
Curcumin Remedies Testicular Function and Spermatogenesis in Male Mice with Low-Carbohydrate-Diet-Induced Metabolic Dysfunction



Abstract
:1. Introduction
2. Results
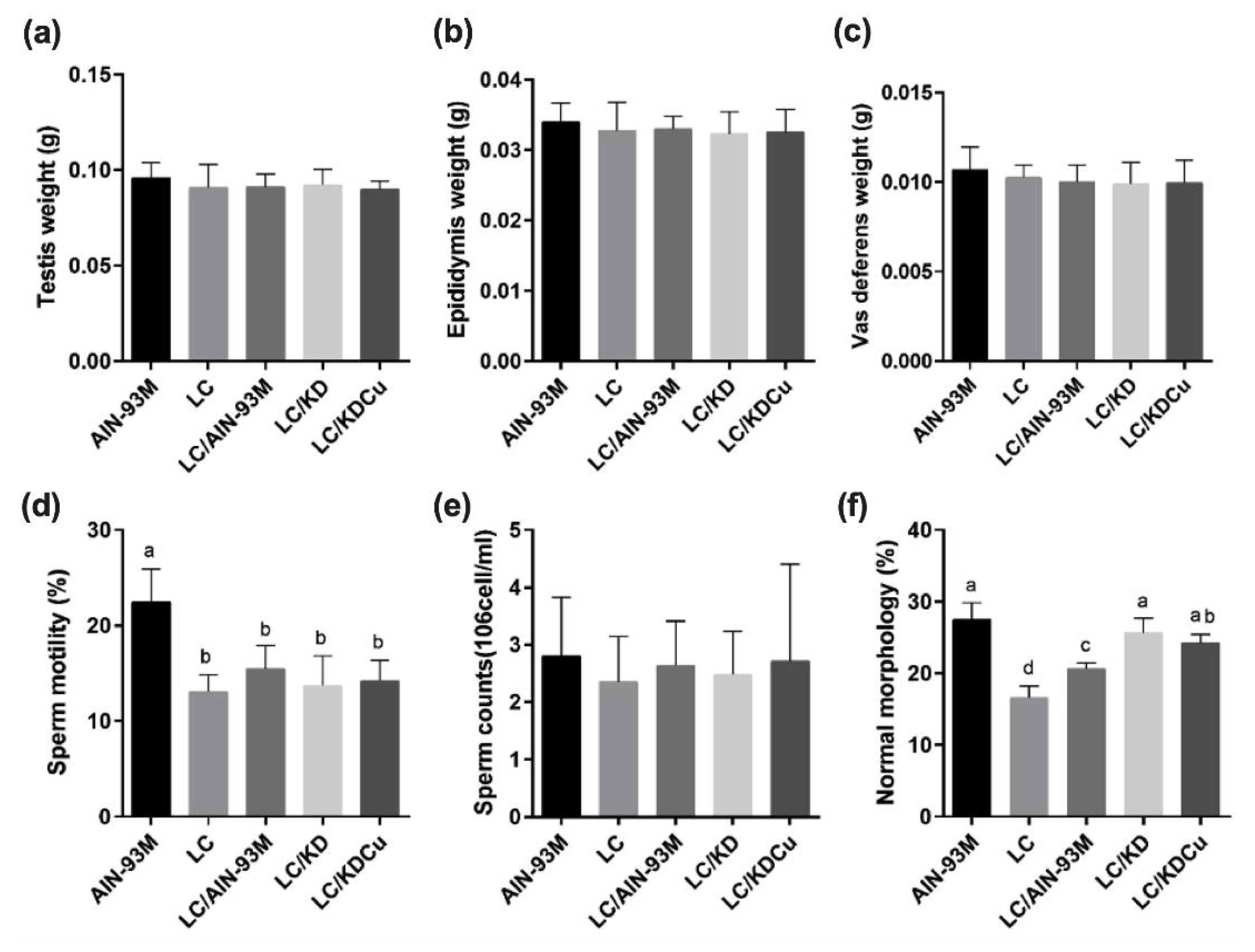
2.1. Curcumin Does Not Affect Reproductive Organ Weight and Lipid Profiles
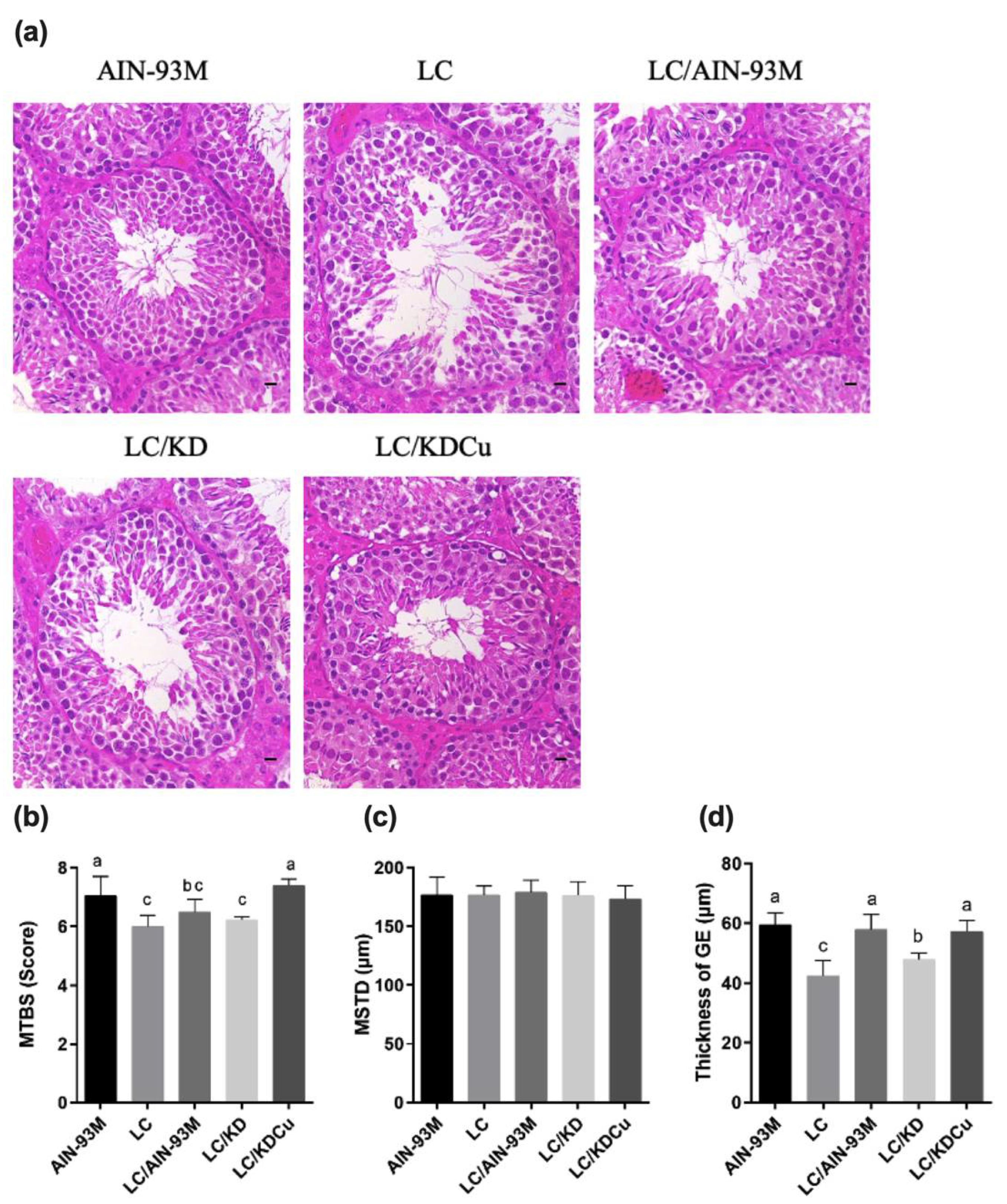
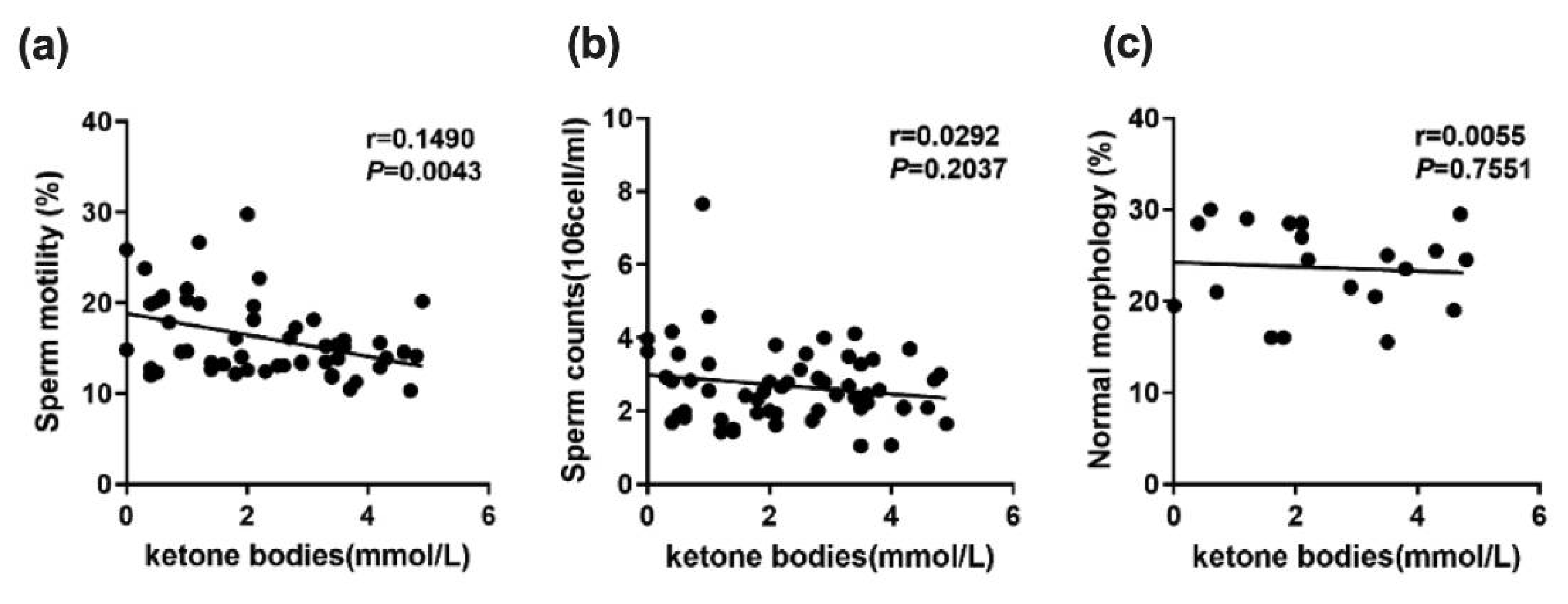
2.2. Curcumin Ameliorates the Decrease in Testicular Spermatogenesis and Sperm Quality Caused by a Low-Carbohydrate Diet
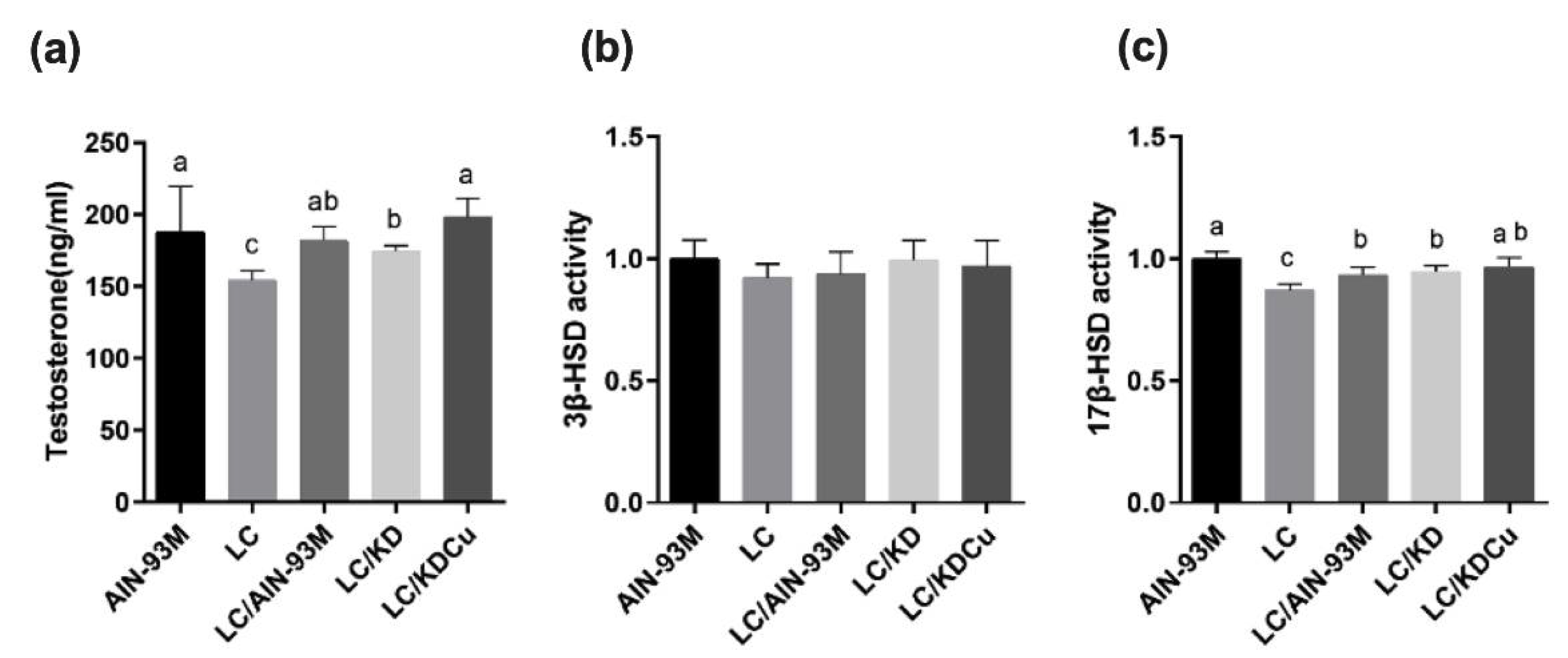
2.3. Curcumin Increases the Testosterone Level through Upregulating 17β-HSD Expression
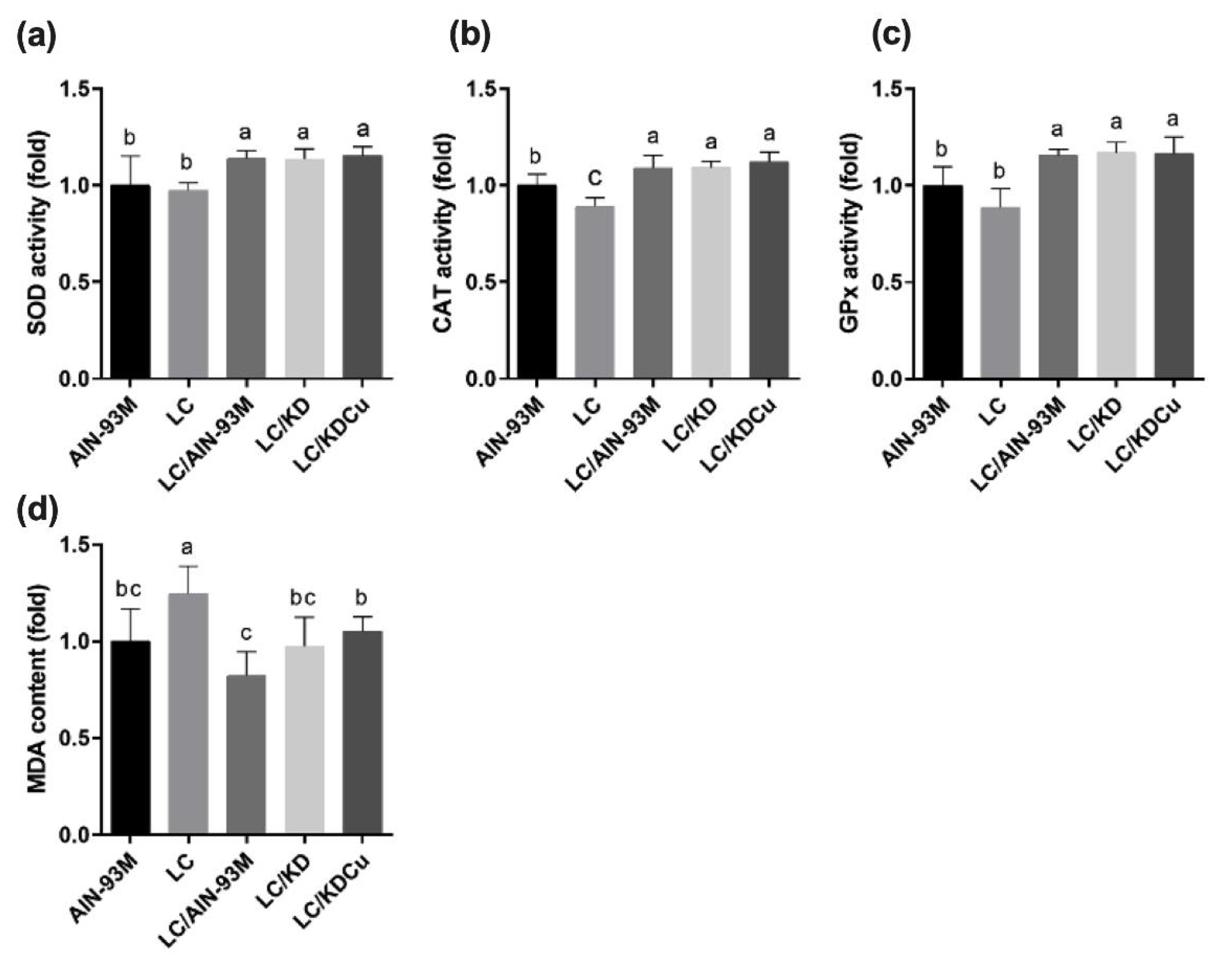
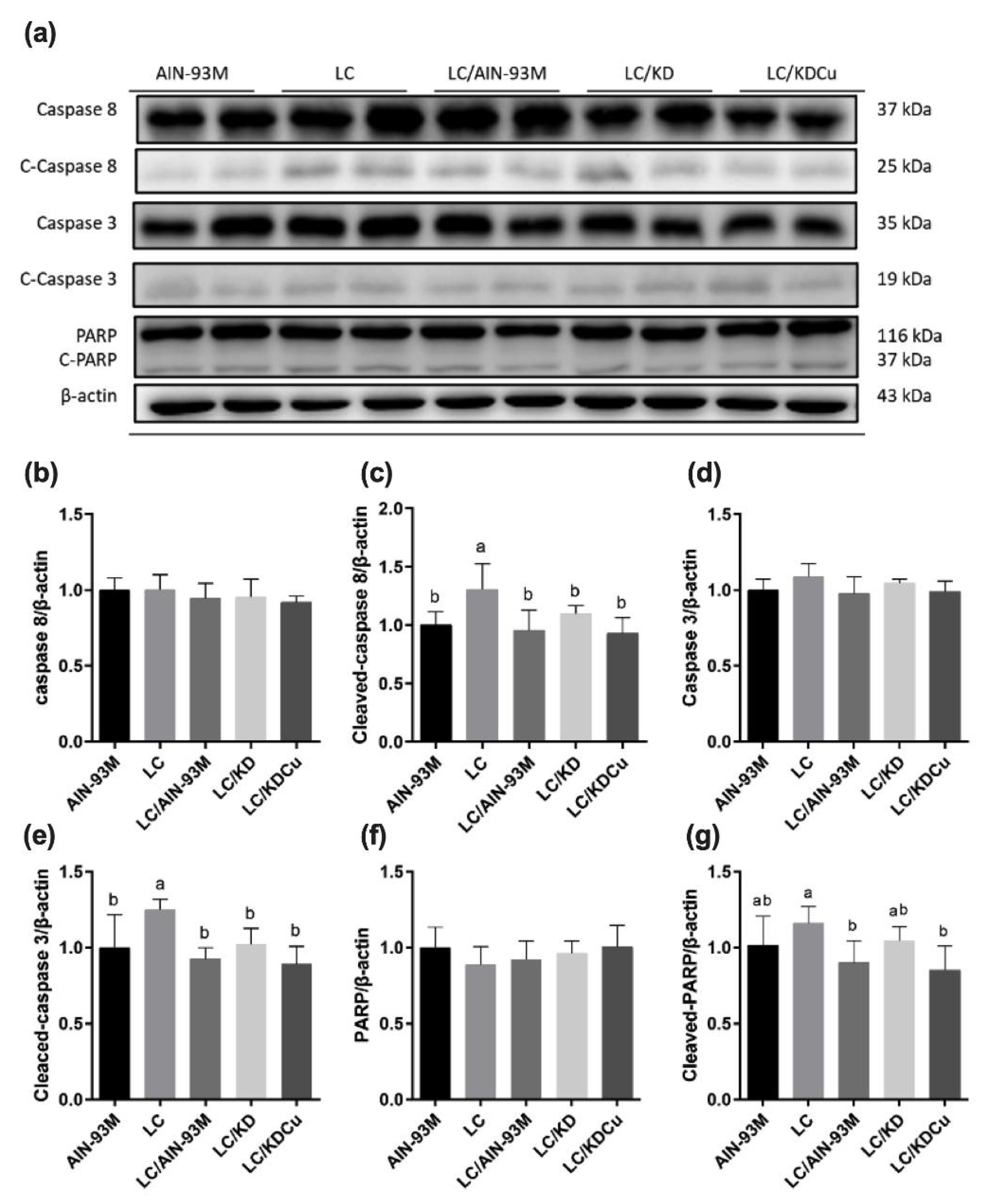
2.4. Curcumin Activates Antioxidant Capacity and Suppresses Lipid Peroxidation, Apoptosis and Inflammation in the Testis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Study Design
4.3. Histological Analysis
4.4. Serum Analysis
4.5. Sperm Quality Analysis
4.6. Testicular Hormone, Antioxidants and Marker of Lipid Peroxidation Analysis
4.7. Western Blotting Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katz, D.J.; Teloken, P.; Shoshany, O. Male infertility-The other side of the equation. Aust. Fam. Physician 2017, 46, 641–646. [Google Scholar] [PubMed]
- Skoracka, K.; Eder, P.; Łykowska-Szuber, L.; Dobrowolska, A.; Krela-Kaźmierczak, I. Diet and Nutritional Factors in Male (In)fertility-Underestimated Factors. J. Clin. Med. 2020, 9, 1400. