
Lumican, an Exerkine, Protects against Skeletal Muscle Loss

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
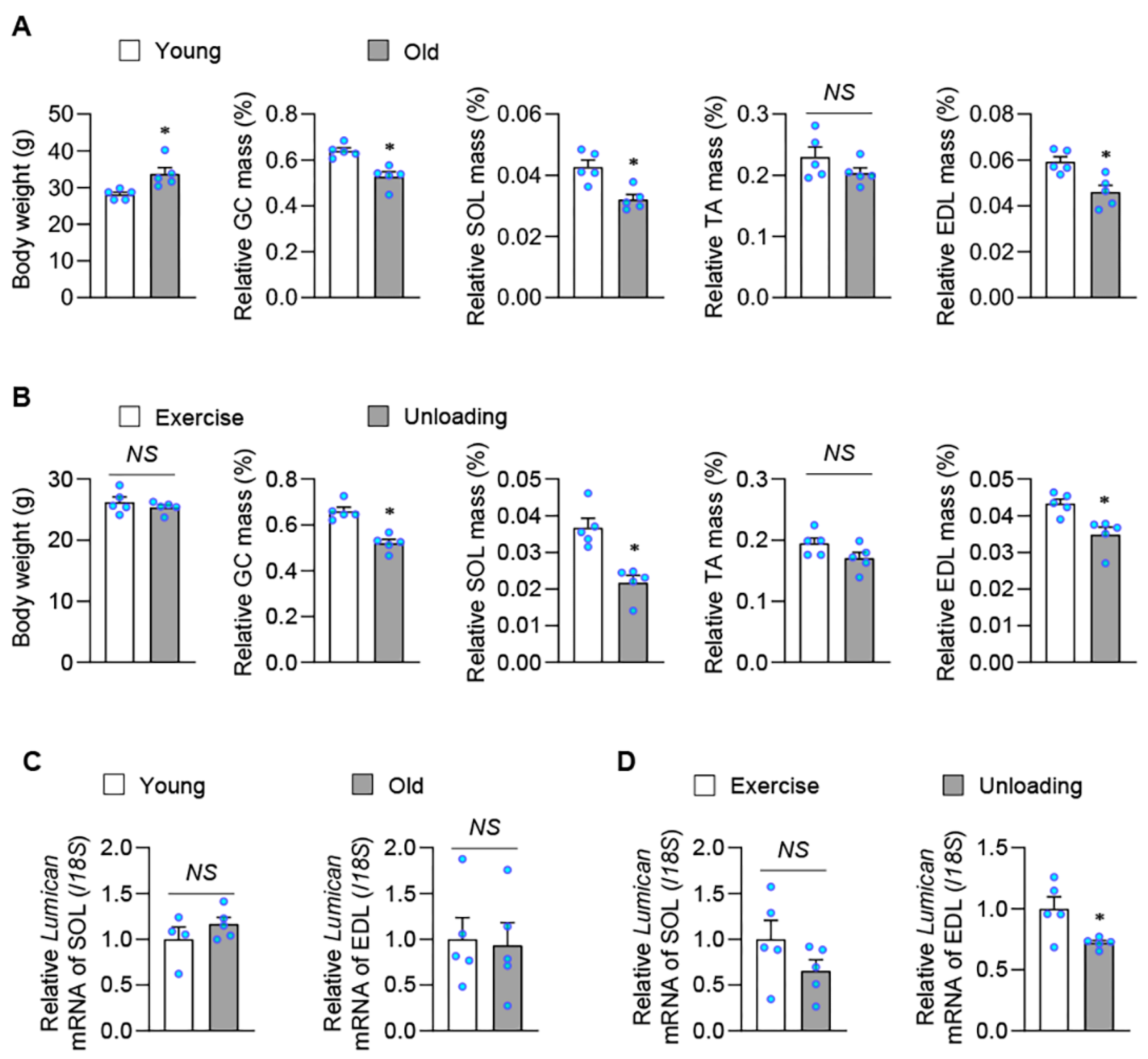
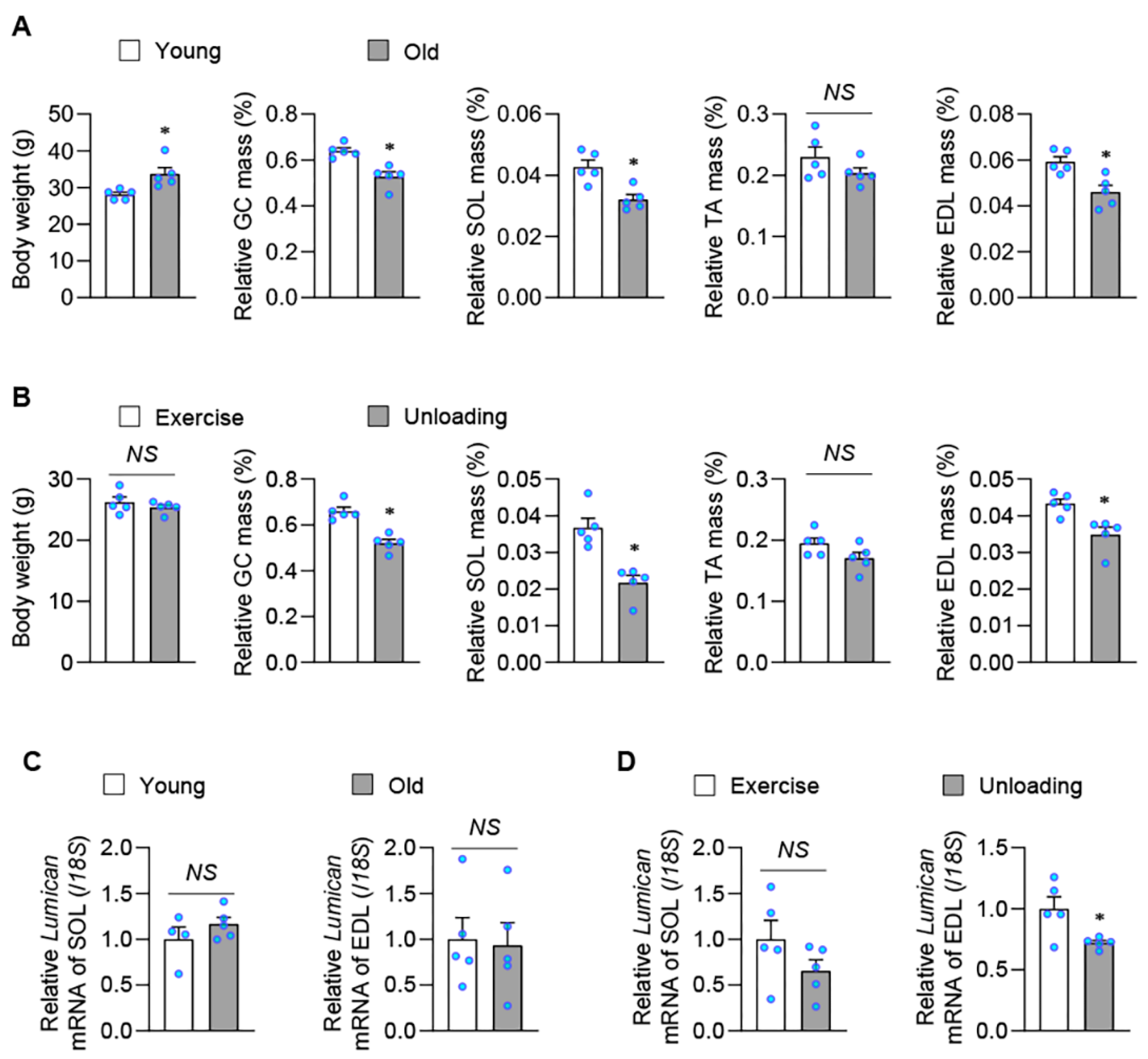
2.1. Exercise Stimulates Lumican Expression in Skeletal Muscles
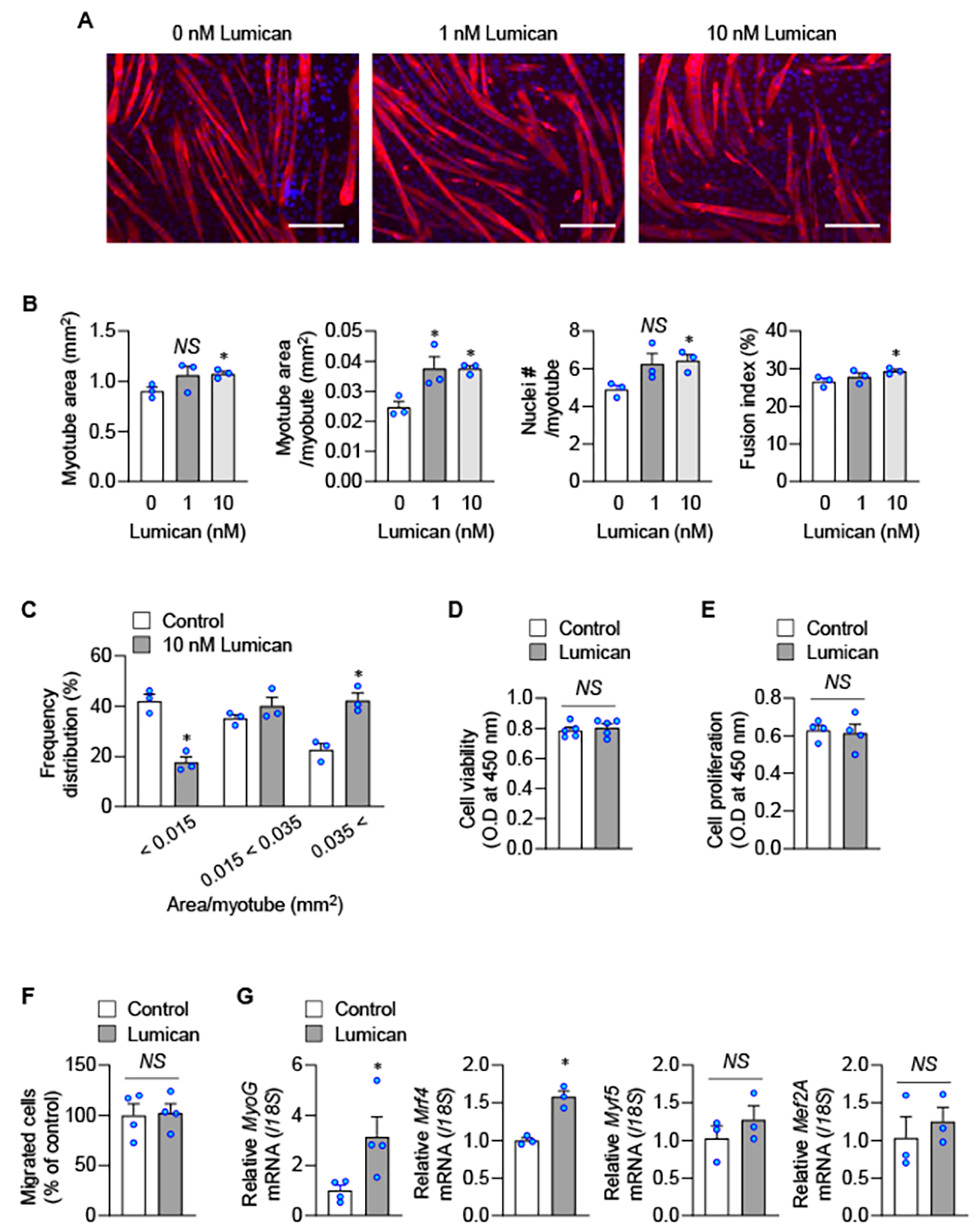
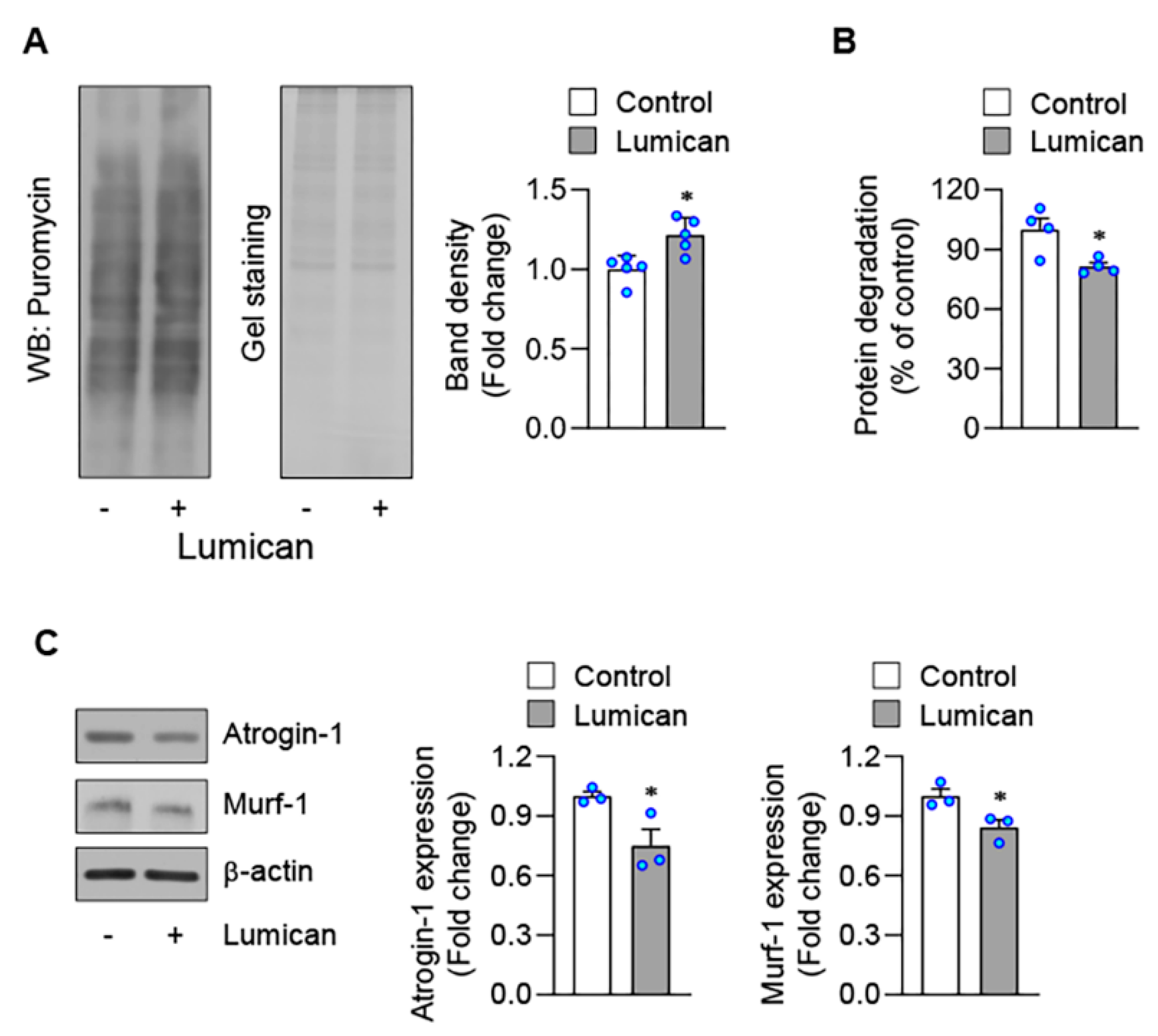
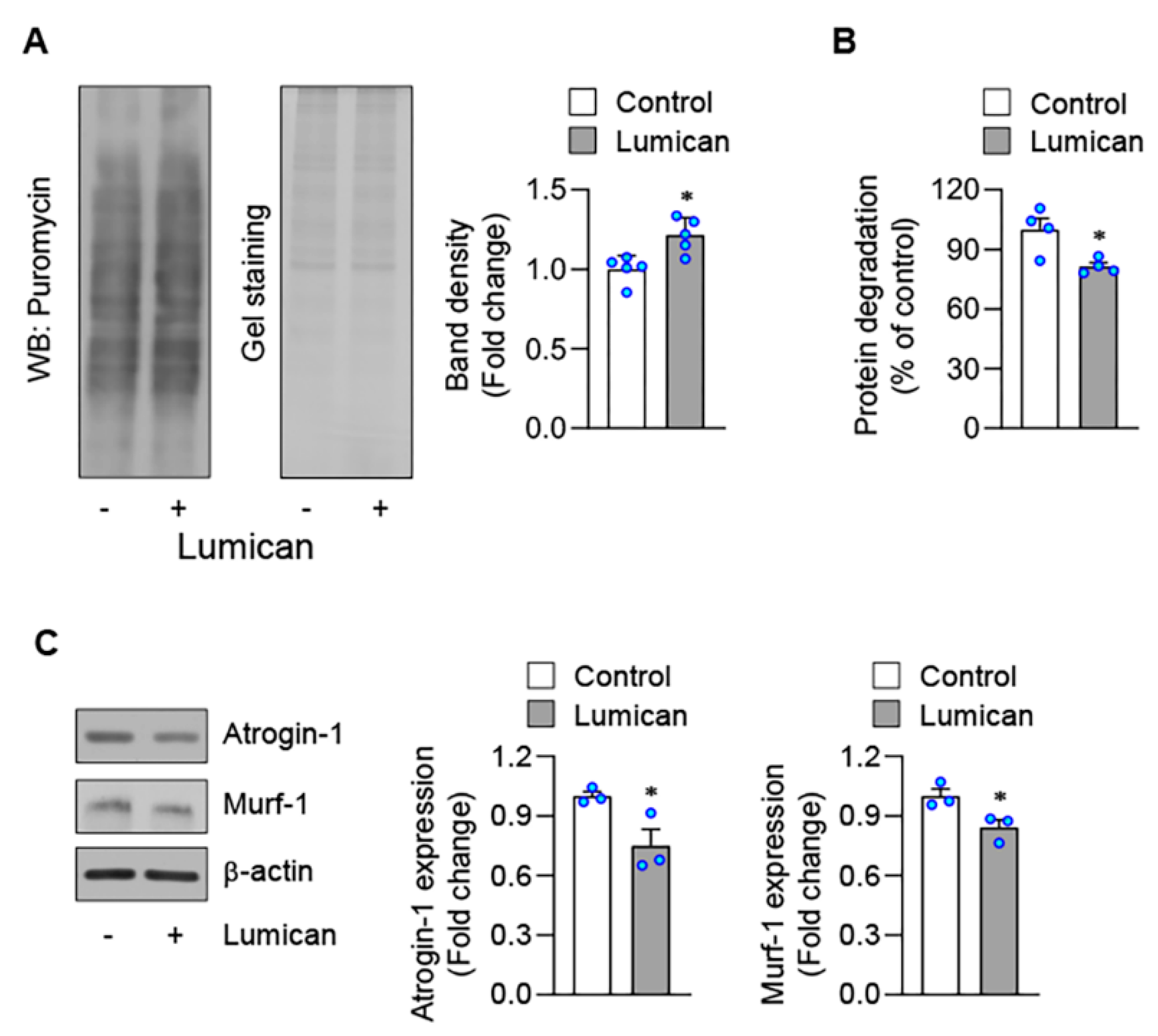
2.2. Lumican Stimulates Myogenesis In Vitro
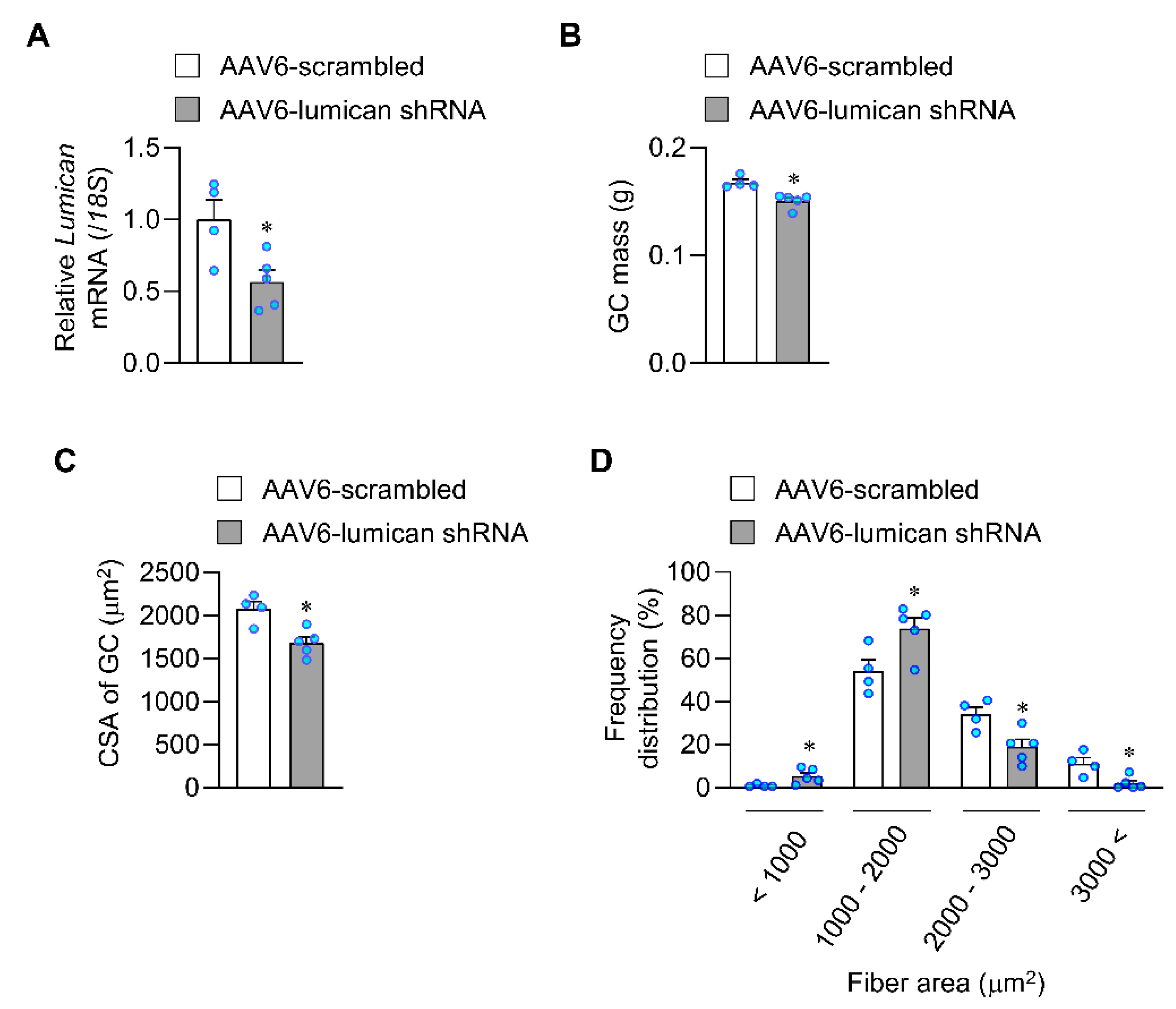
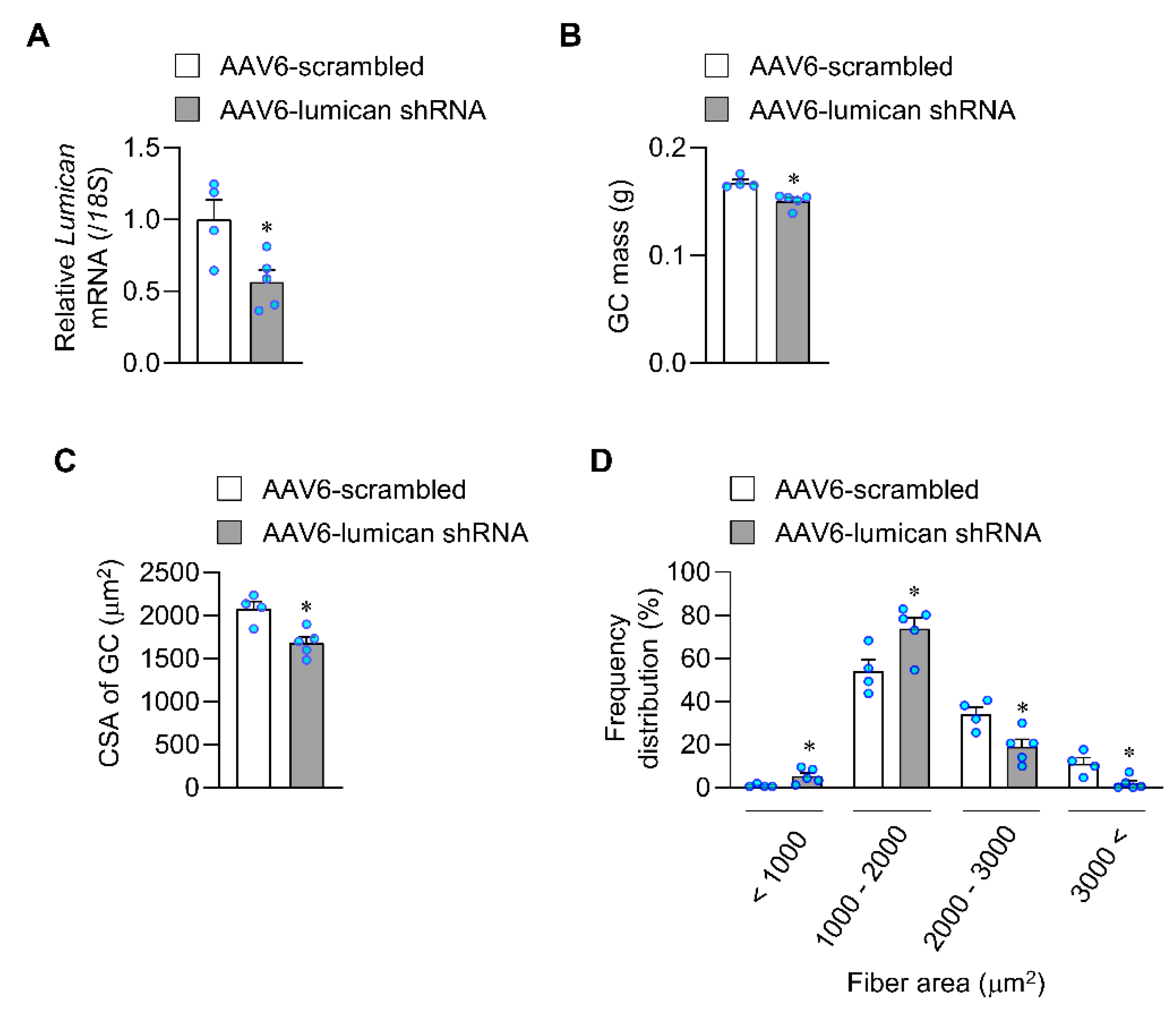
2.3. Lack of Lumican Contributes to Muscle Atrophy in Exercised Mice
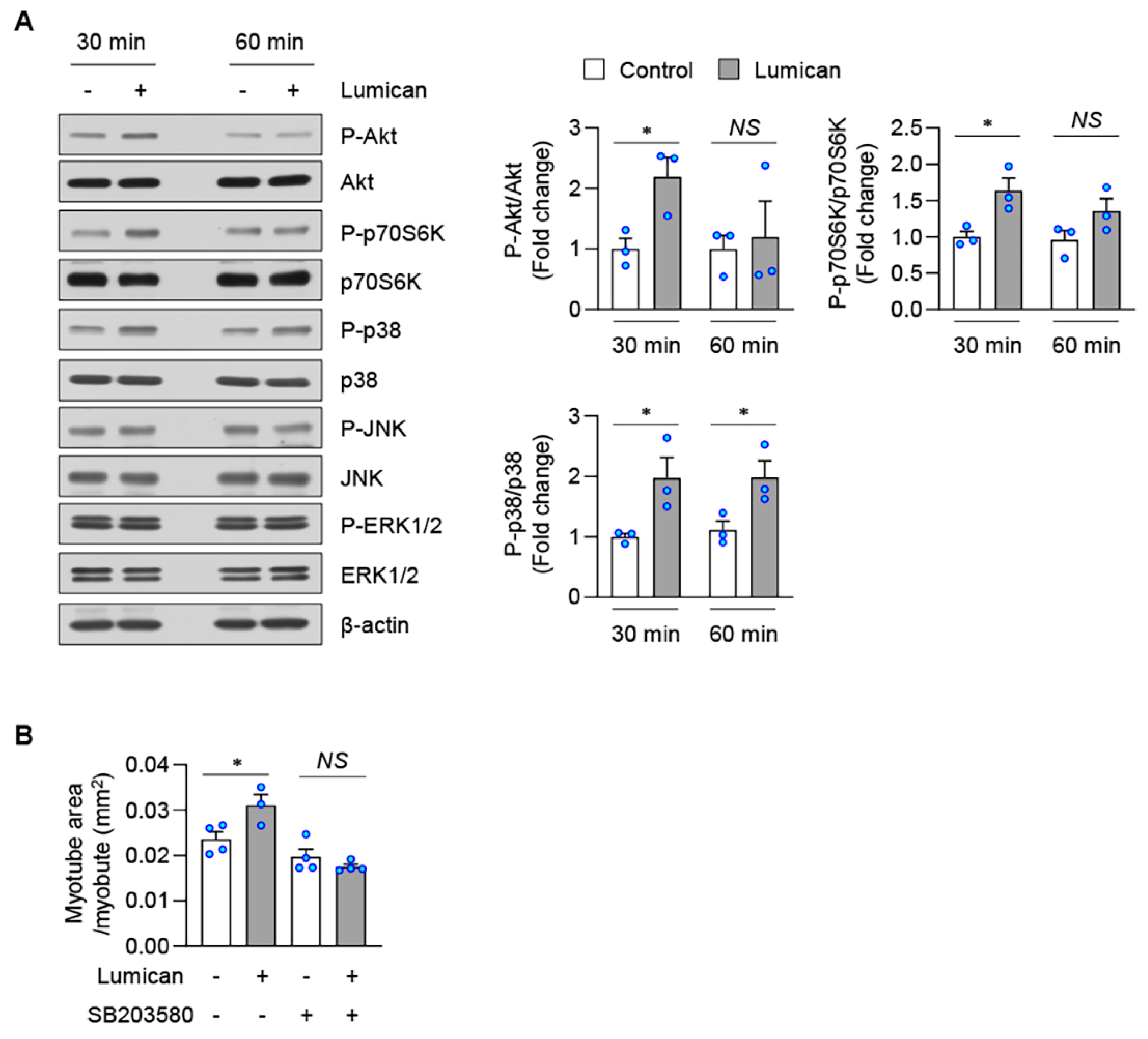
2.4. Lumican Stimulates Myogenesis by Activating the p38 Mitogen-Activated Protein Kinase (MAPK) Signaling Pathway
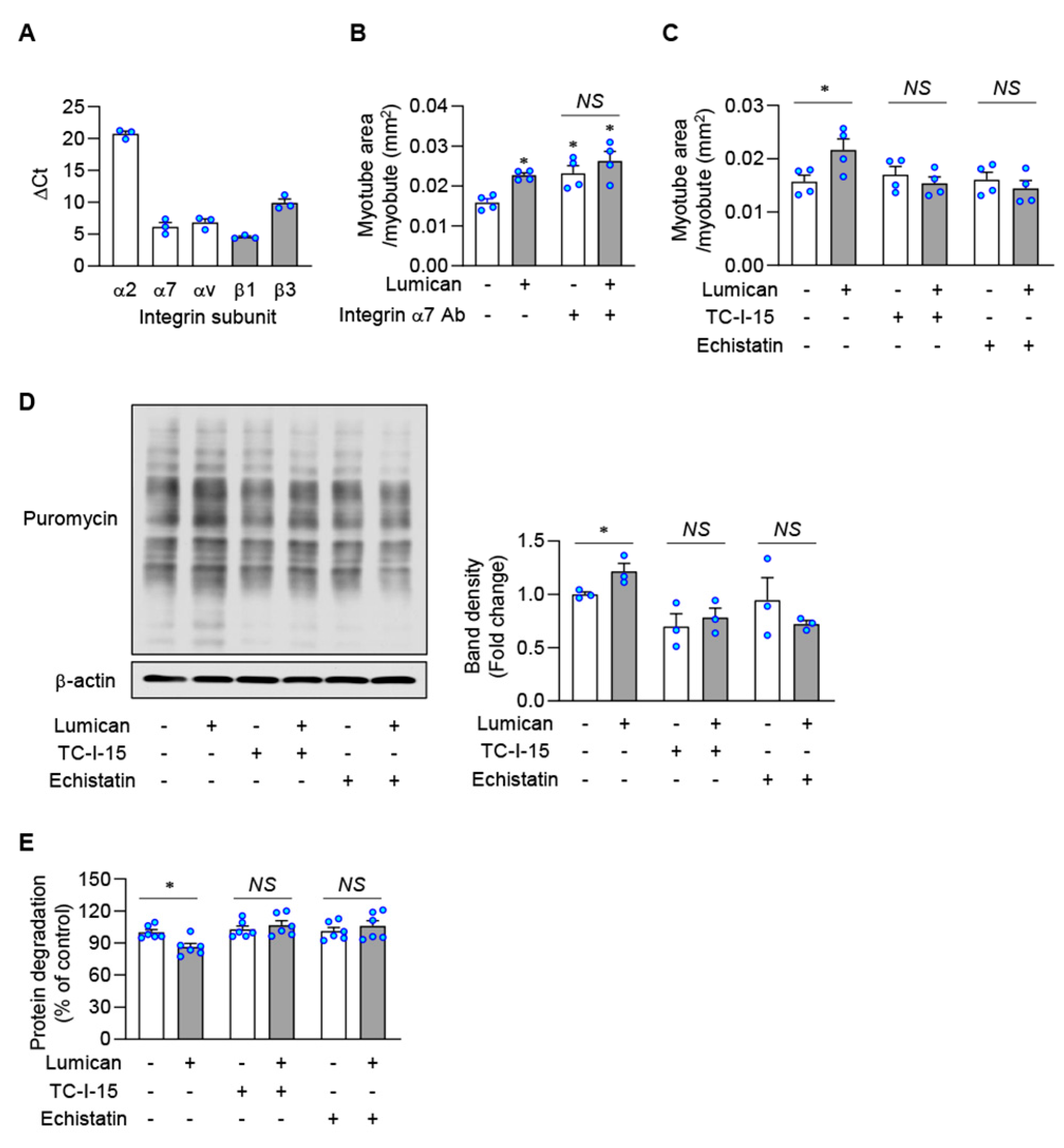
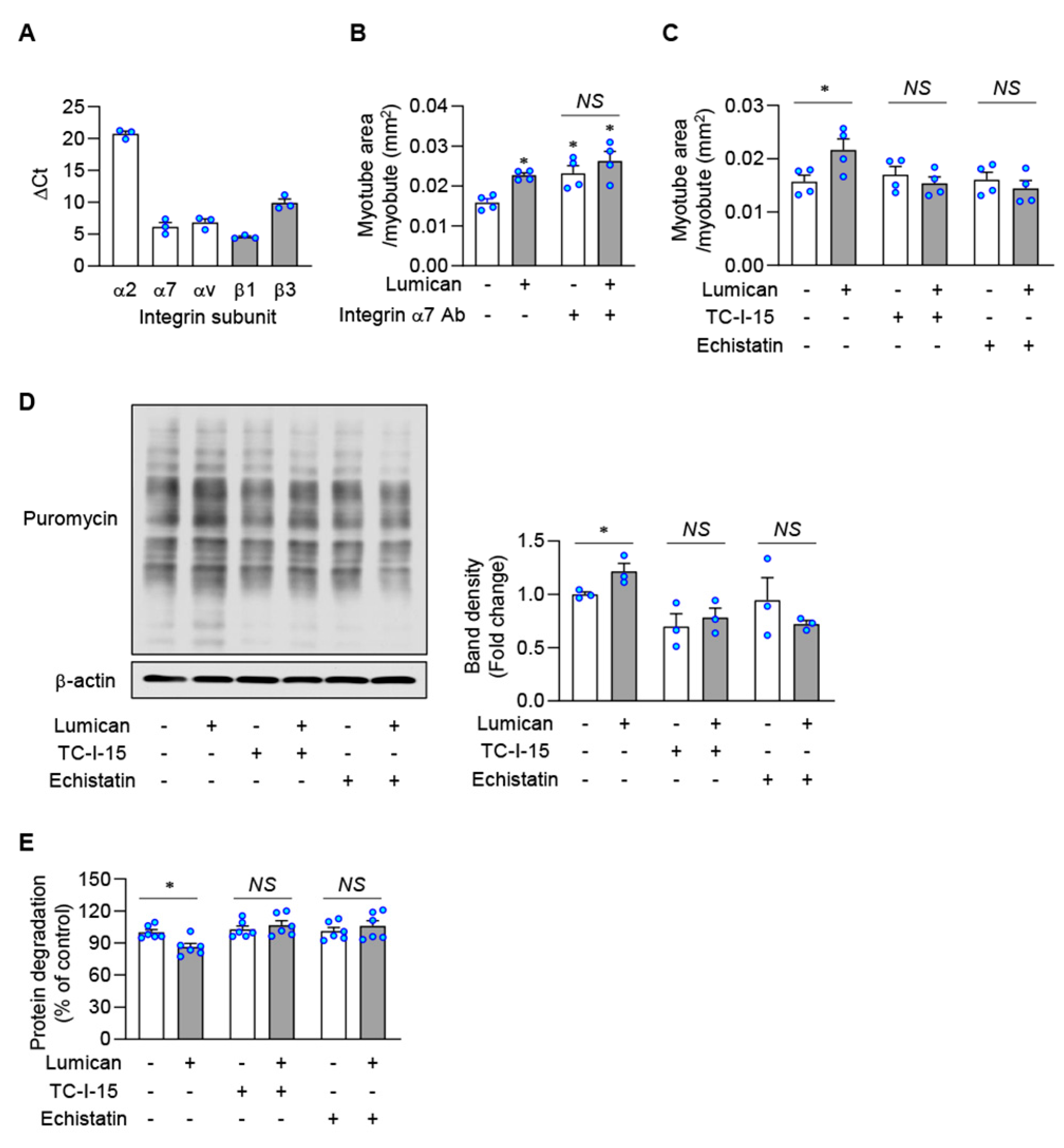
2.5. Integrins Are Putative Lumican Receptors in Muscle Cells
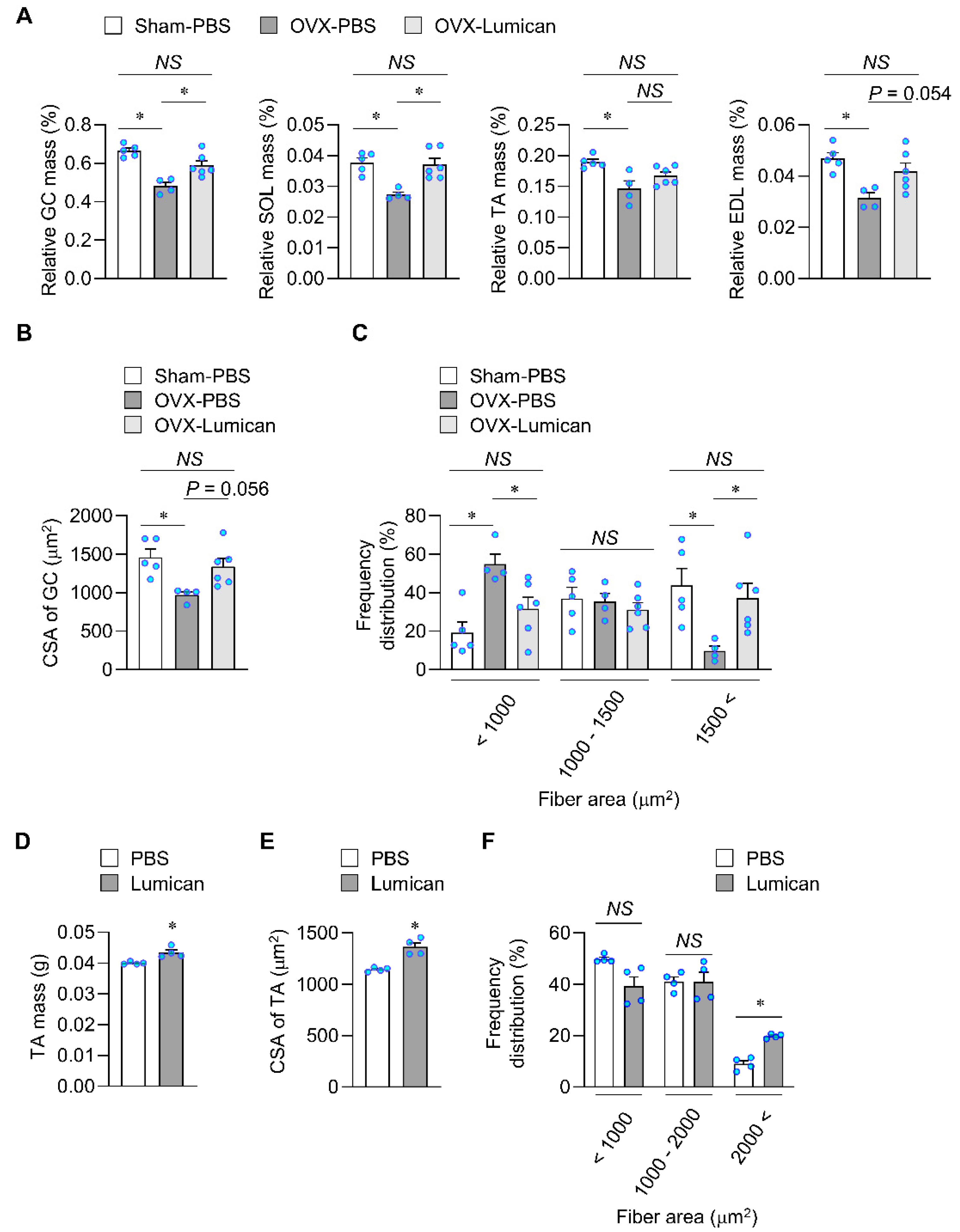
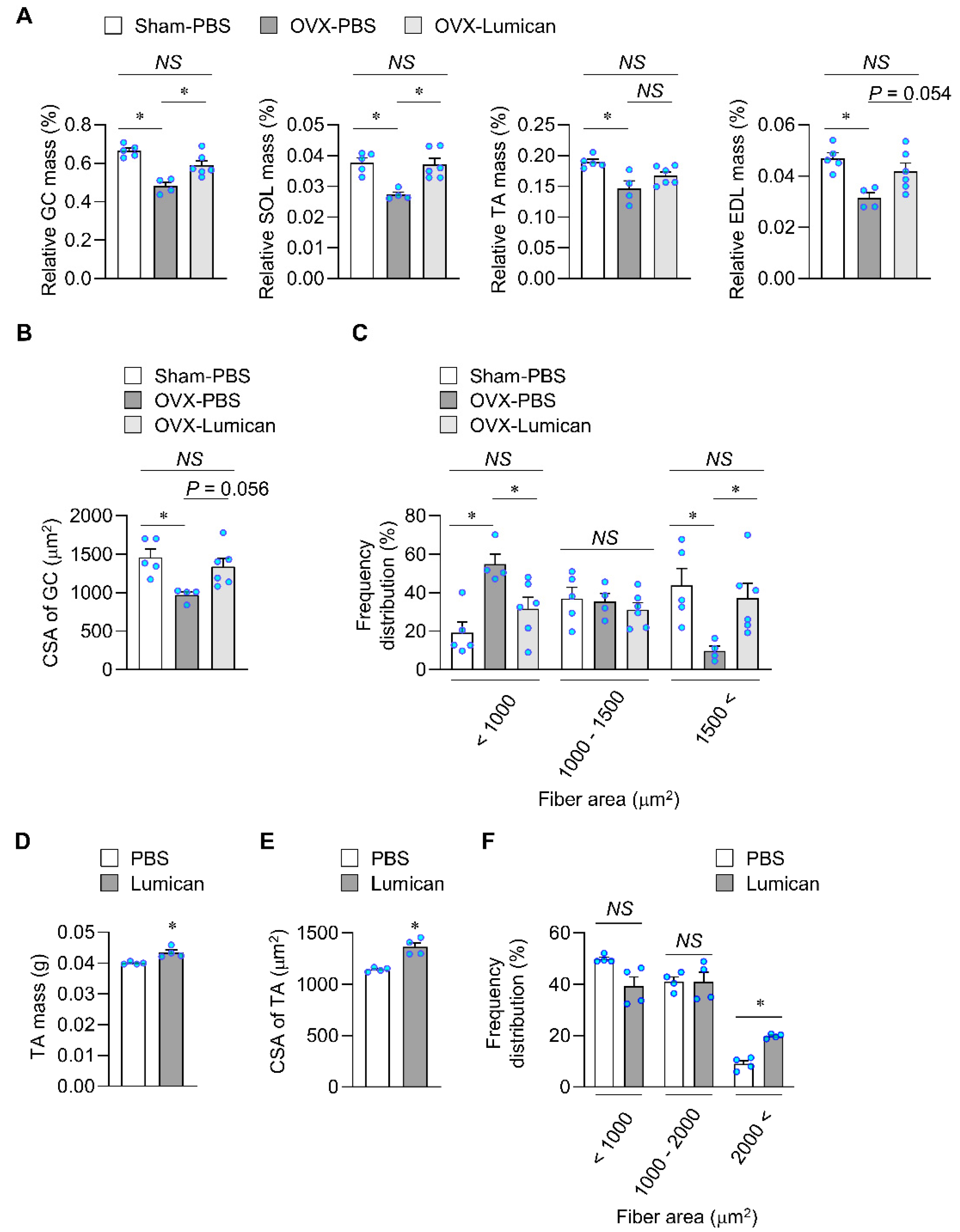
2.6. Lumican Treatment Increases Muscle Mass in Mouse Models Exhibiting Muscle Loss
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. In Vitro Myotube Formation
4.3. Real-Time PCR and Western Blot Analysis
4.4. Protein Synthesis and Degradation Assays
4.5. AAV Cloning and Production
4.6. Animals and Treatments
4.7. Haematoxylin and Eosin (H&E) Staining and Immunofluorescence
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Dennison, E.M.; Sayer, A.A.; Cooper, C. Epidemiology of sarcopenia and insight into possible therapeutic targets. Nat. Rev. Rheumatol. 2017, 13, 340–347. [Google Scholar] [CrossRef]
