
High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
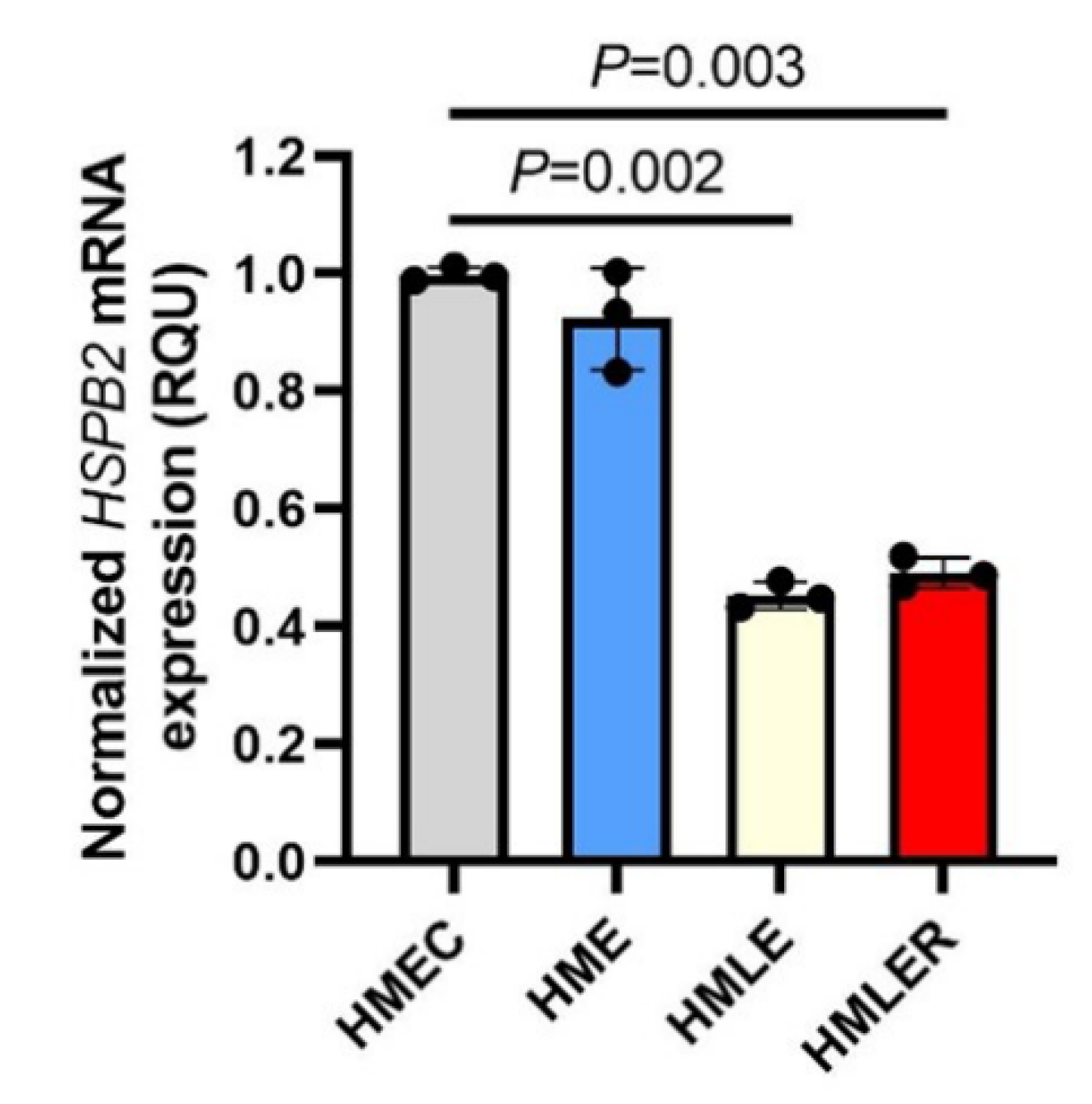
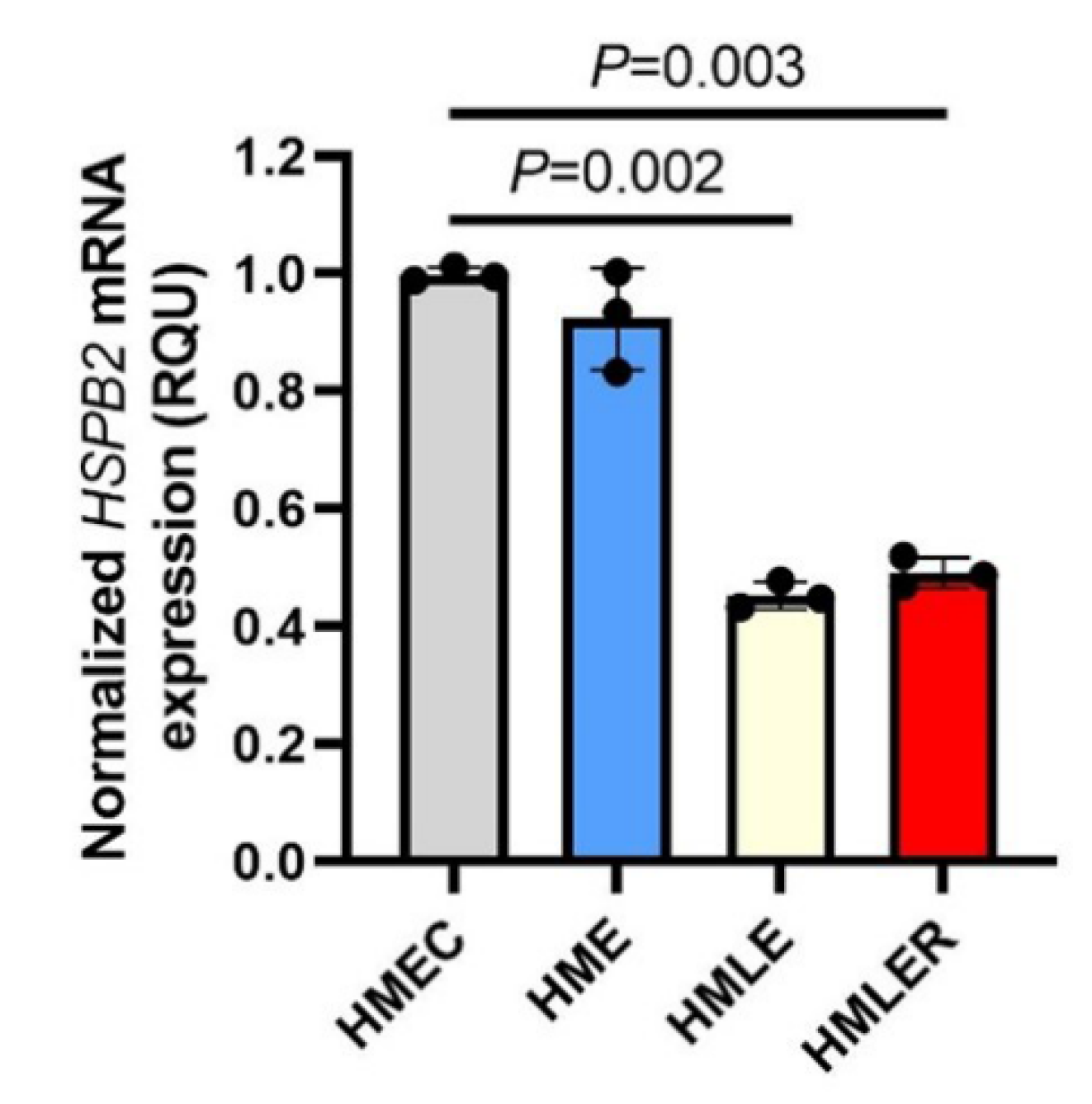
2.1. Downregulation of HSPB2 Gene Expression Levels during Oncogenic Transformation in Mammary Epithelial Cells
2.2. Clinicopathological and Biological Characteristics of BrCa Patients and Used Samples
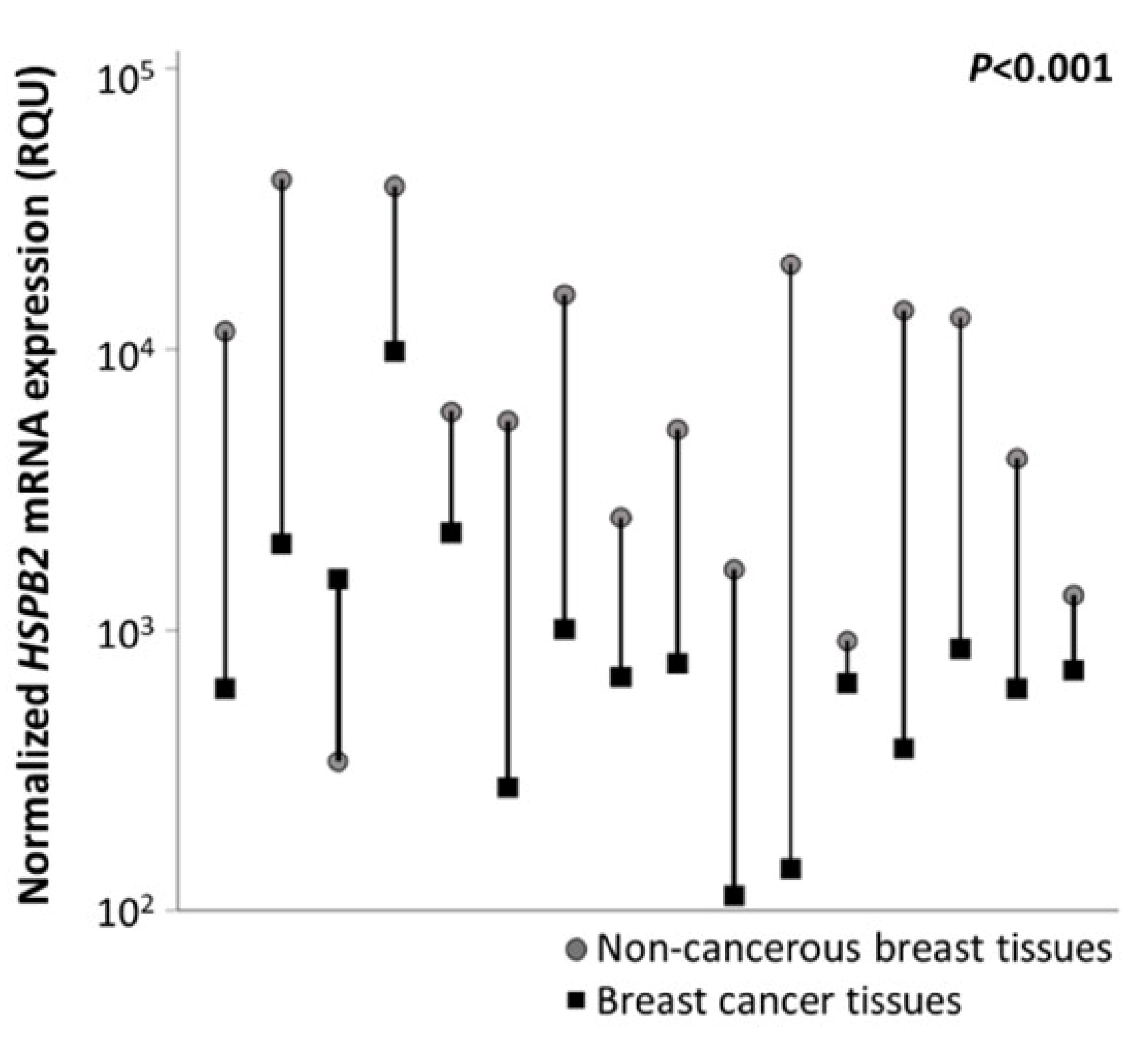
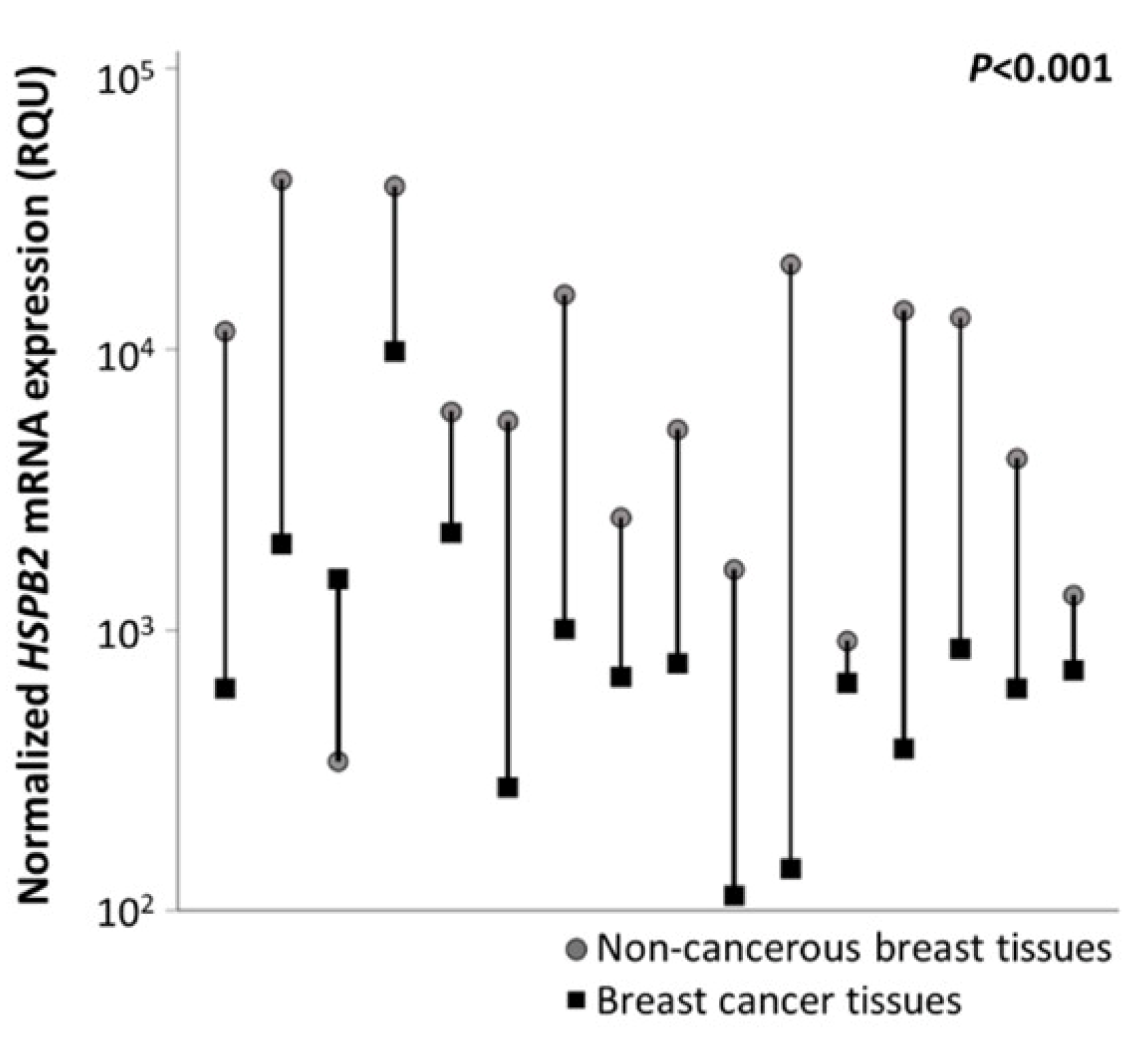
2.3. Reduced Expression Levels of HSPB2 mRNA in Breast Carcinoma Tissues as Compared with Paired Non-Cancerous Tissues
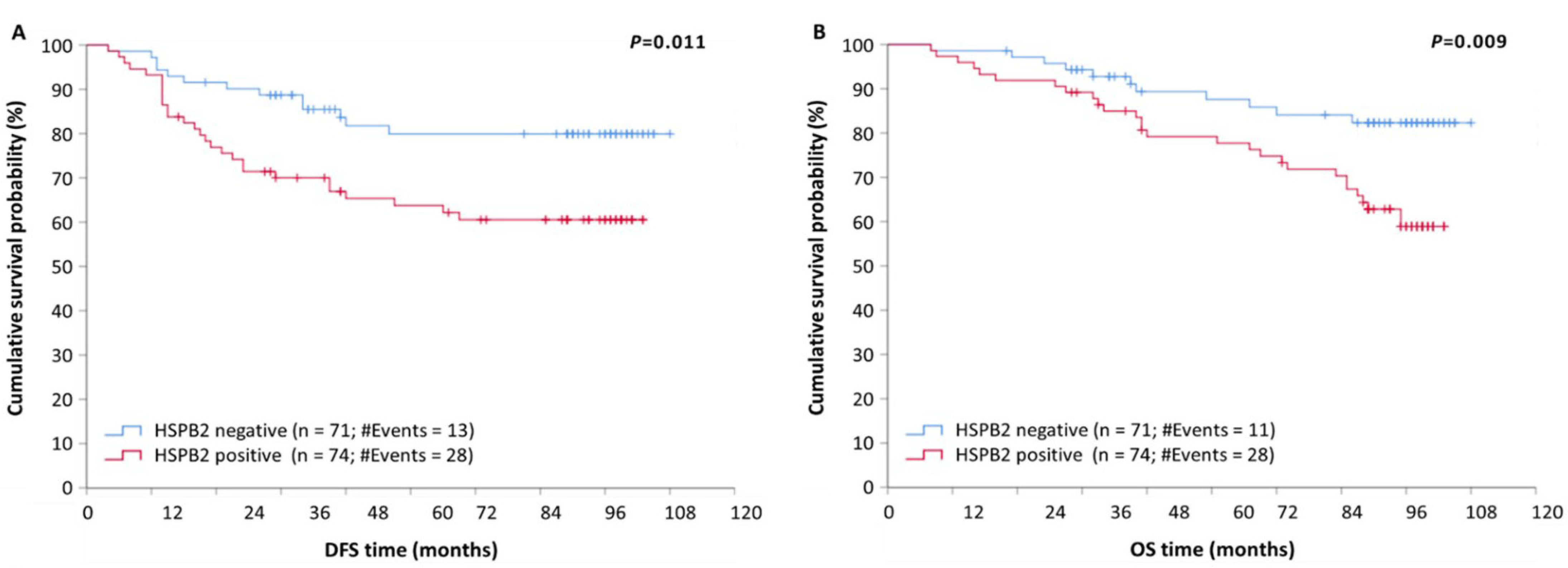
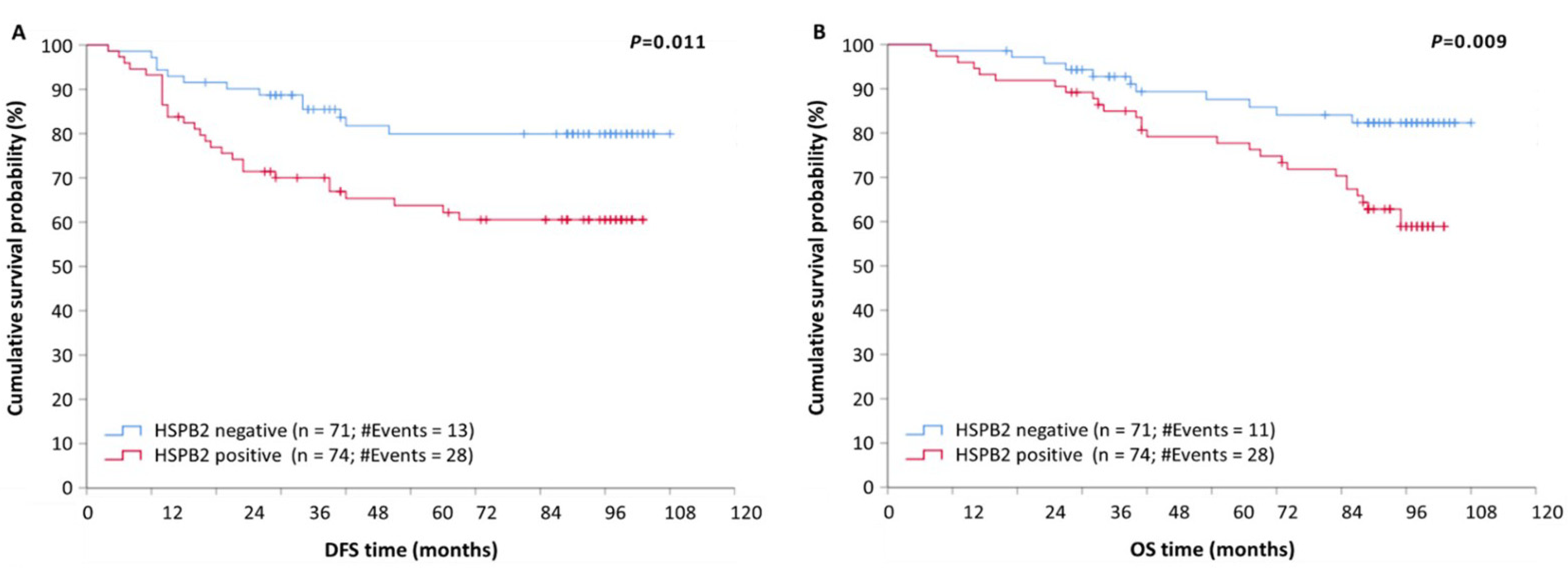
2.4. HSPB2 mRNA Overexpression Is a Reliable Predictor of Poor Prognosis in BrCa Patients, Independent of Other Clinicopathological Factors
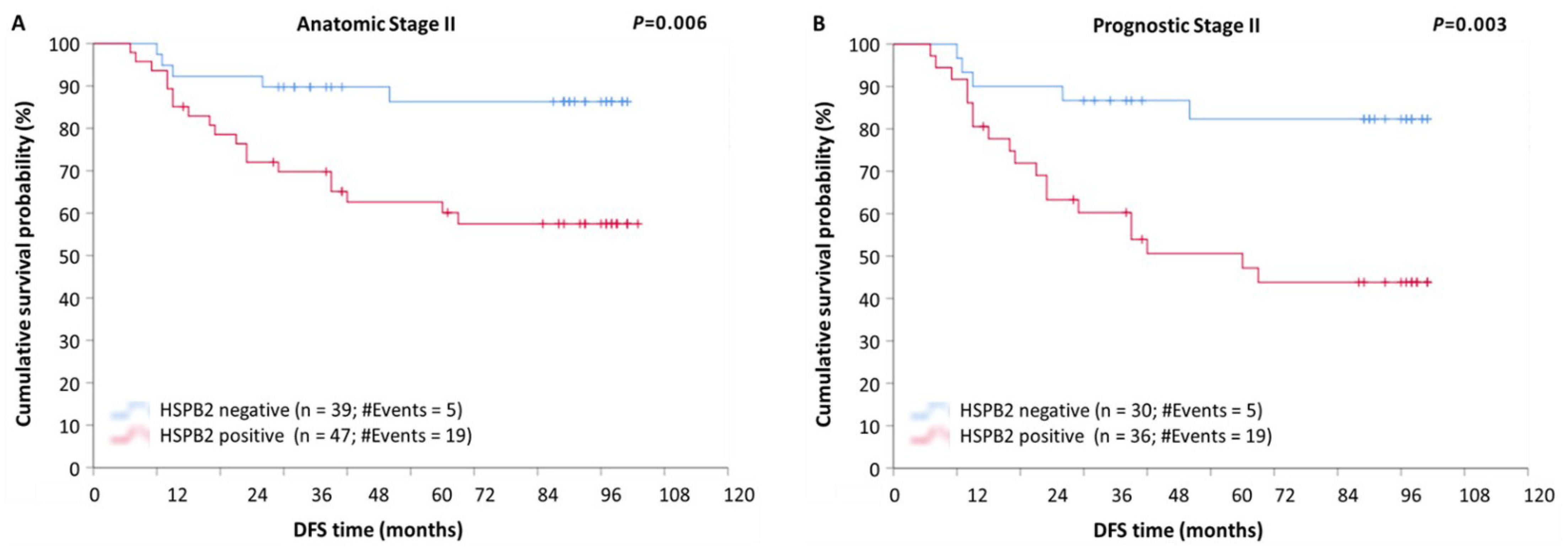
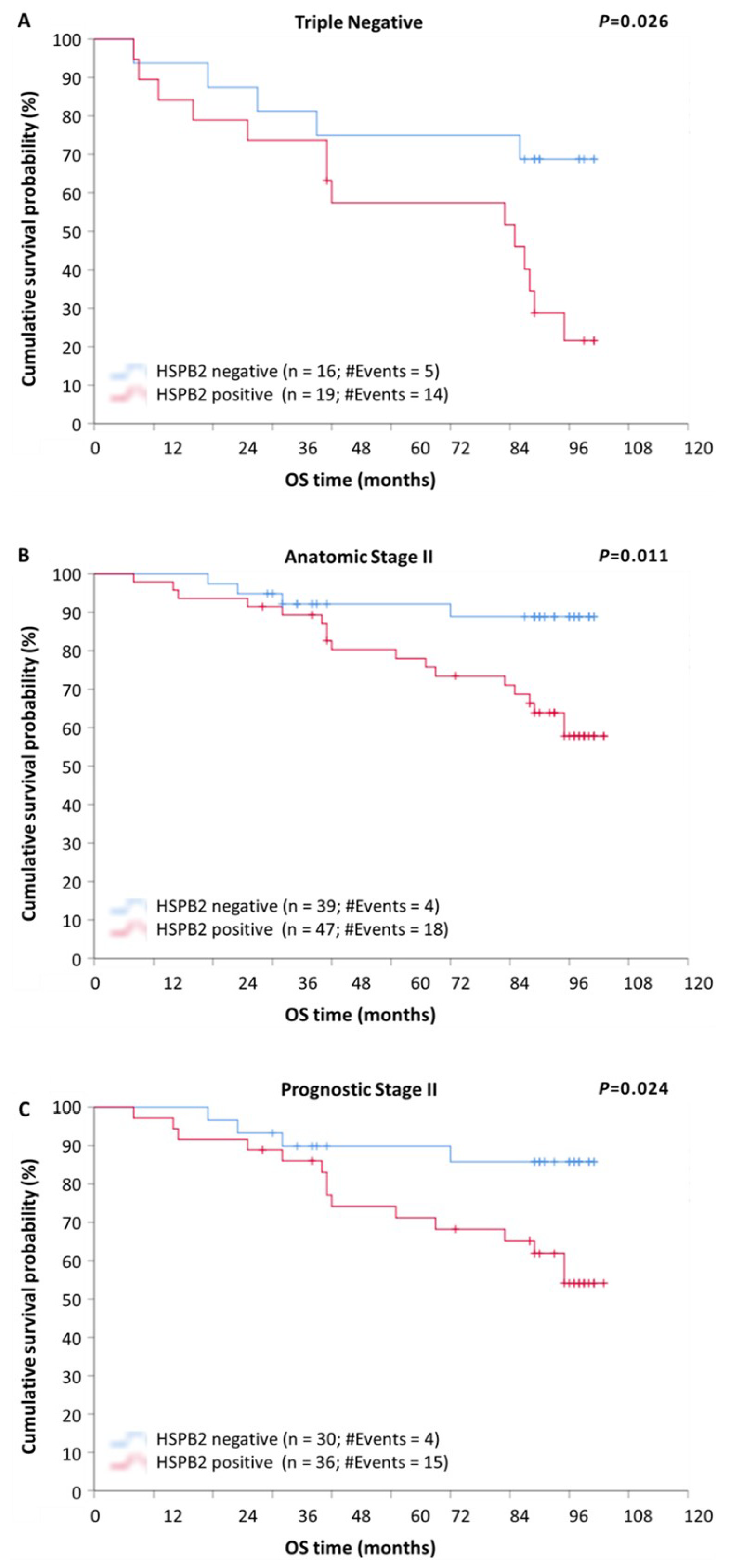
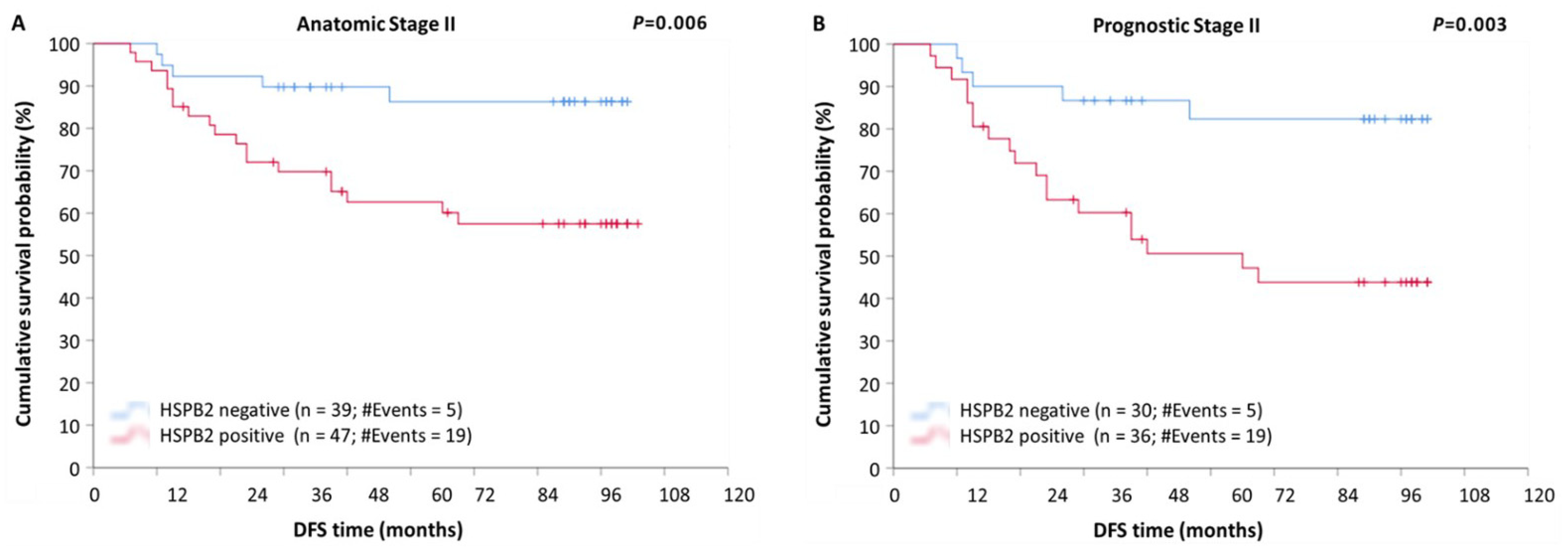
2.5. Prognostic Value of HSPB2 mRNA Expression in Patients with Breast Adenocarcinoma, Stratified According to Molecular Subtype, Anatomic Stage, or Prognostic Stage
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture Conditions
4.2. Patients and Tissue Collection
4.3. Total RNA Extraction and First-Strand cDNA Synthesis
4.4. Quantitative Real-Time PCR (qPCR)
4.5. Biostatistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Lukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanislawek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast cancer. Nat. Rev. Dis. Prim. 2019, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Riggio, A.I.; Varley, K.E.; Welm, A.L. The lingering mysteries of metastatic recurrence in breast cancer. Br. J. Cancer 2021, 124, 13–26. [Google Scholar] [CrossRef]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Loibl, S.; Poortmans, P.; Morrow, M.; Denkert, C.; Curigliano, G. Breast cancer. Lancet 2021, 397, 1750–1769. [Google Scholar] [CrossRef]
- Jolly, C.; Morimoto, R.I. Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J. Natl. Cancer Inst. 2000, 92, 1564–1572. [Google Scholar] [CrossRef]
- Niforou, K.; Cheimonidou, C.; Trougakos, I.P. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol. 2014, 2, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Freilich, R.; Arhar, T.; Abrams, J.L.; Gestwicki, J.E. Protein-Protein Interactions in the Molecular Chaperone Network. Acc Chem. Res. 2018, 51, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, B.; Cristofani, R.; Ferrari, V.; Cozzi, M.; Rusmini, P.; Casarotto, E.; Chierichetti, M.; Mina, F.; Galbiati, M.; Piccolella, M.; et al. Insights on Human Small Heat Shock Proteins and Their Alterations in Diseases. Front. Mol. Biosci. 2022, 9, 842149. [Google Scholar] [CrossRef] [PubMed]
