
Evaluation of the Effect of the Fibroblast Growth Factor Type 2 (FGF-2) Administration on Placental Gene Expression in a Murine Model of Preeclampsia Induced by L-NAME

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
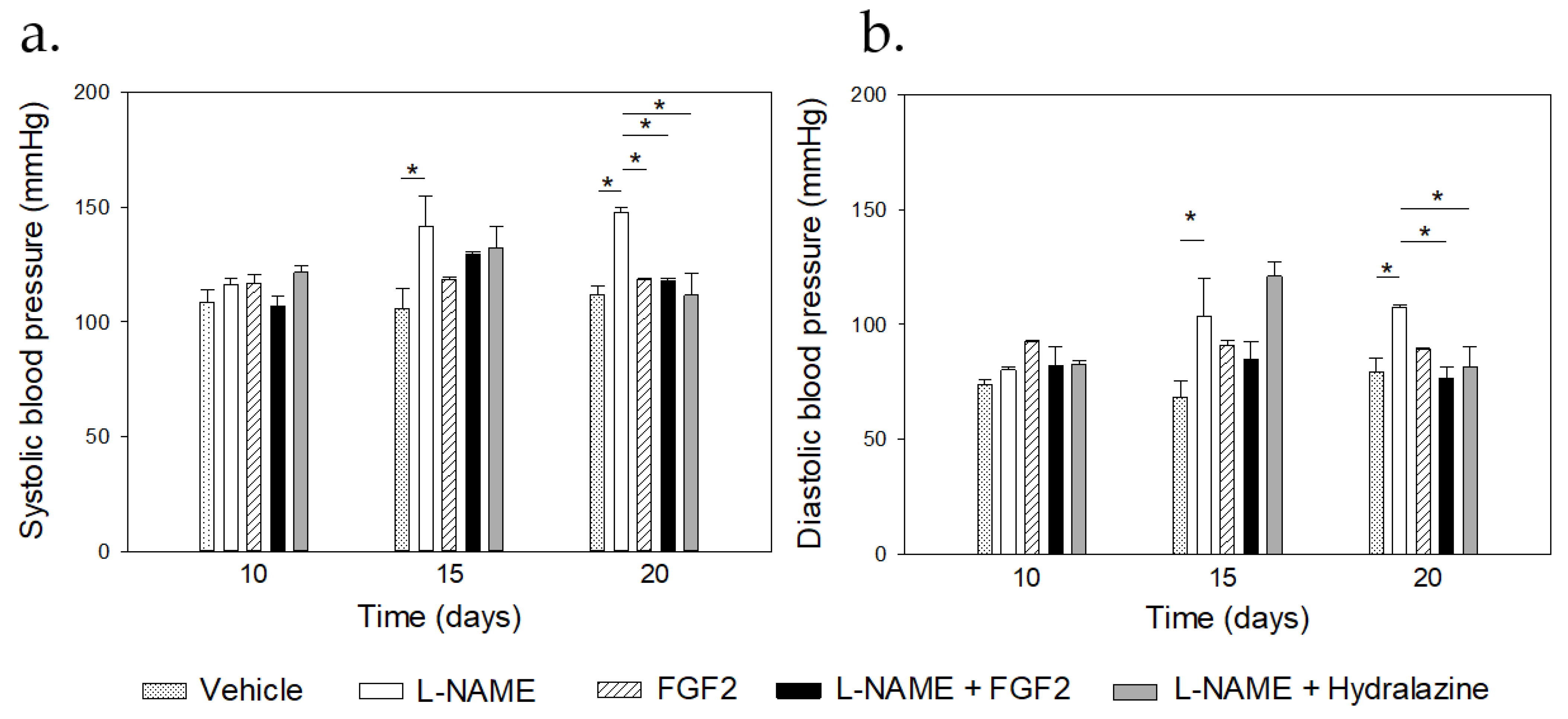
2.1. Modulatory Effect of FGF2 on BP Values and Urine Protein Concentration in the Rat PE-like Model Induced by L-NAME
2.2. FGF2 Administration Induced Changes in Placental Weight in the Rat PE-like Model Induced by L-NAME
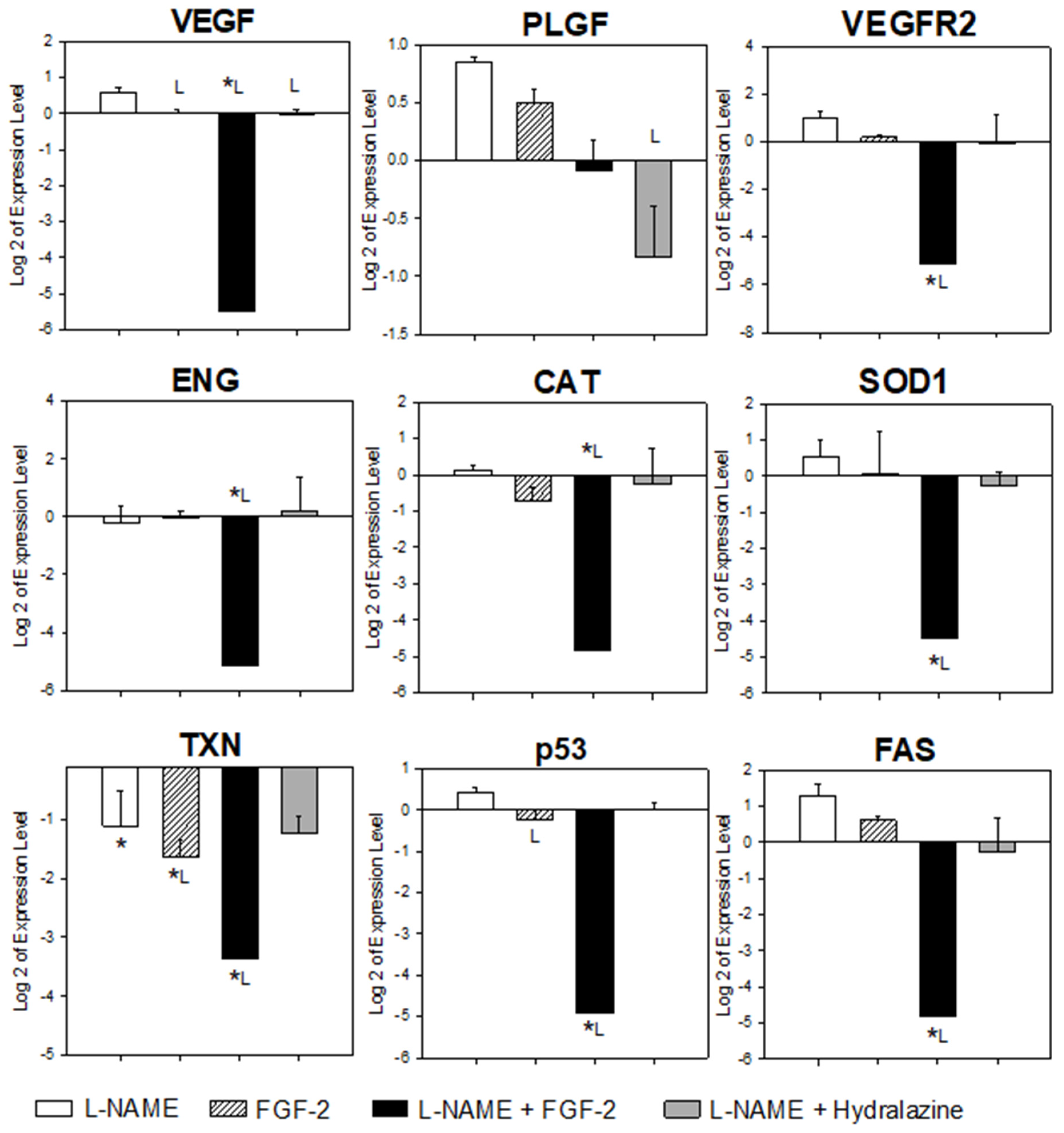
2.3. FGF-2 Administration Modulated the Placental Gene Expression in the Murine PE-like Model Induced by L-NAME
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Animal Treatment
4.3. Biological Samples and Data Collection
4.4. Placental RNA Isolation and cDNA Synthesis
4.5. Gene Selection and Primer Design
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia-Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, J.E.; Fraser, R.; Leslie, K.; Wallace, A.E.; James, J.L. Remodelling at the maternal-fetal interface: Relevance to human pregnancy disorders. Reproduction 2010, 140, 803–813. [Google Scholar] [CrossRef]
- Wu, F.; Tian, F.J.; Lin, Y. Oxidative Stress in Placenta: Health and Diseases. BioMed Res. Int. 2015, 2015, 293271. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Karumanchi, S.A. Angiogenic factors and preeclampsia. Semin. Nephrol. 2011, 31, 33–46. [Google Scholar] [CrossRef]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn Care Res. Off. Publ. Am. Burn Assoc. 2010, 31, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Wanjare, M.; Kusuma, S.; Gerecht, S. Perivascular cells in blood vessel regeneration. Biotechnol. J. 2013, 8, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Poto, R.; Cristinziano, L.; Modestino, L.; de Paulis, A.; Marone, G.; Loffredo, S.; Galdiero, M.R.; Varricchi, G. Neutrophil Extracellular Traps, Angiogenesis and Cancer. Biomedicines 2022, 10, 431. [Google Scholar] [CrossRef]
- Pereira, R.D.; De Long, N.E.; Wang, R.C.; Yazdi, F.T.; Holloway, A.C.; Raha, S. Angiogenesis in the placenta: The role of reactive oxygen species signaling. BioMed Res. Int. 2015, 2015, 814543. [Google Scholar] [CrossRef]
- Munaut, C.; Lorquet, S.; Pequeux, C.; Coulon, C.; Le Goarant, J.; Chantraine, F.; Noel, A.; Goffin, F.; Tsatsaris, V.; Subtil, D.; et al. Differential expression of Vegfr-2 and its soluble form in preeclampsia. PLoS ONE 2012, 7, e33475. [Google Scholar] [CrossRef]