- Hawley, J.A.; Joyner, M.J.; Green, D.J. Mimicking exercise: What matters most and where to next? J. Physiol. 2021, 599, 791–802. [Google Scholar] [CrossRef]
- Safdar, A.; Saleem, A.; Tarnopolsky, M.A. The potential of endurance exercise-derived exosomes to treat metabolic diseases. Nat. Rev. Endocrinol. 2016, 12, 504–517. [Google Scholar] [CrossRef]
- Momenzadeh, S.; Zamani, S.; Pourteymourfard-Tabrizi, Z.; Barreiro, C.; Jami, M.S. Muscles proteome analysis; irisin administration mimics some molecular effects of exercise in quadriceps muscle. Biochimie 2021, 189, 144–157. [Google Scholar] [CrossRef]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostrom, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Vinel, C.; Lukjanenko, L.; Batut, A.; Deleruyelle, S.; Pradere, J.P.; Le Gonidec, S.; Dortignac, A.; Geoffre, N.; Pereira, O.; Karaz, S.; et al. The exerkine apelin reverses age-associated sarcopenia. Nat. Med. 2018, 24, 1360–1371. [Google Scholar] [CrossRef]
- Otaka, N.; Shibata, R.; Ohashi, K.; Uemura, Y.; Kambara, T.; Enomoto, T.; Ogawa, H.; Ito, M.; Kawanishi, H.; Maruyama, S.; et al. Myonectin Is an Exercise-Induced Myokine That Protects the Heart From Ischemia-Reperfusion Injury. Circ. Res. 2018, 123, 1326–1338. [Google Scholar] [CrossRef]
- Pourteymour, S.; Eckardt, K.; Holen, T.; Langleite, T.; Lee, S.; Jensen, J.; Birkeland, K.I.; Drevon, C.A.; Hjorth, M. Global mRNA sequencing of human skeletal muscle: Search for novel exercise-regulated myokines. Mol. Metab. 2017, 6, 352–365. [Google Scholar] [CrossRef]
