
Rational Design of Lipase ROL to Increase Its Thermostability for Production of Structured Tags

Abstract
:1. Introduction
2. Results and Discussion
2.1. Sequence Alignment of ROL and Visualization with WebLogo to Display Consensus Sequence
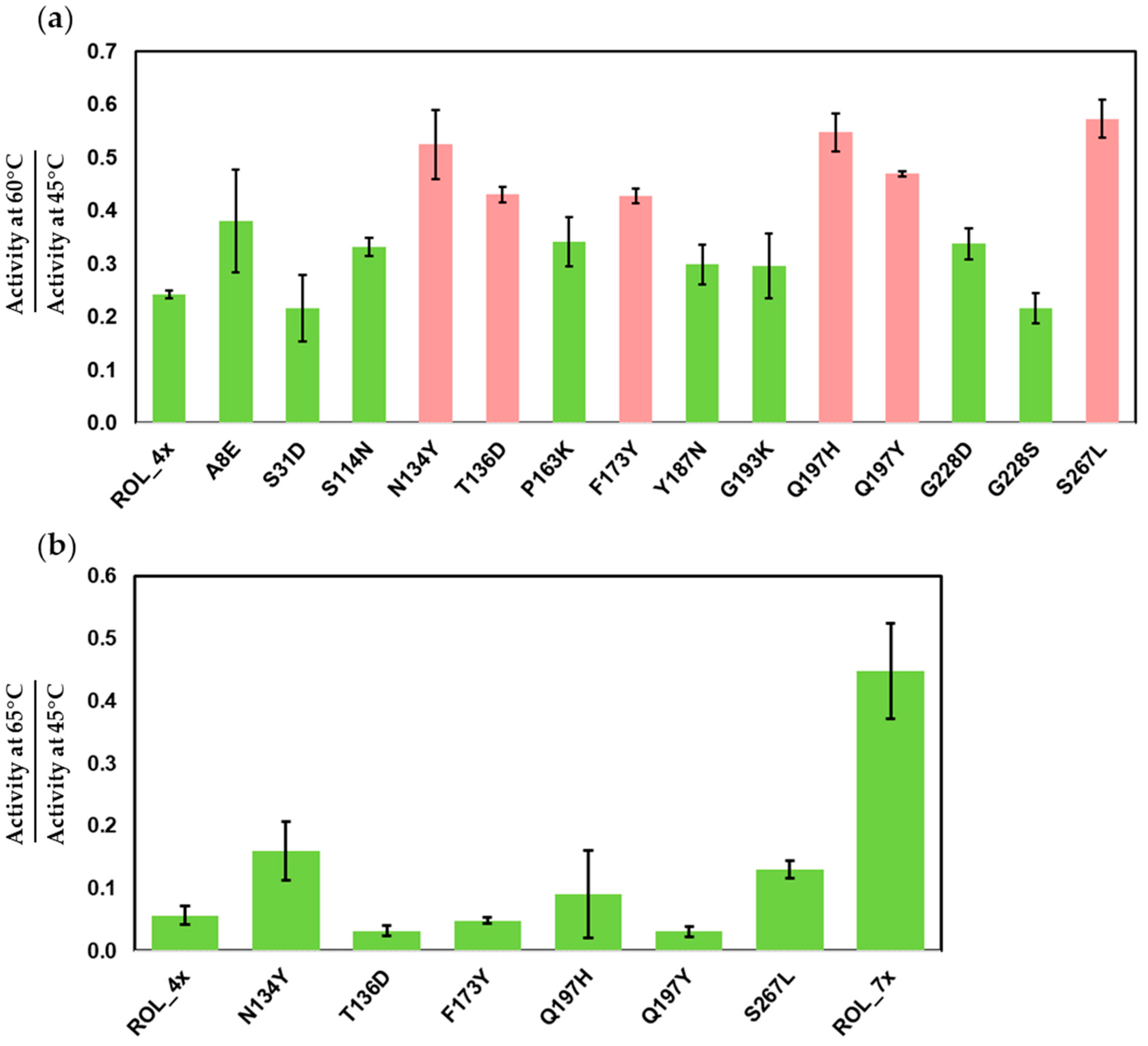
2.2. Expression and Screening of ROL_4x Mutants for Increased Thermostability
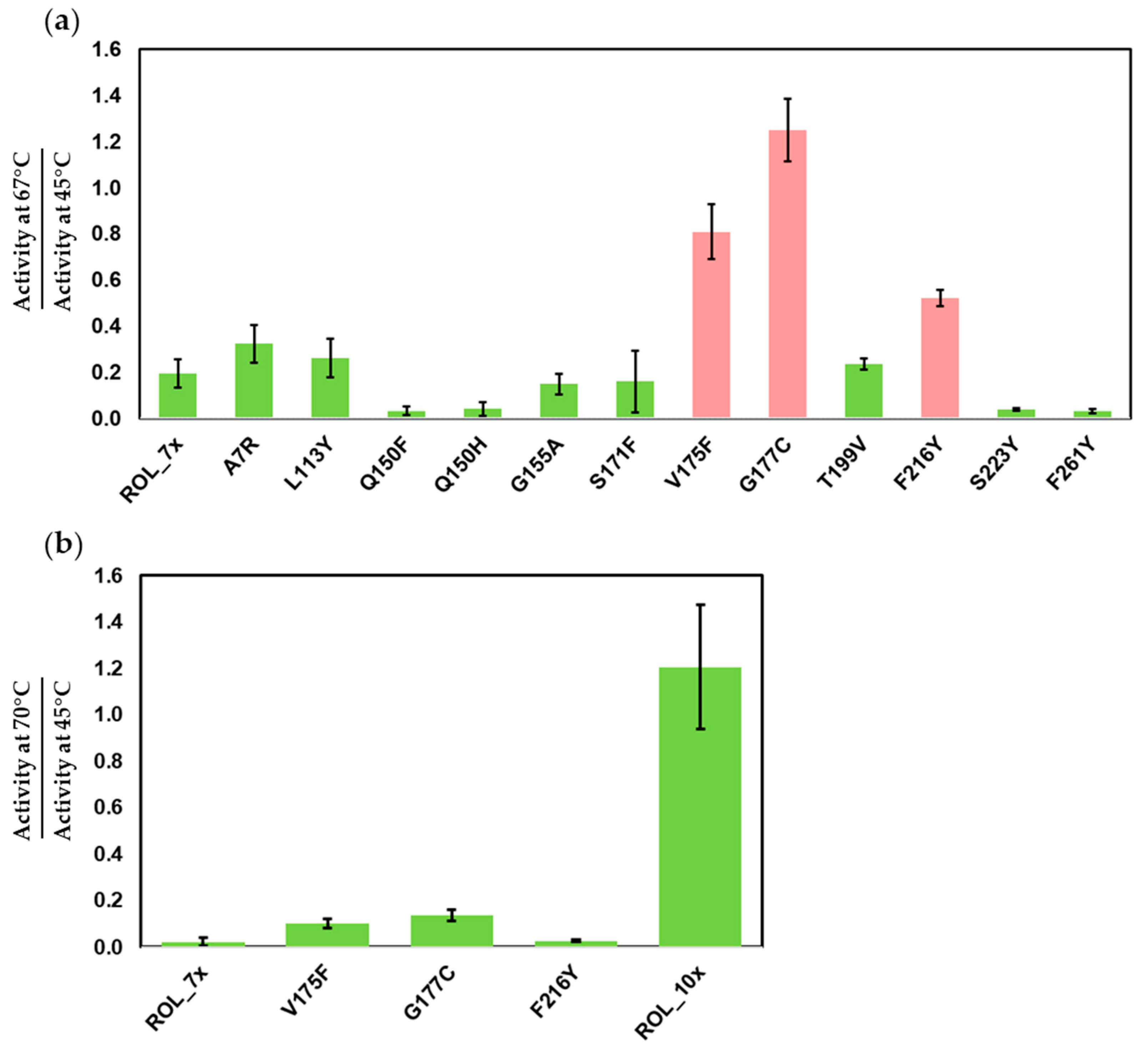
2.3. Screening of ROL_7x Mutants for Increased Thermostability
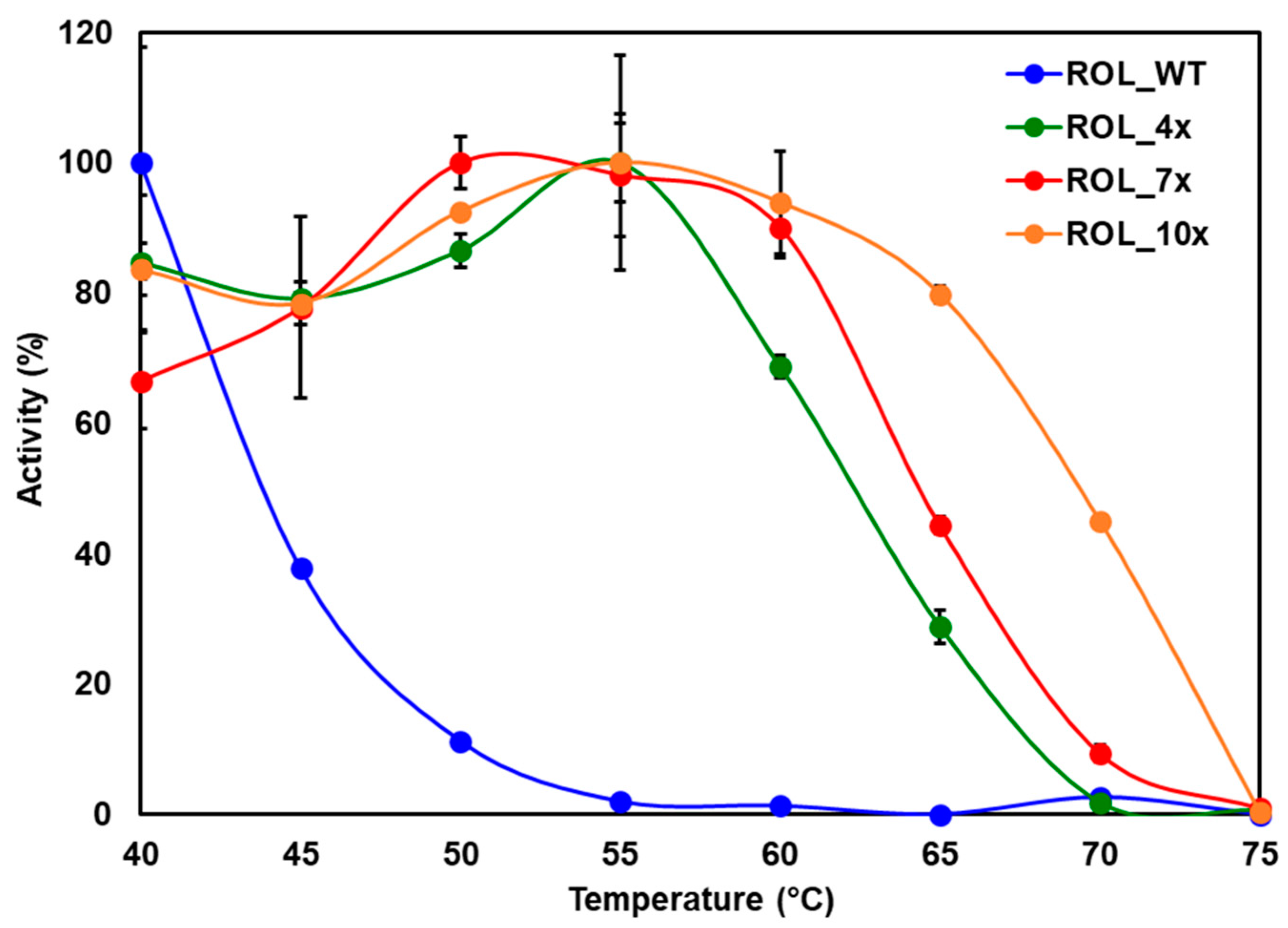
2.4. Large Scale Expression of ROL Mutants and Thermostability Assays
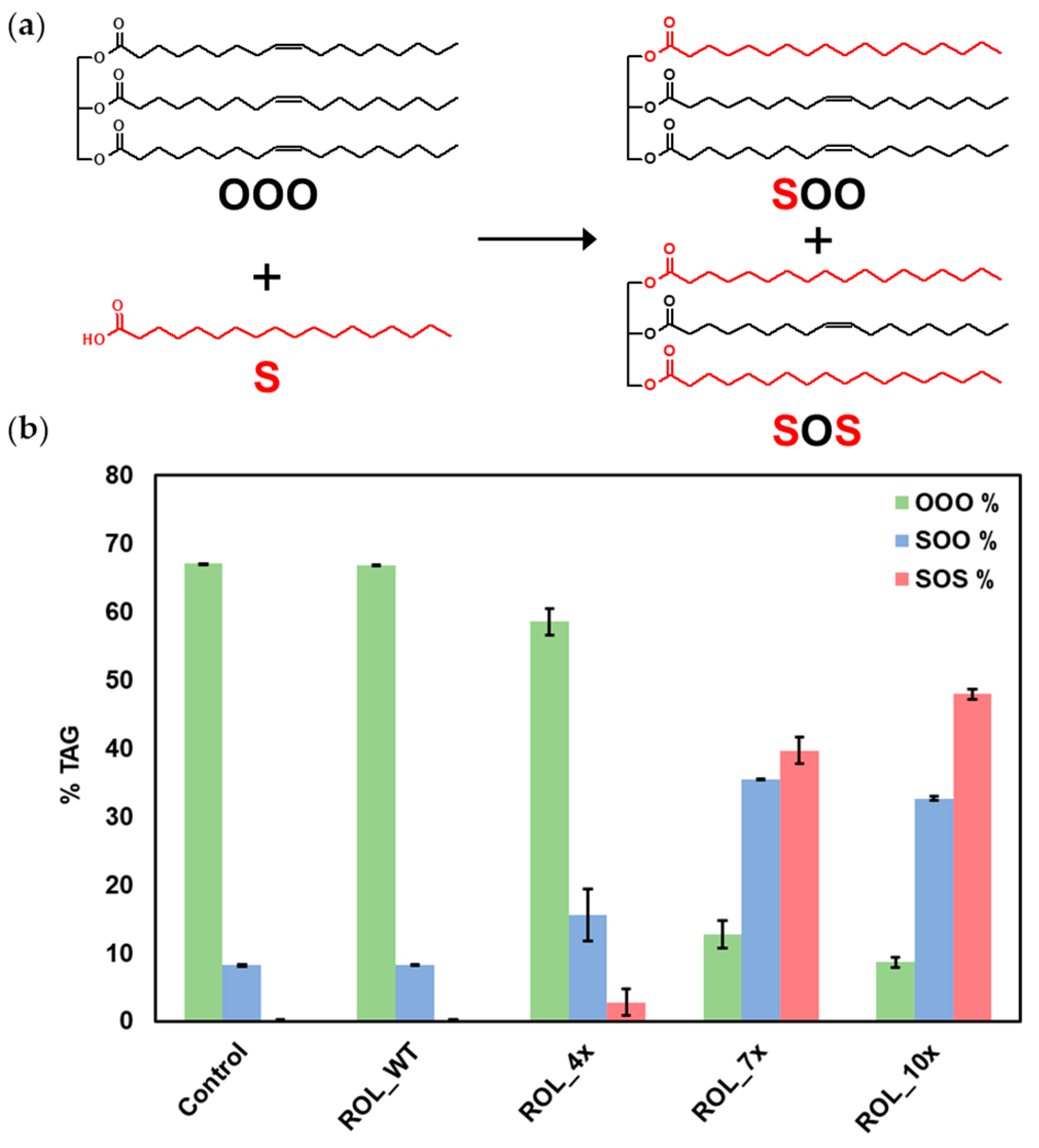
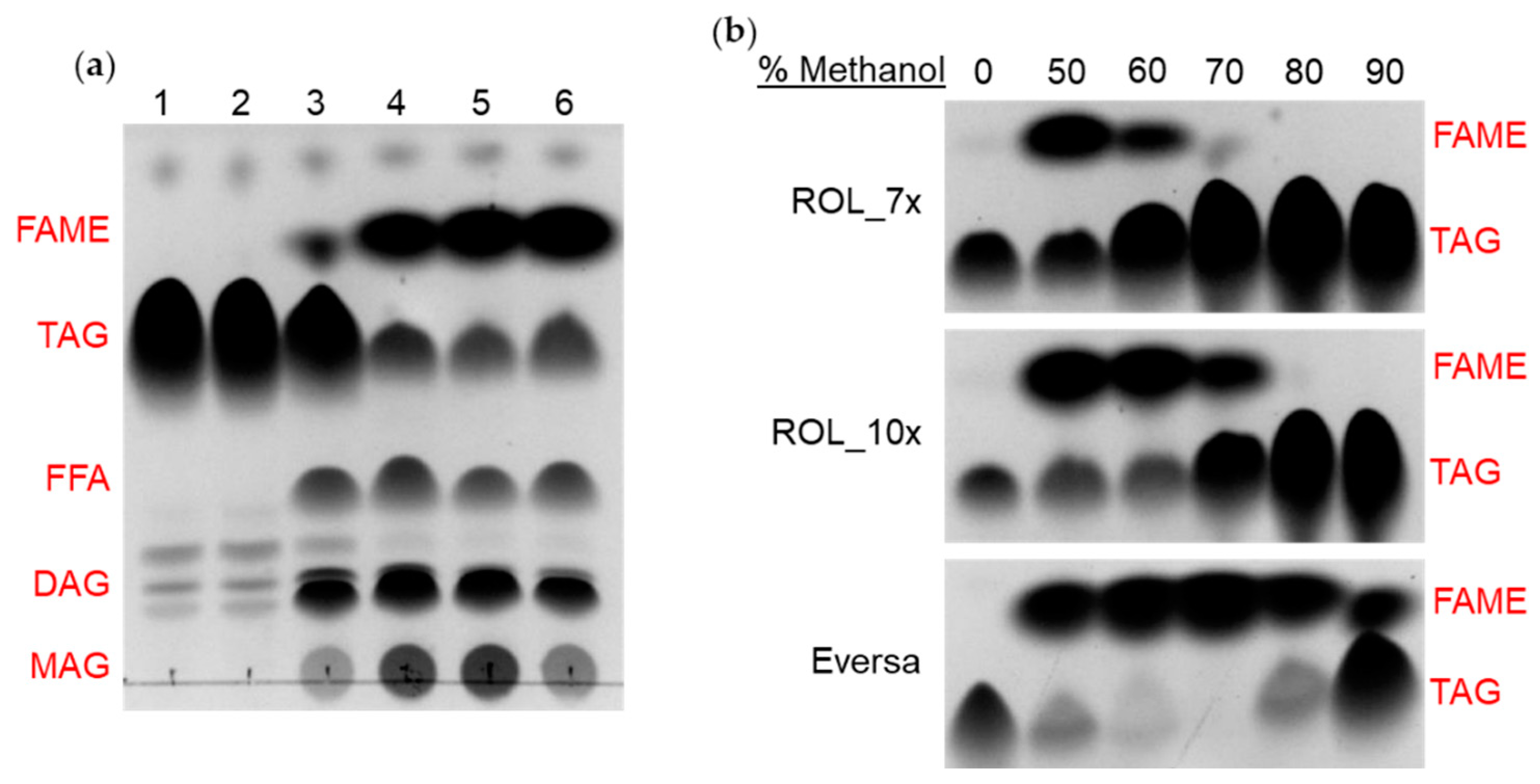
2.5. Methanol Tolerance Assay and Production of Biodiesel
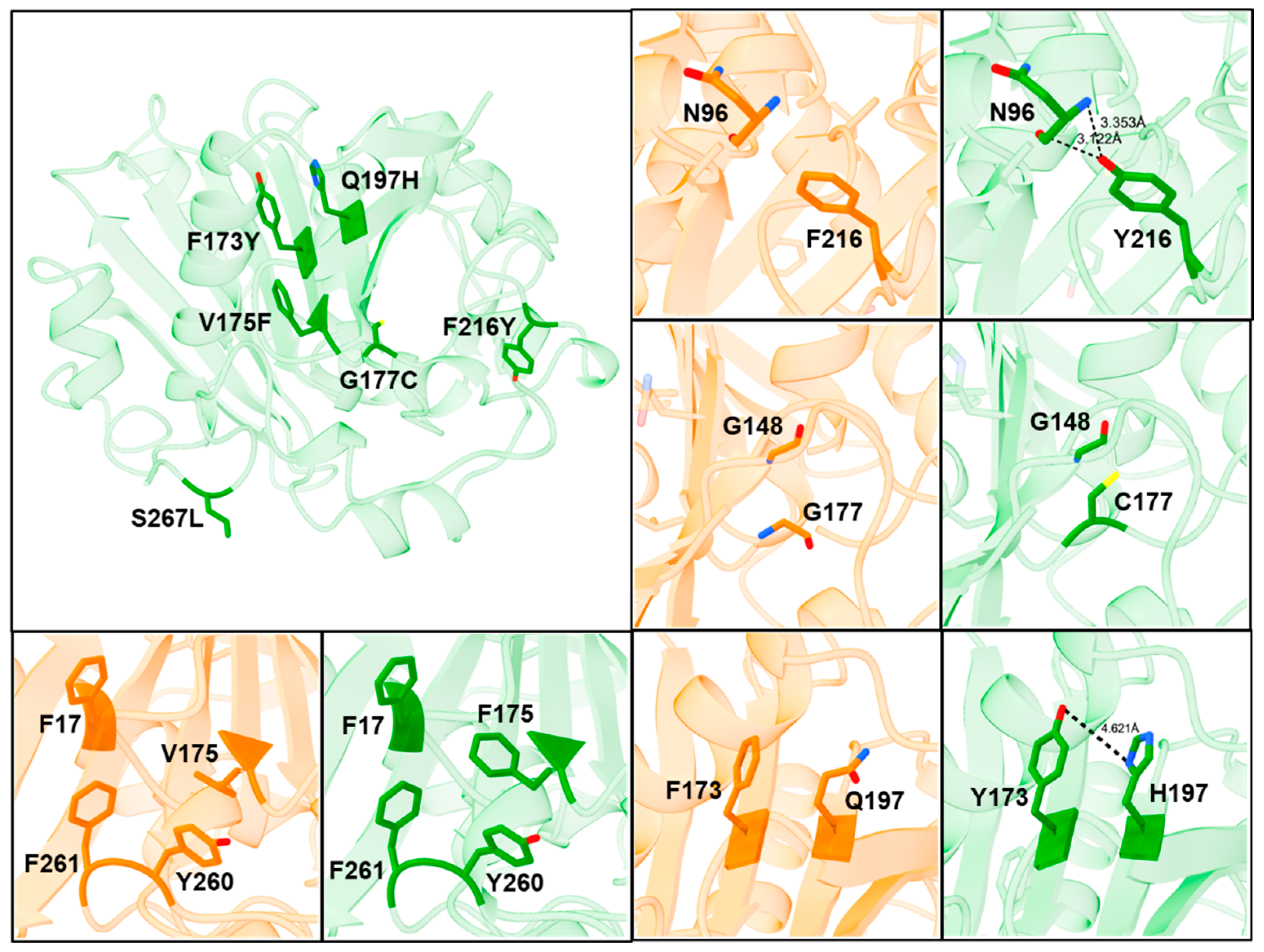
2.6. Mechanism of Increased Stability Observed in the ROL_10x Mutant
2.7. Concluding Remarks
3. Materials and Methods
3.1. Expression of ROL in P. pastoris
3.2. DTNB Lipase Assay for Screening
3.3. Gene Shuffling by PCR and Golden-Gate Cloning
3.4. Determination of the Specific Activity of ROL by Titration
3.5. Analysis of EIE Reaction by TLC Assay
3.6. Analysis of EIE Reaction by GC-FID
3.7. Methanol Tolerance of ROL Mutants
4. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghide, M.K.; Yan, Y. 1,3-Dioleoyl-2-palmitoyl glycerol (OPO)-Enzymatic synthesis and use as an important supplement in infant formulas. J. Food Biochem. 2021, 45, e13799. [Google Scholar] [CrossRef] [PubMed]
- Ghazani, S.M.; Marangoni, A.G. Facile lipase-catalyzed synthesis of a chocolate fat mimetic. Sci. Rep. 2018, 8, 15271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Fernández, J.; Benaiges, M.D.; Valero, F. Rhizopus oryzae Lipase, a Promising Industrial Enzyme: Biochemical Characteristics, Production and Biocatalytic Applications. Catalysts 2020, 10, 1277. [Google Scholar] [CrossRef]
- Kohno, M.; Enatsu, M.; Funatsu, J.; Yoshiizumi, M.; Kugimiya, W. Improvement of the optimum temperature of lipase activity for Rhizopus niveus by random mutagenesis and its structural interpretation. J. Biotechnol. 2001, 87, 203–210. [Google Scholar] [CrossRef]
- Zhao, J.-F.; Wang, Z.; Gao, F.-L.; Lin, J.-P.; Yang, L.-R.; Wu, M.-B. Enhancing the thermostability of Rhizopus oryzae lipase by combined mutation of hot-spots and engineering a disulfide bond. RSC Adv. 2018, 8, 41247–41254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farias, R.N.; Torres, M.; Canela, R. Spectrophotometric determination of the positional specificity of nonspecific and 1,3-specific lipases. Anal. Biochem. 