
Characterization in Effective Stimulation on the Magnitude, Gating, Frequency Dependence, and Hysteresis of INa Exerted by Picaridin (or Icaridin), a Known Insect Repellent

 , ,
, ,
Abstract
:1. Introduction
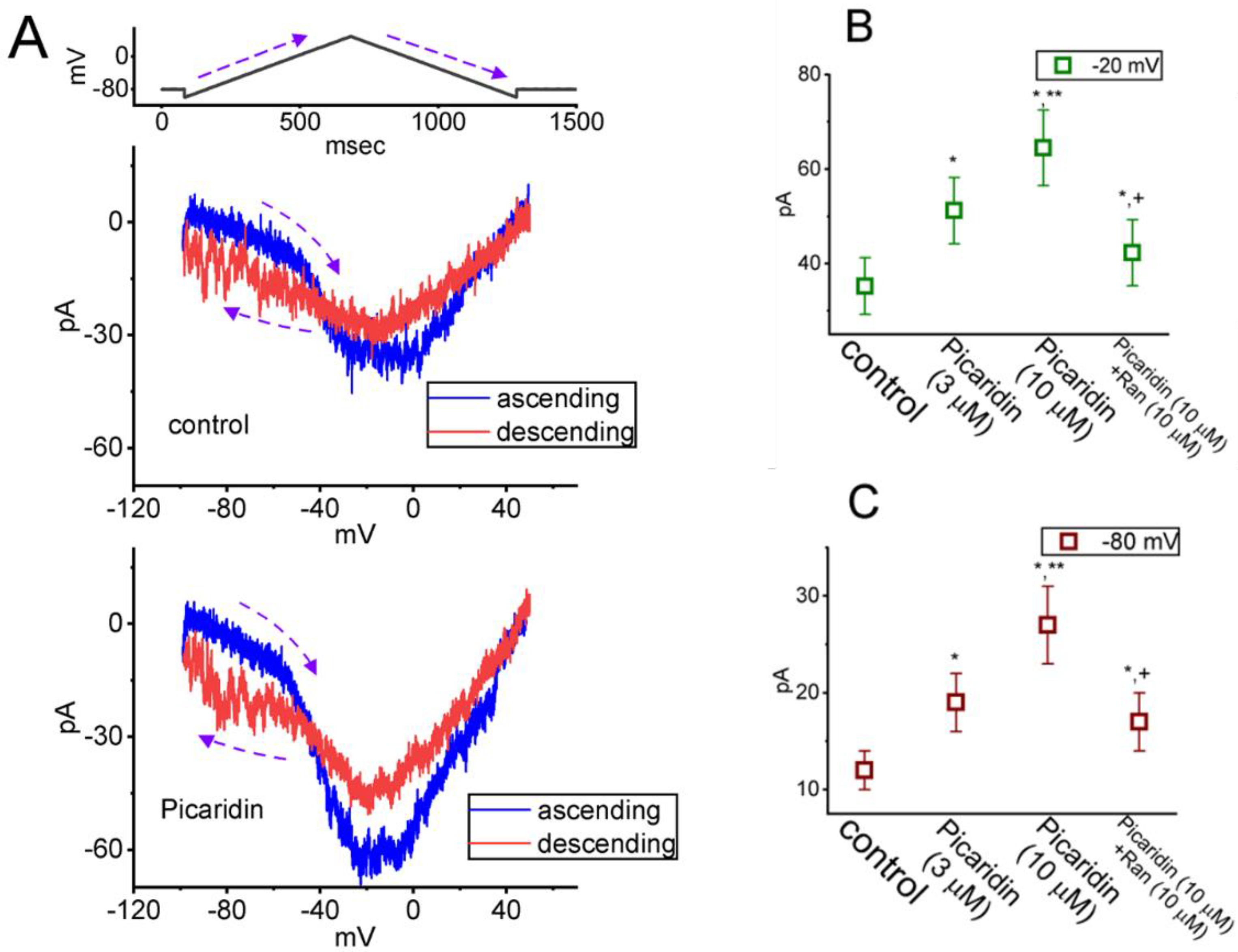
2. Results
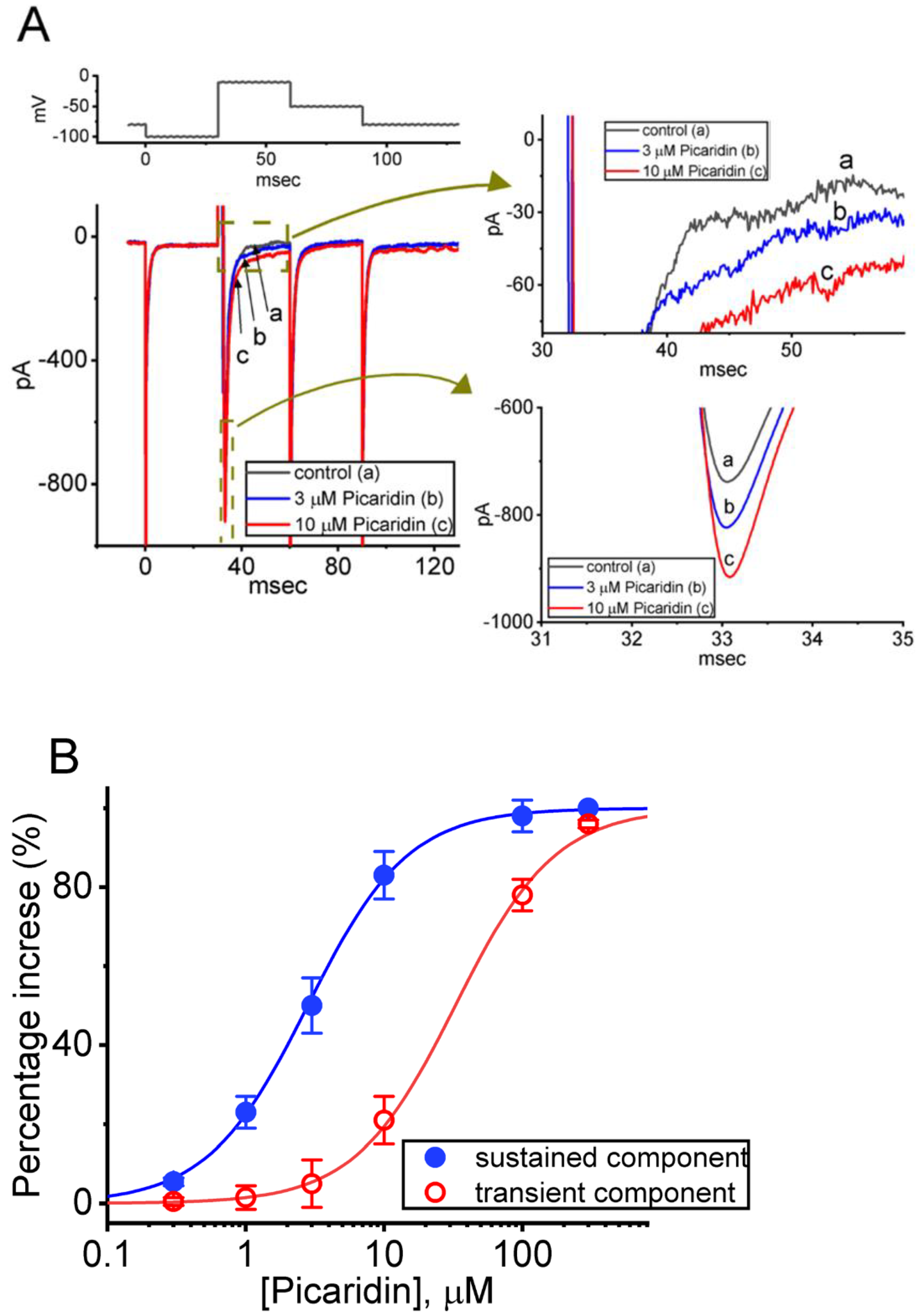
2.1. Stimulatory Effect of Picaridin on Voltage-Gated Na+ Current (INa) Experimentally Recorded from Pituitary GH3 Cells
2.2. Effect of Picaridin on Mean Current versus Voltage (I–V) Relationship of INa(T)
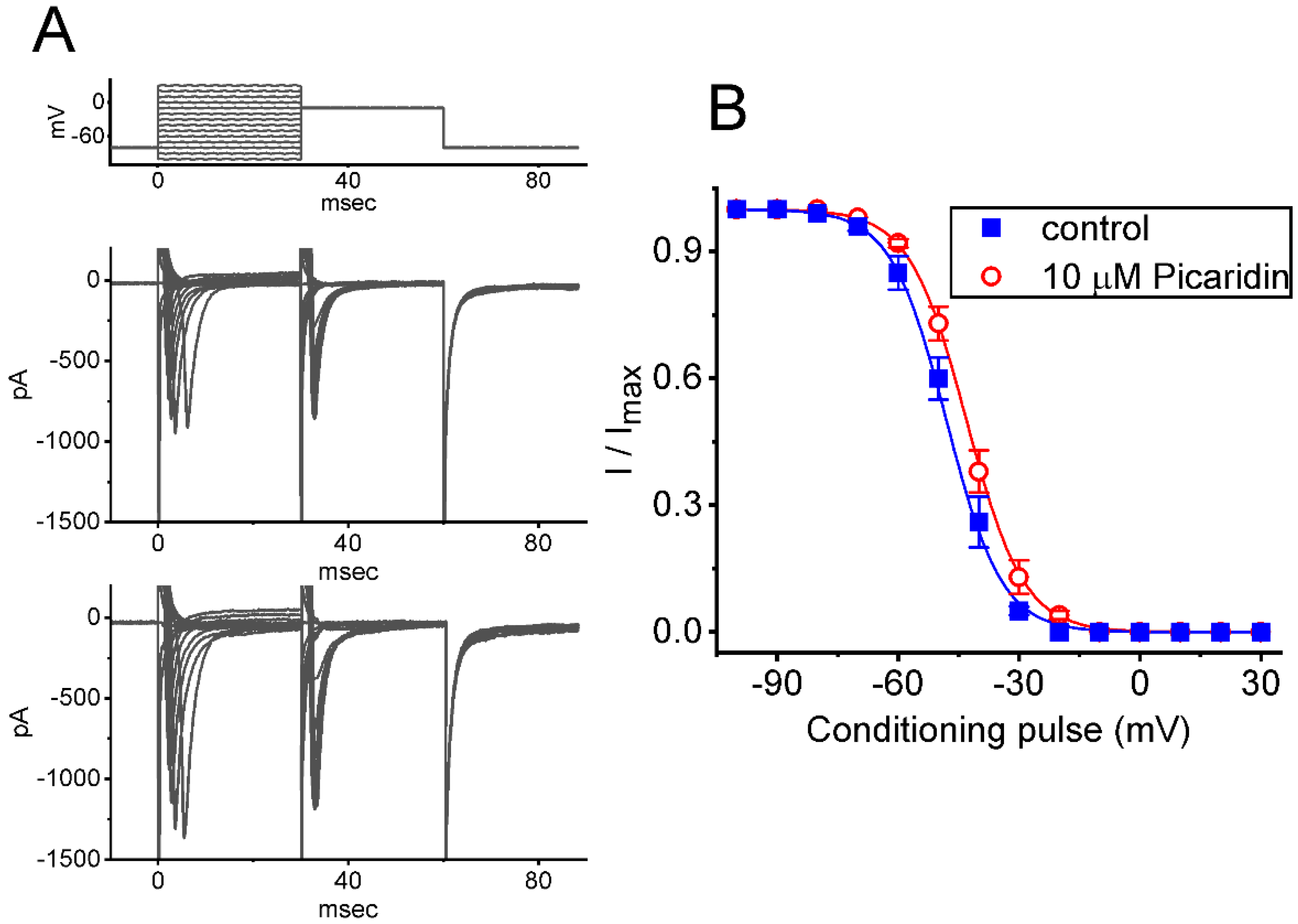
2.3. Characterization of Picaridin-Induced Modifications on the Steady-State Inactivation Curve of INa(T)
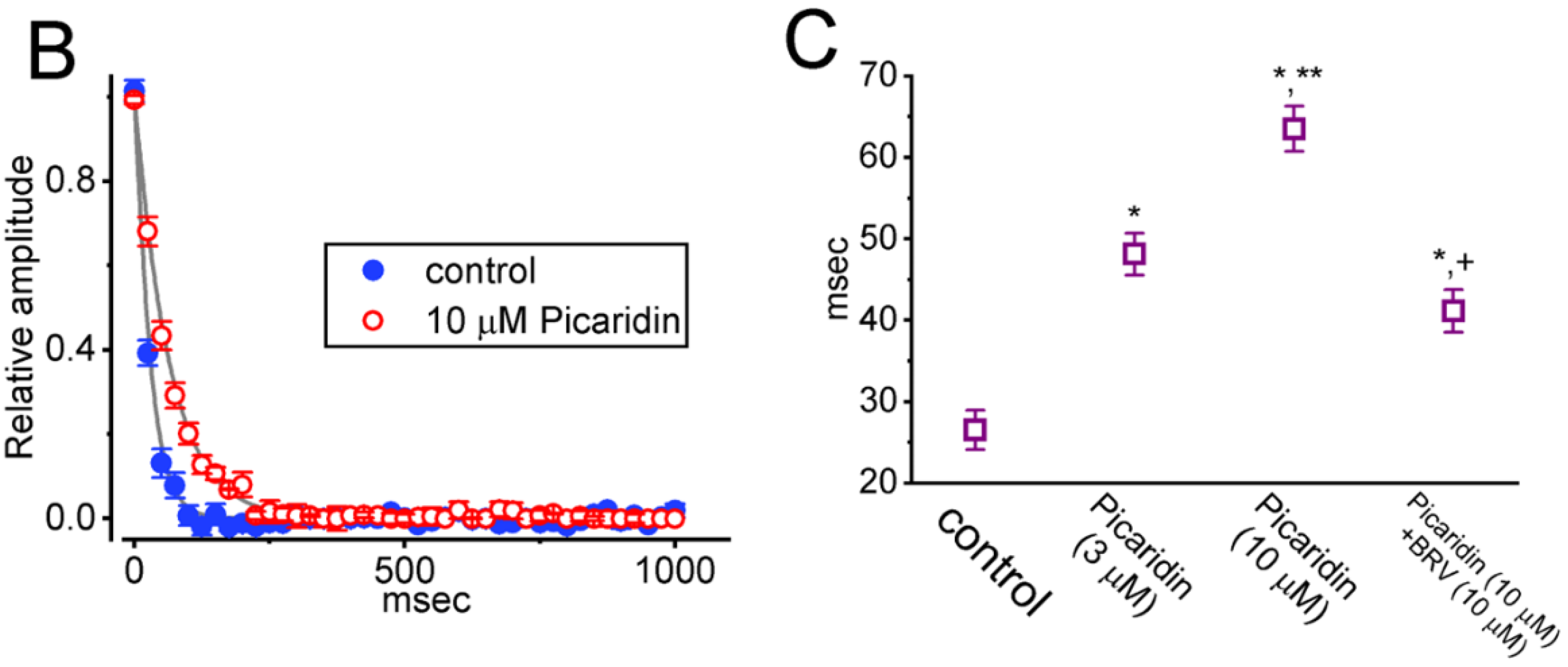
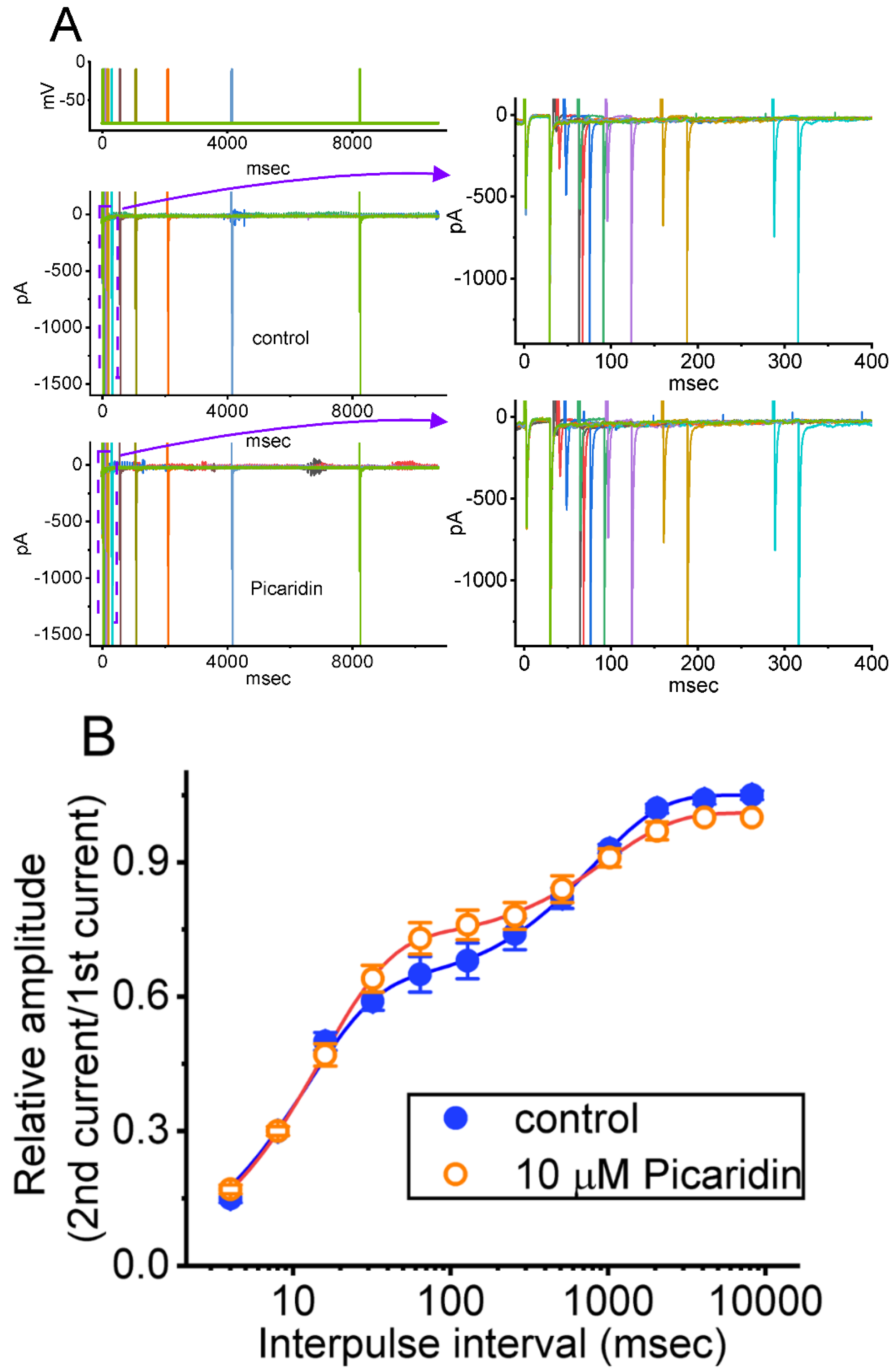
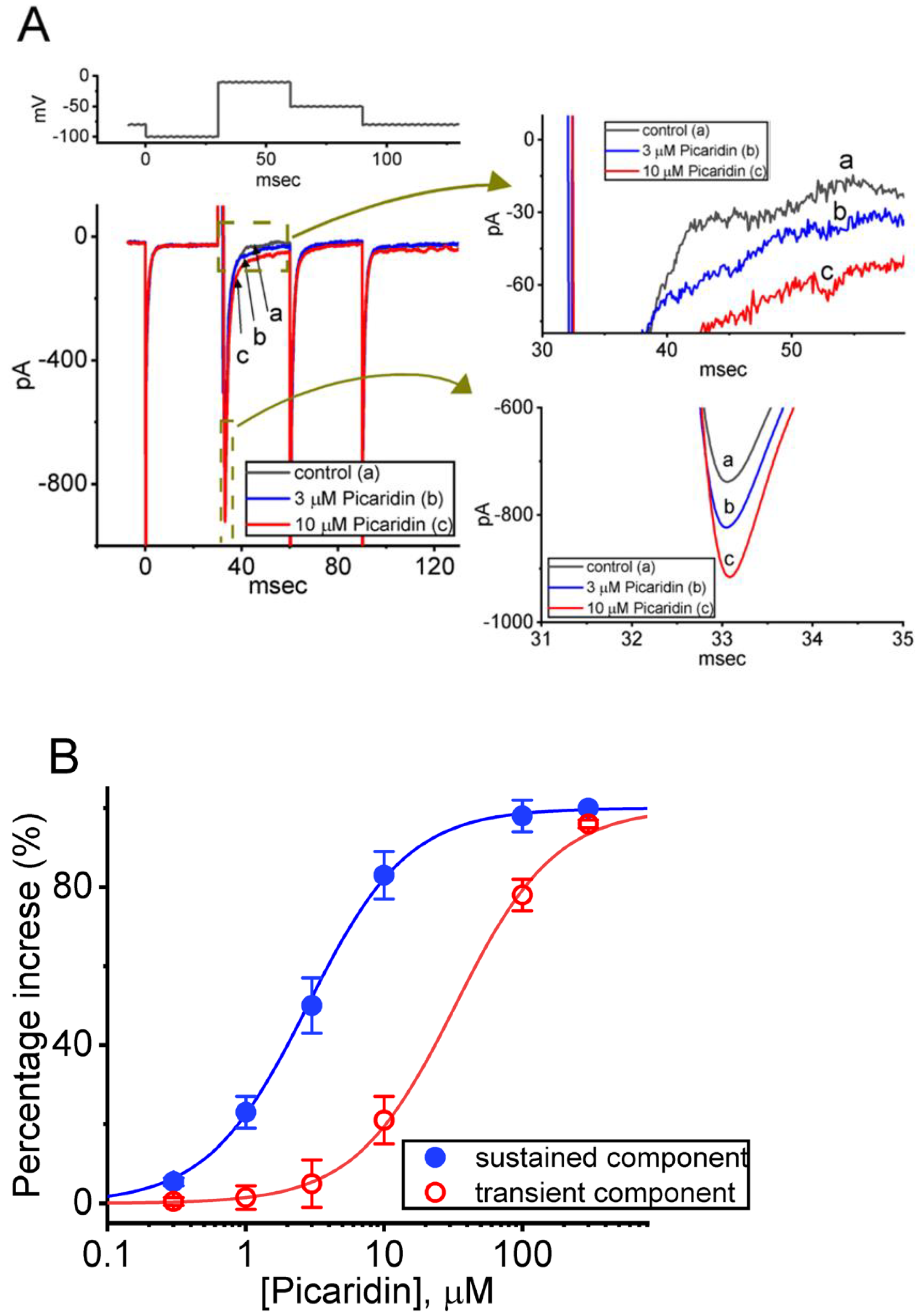
2.4. Picaridin-Induced Retardation in Cumulative Inhibition of INa(T) Inactivation during Rapid Depolarizing Stimuli
2.5. Effect of Picaridin on the Recovery Time Course of INa(T) Inactivation after the Conditioning Train of Depolarizing Stimuli
2.6. Effect of Picaridin on the Strength of Voltage-Dependent Hysteresis (Hys(V)) of Persistent INa (INa(P)) Elicited by an Upright Isosceles-Triangular Vramp
2.7. Picaridin-Mediated Increase of Resurgent INa (INa(R)) Measured from GH3 Cells
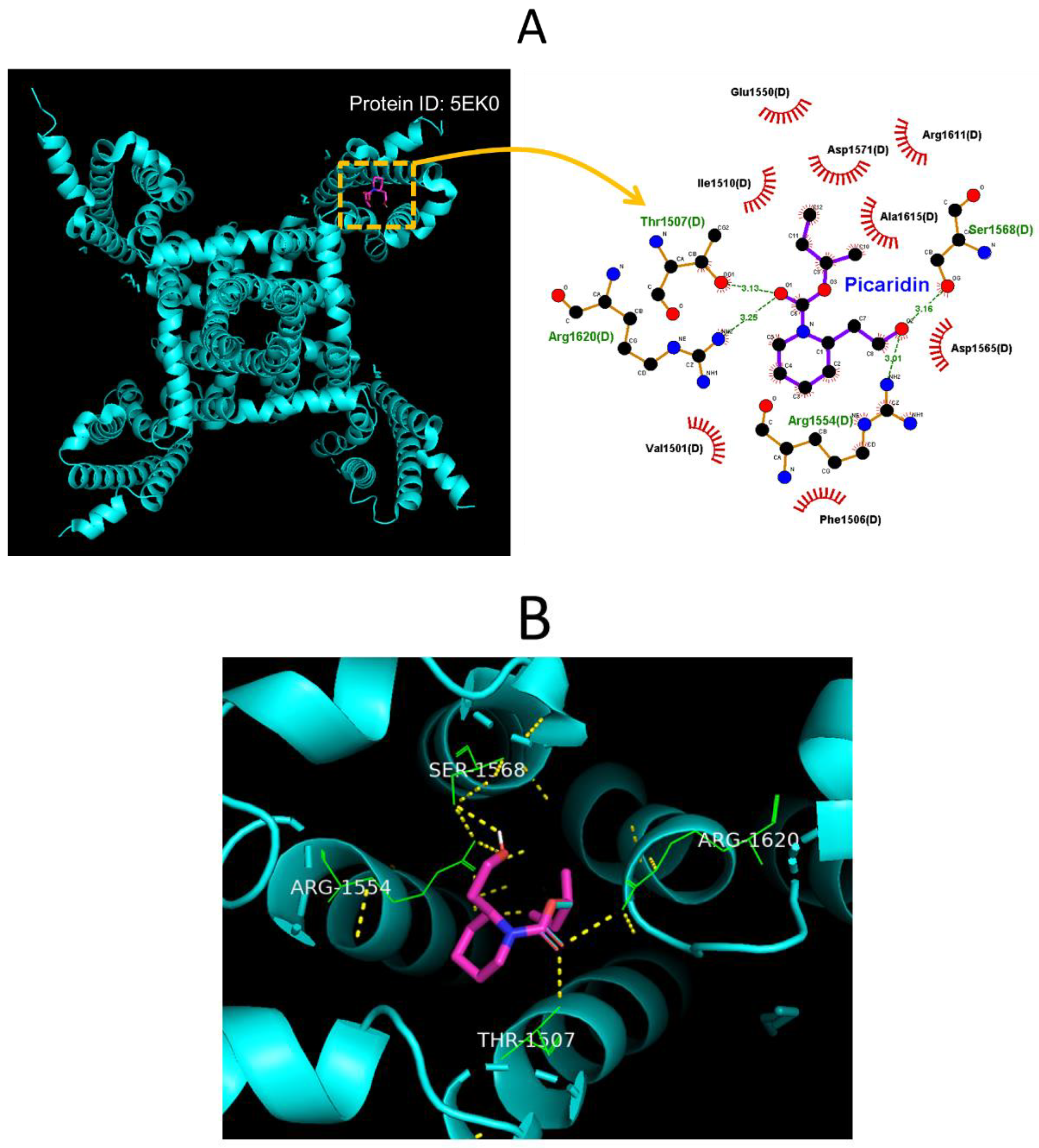
2.8. Molecular Docking on Interaction between NaV1.7 Channel and Picaridin
3. Discussion
4. Materials and Methods
4.1. Chemicals, Drugs, REAGENTS and Solutions Used in This Work
4.2. Cell Preparations
4.3. Electrophysiological Measurements
4.4. Data Recordings and Processing
4.5. Data Analyses for Whole-CELL Ionic Currents
4.6. Curve-Fitting Approximations and Statistical Analyse
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BRV | brivaracetam |
| EC50 | concentration required for half-maximal stimulation |
| Hys(V) | voltage-dependent hysteresis |
| INa(L) | late Na+ current |
| INa(T) | transient (peak) Na+ current |
| NaV channel | voltage-gated Na+ channel |
| Ran | ranolazine |
| SEM | standard error of mean |
| TEA | tetraethylammonium chloride |
| τinact(S) | slow component in inactivation time constant |
| Vramp | ramp voltage |
| TTX | tetrodotoxin |
| Vramp | ramp voltage |
References
- Katz, T.M.; Miller, J.H.; Hebert, A.A. Insect repellents: Historical perspectives and new developments. J. Am. Acad. Dermatol. 2008, 58, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Pages, F.; Dautel, H.; Duvallet, G.; Kahl, O.; de Gentile, L.; Boulanger, N. Tick repellents for human use: Prevention of tick bites and tick-borne diseases. Vector Borne Zoonotic Dis. 2014, 14, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.H. Chemical and Plant-Based Insect Repellents: Efficacy, Safety, and Toxicity. Wilderness Environ. Med. 2016, 27, 153–163. [Google Scholar] [CrossRef]