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Bulló, M.; Salas-Salvadó, J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: A systematic review of observational studies. Hum. Reprod. Update 2017, 23, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Nassan, F.L.; Chavarro, J.E.; Tanrikut, C. Diet and men’s fertility: Does diet affect sperm quality? Fertil. Steril. 2018, 110, 570–577. [Google Scholar] [CrossRef]
- Varani, J. Healthful Eating, the Western Style Diet and Chronic Disease. Appro. Poult. Dairy Vet. Sci. 2017, 1. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: A Comprehensive Review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef] [PubMed]
- Del Razo Olvera, F.M.; Melgarejo Hernández, M.A.; Mehta, R.; Aguilar Salinas, C.A. Setting the Lipid Component of the Diet: A Work in Process. Adv. Nutr. 2017, 8, 165s–172s. [Google Scholar] [CrossRef]
- Bilsborough, S.A.; Crowe, T.C. Low-carbohydrate diets: What are the potential short- and long-term health implications? Asia Pac. J. Clin. Nutr. 2003, 12, 396–404. [Google Scholar]
- Trumbo, P.; Schlicker, S.; Yates, A.A.; Poos, M. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J. Am. Diet. Assoc. 2002, 102, 1621–1630. [Google Scholar] [CrossRef]
- Liu, C.Y.; Chou, Y.C.; Chao, J.C.; Hsu, C.Y.; Cha, T.L.; Tsao, C.W. The Association between Dietary Patterns and Semen Quality in a General Asian Population of 7282 Males. PLoS ONE 2015, 10, e0134224. [Google Scholar] [CrossRef] [PubMed]