1997, 252, 186–189. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Sumbalova, L.; Stourac, J.; Martinek, T.; Bednar, D.; Damborsky, J. HotSpot Wizard 3.0: Web server for automated design of mutations and smart libraries based on sequence input information. Nucleic Acids Res. 2018, 46, W356–W362. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Heterologous Protein Expression in Pichia pastoris: Latest Research Progress and Applications. Chembiochem 2018, 19, 7–21. [Google Scholar] [CrossRef]
- Obst, U.; Lu, T.K.; Sieber, V. A Modular Toolkit for Generating Pichia pastoris Secretion Libraries. ACS Synth. Biol. 2017, 6, 1016–1025. [Google Scholar] [CrossRef]
- Prielhofer, R.; Barrero, J.J.; Steuer, S.; Gassler, T.; Zahrl, R.J.; Baumann, K.; Sauer, M.; Mattanovich, D.; Gasser, B.; Marx, H. GoldenPiCS: A Golden Gate-derived modular cloning system for applied synthetic biology in the yeast Pichia pastoris. BMC Syst. Biol. 2017, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fang, X.; Su, F.; Chen, Y.; Xu, L.; Yan, Y. Enhancing the Thermostability of Rhizomucor miehei Lipase with a Limited Screening Library by Rational-Design Point Mutations and Disulfide Bonds. Appl. Environ. Microbiol. 