- Heng, S.; Sluydts, V.; Durnez, L.; Mean, V.; Polo, K.; Tho, S.; Coosemans, M.; van Griensven, J. Safety of a topical insect repellent (picaridin) during community mass use for malaria control in rural Cambodia. PLoS ONE 2017, 12, e0172566. [Google Scholar] [CrossRef] [PubMed]
- Islam, J.; Zaman, K.; Duarah, S.; Raju, P.S.; Chattopadhyay, P. Mosquito repellents: An insight into the chronological perspectives and novel discoveries. Acta Trop. 2017, 167, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Nentwig, G.; Frohberger, S.; Sonneck, R. Evaluation of Clove Oil, Icaridin, and Transfluthrin for Spatial Repellent Effects in Three Tests Systems Against the Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2017, 54, 150–158. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R. Repellence of essential oils and selected compounds against ticks—A systematic review. Acta Trop. 2018, 179, 47–54. [Google Scholar] [CrossRef]
- Devillers, J. 2D and 3D structure-activity modelling of mosquito repellents: A review ($). SAR QSAR Environ. Res. 2018, 29, 693–723. [Google Scholar] [CrossRef]
- Nguyen, Q.D.; Vu, M.N.; Hebert, A.A. Insect Repellents: An Updated Review for the Clinician. J. Am. Acad Dermatol. 2018. [Google Scholar] [CrossRef]
- Tavares, M.; da Silva, M.R.M.; de Oliveira de Siqueira, L.B.; Rodrigues, R.A.S.; Bodjolle-d’Almeida, L.; Dos Santos, E.P.; Ricci-Júnior, E. Trends in insect repellent formulations: A review. Int J. Pharm. 2018, 539, 190–209. [Google Scholar] [CrossRef]
- Chauhan, K.R.; McPhatter, L.P.; O’Dell, K.; Syed, Z.; Wheeler, A.; Debboun, M. Evaluation of a Novel User-Friendly Arthropod Repellent Gel, Verdegen. J. Med. Entomol. 2021, 58, 2479–2483. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, T.C.; Janich, A.J.; Sanchez-Vargas, I.; Markle, E.D.; Gray, M.; Foster, J.R.; Black Iv, W.C.; Foy, B.D.; Olson, K.E. Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus. Insects 2021, 12, 386. [Google Scholar] [CrossRef] [PubMed]
- Germano-Costa, T.; Bilesky-José, N.; Guilger-Casagrande, M.; Pasquoto-Stigliani, T.; Rogério, C.B.; Abrantes, D.C.; Maruyama, C.R.; Oliveira, J.L.; Fraceto, L.F.; Lima, R. Use of 2D and co-culture cell models to assess the toxicity of zein nanoparticles loading insect repellents icaridin and geraniol. Colloids Surf. B Biointerfaces 2022, 216, 112564. [Google Scholar] [CrossRef]