- Reza, M.M.; Subramaniyam, N.; Sim, C.M.; Ge, X.; Sathiakumar, D.; McFarlane, C.; Sharma, M.; Kambadur, R. Irisin is a pro-myogenic factor that induces skeletal muscle hypertrophy and rescues denervation-induced atrophy. Nat. Commun. 2017, 8, 1104. [Google Scholar] [CrossRef] [Green Version]
- Blochberger, T.C.; Cornuet, P.K.; Hassell, J.R. Isolation and partial characterization of lumican and decorin from adult chicken corneas. A keratan sulfate-containing isoform of decorin is developmentally regulated. J. Biol. Chem. 1992, 267, 20613–20619. [Google Scholar] [CrossRef]
- Chakravarti, S.; Magnuson, T.; Lass, J.H.; Jepsen, K.J.; LaMantia, C.; Carroll, H. Lumican regulates collagen fibril assembly: Skin fragility and corneal opacity in the absence of lumican. J. Cell Biol. 1998, 141, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, S.; Petroll, W.M.; Hassell, J.R.; Jester, J.V.; Lass, J.H.; Paul, J.; Birk, D.E. Corneal opacity in lumican-null mice: Defects in collagen fibril structure and packing in the posterior stroma. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3365–3373. [Google Scholar]
- Liu, X.J.; Kong, F.Z.; Wang, Y.H.; Zheng, J.H.; Wan, W.D.; Deng, C.L.; Mao, G.Y.; Li, J.; Yang, X.M.; Zhang, Y.L.; et al. Lumican Accelerates Wound Healing by Enhancing alpha2beta1 Integrin-Mediated Fibroblast Contractility. PLoS ONE 2013, 8, e67124. [Google Scholar] [CrossRef]
- Yamanaka, O.; Yuan, Y.; Coulson-Thomas, V.J.; Gesteira, T.F.; Call, M.K.; Zhang, Y.; Zhang, J.; Chang, S.H.; Xie, C.; Liu, C.Y.; et al. Lumican binds ALK5 to promote epithelium wound healing. PLoS ONE 2013, 8, e82730. [Google Scholar] [CrossRef] [PubMed]