- Silvestro, S.; Calcaterra, V.; Pelizzo, G.; Bramanti, P.; Mazzon, E. Prenatal Hypoxia and Placental Oxidative Stress: Insights from Animal Models to Clinical Evidences. Antioxidants 2020, 9, 414. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-induced angiogenesis: Good and evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fierro, M.L.; Hernadez-Delgadillo, G.P.; Flores-Mendoza, J.F.; Alvarez-Zuniga, C.D.; Diaz-Lozano, M.L.; Delgado-Enciso, I.; Romero-Diaz, V.J.; Lopez-Saucedo, A.; Rodriguez-Sanchez, I.P.; Marino-Martinez, I.A.; et al. Fibroblast Growth Factor Type 2 (FGF2) Administration Attenuated the Clinical Manifestations of Preeclampsia in a Murine Model Induced by L-NAME. Front. Pharmacol. 2021, 12, 663044. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Yonemitsu, Y.; Onimaru, M.; Inoue, M.; Hasegawa, M.; Kuwano, H.; Sueishi, K. VEGF function for upregulation of endogenous PlGF expression during FGF-2-mediated therapeutic angiogenesis. Atherosclerosis 2008, 200, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Fierro, M.L.; Garza-Veloz, I.; Castruita-Dela Rosa, C.; Ortiz-Castro, Y.; Aceves-Medina, M.C.; Vazquez-Castro, R.; Delgado-Enciso, I.; Castaneda-Lopez, M.E. Plasma cancer biomarker multiplex screening and the risk of subsequent preeclampsia. Int. J. Cardiol. 2015, 179, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Lieu, C.; Heymach, J.; Overman, M.; Tran, H.; Kopetz, S. Beyond VEGF: Inhibition of the fibroblast growth factor pathway and antiangiogenesis. Clin. Cancer Res. 2011, 17, 6130–6139. [Google Scholar] [CrossRef]
- Kienast, Y.; Klein, C.; Scheuer, W.; Raemsch, R.; Lorenzon, E.; Bernicke, D.; Herting, F.; Yu, S.; The, H.H.; Martarello, L.; et al. Ang-2-VEGF-A CrossMab, a novel bispecific human IgG1 antibody blocking VEGF-A and Ang-2 functions simultaneously, mediates potent antitumor, antiangiogenic, and antimetastatic efficacy. Clin. Cancer Res. 2013, 19, 6730–6740. [Google Scholar] [CrossRef]
- Baijnath, S.; Soobryan, N.; Mackraj, I.; Gathiram, P.; Moodley, J. The optimization of a chronic nitric oxide synthase (NOS) inhibition model of pre-eclampsia by evaluating physiological changes. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 182, 71–75. [Google Scholar] [CrossRef]