2018, 84, e02129-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, P.; Holmberg, K.; Watzke, H.; Leser, M.E.; Miller, R. Lipases at interfaces: A review. Adv. Colloid Interface Sci. 2009, 147–148, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Delic, M.; Valli, M.; Graf, A.B.; Pfeffer, M.; Mattanovich, D.; Gasser, B. The secretory pathway: Exploring yeast diversity. FEMS Microbiol. Rev. 2013, 37, 872–914. [Google Scholar] [CrossRef] [Green Version]
- Beer, H.D.; McCarthy, J.E.; Bornscheuer, U.T.; Schmid, R.D. Cloning, expression, characterization and role of the leader sequence of a lipase from Rhizopus oryzae. Biochim. Biophys. Acta 1998, 1399, 173–180. [Google Scholar] [CrossRef]
- Satomura, A.; Kuroda, K.; Ueda, M. Generation of a Functionally Distinct Rhizopus oryzae Lipase through Protein Folding Memory. PLoS ONE 2015, 10, e0124545. [Google Scholar] [CrossRef] [Green Version]
- Cowan, D.A. Thermophilic proteins: Stability and function in aqueous and organic solvents. Comp. Biochem. Physiol. A Physiol. 1997, 118, 429–438. [Google Scholar] [CrossRef]