- Niewiarowska, J.; Brezillon, S.; Sacewicz-Hofman, I.; Bednarek, R.; Maquart, F.X.; Malinowski, M.; Wiktorska, M.; Wegrowski, Y.; Cierniewski, C.S. Lumican inhibits angiogenesis by interfering with alpha2beta1 receptor activity and downregulating MMP-14 expression. Thromb. Res. 2011, 128, 452–457. [Google Scholar] [CrossRef]
- Zeltz, C.; Brezillon, S.; Kapyla, J.; Eble, J.A.; Bobichon, H.; Terryn, C.; Perreau, C.; Franz, C.M.; Heino, J.; Maquart, F.X.; et al. Lumican inhibits cell migration through alpha2beta1 integrin. Exp. Cell Res. 2010, 316, 2922–2931. [Google Scholar] [CrossRef]
- D’Onofrio, M.F.; Brezillon, S.; Baranek, T.; Perreau, C.; Roughley, P.J.; Maquart, F.X.; Wegrowski, Y. Identification of beta1 integrin as mediator of melanoma cell adhesion to lumican. Biochem. Biophys. Res. Commun. 2008, 365, 266–272. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, S.J.; Kim, D.A.; Lee, S.H.; Koh, J.M.; Kim, B.J. Muscle-Derived Lumican Stimulates Bone Formation via Integrin alpha2beta1 and the Downstream ERK Signal. Front. Cell Dev. Biol. 2020, 8, 565826. [Google Scholar] [CrossRef]
- Norheim, F.; Raastad, T.; Thiede, B.; Rustan, A.C.; Drevon, C.A.; Haugen, F. Proteomic identification of secreted proteins from human skeletal muscle cells and expression in response to strength training. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1013–E1021. [Google Scholar] [CrossRef]
- Wang, Y.; Song, J.; Liu, X.; Liu, J.; Zhang, Q.; Yan, X.; Yuan, X.; Ren, D. Multiple Effects of Mechanical Stretch on Myogenic Progenitor Cells. Stem Cells Dev. 2020, 29, 336–352. [Google Scholar] [CrossRef] [Green Version]
