
Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
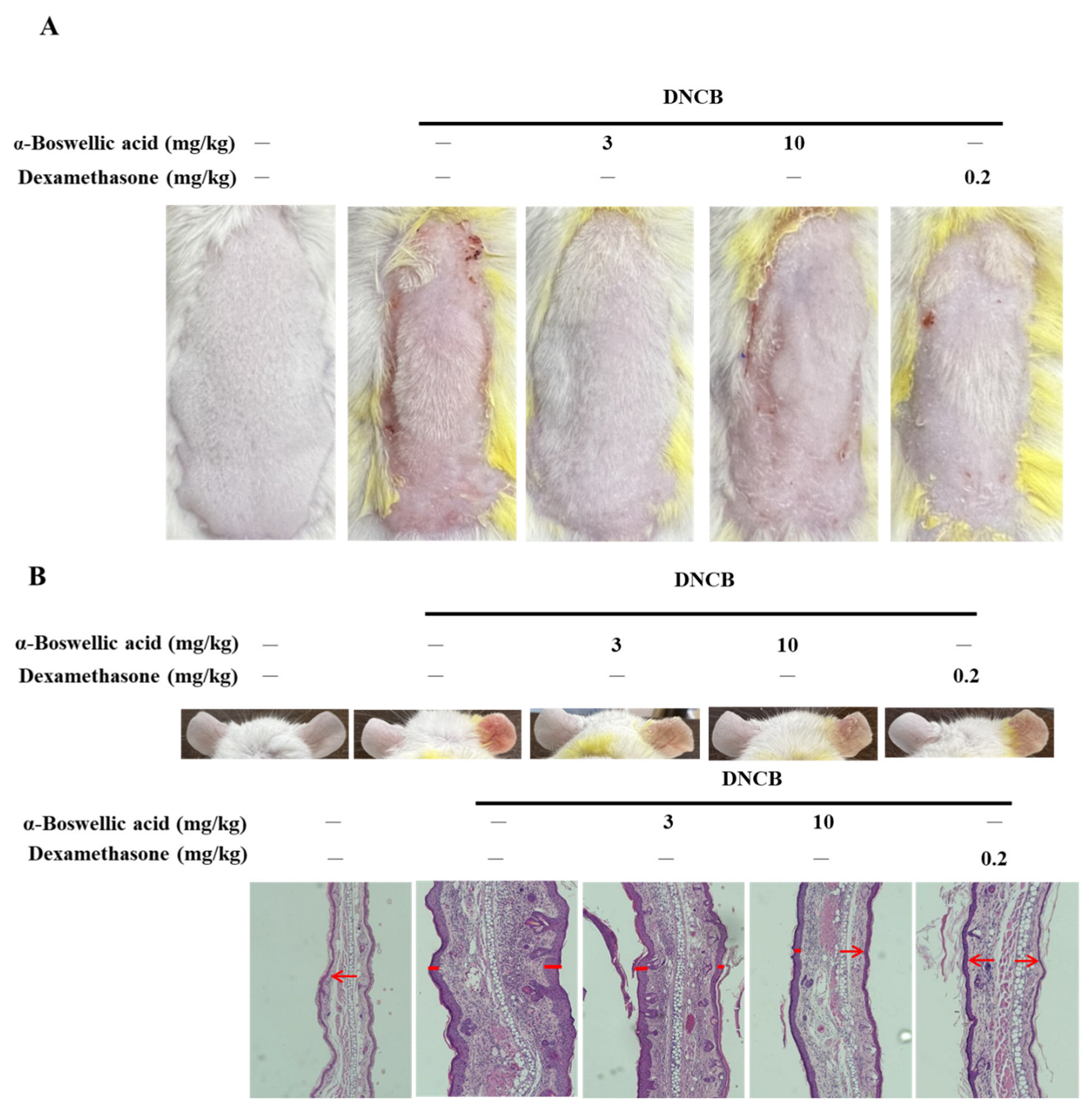
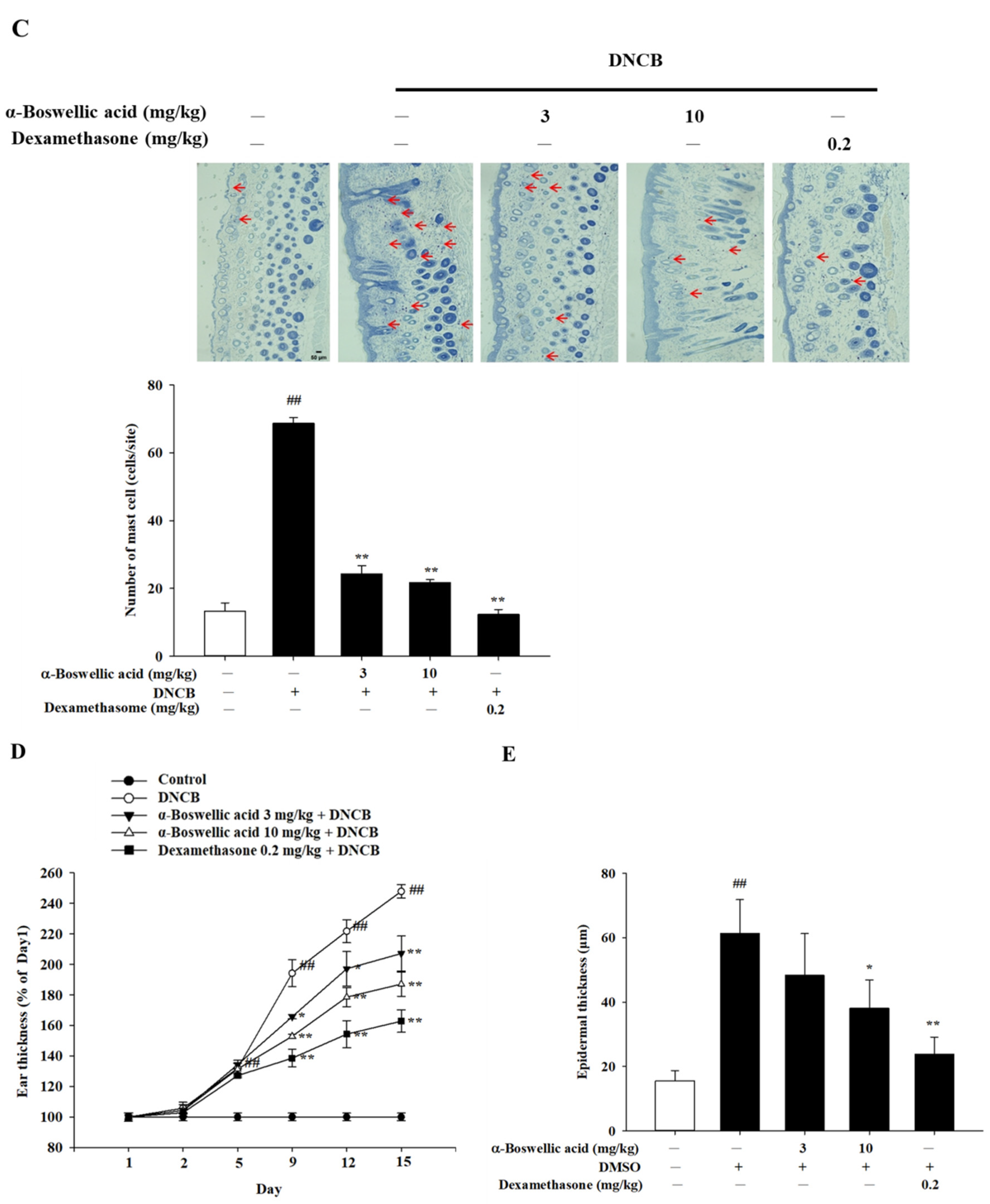
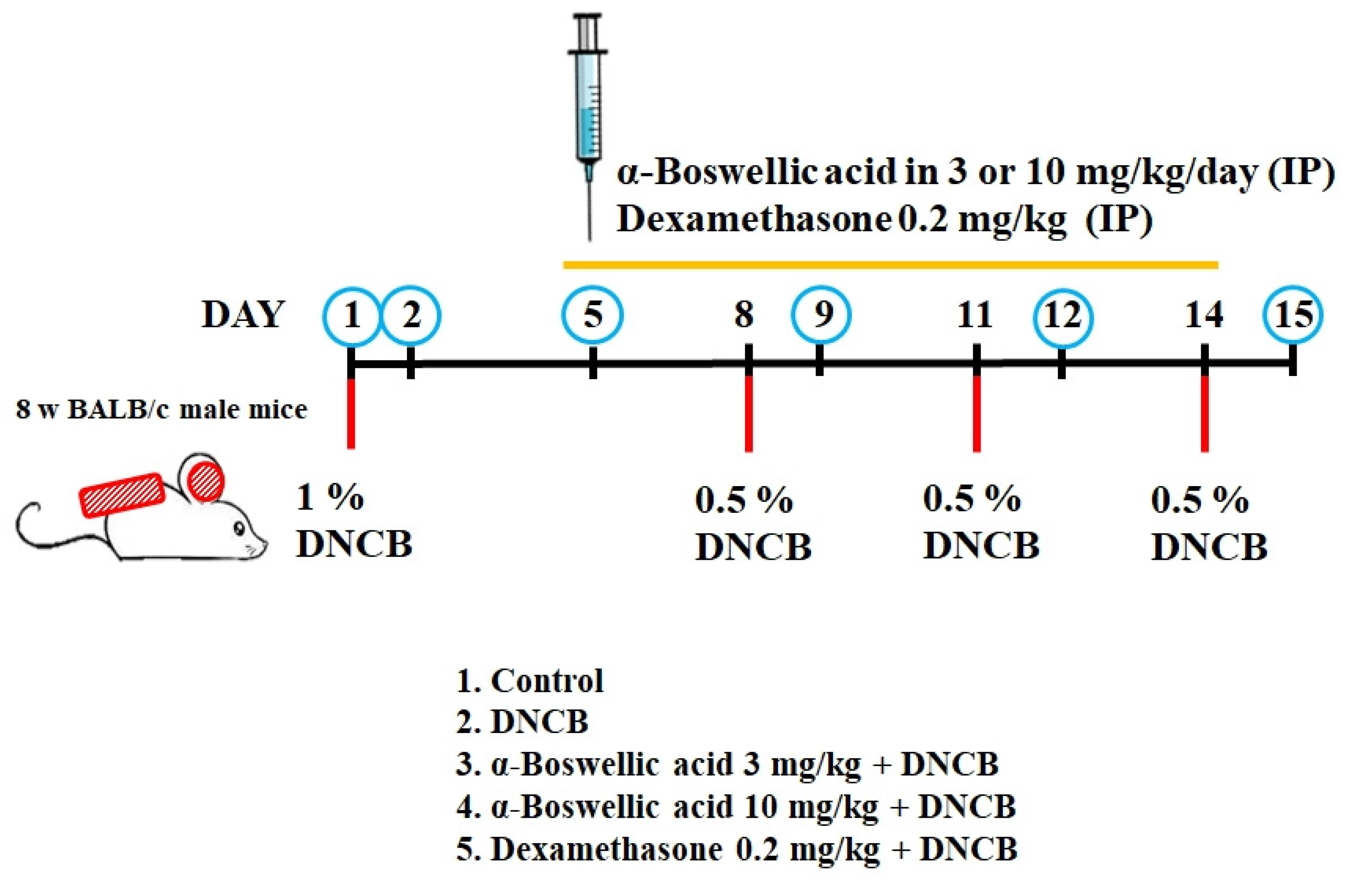
2.1. Effects of α-Boswellic Acid on 2,4-Dinitrochlorobenzene-Induced Atopic-like Dermatitis in BALB/c Mice
2.2. Effects of α-Boswellic Acid on TEWL, Hydration, and Erythema on 2,4-Dinitrochlorobenzene Induced Atopic-like Dermatitis in BALB/c Mice
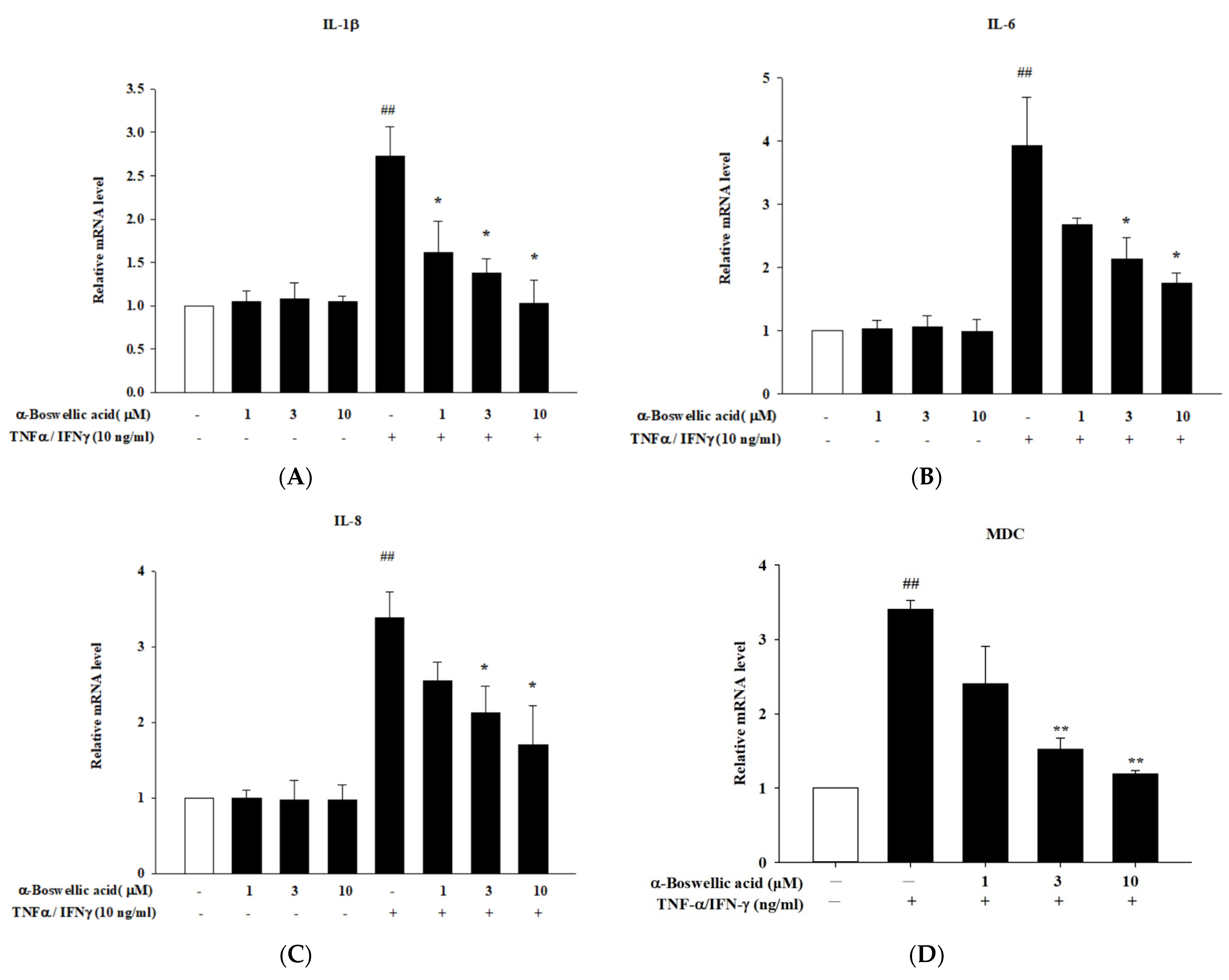
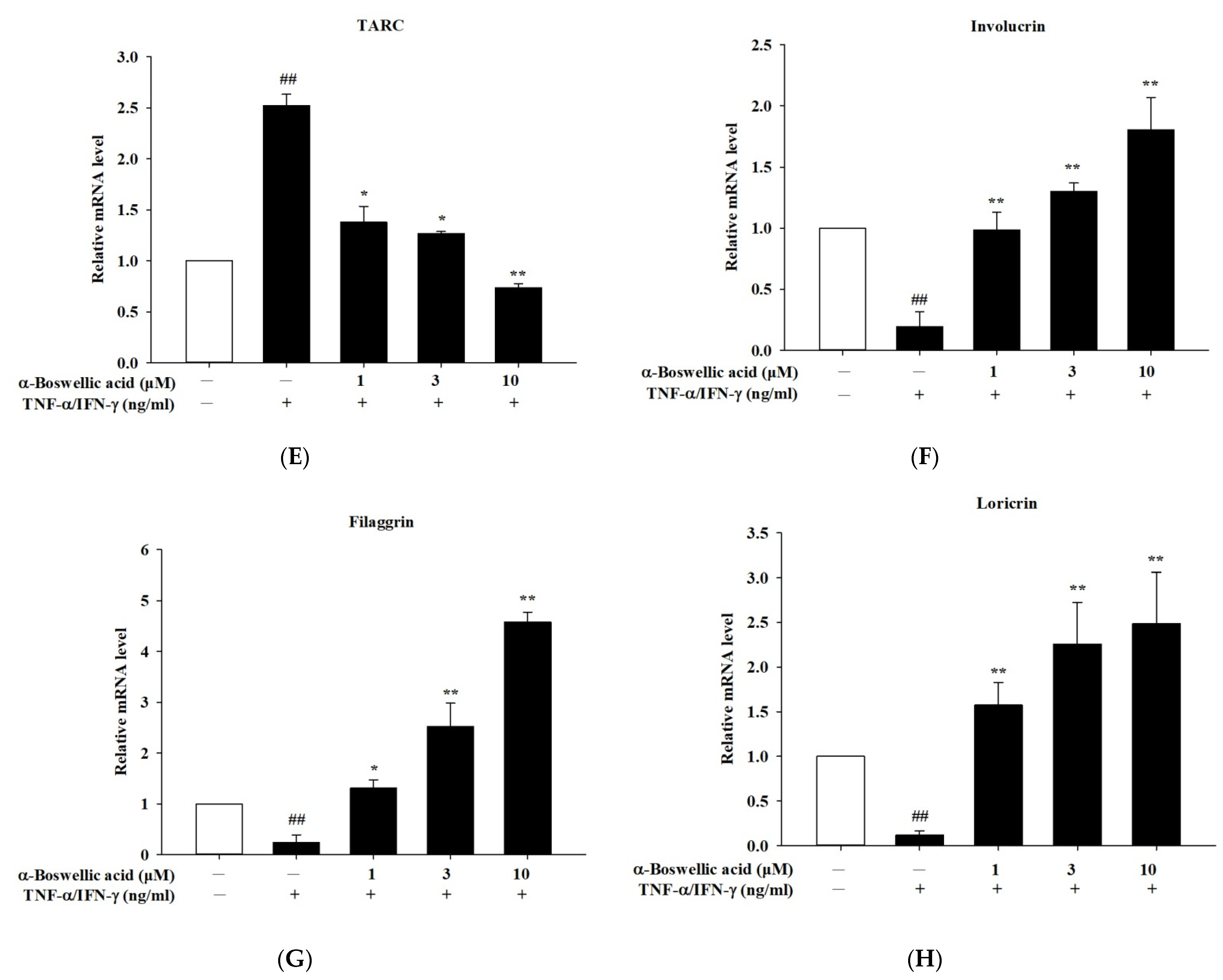
2.3. Effects of α-Boswellic Acid on TNF-α/IFN-γ-Induced Proinflammatory Cytokines, Chemokines, and Barrier Protein mRNA Expressions in HaCaT Cells
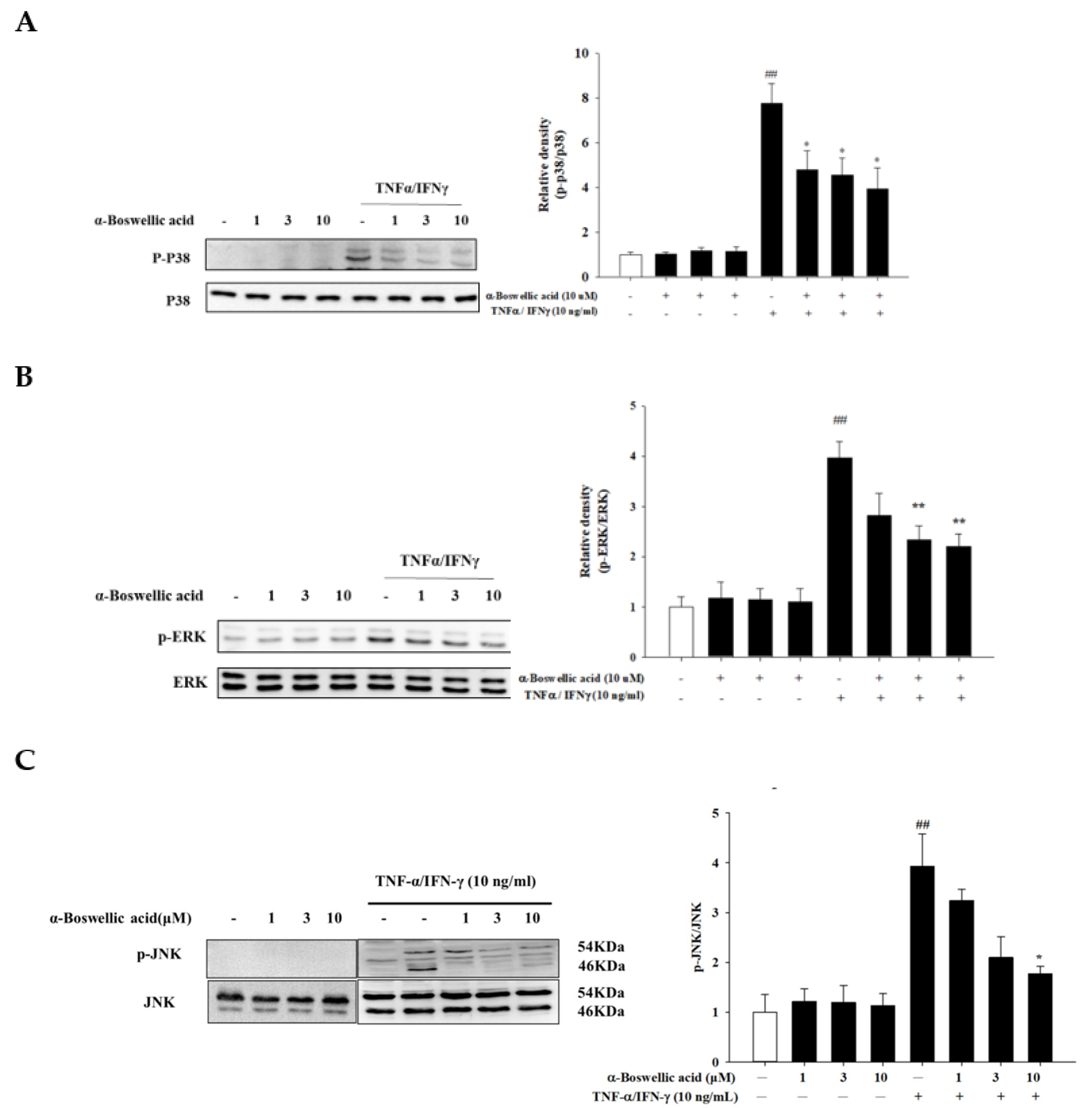
2.4. Effects of α-Boswellic Acid on the MAP Kinase Signaling Pathway in TNF-α/IFN-γ-Induced HaCaT Cells
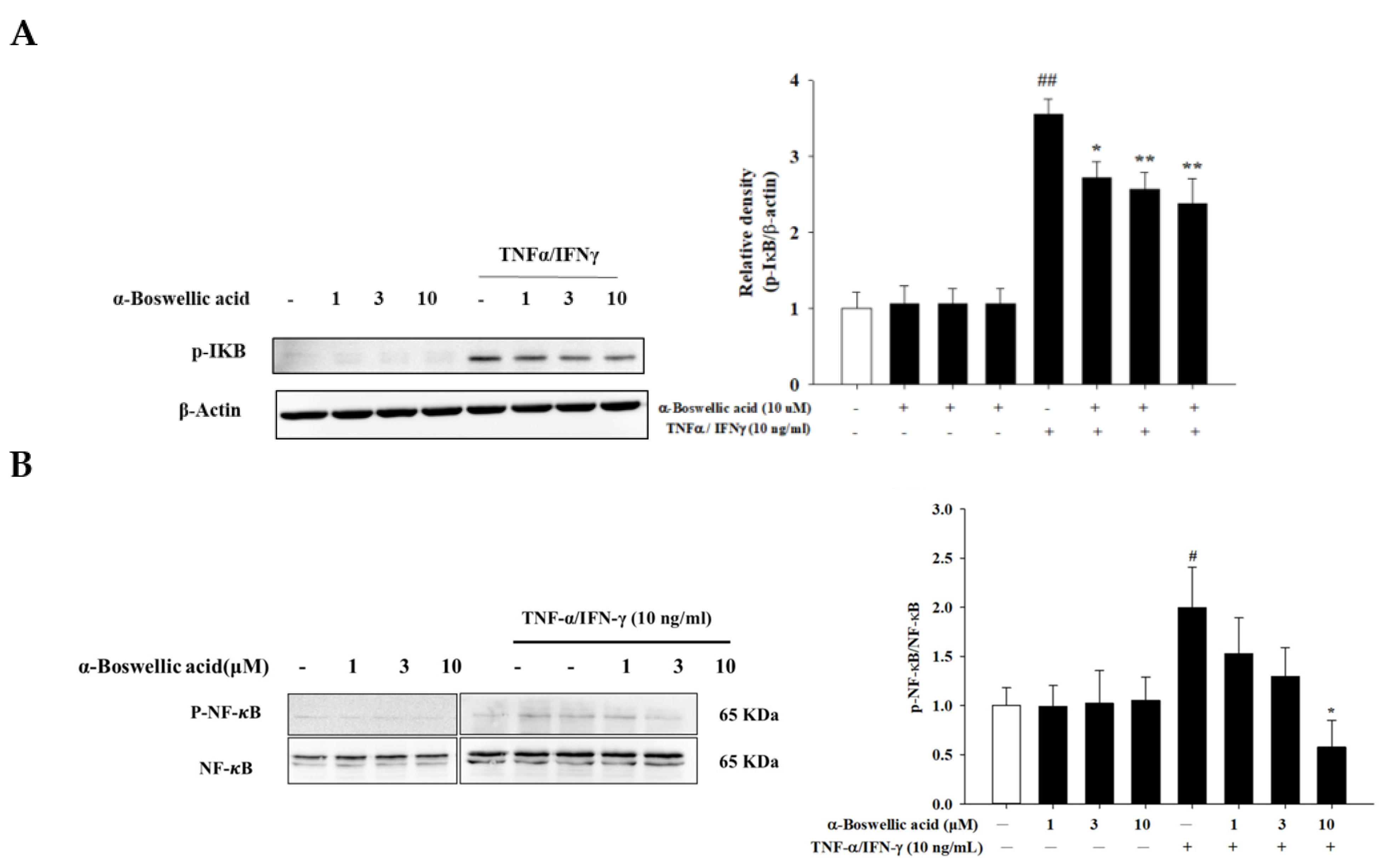
2.5. Effects of α-Boswellic Acid on the Degradation of IκB and Phosphorylation of NF-κB/p65 Protein in TNF-α-/IFN-γ-Induced HaCaT Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. MTT Assay
4.4. mRNA Expression Assay
4.5. Western Blotting Analyses
4.6. Animal
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Efferth, T.; Oesch, F. Anti-inflammatory and anti-cancer activities of frankincense: Targets, treatments and toxicities. Semin. Cancer Biol. 2022, 80, 39–57. [Google Scholar] [CrossRef] [PubMed]