- Eslamian, G.; Amirjannati, N.; Rashidkhani, B.; Sadeghi, M.R.; Hekmatdoost, A. Intake of food groups and idiopathic asthenozoospermia: A case-control study. Hum. Reprod. 2012, 27, 3328–3336. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.L.; Liu, Y.; Liu, M.E.; Pan, J.X.; Guo, M.X.; Sheng, J.Z.; Huang, H.F. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 2015, 17, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Takahashi, T.; Iguchi, N.; Kitamura, K.; Miyagawa, Y.; Tsujimura, A.; Matsumiya, K.; Okuyama, A.; Nishimune, Y. Ketone bodies could support the motility but not the acrosome reaction of mouse sperm. Int. J. Androl. 2004, 27, 172–177. [Google Scholar] [CrossRef]
- Bone, W.; Jones, A.R.; Morin, C.; Nieschlag, E.; Cooper, T.G. Susceptibility of glycolytic enzyme activity and motility of spermatozoa from rat, mouse, and human to inhibition by proven and putative chlorinated antifertility compounds in vitro. J. Androl. 2001, 22, 464–470. [Google Scholar]
- Ghosh, S.; Banerjee, S.; Sil, P.C. The beneficial role of curcumin on inflammation, diabetes and neurodegenerative disease: A recent update. Food Chem. Toxicol. 2015, 83, 111–124. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden nutraceutical: Multitargeting for multiple chronic diseases. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef]
- Ak, T.; Gülçin, I. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef]
- Yarru, L.P.; Settivari, R.S.; Gowda, N.K.; Antoniou, E.; Ledoux, D.R.; Rottinghaus, G.E. Effects of turmeric (Curcuma longa) on the expression of hepatic genes associated with biotransformation, antioxidant, and immune systems in broiler chicks fed aflatoxin. Poult. Sci. 2009, 88, 2620–2627. [Google Scholar] [CrossRef]