- Soobryan, N.; Murugesan, S.; Phoswa, W.; Gathiram, P.; Moodley, J.; Mackraj, I. The effects of sildenafil citrate on uterine angiogenic status and serum inflammatory markers in an L-NAME rat model of pre-eclampsia. Eur. J. Pharmacol. 2017, 795, 101–107. [Google Scholar] [CrossRef]
- Pfeiffer, S.; Leopold, E.; Schmidt, K.; Brunner, F.; Mayer, B. Inhibition of nitric oxide synthesis by NG-nitro-L-arginine methyl ester (L-NAME): Requirement for bioactivation to the free acid, NG-nitro-L-arginine. Br. J. Pharmacol. 1996, 118, 1433–1440. [Google Scholar] [CrossRef]
- Alshami, A.; Romero, C.; Avila, A.; Varon, J. Management of hypertensive crises in the elderly. J. Geriatr. Cardiol. JGC 2018, 15, 504–512. [Google Scholar] [CrossRef]
- Tassi, E.; Lai, E.Y.; Li, L.; Solis, G.; Chen, Y.; Kietzman, W.E.; Ray, P.E.; Riegel, A.T.; Welch, W.J.; Wilcox, C.S.; et al. Blood Pressure Control by a Secreted FGFBP1 (Fibroblast Growth Factor-Binding Protein). Hypertension 2018, 71, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Airoldi, J.; Weinstein, L. Clinical significance of proteinuria in pregnancy. Obstet. Gynecol. Surv. 2007, 62, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.J.; Gao, L.L. Effect of epidural block on 24-hour urine protein in pregnant rat models with preeclampsia. Arch. Gynecol. Obstet. 2012, 286, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.J.d.C.; Silva, A.C.O.; Frazão, I.d.S.; Perrelli, J.G.A.; Silva, T.T.d.M.; Cavalcanti, A.M. Family functionality and burden of family caregivers of users with mental disorders. Rev. Bras. Enferm. 2021, 74, e20201061. [Google Scholar] [CrossRef]
- Can, M.; Guven, B.; Bektas, S.; Arikan, I. Oxidative stress and apoptosis in preeclampsia. Tissue Cell 2014, 46, 477–481. [Google Scholar] [CrossRef]
- Dulak, J.; Jozkowicz, A.; Dembinska-Kiec, A.; Guevara, I.; Zdzienicka, A.; Zmudzinska-Grochot, D.; Florek, I.; Wojtowicz, A.; Szuba, A.; Cooke, J.P. Nitric oxide induces the synthesis of vascular endothelial growth factor by rat vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 659–666. [Google Scholar] [CrossRef]
- Mortensen, K.; Skouv, J.; Hougaard, D.M.; Larsson, L.I. Endogenous endothelial cell nitric-oxide synthase modulates apoptosis in cultured breast cancer cells and is transcriptionally regulated by p53. J. Biol. Chem. 1999, 274, 37679–37684. [Google Scholar] [CrossRef]
- Silva Carmona, A.; Mendieta Zeron, H. NF-kappaBeta and SOD expression in preeclamptic placentas. Turk. J. Med. Sci. 2016, 46, 783–788. [Google Scholar] [CrossRef]