- Treweek, T.M.; Meehan, S.; Ecroyd, H.; Carver, J.A. Small heat-shock proteins: Important players in regulating cellular proteostasis. Cell. Mol. Life Sci. 2015, 72, 429–451. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.J.; Bott, L.C.; Morimoto, R.I. Shaping proteostasis at the cellular, tissue, and organismal level. J. Cell Biol. 2017, 216, 1231–1241. [Google Scholar] [CrossRef]
- Artemaki, P.I.; Sklirou, A.D.; Kontos, C.K.; Liosi, A.A.; Gianniou, D.D.; Papadopoulos, I.N.; Trougakos, I.P.; Scorilas, A. High clusterin (CLU) mRNA expression levels in tumors of colorectal cancer patients predict a poor prognostic outcome. Clin. Biochem. 2020, 75, 62–69. [Google Scholar] [CrossRef]
- Cyran, A.M.; Zhitkovich, A. Heat Shock Proteins and HSF1 in Cancer. Front. Oncol. 2022, 12, 860320. [Google Scholar] [CrossRef]
- Kalioraki, M.A.; Artemaki, P.I.; Sklirou, A.D.; Kontos, C.K.; Adamopoulos, P.G.; Papadopoulos, I.N.; Trougakos, I.P.; Scorilas, A. Heat shock protein beta 3 (HSPB3) is an unfavorable molecular biomarker in colorectal adenocarcinoma. Mol. Carcinog. 2020, 59, 116–125. [Google Scholar] [CrossRef]
- Sheng, B.; Qi, C.; Liu, B.; Lin, Y.; Fu, T.; Zeng, Q. Increased HSP27 correlates with malignant biological behavior of non-small cell lung cancer and predicts patient’s survival. Sci. Rep. 2017, 7, 13807. [Google Scholar] [CrossRef]
- Yun, C.W.; Kim, H.J.; Lim, J.H.; Lee, S.H. Heat Shock Proteins: Agents of Cancer Development and Therapeutic Targets in Anti-Cancer Therapy. Cells 2019, 9, 60. [Google Scholar] [CrossRef]
- Zhang, Z.; Jing, J.; Ye, Y.; Chen, Z.; Jing, Y.; Li, S.; Hong, W.; Ruan, H.; Liu, Y.; Hu, Q.; et al. Characterization of the dual functional effects of heat shock proteins (HSPs) in cancer hallmarks to aid development of HSP inhibitors. Genome Med. 2020, 12, 101. [Google Scholar] [CrossRef]
- Bakthisaran, R.; Tangirala, R.; Rao Ch, M. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta 2015, 1854, 291–319. [Google Scholar] [CrossRef]
- Janowska, M.K.; Baughman, H.E.R.; Woods, C.N.; Klevit, R.E. Mechanisms of Small Heat Shock Proteins. Cold Spring Harb. Perspect Biol. 2019, 11, a034025. [Google Scholar] [CrossRef] [PubMed]