- Barzegar, A.; Moosavi-Movahedi, A.A. Intracellular ROS protection efficiency and free radical-scavenging activity of curcumin. PLoS ONE 2011, 6, e26012. [Google Scholar] [CrossRef] [PubMed]
- Panchal, H.D.; Vranizan, K.; Lee, C.Y.; Ho, J.; Ngai, J.; Timiras, P.S. Early anti-oxidative and anti-proliferative curcumin effects on neuroglioma cells suggest therapeutic targets. Neurochem. Res. 2008, 33, 1701–1710. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.F.; Hou, Z.Q.; Zhong, L.M.; Zhang, Q.Q. Effect of curcumin on the induction of glutathione S-transferases and NADP(H):quinone oxidoreductase and its possible mechanism of action. Yao Xue Xue Bao 2007, 42, 376–380. [Google Scholar] [PubMed]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.A.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin—From Molecule to Biological Function. Angew. Chem. Int. Ed. 2012, 51, 5308–5332. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Horbańczuk, M.; Tzvetkov, N.T.; Mocan, A.; Carradori, S.; Maggi, F.; Marchewka, J.; Sut, S.; Dall’Acqua, S.; Gan, R.Y.; et al. Curcumin: Total-Scale Analysis of the Scientific Literature. Molecules 2019, 24, 1393. [Google Scholar] [CrossRef]
- Alizadeh, F.; Javadi, M.; Karami, A.A.; Gholaminejad, F.; Kavianpour, M.; Haghighian, H.K. Curcumin nanomicelle improves semen parameters, oxidative stress, inflammatory biomarkers, and reproductive hormones in infertile men: A randomized clinical trial. Phytother. Res. 2018, 32, 514–521. [Google Scholar] [CrossRef]