- Enquobahrie, D.A.; Meller, M.; Rice, K.; Psaty, B.M.; Siscovick, D.S.; Williams, M.A. Differential placental gene expression in preeclampsia. Am. J. Obstet. Gynecol. 2008, 199, 566-e1. [Google Scholar] [CrossRef]
- Hansson, S.R.; Chen, Y.; Brodszki, J.; Chen, M.; Hernandez-Andrade, E.; Inman, J.M.; Kozhich, O.A.; Larsson, I.; Marsal, K.; Medstrand, P.; et al. Gene expression profiling of human placentas from preeclamptic and normotensive pregnancies. Mol. Hum. Reprod. 2006, 12, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Tandon, V.; Hiwale, S.; Amle, D.; Nagaria, T.; Patra, P.K. Assessment of Serum Vascular Endothelial Growth Factor Levels in Pregnancy-Induced Hypertension Patients. J. Pregnancy 2017, 2017, 3179670. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Ishikawa, W.; Kushima, T.; Nishimura, T.; Mori, C.; Onuki, A.; Suzuki, T.; Ishii, Y.; Kansaku, N.; Miyazaki, Y.; et al. Nitric oxide induces vascular endothelial growth factor expression in the rat placenta in vivo and in vitro. Biosci. Biotechnol. Biochem. 2013, 77, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, K.; Watanabe Miyano, S.; Minoshima, Y.; Matsui, J.; Funahashi, Y. Activated FGF2 signaling pathway in tumor vasculature is essential for acquired resistance to anti-VEGF therapy. Sci. Rep. 2020, 10, 2939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Neiva, K.G.; Lingen, M.W.; Ellis, L.M.; Nor, J.E. VEGF-dependent tumor angiogenesis requires inverse and reciprocal regulation of VEGFR1 and VEGFR2. Cell Death Differ. 2010, 17, 499–512. [Google Scholar] [CrossRef]
- Seghezzi, G.; Patel, S.; Ren, C.J.; Gualandris, A.; Pintucci, G.; Robbins, E.S.; Shapiro, R.L.; Galloway, A.C.; Rifkin, D.B.; Mignatti, P. Fibroblast growth factor-2 (FGF-2) induces vascular endothelial growth factor (VEGF) expression in the endothelial cells of forming capillaries: An autocrine mechanism contributing to angiogenesis. J. Cell Biol. 1998, 141, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Nakatsukasa, Y.; Tsukahara, H.; Tabuchi, K.; Tabuchi, M.; Magami, T.; Yamada, M.; Fujii, Y.; Yashiro, M.; Tsuge, M.; Morishima, T. Thioredoxin-1 and oxidative stress status in pregnant women at early third trimester of pregnancy: Relation to maternal and neonatal characteristics. J. Clin. Biochem. Nutr. 2013, 52, 27–31. [Google Scholar] [CrossRef][Green Version]