- Banno, N.; Akihisa, T.; Yasukawa, K.; Tokuda, H.; Tabata, K.; Nakamura, Y.; Nishimura, R.; Kimura, Y.; Suzuki, T. Anti-inflammatory activities of the triterpene acids from the resin of Boswellia carteri. J. Ethnopharmacol. 2006, 107, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Langmead, L.; Rampton, D. Complementary and alternative therapies for inflammatory bowel disease. Aliment. Pharmacol. Ther. 2006, 23, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Al-Saidi, S.; Rameshkumar, K.; Hisham, A.; Sivakumar, N.; Al-Kindy, S. Composition and Antibacterial Activity of the Essential Oils of Four Commercial Grades of Omani Luban, the Oleo-Gum Resin of Boswellia sacra Flueck. Chem. Biodivers. 2012, 9, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.J. Medicinal properties of frankincense. Int. J. Nutr. Pharmacol. Neurol. Dis. 2012, 2, 79. [Google Scholar] [CrossRef]
- Mikhaeil, B.R.; Maatooq, G.T.; Badria, F.A.; Amer, M.M. Chemistry and immunomodulatory activity of frankincense oil. Z. Nat. C 2003, 58, 230–238. [Google Scholar] [CrossRef]
- Mishra, N.; Bstia, S.; Mishra, G.; Chowdary, K.; Patra, S. Anti-arthritic activity of Glycyrrhiza glabra, Boswellia serrata and their synergistic activity in combined formulation studied in freund’s adjuvant induced arthritic rats. J. Pharm. Educ. Res. 2011, 2, 92. [Google Scholar]
- Mothana, R.A. Anti-inflammatory, antinociceptive and antioxidant activities of the endemic Soqotraen Boswellia elongata Balf. f. and Jatropha unicostata Balf. f. in different experimental models. Food Chem. Toxicol. 2011, 49, 2594–2599. [Google Scholar] [CrossRef]