- Baldascini, H.; Janssen, D.B. Interfacial inactivation of epoxide hydrolase in a two-liquid-phase system. Enzym. Microb. Technol. 2005, 36, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Maffucci, I.; Laage, D.; Sterpone, F.; Stirnemann, G. Thermal Adaptation of Enzymes: Impacts of Conformational Shifts on Catalytic Activation Energy and Optimum Temperature. Chemistry 2020, 26, 10045–10056. [Google Scholar] [CrossRef]
- Chow, J.Y.; Shi Choo, K.L.; Lim, Y.P.; Ling, L.H.; Truc Nguyen, G.K.; Xue, B.; Chua, N.H.; Yew, W.S. Scalable Workflow for Green Manufacturing: Discovery of Bacterial Lipases for Biodiesel Production. ACS Sustain. Chem. Eng. 2021, 9, 13450–13459. [Google Scholar] [CrossRef]
- Loh, J.M.; Amelia; Gourich, W.; Chew, C.L.; Song, C.P.; Chan, E.-S. Improved biodiesel production from sludge palm oil catalyzed by a low-cost liquid lipase under low-input process conditions. Renew. Energy 2021, 177, 348–358. [Google Scholar] [CrossRef]
- Ketzer, F.; Wancura, J.H.C.; Tres, M.V.; de Oliveira, J.V. Kinetic and thermodynamic study of enzymatic hydroesterification mechanism to fatty acid methyl esters synthesis. Bioresour. Technol. 2022, 356, 127335. [Google Scholar] [CrossRef] [PubMed]