- Ellacott, K.L.; Morton, G.J.; Woods, S.C.; Tso, P.; Schwartz, M.W. Assessment of feeding behavior in laboratory mice. Cell Metab. 2010, 12, 10–17. [Google Scholar] [CrossRef]
- Li, X.; Lian, F.; Liu, C.; Hu, K.-Q.; Wang, X.-D. Isocaloric Pair-Fed High-Carbohydrate Diet Induced More Hepatic Steatosis and Inflammation than High-Fat Diet Mediated by miR-34a/SIRT1 Axis in Mice. Sci. Rep. 2015, 5, 16774. [Google Scholar] [CrossRef]
- Volek, J.S.; Sharman, M.J.; Forsythe, C.E. Modification of lipoproteins by very low-carbohydrate diets. J. Nutr. 2005, 135, 1339–1342. [Google Scholar] [CrossRef]
- Attaman, J.A.; Toth, T.L.; Furtado, J.; Campos, H.; Hauser, R.; Chavarro, J.E. Dietary fat and semen quality among men attending a fertility clinic. Hum. Reprod. 2012, 27, 1466–1474. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Hill, D.L.; Wang, H.; Zhang, R. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway. Cancer Res. 2007, 67, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.; Chirino, Y.I.; Molina-Jijón, E.; Andérica-Romero, A.C.; Tapia, E.; Pedraza-Chaverrí, J. Renoprotective effect of the antioxidant curcumin: Recent findings. Redox Biol. 2013, 1, 448–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Li, J.; Song, B.; Xiao, X.; Zhang, B.; Qi, M.; Huang, W.; Yang, L.; Wang, Z. Curcumin rescues high fat diet-induced obesity and insulin sensitivity in mice through regulating SREBP pathway. Toxicol. Appl. Pharmacol. 2016, 304, 99–109. [Google Scholar] [CrossRef]