- Mothana, R.A.; Hasson, S.S.; Schultze, W.; Mowitz, A.; Lindequist, U. Phytochemical composition and in vitro antimicrobial and antioxidant activities of essential oils of three endemic Soqotraen Boswellia species. Food Chem. 2011, 126, 1149–1154. [Google Scholar] [CrossRef]
- Shehata, A.M.; Quintanilla-Fend, L.; Bettio, S.; Singh, C.; Ammon, H. Prevention of multiple low-dose streptozotocin (MLD-STZ) diabetes in mice by an extract from gum resin of Boswellia serrata (BE). Phytomedicine 2011, 18, 1037–1044. [Google Scholar] [CrossRef]
- Al-Yasiry, A.R.; Kiczorowska, B. Frankincense—Therapeutic properties. Postepy Hig. Med. Dosw 2016, 70, 380–391. [Google Scholar] [CrossRef]
- Roy, N.K.; Parama, D.; Banik, K.; Bordoloi, D.; Devi, A.K.; Thakur, K.K.; Padmavathi, G.; Shakibaei, M.; Fan, L.; Sethi, G.; et al. An Update on Pharmacological Potential of Boswellic Acids against Chronic Diseases. Int. J. Mol. Sci. 2019, 20, 4101. [Google Scholar] [CrossRef]
- Roy, N.K.; Deka, A.; Bordoloi, D.; Mishra, S.; Kumar, A.P.; Sethi, G.; Kunnumakkara, A.B. The potential role of boswellic acids in cancer prevention and treatment. Cancer Lett. 2016, 377, 74–86. [Google Scholar] [CrossRef]
- Wang, D.; Ge, S.; Bai, J.; Song, Y. Boswellic acid exerts potent anticancer effects in HCT-116 human colon cancer cells mediated via induction of apoptosis, cell cycle arrest, cell migration inhibition and inhibition of PI3K/AKT signalling pathway. J. BUON 2018, 23, 340–345. [Google Scholar]