- Korman, T.P.; Sahachartsiri, B.; Charbonneau, D.M.; Huang, G.L.; Beauregard, M.; Bowie, J.U. Dieselzymes: Development of a stable and methanol tolerant lipase for biodiesel production by directed evolution. Biotechnol. Biofuels 2013, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehrmann, M.; Klebensberger, J. Engineering thermal stability and solvent tolerance of the soluble quinoprotein PedE from Pseudomonas putida KT2440 with a heterologous whole-cell screening approach. Microb. Biotechnol. 2018, 11, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Yang, L.; Wang, Z.; Lv, P.; Fu, J.; Miao, C.; Li, M.; Liu, T.; Luo, W. Improved methanol tolerance of Rhizomucor miehei lipase based on N-glycosylation within the α-helix region and its application in biodiesel production. Biotechnol. Biofuels 2021, 14, 237. [Google Scholar] [CrossRef]
- He, D.; Luo, W.; Wang, Z.; Lv, P.; Yuan, Z.; Huang, S.; Xv, J. Establishment and application of a modified membrane-blot assay for Rhizomucor miehei lipases aimed at improving their methanol tolerance and thermostability. Enzym. Microb. Technol. 2017, 102, 35–40. [Google Scholar] [CrossRef]
- Batra, J.; Mishra, S. Organic solvent tolerance and thermostability of a β-glucosidase co-engineered by random mutagenesis. J. Mol. Catal. B Enzym. 2013, 96, 61–66. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; De Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Iyer, B.R.; Mahalakshmi, R. Hydrophobic Characteristic Is Energetically Preferred for Cysteine in a Model Membrane Protein. Biophys. J. 2019, 117, 25–35. [Google Scholar] [CrossRef] [Green Version]
- You, C.; Huang, Q.; Xue, H.; Xu, Y.; Lu, H. Potential hydrophobic interaction between two cysteines in interior hydrophobic region improves thermostability of a family 11 xylanase from Neocallimastix patriciarum. Biotechnol. Bioeng. 2010, 105, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, S.; Kar, T.; Pattanayak, J. Comparison of various types of hydrogen bonds involving aromatic amino acids. J. Am. Chem. Soc. 2002, 124, 13257–13264. [Google Scholar] [CrossRef] [PubMed]
- Andualema, B.; Andualema, B.; Gessesse, A. Microbial Lipases and Their Industrial Applications: Review. Biotechnology 2012, 11, 100–118. [Google Scholar] [CrossRef] [Green Version]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 st Round Mutagenesis * | 2 nd Round Mutagenesis ** | ||