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Gumucio, J.P.; Mendias, C.L. Atrogin-1, MuRF-1, and sarcopenia. Endocrine 2013, 43, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Truty, M.A.; Kang, Y.; Chopin-Laly, X.; Zhang, R.; Roife, D.; Chatterjee, D.; Lin, E.; Thomas, R.M.; Wang, H.; et al. Extracellular lumican inhibits pancreatic cancer cell growth and is associated with prolonged survival after surgery. Clin. Cancer Res. 2014, 20, 6529–6540. [Google Scholar] [CrossRef] [PubMed]
- Maiti, G.; Frikeche, J.; Lam, C.Y.; Biswas, A.; Shinde, V.; Samanovic, M.; Kagan, J.C.; Mulligan, M.J.; Chakravarti, S. Matrix lumican endocytosed by immune cells controls receptor ligand trafficking to promote TLR4 and restrict TLR9 in sepsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2100999118. [Google Scholar] [CrossRef]
- Tarabees, R.; Hill, D.; Rauch, C.; Barrow, P.A.; Loughna, P.T. Endotoxin transiently inhibits protein synthesis through Akt and MAPK mediating pathways in C2C12 myotubes. Am. J. Physiol. Cell Physiol. 2011, 301, C895–C902. [Google Scholar] [CrossRef]
- Cabane, C.; Coldefy, A.S.; Yeow, K.; Derijard, B. The p38 pathway regulates Akt both at the protein and transcriptional activation levels during myogenesis. Cell Signal. 2004, 16, 1405–1415. [Google Scholar] [CrossRef]
- Wu, Z.; Woodring, P.J.; Bhakta, K.S.; Tamura, K.; Wen, F.; Feramisco, J.R.; Karin, M.; Wang, J.Y.; Puri, P.L. p38 and extracellular signal-regulated kinases regulate the myogenic program at multiple steps. Mol. Cell. Biol. 2000, 20, 3951–3964. [Google Scholar] [CrossRef]
- Bahrami, B.F.; Ataie-Kachoie, P.; Pourgholami, M.H.; Morris, D.L. p70 Ribosomal protein S6 kinase (Rps6kb1): An update. J. Clin. Pathol. 2014, 67, 1019–1025. [Google Scholar] [CrossRef]