- Wang, M.X.; Zhao, J.X.; Meng, Y.J.; Di, T.T.; Xu, X.L.; Xie, X.J.; Lin, Y.; Zhang, L.; Wang, N.; Li, P.; et al. Acetyl-11-keto-β-boswellic acid inhibits the secretion of cytokines by dendritic cells via the TLR7/8 pathway in an imiquimod-induced psoriasis mouse model and in vitro. Life Sci. 2018, 207, 90–104. [Google Scholar] [CrossRef]
- Shamraiz, U.; Hussain, H.; Ur Rehman, N.; Al-Shidhani, S.; Saeed, A.; Khan, H.Y.; Khan, A.; Fischer, L.; Csuk, R.; Badshah, A.; et al. Synthesis of new boswellic acid derivatives as potential antiproliferative agents. Nat. Prod. Res. 2020, 34, 1845–1852. [Google Scholar] [CrossRef]
- Zhao, W.; Entschladen, F.; Liu, H.; Niggemann, B.; Fang, Q.; Zaenker, K.S.; Han, R. Boswellic acid acetate induces differentiation and apoptosis in highly metastatic melanoma and fibrosarcoma cells. Cancer Detect. Prev. 2003, 27, 67–75. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis: An expanding therapeutic pipeline for a complex disease. Nat. Rev. Drug Discov. 2022, 21, 21–40. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z.; Zhang, H.; Guo, Y.; Yao, Z. Update on the Pathogenesis and Therapy of Atopic Dermatitis. Clin. Rev. Allergy Immunol. 2021, 61, 324–338. [Google Scholar] [CrossRef]
- Wollenberg, A.; Weidinger, S.; Worm, M.; Bieber, T. Tralokinumab in atopic dermatitis. J. Dtsch Dermatol Ges. 2021, 19, 1435–1442. [Google Scholar] [CrossRef]