- Kopsco, H.L.; Mather, T.N. Tick-Borne Disease Prevention Behaviors Among Participants in a Tick Surveillance System Compared with a Sample Of Master Gardeners. J. Community Health 2022, 47, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Rogerio, C.B.; Carvalho Abrantes, D.; de Oliveira, J.L.; Ribeiro de Araújo, D.; Germano da Costa, T.; de Lima, R.; Fernandes Fraceto, L. Cellulose Hydrogels Containing Geraniol and Icaridin Encapsulated in Zein Nanoparticles for Arbovirus Control. ACS Appl. Bio Mater. 2022, 5, 1273–1283. [Google Scholar] [CrossRef]
- Zeng, W.; Ren, H.; Zhu, H.; Shi, W.; Xu, L. Icaridin-Loading Nitrocellulose As a New Repellent Against Aquatic Leech with Waterproof and Long-Acting Properties. Vector Borne Zoonotic Dis. 2022, 22, 114–119. [Google Scholar] [CrossRef]
- Bohbot, J.D.; Dickens, J.C. Insect repellents: Modulators of mosquito odorant receptor activity. PLoS ONE 2010, 5, e12138. [Google Scholar] [CrossRef]
- Xu, P.; Choo, Y.M.; De La Rosa, A.; Leal, W.S. Mosquito odorant receptor for DEET and methyl jasmonate. Proc. Natl. Acad. Sci. USA 2014, 111, 16592–16597. [Google Scholar] [CrossRef]
- Sparks, J.T.; Dickens, J.C. Bitter-sensitive gustatory receptor neuron responds to chemically diverse insect repellents in the common malaria mosquito Anopheles quadrimaculatus. Naturwissenschaften 2016, 103, 39. [Google Scholar] [CrossRef]
- Drakou, C.E.; Tsitsanou, K.E.; Potamitis, C.; Fessas, D.; Zervou, M.; Zographos, S.E. The crystal structure of the AgamOBP1 Icaridin complex reveals alternative binding modes and stereo-selective repellent recognition. Cell Mol. Life Sci. 2017, 74, 319–338. [Google Scholar] [CrossRef]
- Afify, A.; Betz, J.F.; Riabinina, O.; Lahondère, C.; Potter, C.J. Commonly Used Insect Repellents Hide Human Odors from Anopheles Mosquitoes. Curr. Biol. 2019, 29, 3669–3680.e3665. [Google Scholar] [CrossRef] [PubMed]
- Kritsi, E.; Liggri, P.G.V.; Stamati, E.C.V.; Tsitsanou, K.E.; Zographos, S.E.; Michaelakis, A.; Papachristos, D.; Zoumpoulakis, P. A Combined Computational Methodology for the Discovery of Hit Compounds with Putative Insect Repellency Properties. ChemMedChem 2022, 17, e202200271. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy. Neurochem. Res. 2017, 42, 2495–2504. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endocr. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Bjelobaba, I.; Zemkova, H. Ion Channels of Pituitary Gonadotrophs and Their Roles in Signaling and Secretion. Front. Endocrinol. 2017, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Chen, B.S.; Hsu, T.I.; Peng, H.; Wu, Y.H.; Lo, Y.C. Analytical studies of rapidly inactivating and noninactivating sodium currents in differentiated NG108-15 neuronal cells. J. Theor. Biol. 2009, 259, 828–836. [Google Scholar] [CrossRef]