- Webster, J.M.; Darling, A.L.; Uversky, V.N.; Blair, L.J. Small Heat Shock Proteins, Big Impact on Protein Aggregation in Neurodegenerative Disease. Front. Pharmacol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Sugiyama, Y.; Hayashi, Y.; Nyu-i, N.; Yoshida, M.; Nonaka, I.; Ishiura, S.; Arahata, K.; Ohno, S. MKBP, a novel member of the small heat shock protein family, binds and activates the myotonic dystrophy protein kinase. J. Cell Biol. 1998, 140, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Li, Y.; Tan, X.; Fu, L. Small Heat Shock Proteins in Cancers: Functions and Therapeutic Potential for Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 6611. [Google Scholar] [CrossRef]
- Oshita, S.E.; Chen, F.; Kwan, T.; Yehiely, F.; Cryns, V.L. The small heat shock protein HspB2 is a novel anti-apoptotic protein that inhibits apical caspase activation in the extrinsic apoptotic pathway. Breast Cancer Res. Treat. 2010, 124, 307–315. [Google Scholar] [CrossRef]
- Elenbaas, B.; Spirio, L.; Koerner, F.; Fleming, M.D.; Zimonjic, D.B.; Donaher, J.L.; Popescu, N.C.; Hahn, W.C.; Weinberg, R.A. Human breast cancer cells generated by oncogenic transformation of primary mammary epithelial cells. Genes Dev. 2001, 15, 50–65. [Google Scholar] [CrossRef]
- Colzani, E.; Liljegren, A.; Johansson, A.L.; Adolfsson, J.; Hellborg, H.; Hall, P.F.; Czene, K. Prognosis of patients with breast cancer: Causes of death and effects of time since diagnosis, age, and tumor characteristics. J. Clin. Oncol. 2011, 29, 4014–4021. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Kim, Y.E.; Hipp, M.S.; Bracher, A.; Hayer-Hartl, M.; Hartl, F.U. Molecular chaperone functions in protein folding and proteostasis. Annu. Rev. Biochem. 2013, 82, 323–355. [Google Scholar] [CrossRef]
- Bakthisaran, R.; Akula, K.K.; Tangirala, R.; Rao Ch, M. Phosphorylation of alphaB-crystallin: Role in stress, aging and patho-physiological conditions. Biochim. Biophys. Acta 2016, 1860, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Carra, S.; Alberti, S.; Arrigo, P.A.; Benesch, J.L.; Benjamin, I.J.; Boelens, W.; Bartelt-Kirbach, B.; Brundel, B.; Buchner, J.; Bukau, B.; et al. The growing world of small heat shock proteins: From structure to functions. Cell Stress Chaperones 2017, 22, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Shevtsov, M.; Balogi, Z.; Khachatryan, W.; Gao, H.; Vigh, L.; Multhoff, G. Membrane-Associated Heat Shock Proteins in Oncology: From Basic Research to New Theranostic Targets. Cells 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell. 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Luo, J.; Solimini, N.L.; Elledge, S.J. Principles of cancer therapy: Oncogene and non-oncogene addiction. Cell 2009, 136, 823–837. [Google Scholar] [CrossRef]