- Kwon, D.-J.; Bae, Y.-S.; Ju, S.M.; Goh, A.R.; Youn, G.S.; Choi, S.Y.; Park, J. Casuarinin suppresses TARC/CCL17 and MDC/CCL22 production via blockade of NF-κB and STAT1 activation in HaCaT cells. Biochem. Biophys. Res. Commun. 2012, 417, 1254–1259. [Google Scholar] [CrossRef]
- Čepelak, I.; Dodig, S.; Pavić, I. Filaggrin and atopic march. Biochem. Med. 2019, 29, 20501. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Kadono, T. "Inflammatory skin march" in atopic dermatitis and psoriasis. Inflamm. Res. 2017, 66, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Axon, E.; Chalmers, J.R.; Santer, M.; Ridd, M.J.; Lawton, S.; Langan, S.M.; Grindlay, D.J.C.; Muller, I.; Roberts, A.; Ahmed, A.; et al. Safety of topical corticosteroids in atopic eczema: An umbrella review. BMJ Open 2021, 11, e046476. [Google Scholar] [CrossRef] [PubMed]
- Moyle, M.; Cevikbas, F.; Harden, J.L.; Guttman-Yassky, E. Understanding the immune landscape in atopic dermatitis: The era of biologics and emerging therapeutic approaches. Exp. Derm. 2019, 28, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Buyanravjikh, S.; Han, S.; Lee, S.; Jeong, A.L.; Ka, H.I.; Park, J.Y.; Boldbaatar, A.; Lim, J.S.; Lee, M.S.; Yang, Y. Cryptotanshinone inhibits IgE-mediated degranulation through inhibition of spleen tyrosine kinase and tyrosine-protein kinase phosphorylation in mast cells. Mol. Med. Rep. 2018, 18, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Jeon, W.J.; Lee, J.H.; Lee, J.E.; Son, S.E.; Okajima, F.; Chung, K.W.; Im, D.S. Salvianolic Acid A Suppresses DNCB-Induced Atopic Dermatitis-Like Symptoms in BALB/c Mice. Evid. Based Complement. Alternat. Med. 2021, 2021, 7902592. [Google Scholar] [CrossRef]