- Huang, C.W.; Hung, T.Y.; Wu, S.N. The inhibitory actions by lacosamide, a functionalized amino acid, on voltage-gated Na+ currents. Neuroscience 2015, 287, 125–136. [Google Scholar] [CrossRef]
- Chen, B.S.; Lo, Y.C.; Peng, H.; Hsu, T.I.; Wu, S.N. Effects of ranolazine, a novel anti-anginal drug, on ion currents and membrane potential in pituitary tumor GH(3) cells and NG108-15 neuronal cells. J. Pharmacol. Sci. 2009, 110, 295–305. [Google Scholar] [CrossRef]
- Maier, L.S. A novel mechanism for the treatment of angina, arrhythmias, and diastolic dysfunction: Inhibition of late I(Na) using ranolazine. J. Cardiovasc. Pharmacol. 2009, 54, 279–286. [Google Scholar] [CrossRef]
- Lo, Y.C.; Tseng, Y.T.; Liu, C.M.; Wu, B.N.; Wu, S.N. Actions of KMUP-1, a xanthine and piperazine derivative, on voltage-gated Na+ and Ca2+ -activated K+ currents in GH3 pituitary tumour cells. Br. J. Pharmacol. 2015, 172, 5110–5122. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.N.; Wu, Y.H.; Chen, B.S.; Lo, Y.C.; Liu, Y.C. Unnderlying mechanism of actions of tefluthrin, a pyrethroid insecticide, on voltage-gated ion currents and on action currents in pituitary tumor (GH3) cells and GnRH-secreting (GT1-7) neurons. Toxicology 2009, 258, 70–77. [Google Scholar] [CrossRef] [PubMed]
- So, E.C.; Wu, S.N.; Lo, Y.C.; Su, K. Differential regulation of tefluthrin and telmisartan on the gating charges of I(Na) activation and inactivation as well as on resurgent and persistent I(Na) in a pituitary cell line (GH(3)). Toxicol. Lett. 2018, 285, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. Effective Accentuation of Voltage-Gated Sodium Current Caused by Apocynin (4’-Hydroxy-3’-methoxyacetophenone), a Known NADPH-Oxidase Inhibitor. Biomedicines 2021, 9, 1146. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Charles, I.; James, A.F.; Abdala, A.P.; Hancox, J.C. Delayed Ventricular Repolarization and Sodium Channel Current Modification in a Mouse Model of Rett Syndrome. Int. J. Mol. Sci. 2022, 23, 5735. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhou, L.; Li, D.; Lai, Q.; Shi, H.; Wang, M. Ecotoxicological effects of the pyrethroid insecticide tefluthrin to the earthworm Eisenia fetida: A chiral view. Environ. Res. 2020, 190, 109991. [Google Scholar] [CrossRef]
- Wu, G.; Li, Q.; Liu, X.; Li-Byarlay, H.; He, B. Differential state-dependent effects of deltamethrin and tefluthrin on sodium channels in central neurons of Helicoverpa armigera. Pestic. Biochem. Physiol. 2021, 175, 104836. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Effectiveness of Columbianadin, a Bioactive Coumarin Derivative, in Perturbing Transient and Persistent I(Na). Int. J. Mol. Sci. 2021, 22, 621. [Google Scholar] [CrossRef]