- Sklirou, A.; Papanagnou, E.D.; Fokialakis, N.; Trougakos, I.P. Cancer chemoprevention via activation of proteostatic modules. Cancer Lett. 2018, 413, 110–121. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Breast Cancer: A Molecularly Heterogenous Disease Needing Subtype-Specific Treatments. Med. Sci. 2020, 8, 18. [Google Scholar] [CrossRef]
- Rugo, H.S.; Rumble, R.B.; Macrae, E.; Barton, D.L.; Connolly, H.K.; Dickler, M.N.; Fallowfield, L.; Fowble, B.; Ingle, J.N.; Jahanzeb, M.; et al. Endocrine Therapy for Hormone Receptor-Positive Metastatic Breast Cancer: American Society of Clinical Oncology Guideline. J. Clin. Oncol. 2016, 34, 3069–3103. [Google Scholar] [CrossRef]
- Wang, J.; Xu, B. Targeted therapeutic options and future perspectives for HER2-positive breast cancer. Signal Transduct. Target. Ther. 2019, 4, 34. [Google Scholar] [CrossRef]
- Afzal, S.; Hassan, M.; Ullah, S.; Abbas, H.; Tawakkal, F.; Khan, M.A. Breast Cancer; Discovery of Novel Diagnostic Biomarkers, Drug Resistance, and Therapeutic Implications. Front. Mol. Biosci. 2022, 9, 783450. [Google Scholar] [CrossRef]
- Wu, H.J.; Chu, P.Y. Recent Discoveries of Macromolecule- and Cell-Based Biomarkers and Therapeutic Implications in Breast Cancer. Int. J. Mol. Sci. 2021, 22, 636. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Marino Gammazza, A.; Campanella, C.; Bucchieri, F.; Cappello, F. Extracellular heat shock proteins in cancer: From early diagnosis to new therapeutic approach. Semin. Cancer Biol. 2021; accepted. [Google Scholar] [CrossRef] [PubMed]
- Lianos, G.D.; Alexiou, G.A.; Mangano, A.; Mangano, A.; Rausei, S.; Boni, L.; Dionigi, G.; Roukos, D.H. The role of heat shock proteins in cancer. Cancer Lett. 2015, 360, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xiao, H.; Cao, L. Recent advances in heat shock proteins in cancer diagnosis, prognosis, metabolism and treatment. Biomed. Pharmacother. 2021, 142, 112074. [Google Scholar] [CrossRef]
- Ciocca, D.R.; Calderwood, S.K. Heat shock proteins in cancer: Diagnostic, prognostic, predictive, and treatment implications. Cell Stress Chaperones 2005, 10, 86–103. [Google Scholar] [CrossRef]
- Zoppino, F.C.M.; Guerrero-Gimenez, M.E.; Castro, G.N.; Ciocca, D.R. Comprehensive transcriptomic analysis of heat shock proteins in the molecular subtypes of human breast cancer. BMC Cancer 2018, 18, 700. [Google Scholar] [CrossRef] [PubMed]
- Salhia, B.; Kiefer, J.; Ross, J.T.; Metapally, R.; Martinez, R.A.; Johnson, K.N.; DiPerna, D.M.; Paquette, K.M.; Jung, S.; Nasser, S.; et al. Integrated genomic and epigenomic analysis of breast cancer brain metastasis. PLoS ONE 2014, 9, e85448. [Google Scholar] [CrossRef]