- Yang, C.C.; Hung, Y.L.; Li, H.J.; Lin, Y.F.; Wang, S.J.; Chang, D.C.; Pu, C.M.; Hung, C.F. Quercetin inhibits histamine-induced calcium influx in human keratinocyte via histamine H4 receptors. Int. Immunopharmacol. 2021, 96, 107620. [Google Scholar] [CrossRef]
- Chiu, K.M.; Hung, Y.L.; Wang, S.J.; Tsai, Y.J.; Wu, N.L.; Liang, C.W.; Chang, D.C.; Hung, C.F. Anti-Allergic and Anti-Inflammatory Effects of Neferine on RBL-2H3 Cells. Int. J. Mol. Sci. 2021, 22, 10944. [Google Scholar] [CrossRef]
- Yang, C.C.; Hung, Y.L.; Ko, W.C.; Tsai, Y.J.; Chang, J.F.; Liang, C.W.; Chang, D.C.; Hung, C.F. Effect of Neferine on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Int. J. Mol. Sci. 2021, 22, 8237. [Google Scholar] [CrossRef]
- Wang, C.-C.; Hsiao, C.-Y.; Hsu, Y.-J.; Ko, H.-H.; Chang, D.-C.; Hung, C.-F. Anti-Inflammatory Effects of Cycloheterophyllin on Dinitrochlorobenzene-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Molecules 2022, 27, 2610. [Google Scholar] [CrossRef]
- Silverberg, N.B.; Silverberg, J.I. Inside out or outside in: Does atopic dermatitis disrupt barrier function or does disruption of barrier function trigger atopic dermatitis? Cutis 2015, 96, 359–361. [Google Scholar]