- Taddese, A.; Bean, B.P. Subthreshold sodium current from rapidly inactivating sodium channels drives spontaneous firing of tuberomammillary neurons. Neuron 2002, 33, 587–600. [Google Scholar] [CrossRef]
- Tsai, D.; Morley, J.W.; Suaning, G.J.; Lovell, N.H. Frequency-dependent reduction of voltage-gated sodium current modulates retinal ganglion cell response rate to electrical stimulation. J. Neural Eng. 2011, 8, 066007. [Google Scholar] [CrossRef]
- Wu, P.M.; Cho, H.Y.; Chiang, C.W.; Chuang, T.H.; Wu, S.N.; Tu, Y.F. Characterization in Inhibitory Effectiveness of Carbamazepine in Voltage-Gated Na+ and Erg-Mediated K+ Currents in a Mouse Neural Crest-Derived (Neuro-2a) Cell Line. Int. J. Mol. Sci. 2022, 23, 7892. [Google Scholar] [CrossRef]
- Wu, C.L.; Chuang, C.W.; Cho, H.Y.; Chuang, T.H.; Wu, S.N. The Evidence for Effective Inhibition of I(Na) Produced by Mirogabalin ((1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid), a Known Blocker of Ca(V) Channels. Int. J. Mol. Sci. 2022, 23, 3845. [Google Scholar] [CrossRef]
- Carter, B.C.; Bean, B.P. Incomplete inactivation and rapid recovery of voltage-dependent sodium channels during high-frequency firing in cerebellar Purkinje neurons. J. Neurophysiol. 2011, 105, 860–871. [Google Scholar] [CrossRef]
- Hung, T.Y.; Wu, S.N.; Huang, C.W. The Integrated Effects of Brivaracetam, a Selective Analog of Levetiracetam, on Ionic Currents and Neuronal Excitability. Biomedicines 2021, 9, 369. [Google Scholar] [CrossRef]
- Wang, D.W.; Mistry, A.M.; Kahlig, K.M.; Kearney, J.A.; Xiang, J.; George, A.L., Jr. Propranolol blocks cardiac and neuronal voltage-gated sodium channels. Front. Pharmacol. 2010, 1, 144. [Google Scholar] [CrossRef]
- Powers, R.K.; Binder, M.D. Persistent sodium and calcium currents in rat hypoglossal motoneurons. J. Neurophysiol. 2003, 89, 615–624. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Characterization of Direct Perturbations on Voltage-Gated Sodium Current by Esaxerenone, a Nonsteroidal Mineralocorticoid Receptor Blocker. Biomedicines 2021, 9, 549. [Google Scholar] [CrossRef]
- Stafstrom, C.E. Persistent sodium current and its role in epilepsy. Epilepsy Curr. 2007, 7, 15–22. [Google Scholar] [CrossRef]
- Boeri, J.; Le Corronc, H.; Lejeune, F.X.; Le Bras, B.; Mouffle, C.; Angelim, M.; Mangin, J.M.; Branchereau, P.; Legendre, P.; Czarnecki, A. Persistent Sodium Current Drives Excitability of Immature Renshaw Cells in Early Embryonic Spinal Networks. J. Neurosci. 2018, 38, 7667–7682. [Google Scholar] [CrossRef]