- Chang, X.; Yamashita, K.; Sidransky, D.; Kim, M.S. Promoter methylation of heat shock protein B2 in human esophageal squamous cell carcinoma. Int. J. Oncol. 2011, 38, 1129–1135. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, H.; Fang, Y.; Lu, L.; Li, M.; Yan, B.; Nie, Y.; Teng, C. Molecular chaperone HspB2 inhibited pancreatic cancer cell proliferation via activating p53 downstream gene RPRM, BAI1, and TSAP6. J. Cell Biochem. 2020, 121, 2318–2329. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Fonseca, N.A.; Keays, M.; Tang, Y.A.; Barrera, E.; Bazant, W.; Burke, M.; Füllgrabe, A.; Fuentes, A.M.-P.; George, N.; et al. Expression Atlas: Gene and protein expression across multiple studies and organisms. Nucleic Acids Res. 2018, 46, D246–D251. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Khaleque, M.A.; Sawyer, D.B.; Ciocca, D.R. Heat shock proteins in cancer: Chaperones of tumorigenesis. Trends Biochem. Sci. 2006, 31, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Burns, T.F. Targeting Heat Shock Proteins in Cancer: A Promising Therapeutic Approach. Int. J. Mol. Sci. 2017, 18, 1978. [Google Scholar] [CrossRef] [PubMed]
- Zoubeidi, A.; Gleave, M. Small heat shock proteins in cancer therapy and prognosis. Int. J. Biochem. Cell Biol. 2012, 44, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Labbadia, J.; Morimoto, R.I. The biology of proteostasis in aging and disease. Annu. Rev. Biochem. 2015, 84, 435–464. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Cao, P.; Fan, Y.; Tan, K. Emerging roles of HSF1 in cancer: Cellular and molecular episodes. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188390. [Google Scholar] [CrossRef]
- Mendillo, M.L.; Santagata, S.; Koeva, M.; Bell, G.W.; Hu, R.; Tamimi, R.M.; Fraenkel, E.; Ince, T.A.; Whitesell, L.; Lindquist, S. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. Cell 2012, 150, 549–562. [Google Scholar] [CrossRef]
- Santagata, S.; Hu, R.; Lin, N.U.; Mendillo, M.L.; Collins, L.C.; Hankinson, S.E.; Schnitt, S.J.; Whitesell, L.; Tamimi, R.M.; Lindquist, S.; et al. High levels of nuclear heat-shock factor 1 (HSF1) are associated with poor prognosis in breast cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 18378–18383. [Google Scholar] [CrossRef]
- Yu, W.; Wang, J.; Li, C.; Xuan, M.; Han, S.; Zhang, Y.; Liu, P.; Zhao, Z. miR-17-5p promotes the invasion and migration of colorectal cancer by regulating HSPB2. J. Cancer 2022, 13, 918–931. [Google Scholar] [CrossRef]
- Calderwood, S.K.; Gong, J. Heat Shock Proteins Promote Cancer: It’s a Protection Racket. Trends Biochem. Sci. 2016, 41, 311–323. [Google Scholar] [CrossRef]
