structural relatedness of distinct determinants recognized by monoclonal antibody TP25.99 on beta 2-microglobulin-associated and beta 2-microglobulin-free HLA class I heavy chains. J. Immunol. 2000, 165, 3275–3283. [Google Scholar] [CrossRef]
- Lampson, L.A.; Levy, R. Two populations of Ia-like molecules on a human B cell line. J. Immunol. 1980, 125, 293–299. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| wtGISTc | Gene Name | Variant Type | Single Nucleotide Variant ID | VAF (%) | Fold Change |
|---|---|---|---|---|---|
| S012 | SPEN | inframe_deletion | chr1_16260673_CCTCACTGGTCTGGTGAGCGCA_C | 27 | |
| LATS2 | missense_variant | chr13_21562627_G_A | 36 | ||
| JAK2 | amplification | chr9 | 1.946 | ||
| PTEN | amplification | chr10 | 1.445 | ||
| S061 | / | / | / | / | |
| S075 | / | / | / | / | |
| S108 | NOTCH1 | missense variant:splice region variant | chr9_139401170_G_A | 51 | |
| S125 | / | / | / | / | |
| S148 | ARID1B | missense_variant | chr6_157100551_G_C | 47 | |
| PRKDC | missense_variant | chr8_48805728_C_G | 24 | ||
| S173 | MYCN | missense_variant | chr2_16082319_C_T | 20 | |
| GNAS | missense_variant | chr20_57474036_G_T | 19 | ||
| S188 | PGR | missense_variant | chr11_100998322_C_G | 31 | |
| S220 | HIST3H3 | missense_variant | chr1_228612992_G_A | 16 | |
| LRP1B | missense_variant | chr2_141201961_G_T | 19 | ||
| CUL3 | missense_variant | chr2_225379486_G_A | 20 | ||
| PRKDC | missense_variant | chr8_48866988_T_G | 58 | ||
| MALT1 | missense_variant | chr18_56363602_A_G | 16 | ||
| NF2 | stop_gained | chr22_30038226_C_A | 50 | ||
| FGFR1 | amplification | chr8 | 1.533 | ||
| MYC | amplification | chr8 | 1.637 | ||
| S243 | IRS1 | stop_gained | chr2_227661875_G_T | 9 | |
| PDGFRB | inframe_deletion | chr5_149495390_AGCTCTG_A | 7 | ||
| NOTCH4 | missense_variant | chr6_32189007_G_A | 7 | ||
| S288 | FGF2 | missense_variant | chr4_123813429_C_T | 51 | |
| FAT1 | missense_variant | chr4_187628598_T_C | 46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiorino, E.; Merlini, A.; D’Ambrosio, L.; Cerviere, I.; Berrino, E.; Marchiò, C.; Giraudo, L.; Basiricò, M.; Massa, A.; Donini, C.; et al. Integrated Antitumor Activities of Cellular Immunotherapy with CIK Lymphocytes and Interferons against KIT/PDGFRA Wild Type GIST. Int. J. Mol. Sci. 2022, 23, 10368. https://doi.org/10.3390/ijms231810368
Fiorino E, Merlini A, D’Ambrosio L, Cerviere I, Berrino E, Marchiò C, Giraudo L, Basiricò M, Massa A, Donini C, et al. Integrated Antitumor Activities of Cellular Immunotherapy with CIK Lymphocytes and Interferons against KIT/PDGFRA Wild Type GIST. International Journal of Molecular Sciences. 2022; 23(18):10368. https://doi.org/10.3390/ijms231810368
Chicago/Turabian StyleFiorino, Erika, Alessandra Merlini, Lorenzo D’Ambrosio, Ilaria Cerviere, Enrico Berrino, Caterina Marchiò, Lidia Giraudo, Marco Basiricò, Annamaria Massa, Chiara Donini, and et al. 2022. "Integrated Antitumor Activities of Cellular Immunotherapy with CIK Lymphocytes and Interferons against KIT/PDGFRA Wild Type GIST" International Journal of Molecular Sciences 23, no. 18: 10368. https://doi.org/10.3390/ijms231810368
APA StyleFiorino, E., Merlini, A., D’Ambrosio, L., Cerviere, I., Berrino, E., Marchiò, C., Giraudo, L., Basiricò, M., Massa, A., Donini, C., Leuci, V., Rotolo, R., Galvagno, F., Vitali, L., Proment, A., Ferrone, S., Pisacane, A., Pignochino, Y., Aglietta, M., ... Sangiolo, D. (2022). Integrated Antitumor Activities of Cellular Immunotherapy with CIK Lymphocytes and Interferons against KIT/PDGFRA Wild Type GIST. International Journal of Molecular Sciences, 23(18), 10368. https://doi.org/10.3390/ijms231810368