- El-Moselhy, M.A.; Taye, A.; Sharkawi, S.S.; El-Sisi, S.F.; Ahmed, A.F. The antihyperglycemic effect of curcumin in high fat diet fed rats. Role of TNF-alpha and free fatty acids. Food Chem. Toxicol. 2011, 49, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Shin, D.G.; Park, S.G.; Chu, S.B.; Gwon, L.W.; Lee, J.G.; Yon, J.M.; Baek, I.J.; Nam, S.Y. Curcumin dose-dependently improves spermatogenic disorders induced by scrotal heat stress in mice. Food Funct. 2015, 6, 3770–3777. [Google Scholar] [CrossRef] [PubMed]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Liu, W.; Zhai, Y.; Heng, X.; Che, F.Y.; Chen, W.; Sun, D.; Zhai, G. Oral bioavailability of curcumin: Problems and advancements. J. Drug Target. 2016, 24, 694–702. [Google Scholar] [CrossRef]
- Rahimi, H.R.; Nedaeinia, R.; Sepehri Shamloo, A.; Nikdoust, S.; Kazemi Oskuee, R. Novel delivery system for natural products: Nano-curcumin formulations. Avicenna J. Phytomed. 2016, 6, 383–398. [Google Scholar]
- McLachlan, R.I.; O’Donnell, L.; Meachem, S.J.; Stanton, P.G.; de Kretser, D.M.; Pratis, K.; Robertson, D.M. Hormonal regulation of spermatogenesis in primates and man: Insights for development of the male hormonal contraceptive. J. Androl. 2002, 23, 149–162. [Google Scholar]