- Hsu, C.L.; Zhao, X.; Milstein, A.D.; Spruston, N. Persistent Sodium Current Mediates the Steep Voltage Dependence of Spatial Coding in Hippocampal Pyramidal Neurons. Neuron 2018, 99, 147–162.e148. [Google Scholar] [CrossRef]
- Müller, P.; Draguhn, A.; Egorov, A.V. Persistent sodium current modulates axonal excitability in CA1 pyramidal neurons. J. Neurochem. 2018, 146, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Meredith, F.L.; Rennie, K.J. Persistent and resurgent Na+ currents in vestibular calyx afferents. J. Neurophysiol. 2020, 124, 510–524. [Google Scholar] [CrossRef]
- Enomoto, A.; Han, J.M.; Hsiao, C.F.; Wu, N.; Chandler, S.H. Participation of sodium currents in burst generation and control of membrane excitability in mesencephalic trigeminal neurons. J. Neurosci. 2006, 26, 3412–3422. [Google Scholar] [CrossRef]
- Hong, H.; Lu, T.; Wang, X.; Wang, Y.; Sanchez, J.T. Resurgent sodium current promotes action potential firing in the avian auditory brainstem. J. Physiol. 2018, 596, 423–443. [Google Scholar] [CrossRef]
- Kuo, P.C.; Kao, Z.H.; Lee, S.W.; Wu, S.N. Effects of Sesamin, the Major Furofuran Lignan of Sesame Oil, on the Amplitude and Gating of Voltage-Gated Na+ and K+ Currents. Molecules 2020, 25, 3062. [Google Scholar] [CrossRef]
- Tidball, A.M.; Lopez-Santiago, L.F.; Yuan, Y.; Glenn, T.W.; Margolis, J.L.; Clayton Walker, J.; Kilbane, E.G.; Miller, C.A.; Martina Bebin, E.; Scott Perry, M.; et al. Variant-specific changes in persistent or resurgent sodium current in SCN8A-related epilepsy patient-derived neurons. Brain 2020, 143, 3025–3040. [Google Scholar] [CrossRef]
- Quattrocolo, G.; Dunville, K.; Nigro, M.J. Resurgent Sodium Current in Neurons of the Cerebral Cortex. Front. Cell Neurosci. 2021, 15, 760610. [Google Scholar] [CrossRef]
- Zemel, B.M.; Nevue, A.A.; Dagostin, A.; Lovell, P.V.; Mello, C.V.; von Gersdorff, H. Resurgent Na+ currents promote ultrafast spiking in projection neurons that drive fine motor control. Nat. Commun. 2021, 12, 6762. [Google Scholar] [CrossRef]
- Ransdell, J.L.; Moreno, J.D.; Bhagavan, D.; Silva, J.R.; Nerbonne, J.M. Intrinsic mechanisms in the gating of resurgent Na+ currents. Elife 2022, 11, e70173. [Google Scholar] [CrossRef]
- Ahuja, S.; Mukund, S.; Deng, L.; Khakh, K.; Chang, E.; Ho, H.; Shriver, S.; Young, C.; Lin, S.; Johnson, J.P., Jr.; et al. Structural basis of Nav1.7 inhibition by an isoform-selective small-molecule antagonist. Science 2015, 350, aac5464. [Google Scholar] [CrossRef]