- Jego, G.; Hazoume, A.; Seigneuric, R.; Garrido, C. Targeting heat shock proteins in cancer. Cancer Lett. 2013, 332, 275–285. [Google Scholar] [CrossRef]
- Chang, H.R.; Jung, E.; Cho, S.; Jeon, Y.J.; Kim, Y. Targeting Non-Oncogene Addiction for Cancer Therapy. Biomolecules 2021, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; Semenova, E.A.; Berns, A. Drugging the addict: Non-oncogene addiction as a target for cancer therapy. EMBO Rep. 2016, 17, 1516–1531. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, M.; Zhou, J.; Zhang, X. HSP27, 70 and 90, anti-apoptotic proteins, in clinical cancer therapy (Review). Int. J. Oncol. 2014, 45, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, T.; Rios, Z.; Mei, Q.; Lin, X.; Cao, S. Heat Shock Proteins and Cancer. Trends Pharmacol. Sci. 2017, 38, 226–256. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Cortes, J. Breast cancer and HSP90 inhibitors: Is there a role beyond the HER2-positive subtype? Breast 2012, 21, 604–607. [Google Scholar] [CrossRef]
- Modi, S.; Stopeck, A.; Linden, H.; Solit, D.; Chandarlapaty, S.; Rosen, N.; D’Andrea, G.; Dickler, M.; Moynahan, M.E.; Sugarman, S.; et al. HSP90 inhibition is effective in breast cancer: A phase II trial of tanespimycin (17-AAG) plus trastuzumab in patients with HER2-positive metastatic breast cancer progressing on trastuzumab. Clin. Cancer Res. 2011, 17, 5132–5139. [Google Scholar] [CrossRef]
- Park, J.M.; Kim, Y.J.; Park, S.; Park, M.; Farrand, L.; Nguyen, C.T.; Ann, J.; Nam, G.; Park, H.J.; Lee, J.; et al. A novel HSP90 inhibitor targeting the C-terminal domain attenuates trastuzumab resistance in HER2-positive breast cancer. Mol. Cancer 2020, 19, 161. [Google Scholar] [CrossRef]
- Henry, N.L.; Hayes, D.F. Cancer biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef]
- Kamel, H.F.M.; Al-Amodi, H. Exploitation of Gene Expression and Cancer Biomarkers in Paving the Path to Era of Personalized Medicine. Genom. Proteom. Bioinform. 2017, 15, 220–235. [Google Scholar] [CrossRef]
- Vargas, A.J.; Harris, C.C. Biomarker development in the precision medicine era: Lung cancer as a case study. Nat. Rev. Cancer 2016, 16, 525–537. [Google Scholar] [CrossRef]
- de Kok, J.B.; Roelofs, R.W.; Giesendorf, B.A.; Pennings, J.L.; Waas, E.T.; Feuth, T.; Swinkels, D.W.; Span, P.N. Normalization of gene expression measurements in tumor tissues: Comparison of 13 endogenous control genes. Lab. Investig. 2005, 85, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Patients (%) | |
|---|---|
| Patients | 150 |
| HSPB2 mRNA Expression Status | |
| Negative | 75 (50.0%) |