- Leung, D.Y.M.; Berdyshev, E.; Goleva, E. Cutaneous barrier dysfunction in allergic diseases. J. Allergy Clin. Immunol. 2020, 145, 1485–1497. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef]
- Salvati, L.; Cosmi, L.; Annunziato, F. From Emollients to Biologicals: Targeting Atopic Dermatitis. Int. J. Mol. Sci. 2021, 22, 10381. [Google Scholar] [CrossRef]
- Lee, J.H.; Lim, J.Y.; Jo, E.H.; Noh, H.M.; Park, S.; Park, M.C.; Kim, D.K. Chijabyukpi-Tang Inhibits Pro-Inflammatory Cytokines and Chemokines via the Nrf2/HO-1 Signaling Pathway in TNF-α/IFN-γ-Stimulated HaCaT Cells and Ameliorates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in Mice. Front. Pharm. 2020, 11, 1018. [Google Scholar] [CrossRef]
- Tüzün, Y.; Antonov, M.; Dolar, N.; Wolf, R. Keratinocyte cytokine and chemokine receptors. Dermatol Clin. 2007, 25, 467–476. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis. Ann. Derm. 2010, 22, 125–137. [Google Scholar] [CrossRef]
- Lee, H.S.; Park, J.W.; Kwon, O.K.; Lim, Y.; Kim, J.H.; Kim, S.Y.; Zamora, N.; Rosales, K.; Choi, S.; Oh, S.R.; et al. Anti-inflammatory effects of ethanol extract from the leaves and shoots of Cedrela odorata L. in cytokine-stimulated keratinocytes. Exp. Ther. Med. 2019, 18, 833–840. [Google Scholar] [CrossRef]
- Lim, J.S.; Kim, J.Y.; Lee, S.; Choi, J.K.; Kim, E.N.; Choi, Y.A.; Jang, Y.H.; Jeong, G.S.; Kim, S.H. Bakuchicin attenuates atopic skin inflammation. Biomed. Pharm. 2020, 129, 110466. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.H. STAT signaling in inflammation. Jakstat 2013, 2, e24198. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A.; Nf-kappa, B. Chemokine Gene Transcription and Tumour Growth. Nat. Rev. Immunol. 2002, 2, 664–674. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, Y.-C.; Chang, H.-H.; Chou, S.-C.; Chu, T.W.; Hsu, Y.-J.; Hsiao, C.-Y.; Lo, Y.-H.; Wu, N.-L.; Chang, D.-C.; Hung, C.-F. Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice. Int. J. Mol. Sci. 2022, 23, 9863. https://doi.org/10.3390/ijms23179863
Tsai Y-C, Chang H-H, Chou S-C, Chu TW, Hsu Y-J, Hsiao C-Y, Lo Y-H, Wu N-L, Chang D-C, Hung C-F. Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice. International Journal of Molecular Sciences. 2022; 23(17):9863. https://doi.org/10.3390/ijms23179863
Chicago/Turabian StyleTsai, Ya-Chu, Hsun-Hao Chang, Sheng-Chieh Chou, Thomas W. Chu, Yu-Jou Hsu, Chien-Yu Hsiao, Yuan-Hsin Lo, Nan-Lin Wu, Der-Chen Chang, and Chi-Feng Hung. 2022. "Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice" International Journal of Molecular Sciences 23, no. 17: 9863. https://doi.org/10.3390/ijms23179863
APA StyleTsai, Y.-C., Chang, H.-H., Chou, S.-C., Chu, T. W., Hsu, Y.-J., Hsiao, C.-Y., Lo, Y.-H., Wu, N.-L., Chang, D.-C., & Hung, C.-F. (2022). Evaluation of the Anti-Atopic Dermatitis Effects of α-Boswellic Acid on Tnf-α/Ifn-γ-Stimulated HaCat Cells and DNCB-Induced BALB/c Mice. International Journal of Molecular Sciences, 23(17), 9863. https://doi.org/10.3390/ijms23179863