|---|---|---|---|
| A8E | Y187N | A7R | V175F |
| S31D | G193K | L113Y | G177C |
| S114N | Q197H | Q150F | T199V |
| N134Y | Q197Y | Q150H | F216Y |
| T136D | |||
| P163K | |||
| F173Y | S267L | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chow, J.Y.; Nguyen, G.K.T. Rational Design of Lipase ROL to Increase Its Thermostability for Production of Structured Tags. Int. J. Mol. Sci. 2022, 23, 9515. https://doi.org/10.3390/ijms23179515
Chow JY, Nguyen GKT. Rational Design of Lipase ROL to Increase Its Thermostability for Production of Structured Tags. International Journal of Molecular Sciences. 2022; 23(17):9515. https://doi.org/10.3390/ijms23179515
Chicago/Turabian StyleChow, Jeng Yeong, and Giang Kien Truc Nguyen. 2022. "Rational Design of Lipase ROL to Increase Its Thermostability for Production of Structured Tags" International Journal of Molecular Sciences 23, no. 17: 9515. https://doi.org/10.3390/ijms23179515
APA StyleChow, J. Y., & Nguyen, G. K. T. (2022). Rational Design of Lipase ROL to Increase Its Thermostability for Production of Structured Tags. International Journal of Molecular Sciences, 23(17), 9515. https://doi.org/10.3390/ijms23179515