- Sharma, P.; Aslam Khan, I.; Singh, R. Curcumin and Quercetin Ameliorated Cypermethrin and Deltamethrin-Induced Reproductive System Impairment in Male Wistar Rats by Upregulating The Activity of Pituitary-Gonadal Hormones and Steroidogenic Enzymes. Int. J. Fertil. Steril. 2018, 12, 72–80. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef] [PubMed]
- Tanrıkulu-Küçük, S.; Başaran-Küçükgergin, C.; Seyithanoğlu, M.; Doğru-Abbasoğlu, S.; Koçak, H.; Beyhan-Özdaş, Ş.; Öner-İyidoğan, Y. Effect of dietary curcumin and capsaicin on testicular and hepatic oxidant–antioxidant status in rats fed a high-fat diet. Appl. Physiol. Nutr. Metab. 2019, 44, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sharma, R.K.; Sikka, S.C.; Thomas, A.J., Jr.; Falcone, T.; Agarwal, A. Oxidative stress is associated with increased apoptosis leading to spermatozoa DNA damage in patients with male factor infertility. Fertil. Steril. 2003, 80, 531–535. [Google Scholar] [CrossRef]
- Mu, Y.; Yan, W.J.; Yin, T.L.; Yang, J. Curcumin ameliorates high-fat diet-induced spermatogenesis dysfunction. Mol. Med. Rep. 2016, 14, 3588–3594. [Google Scholar] [CrossRef]
- Park, J.Y.; Chung, T.W.; Jeong, Y.J.; Kwak, C.H.; Ha, S.H.; Kwon, K.M.; Abekura, F.; Cho, S.H.; Lee, Y.C.; Ha, K.T.; et al. Ascofuranone inhibits lipopolysaccharide-induced inflammatory response via NF-kappaB and AP-1, p-ERK, TNF-α, IL-6 and IL-1β in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0171322. [Google Scholar] [CrossRef]
- Rosenbaum, M.; Hall, K.D.; Guo, J.; Ravussin, E.; Mayer, L.S.; Reitman, M.L.; Smith, S.R.; Walsh, B.T.; Leibel, R.L. Glucose and Lipid Homeostasis and Inflammation in Humans Following an Isocaloric Ketogenic Diet. Obesity 2019, 27, 971–981. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/L) | AIN-93M | LC | KD | KDCu |