- Sahay, A.S.; Sundrani, D.P.; Wagh, G.N.; Mehendale, S.S.; Joshi, S.R. Regional differences in the placental levels of oxidative stress markers in pre-eclampsia. Int. J. Gynaecol. Obstet. Off. Organ. Int. Fed. Gynaecol. Obstet. 2015, 129, 213–218. [Google Scholar] [CrossRef]
- Myatt, L.; Eis, A.L.; Brockman, D.E.; Kossenjans, W.; Greer, I.A.; Lyall, F. Differential localization of superoxide dismutase isoforms in placental villous tissue of normotensive, pre-eclamptic, and intrauterine growth-restricted pregnancies. J. Histochem. Cytochem. 1997, 45, 1433–1438. [Google Scholar] [CrossRef]
- Roland, L.; Beauchemin, D.; Acteau, G.; Fradette, C.; St-Pierre, I.; Bilodeau, J.F. Effects of labor on placental expression of superoxide dismutases in preeclampsia. Placenta 2010, 31, 392–400. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, S.W. Antioxidant activities and mRNA expression of superoxide dismutase, catalase, and glutathione peroxidase in normal and preeclamptic placentas. J. Soc. Gynecol. Investig. 1996, 3, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Perkins, A.V. Endogenous anti-oxidants in pregnancy and preeclampsia. Aust. N. Z. J. Obstet. Gynaecol. 2006, 46, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Wiktor, H.; Kankofer, M. Catalase activity in normal and preeclamptic placentas. Ginekol. Pol. 2001, 72, 1228–1232. [Google Scholar] [PubMed]
- Krishna, T.S.; Rao, E.V.; Naidu, J.N. Alterations of antioxidant enzymes in preeclampsia. IJRMS 2015, 3, 629. [Google Scholar] [CrossRef][Green Version]
- Glorieux, C.; Zamocky, M.; Sandoval, J.M.; Verrax, J.; Calderon, P.B. Regulation of catalase expression in healthy and cancerous cells. Free Radic. Biol. Med. 2015, 87, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Wei, J.X.; Xue, S.F.; Liu, G.H.; Fu, L.X. Fibrogrowth factor-2 protects against acute lung injury by activating the PI3K/Akt signaling pathway. J. Biol. Regul. Homeost. Agents 2020, 34, 1679–1688. [Google Scholar] [CrossRef]
- Benhar, M. Nitric oxide and the thioredoxin system: A complex interplay in redox regulation. Biochim. Biophys. Acta 2015, 1850, 2476–2484. [Google Scholar] [CrossRef]
- Sahlin, L.; Ostlund, E.; Wang, H.; Holmgren, A.; Fried, G. Decreased expression of thioredoxin and glutaredoxin in placentae from pregnancies with pre-eclampsia and intrauterine growth restriction. Placenta 2000, 21, 603–609. [Google Scholar] [CrossRef]
- Wei, D.; Wu, Q.; Shi, H. Apoptosis and p53 expression in the placental villi of females with unexplained recurrent spontaneous abortion. Exp. Ther. Med. 2014, 7, 191–194. [Google Scholar] [CrossRef]