- Charlton, N.P.; Murphy, L.T.; Parker Cote, J.L.; Vakkalanka, J.P. The toxicity of picaridin containing insect repellent reported to the National Poison Data System. Clin. Toxicol. 2016, 54, 655–658. [Google Scholar] [CrossRef]
- Spirhanzlova, P.; Fini, J.B.; Demeneix, B.; Lardy-Fontan, S.; Vaslin-Reimann, S.; Lalere, B.; Guma, N.; Tindall, A.; Krief, S. Composition and endocrine effects of water collected in the Kibale national park in Uganda. Environ. Pollut. 2019, 251, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Magariños-Ascone, C.; Pazo, J.H.; Macadar, O.; Buño, W. High-frequency stimulation of the subthalamic nucleus silences subthalamic neurons: A possible cellular mechanism in Parkinson’s disease. Neuroscience 2002, 115, 1109–1117. [Google Scholar] [CrossRef]
- Khaliq, Z.M.; Gouwens, N.W.; Raman, I.M. The contribution of resurgent sodium current to high-frequency firing in Purkinje neurons: An experimental and modeling study. J. Neurosci. 2003, 23, 4899–4912. [Google Scholar] [CrossRef] [PubMed]
- Avendano-Coy, J.; Serrano-Munoz, D.; Taylor, J.; Goicoechea-Garcia, C.; Gomez-Soriano, J. Peripheral Nerve Conduction Block by High-Frequency Alternating Currents: A Systematic Review. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.A.; Salari, A.; Lin, J.L.; Cowan, L.M.; Penington, N.J.; Milescu, M.; Milescu, L.S. Sodium channels implement a molecular leaky integrator that detects action potentials and regulates neuronal firing. Elife 2020, 9, e54940. [Google Scholar] [CrossRef]
- Zhu, T.; Wei, S.; Wang, Y. Post-Inhibitory Rebound Firing of Dorsal Root Ganglia Neurons. J. Pain Res. 2022, 15, 2029–2040. [Google Scholar] [CrossRef]
- Leão, R.M.; Kushmerick, C.; Pinaud, R.; Renden, R.; Li, G.L.; Taschenberger, H.; Spirou, G.; Levinson, S.R.; von Gersdorff, H. Presynaptic Na+ channels: Locus, development, and recovery from inactivation at a high-fidelity synapse. J. Neurosci. 2005, 25, 3724–3738. [Google Scholar] [CrossRef] [PubMed]
- Osteen, J.D.; Sampson, K.; Iyer, V.; Julius, D.; Bosmans, F. Pharmacology of the Na(v)1.1 domain IV voltage sensor reveals coupling between inactivation gating processes. Proc. Natl. Acad. Sci. USA 2017, 114, 6836–6841. [Google Scholar] [CrossRef]
- Durey, M. Bifurcations and chaos in a Lorenz-like pilot-wave system. Chaos 2020, 30, 103115. [Google Scholar] [CrossRef]
- Wu, S.N.; Chern, J.H.; Shen, S.; Chen, H.H.; Hsu, Y.T.; Lee, C.C.; Chan, M.H.; Lai, M.C.; Shie, F.S. Stimulatory actions of a novel thiourea derivative on large-conductance, calcium-activated potassium channels. J. Cell Physiol. 2017, 232, 3409–3421. [Google Scholar] [CrossRef]