- Mayer, U. Integrins: Redundant or important players in skeletal muscle? J. Biol. Chem. 2003, 278, 14587–14590. [Google Scholar] [CrossRef]
- Liu, H.; Niu, A.; Chen, S.E.; Li, Y.P. Beta3-integrin mediates satellite cell differentiation in regenerating mouse muscle. FASEB J. 2011, 25, 1914–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Kim, H.; Lee, Y.S.; Moon, S.A.; Kim, J.M.; Kim, H.; Kim, M.J.; Yu, J.; Kim, K.; Baek, I.J.; et al. SLIT3 promotes myogenic differentiation as a novel therapeutic factor against muscle loss. J. Cachexia Sarcopenia Muscle 2021, 12, 1724–1740. [Google Scholar] [CrossRef] [PubMed]
- Sipila, S.; Narici, M.; Kjaer, M.; Pollanen, E.; Atkinson, R.A.; Hansen, M.; Kovanen, V. Sex hormones and skeletal muscle weakness. Biogerontology 2013, 14, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Steensberg, A.; Fischer, C.; Keller, C.; Keller, P.; Plomgaard, P.; Febbraio, M.; Saltin, B. Searching for the exercise factor: Is IL-6 a candidate? J. Muscle. Res. Cell Motil. 2003, 24, 113–119. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle-Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef]
- Hettige, P.; Tahir, U.; Nishikawa, K.C.; Gage, M.J. Comparative analysis of the transcriptomes of EDL, psoas, and soleus muscles from mice. BMC Genomics 2020, 21, 808. [Google Scholar] [CrossRef]
- Perdiguero, E.; Ruiz-Bonilla, V.; Gresh, L.; Hui, L.; Ballestar, E.; Sousa-Victor, P.; Baeza-Raja, B.; Jardi, M.; Bosch-Comas, A.; Esteller, M.; et al. Genetic analysis of p38 MAP kinases in myogenesis: Fundamental role of p38alpha in abrogating myoblast proliferation. EMBO J. 2007, 26, 1245–1256. [Google Scholar] [CrossRef]