| Positive | 75 (50.0%) |
| Age (y) | Median: 60; Range: 31–90 |
| Tumor Size (cm2) | Median: 2.35; Range: (0.60–8.50) |
| Cancer Type | |
| Invasive ductal carcinoma | 121 (80.7%) |
| Lobular carcinoma | 16 (10.7%) |
| Other carcinomas | 13 (8.6%) |
| Molecular Subtype | |
| Luminal A | 59 (39.3%) |
| Luminal B | 40 (26.7%) |
| Triple-negative | 36 (24.0%) |
| HER2-enriched | 15 (10.0%) |
| Histological Grade | |
| I | 8 (5.3%) |
| II | 97 (64.7%) |
| III | 45 (30.0%) |
| HER2 Status | |
| Negative | 115 (79.3%) |
| Positive | 30 (20.7%) |
| Unknown | 5 |
| ER Status | |
| Negative | 56 (37.3%) |
| Positive | 94 (62.7%) |
| PR Status | |
| Negative | 82 (54.7%) |
| Positive | 68 (45.3%) |
| Ki-67 Index | |
| Low (≤14%) | 92 (63.9%) |
| High (>14%) | 52 (36.1%) |
| Unknown | 6 |
| Anatomic Stage | |
| I | 43 (28.7%) |
| II | 89 (59.3%) |
| III | 18 (12.0%) |
| Prognostic Stage | |
| I | 60 (40.0%) |
| II | 68 (45.3%) |
| III | 22 (14.7%) |
| Variable | Mean ± SE | Range | Percentiles | ||
|---|---|---|---|---|---|
| 25th | 50th (Median) | 75th | |||
| HSPB2 mRNA Expression (RQU) | |||||
| in breast cancer tissues (n = 150) | 2215.7 ± 265.8 | 29.0–19,101.0 | 315.5 | 656.5 | 2637.8 |
| in non-cancerous tissues (n = 16) | 11,234.5 ± 3094.54 | 340.0–40,093.0 | 1864.0 | 5778.5 | 15,170.3 |
| Univariate Analysis (n = 145) | Multivariable Analysis 1 (n = 145) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Covariate | HR 2 | 95% CI 3 | p Value 4 | BCa Bootstrap 95% CI 3 | Bootstrap p Value 4 | HR 2 | 95% CI 3 | p Value 4 | BCa Bootstrap 95% CI 3 | Bootstrap p Value 4 |
| HSPB2 mRNA expression status | ||||||||||
| Negative (n = 71) | 1.00 | 1.00 | ||||||||
| Positive (n = 74) | 2.29 | 1.19–4.42 | 0.014 | 1.17–4.82 | 0.014 | 2.61 | 1.34–5.08 | 0.005 | 1.26–6.61 | 0.006 |
| Molecular subtype | <0.001 | 0.071 | ||||||||
| Luminal A (n = 56) | 1.00 | 1.00 | ||||||||
| Luminal B (n = 39) | 1.02 | 0.36–2.88 | 0.97 | 0.31–3.11 | 0.97 | 1.27 | 0.45–3.60 | 0.65 | 0.24–4.65 | 0.66 |
| Triple-negative (n = 35) | 3.63 | 1.60–8.21 | 0.002 | 1.45–10.88 | 0.003 | 2.04 | 0.83–5.02 | 0.12 | 0.78–7.18 | 0.094 |
| HER2-enriched (n = 15) | 6.04 | 2.44–14.96 | <0.001 | 2.40–19.65 | <0.001 | 3.59 | 1.35–9.20 | 0.010 | 1.42–11.78 | 0.005 |
| Prognostic stage | <0.001 | 0.013 | ||||||||
| I (n = 58) | 1.00 | 1.00 | ||||||||
| II (n = 66) | 5.00 | 1.90–13.10 | 0.001 | 1.78–4.89 × 104 | 0.002 | 3.72 | 1.33–10.38 | 0.012 | 0.99–1.10 × 105 | 0.010 |
| III (n = 21) | 10.06 | 3.53–28.63 | <0.001 | 2.87–87.66 | <0.001 | 6.22 | 1.83–21.12 | 0.003 | 1.4–1.45 × 106 | 0.005 |