- Chang, W.T.; Liu, P.Y.; Gao, Z.H.; Lee, S.W.; Lee, W.K.; Wu, S.N. Evidence for the Effectiveness of Remdesivir (GS-5734), a Nucleoside-Analog Antiviral Drug in the Inhibition of I (K(M)) or I (K(DR)) and in the Stimulation of I (MEP). Front. Pharmacol. 2020, 11, 1091. [Google Scholar] [CrossRef] [PubMed]
- Weiser, T.; Qu, Y.; Catterall, W.A.; Scheuer, T. Differential interaction of R-mexiletine with the local anesthetic receptor site on brain and heart sodium channel alpha-subunits. Mol. Pharmacol. 1999, 56, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.V.; Espinosa, J.L.; López-Domínguez, A.M.; López-Santiago, L.F.; Navarrete, A.; Cota, G. L-type calcium channel activation up-regulates the mRNAs for two different sodium channel alpha subunits (Nav1.2 and Nav1.3) in rat pituitary GH3 cells. Brain Res. Mol. Brain Res. 2003, 116, 115–125. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | τfast (ms) | τslow (ms) | A | B | |
|---|---|---|---|---|---|
| Control | 8 | 12.1 ± 0.5 | 801 ± 12 | 0.62 ± 0.04 | 0.43 ± 0.02 |
| Picaridin (3 μM) | 8 | 13.9 ± 0.6 * | 893 ± 14 * | 0.67 ± 0.04 * | 0.36 ± 0.02 * |
| Picaridin (10 μM) | 8 | 15.1 ± 0.7 * | 976 ± 15 * | 0.72 ± 0.04 * | 0.29 ± 0.02 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiau, A.-L.; Liao, C.-S.; Tu, C.-W.; Wu, S.-N.; Cho, H.-Y.; Yu, M.-C. Characterization in Effective Stimulation on the Magnitude, Gating, Frequency Dependence, and Hysteresis of INa Exerted by Picaridin (or Icaridin), a Known Insect Repellent. Int. J. Mol. Sci. 2022, 23, 9696. https://doi.org/10.3390/ijms23179696
Shiau A-L, Liao C-S, Tu C-W, Wu S-N, Cho H-Y, Yu M-C. Characterization in Effective Stimulation on the Magnitude, Gating, Frequency Dependence, and Hysteresis of INa Exerted by Picaridin (or Icaridin), a Known Insect Repellent. International Journal of Molecular Sciences. 2022; 23(17):9696. https://doi.org/10.3390/ijms23179696
Chicago/Turabian StyleShiau, Ai-Li, Chih-Szu Liao, Chi-Wen Tu, Sheng-Nan Wu, Hsin-Yen Cho, and Meng-Cheng Yu. 2022. "Characterization in Effective Stimulation on the Magnitude, Gating, Frequency Dependence, and Hysteresis of INa Exerted by Picaridin (or Icaridin), a Known Insect Repellent" International Journal of Molecular Sciences 23, no. 17: 9696. https://doi.org/10.3390/ijms23179696
APA StyleShiau, A.-L., Liao, C.-S., Tu, C.-W., Wu, S.-N., Cho, H.-Y., & Yu, M.-C. (2022). Characterization in Effective Stimulation on the Magnitude, Gating, Frequency Dependence, and Hysteresis of INa Exerted by Picaridin (or Icaridin), a Known Insect Repellent. International Journal of Molecular Sciences, 23(17), 9696. https://doi.org/10.3390/ijms23179696