- Zetser, A.; Gredinger, E.; Bengal, E. p38 mitogen-activated protein kinase pathway promotes skeletal muscle differentiation. Participation of the Mef2c transcription factor. J. Biol. Chem. 1999, 274, 5193–5200. [Google Scholar] [CrossRef]
- Zbinden-Foncea, H.; Deldicque, L.; Pierre, N.; Francaux, M.; Raymackers, J.M. TLR2 and TLR4 activation induces p38 MAPK-dependent phosphorylation of S6 kinase 1 in C2C12 myotubes. Cell Biol. Int. 2012, 36, 1107–1113. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, Y.; Bode, A.M.; Ma, W.Y.; Dong, Z. Phosphorylation of 4E-BP1 is mediated by the p38/MSK1 pathway in response to UVB irradiation. J. Biol. Chem. 2002, 277, 8810–8816. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, W.W.; Liu, C.Y. Roles of lumican and keratocan on corneal transparency. Glycoconj. J. 2002, 19, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Zutter, M.M.; Santoro, S.A. Widespread histologic distribution of the alpha 2 beta 1 integrin cell-surface collagen receptor. Am. J. Pathol. 1990, 137, 113–120. [Google Scholar] [PubMed]
- Bouvard, D.; Brakebusch, C.; Gustafsson, E.; Aszodi, A.; Bengtsson, T.; Berna, A.; Fassler, R. Functional consequences of integrin gene mutations in mice. Circ. Res. 2001, 89, 211–223. [Google Scholar] [CrossRef]
- Chakravarti, S. Functions of lumican and fibromodulin: Lessons from knockout mice. Glycoconj. J. 2002, 19, 287–293. [Google Scholar] [CrossRef]