| Univariate Analysis (n = 145) | Multivariable Analysis 1 (n = 145) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Covariate | HR 2 | 95% CI 3 | p Value 4 | BCa Bootstrap 95% CI 3 | Bootstrap p Value 4 | HR 2 | 95% CI 3 | p Value 4 | BCa Bootstrap 95% CI 3 | Bootstrap p Value 4 |
| HSPB2 mRNA expression status | ||||||||||
| Negative (n = 71) | 1.00 | 1.00 | ||||||||
| Positive (n = 74) | 2.45 | 1.23–4.96 | 0.011 | 1.21–5.79 | 0.015 | 2.69 | 1.33–5.42 | 0.006 | 1.23–7.98 | 0.007 |
| Molecular subtype | <0.001 | 0.085 | ||||||||
| Luminal A (n = 56) | 1.00 | 1.00 | ||||||||
| Luminal B (n = 39) | 1.16 | 0.40–3.35 | 0.78 | 3.66–38.89 | 0.79 | 1.27 | 0.44–3.66 | 0.66 | 0.30–4.40 | 0.67 |
| Triple-negative (n = 35) | 4.66 | 2.04–10.66 | <0.001 | 15.17–3.89 × 106 | <0.001 | 3.22 | 1.26–8.23 | 0.014 | 1.26–10.23 | 0.008 |
| HER2-enriched (n = 15) | 3.54 | 1.23–10.22 | 0.019 | 12.50–2.69 × 105 | 0.011 | 2.45 | 0.79–7.59 | 0.12 | 0.47–10.17 | 0.14 |
| Prognostic stage | 0.16 | |||||||||
| I (n = 58) | 1.00 | 1.00 | ||||||||
| II (n = 66) | 2.26 | 0.99–5.16 | 0.053 | 0.96–6.20 | 0.046 | 1.62 | 0.67–3.92 | 0.29 | 0.64–4.88 | 0.23 |
| III (n = 21) | 6.00 | 2.44–14.71 | <0.001 | 2.29–17.55 | <0.001 | 2.86 | 0.96–8.53 | 0.059 | 0.82–15.54 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sklirou, A.D.; Gianniou, D.D.; Karousi, P.; Cheimonidi, C.; Papachristopoulou, G.; Kontos, C.K.; Scorilas, A.; Trougakos, I.P. High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival. Int. J. Mol. Sci. 2022, 23, 9758. https://doi.org/10.3390/ijms23179758
Sklirou AD, Gianniou DD, Karousi P, Cheimonidi C, Papachristopoulou G, Kontos CK, Scorilas A, Trougakos IP. High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival. International Journal of Molecular Sciences. 2022; 23(17):9758. https://doi.org/10.3390/ijms23179758
Chicago/Turabian StyleSklirou, Aimilia D., Despoina D. Gianniou, Paraskevi Karousi, Christina Cheimonidi, Georgia Papachristopoulou, Christos K. Kontos, Andreas Scorilas, and Ioannis P. Trougakos. 2022. "High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival" International Journal of Molecular Sciences 23, no. 17: 9758. https://doi.org/10.3390/ijms23179758
APA StyleSklirou, A. D., Gianniou, D. D., Karousi, P., Cheimonidi, C., Papachristopoulou, G., Kontos, C. K., Scorilas, A., & Trougakos, I. P. (2022). High mRNA Expression Levels of Heat Shock Protein Family B Member 2 (HSPB2) Are Associated with Breast Cancer Patients’ Relapse and Poor Survival. International Journal of Molecular Sciences, 23(17), 9758. https://doi.org/10.3390/ijms23179758