- Sohr, S.; Engeland, K. The tumor suppressor p53 induces expression of the pregnancy-supporting human chorionic gonadotropin (hCG) CGB7 gene. Cell Cycle 2011, 10, 3758–3767. [Google Scholar] [CrossRef]
- Abrahams, V.M.; Straszewski-Chavez, S.L.; Guller, S.; Mor, G. First trimester trophoblast cells secrete Fas ligand which induces immune cell apoptosis. Mol. Hum. Reprod. 2004, 10, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Levy, R. The role of apoptosis in preeclampsia. Isr Med. Assoc. J. 2005, 7, 178–181. [Google Scholar] [PubMed]
- Baserga, M.; Hale, M.A.; Ke, X.; Wang, Z.M.; Yu, X.; Callaway, C.W.; McKnight, R.A.; Lane, R.H. Uteroplacental insufficiency increases p53 phosphorylation without triggering the p53-MDM2 functional circuit response in the IUGR rat kidney. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R412–R418. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Song, J.; Rutherford, T.; Naftolin, F.; Brown, S.; Mor, G. Hormonal regulation of apoptosis and the Fas and Fas ligand system in human endometrial cells. Mol. Hum. Reprod. 2002, 8, 447–455. [Google Scholar] [CrossRef]
- Lan, R.; Yang, Y.; Song, J.; Wang, L.; Gong, H. Fas regulates the apoptosis and migration of trophoblast cells by targeting NF-κB. Exp. Ther. Med. 2021, 22, 1055. [Google Scholar] [CrossRef]
- Neale, D.M.; Mor, G. The role of Fas mediated apoptosis in preeclampsia. J. Perinat. Med. 2005, 33, 471–477. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Bank ID | Description | * Primer Sequence (5′→3′) | Tm | Product Size (bp) |
|---|---|---|---|---|---|
| VEGFA | NM_031836.3 | Vascular Endothelial Growth Factor A | Fw: GGAGCAGAAAGCCCATGAAGTGGT Rv: TCATTGCAGCAGCCCGCACA | 65 | 168 |
| VEGFR2 | NM_013062.2 | Vascular Endothelial Growth Factor Receptor 2 | Fw: TTTGCACTGCAGGAGCGCGT Rv: GGAATCGCCAGGCAAACCCACA | 65 | 171 |
| ENG | NM_001010968.3 | Endoglin | Fw: CAGGGCTTCGTACAGGTGAGCA Rv: TCACACAGCTGCCCTTGGCT | 64 | 139 |
| P53 | NM_030989.3 | Tumor Protein P53 | Fw: GTTGCTCTGATGGTGACGGCCT Rv: ACCACCACGCTGTGCCGAAA | 65 | 112 |
| BCL2 | NM_016993.2 | BCL2 Apoptosis Regulator | Fw: TCCAGGATAACGGAGGCTGGGATGC Rv: AGGCTGAGCAGCGTCTTCAGAGACA | 67 | 103 |
| FAS | NM_139194.3 | Fas Cell Surface Death Receptor | Fw: GTCAACCGTGTCAGCCTGGTGAA Rv: TGGGTCCGGGTGCAGTTCGTTT | 65 | 190 |
| CASP3 | NM_012922.2 | Caspase 3 | Fw: GCGGAGCTTGGAACGCGAAGAAA Rv: TCCAGAGTCCATCGACTTGCTTCCA | 65 | 120 |