- Zilio, F.; Di Fusco, S.A.; Flori, M.; Malvezzi Caracciolo D’Aquino, M.; Pollarolo, L.; Ingianni, N.; Luca, F.; Riccio, C.; Gulizia, M.M.; Gabrielli, D.; et al. Physical activity and the heart: From well-established cardiovascular benefits to possible adverse effects. Trends Cardiovasc. Med. 2022, in press. [CrossRef]
- Peters, H.P.; De Vries, W.R.; Vanberge-Henegouwen, G.P.; Akkermans, L.M. Potential benefits and hazards of physical activity and exercise on the gastrointestinal tract. Gut 2001, 48, 435–439. [Google Scholar] [CrossRef]
- Bloemberg, D.; Quadrilatero, J. Rapid determination of myosin heavy chain expression in rat, mouse, and human skeletal muscle using multicolor immunofluorescence analysis. PLoS ONE 2012, 7, e35273. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.J.; Lee, Y.-S.; Kim, D.A.; Moon, S.A.; Lee, S.E.; Lee, S.H.; Koh, J.-M. Lumican, an Exerkine, Protects against Skeletal Muscle Loss. Int. J. Mol. Sci. 2022, 23, 10031. https://doi.org/10.3390/ijms231710031
Cho HJ, Lee Y-S, Kim DA, Moon SA, Lee SE, Lee SH, Koh J-M. Lumican, an Exerkine, Protects against Skeletal Muscle Loss. International Journal of Molecular Sciences. 2022; 23(17):10031. https://doi.org/10.3390/ijms231710031
Chicago/Turabian StyleCho, Han Jin, Young-Sun Lee, Da Ae Kim, Sung Ah Moon, Seung Eun Lee, Seung Hun Lee, and Jung-Min Koh. 2022. "Lumican, an Exerkine, Protects against Skeletal Muscle Loss" International Journal of Molecular Sciences 23, no. 17: 10031. https://doi.org/10.3390/ijms231710031
APA StyleCho, H. J., Lee, Y.-S., Kim, D. A., Moon, S. A., Lee, S. E., Lee, S. H., & Koh, J.-M. (2022). Lumican, an Exerkine, Protects against Skeletal Muscle Loss. International Journal of Molecular Sciences, 23(17), 10031. https://doi.org/10.3390/ijms231710031