| SOD1 | NM_017050.1 | Superoxide Dismutase 1 | Fw: TTCGTTTCCTGCGGCGGCTTCT Rv: GGTTCACCGCTTGCCTTCTGCT | 66 | 169 |
| CAT | NM_012520.2 | Catalase | Fw: GGCACACTTTGACAGAGAGCGGA Rv: TGAGCCTGACTCTCCAGCGACT | 65 | 184 |
| TXN | NM_053800.3 | Thioredoxin | Fw: TCTGCCACGTGGTGTGGACCTT Rv: ACAGTCTGCAGCAACATCCTGGC | 66 | 126 |
| PLGF | NM_053595.2 | Placental Growth Factor | Fw: TGAGGAACCCCACCTGTGATGCT Rv: CATTCAGCAGGGACGAGTTGGCT | 65 | 156 |
| HPRT1 | NM_012583.2 | Hypoxanthine phosphoribosyltransferase 1 | Fw: CAGTCCCAGCGTCGTGATTA Rv: TGGCCTCCCATCTCCTTCAT | 60 | 168 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Fierro, M.L.; Garza-Veloz, I.; Castañeda-Lopez, M.E.; Wasike, D.; Castruita-De la Rosa, C.; Rodriguez-Sanchez, I.P.; Delgado-Enciso, I.; Flores-Mendoza, J. Evaluation of the Effect of the Fibroblast Growth Factor Type 2 (FGF-2) Administration on Placental Gene Expression in a Murine Model of Preeclampsia Induced by L-NAME. Int. J. Mol. Sci. 2022, 23, 10129. https://doi.org/10.3390/ijms231710129
Martinez-Fierro ML, Garza-Veloz I, Castañeda-Lopez ME, Wasike D, Castruita-De la Rosa C, Rodriguez-Sanchez IP, Delgado-Enciso I, Flores-Mendoza J. Evaluation of the Effect of the Fibroblast Growth Factor Type 2 (FGF-2) Administration on Placental Gene Expression in a Murine Model of Preeclampsia Induced by L-NAME. International Journal of Molecular Sciences. 2022; 23(17):10129. https://doi.org/10.3390/ijms231710129
Chicago/Turabian StyleMartinez-Fierro, Margarita L, Idalia Garza-Veloz, Maria Eugenia Castañeda-Lopez, Dorothy Wasike, Claudia Castruita-De la Rosa, Iram Pablo Rodriguez-Sanchez, Ivan Delgado-Enciso, and Jose Flores-Mendoza. 2022. "Evaluation of the Effect of the Fibroblast Growth Factor Type 2 (FGF-2) Administration on Placental Gene Expression in a Murine Model of Preeclampsia Induced by L-NAME" International Journal of Molecular Sciences 23, no. 17: 10129. https://doi.org/10.3390/ijms231710129
APA StyleMartinez-Fierro, M. L., Garza-Veloz, I., Castañeda-Lopez, M. E., Wasike, D., Castruita-De la Rosa, C., Rodriguez-Sanchez, I. P., Delgado-Enciso, I., & Flores-Mendoza, J. (2022). Evaluation of the Effect of the Fibroblast Growth Factor Type 2 (FGF-2) Administration on Placental Gene Expression in a Murine Model of Preeclampsia Induced by L-NAME. International Journal of Molecular Sciences, 23(17), 10129. https://doi.org/10.3390/ijms231710129