|---|---|---|---|---|
| Casein | 28.00 | 28.00 | 28.00 | 28.00 |
| Methionine | 0.30 | 0.30 | 0.30 | 0.30 |
| L-cystine | 0.50 | 0.50 | 0.50 | 0.50 |
| Dextrin | 192.00 | 22.00 | 12.00 | 12.00 |
| Sucrose | - | 23.00 | - | - |
| Soybean oil | 10.00 | 75.00 | 90.00 | 90.00 |
| Cellulose | 10.00 | 10.00 | 10.00 | 10.00 |
| Xanthan gum | 3.00 | 3.00 | 3.00 | 3.00 |
| Mineral | 15.00 | 15.00 | 15.00 | 15.00 |
| Vitamin | 5.00 | 5.00 | 5.00 | 5.00 |
| Choline | 0.53 | 0.53 | 0.53 | 0.53 |
| Curcumin | - | - | - | 80 mg/kg |
| Energy (% of kcal) | AIN-93M | LC | KD | KDCu |
| Carbohydrate | 79.2 | 18.6 | 4.9 | 4.9 |
| Fat | 9.3 | 69.6 | 83.2 | 83.2 |
| Protein | 11.9 | 11.9 | 11.8 | 11.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsao, C.-W.; Ke, P.-S.; Yang, H.-Y.; Chang, T.-C.; Liu, C.-Y. Curcumin Remedies Testicular Function and Spermatogenesis in Male Mice with Low-Carbohydrate-Diet-Induced Metabolic Dysfunction. Int. J. Mol. Sci. 2022, 23, 10009. https://doi.org/10.3390/ijms231710009
Tsao C-W, Ke P-S, Yang H-Y, Chang T-C, Liu C-Y. Curcumin Remedies Testicular Function and Spermatogenesis in Male Mice with Low-Carbohydrate-Diet-Induced Metabolic Dysfunction. International Journal of Molecular Sciences. 2022; 23(17):10009. https://doi.org/10.3390/ijms231710009
Chicago/Turabian StyleTsao, Chih-Wei, Pei-Shan Ke, Hsin-Yi Yang, Ting-Chia Chang, and Chin-Yu Liu. 2022. "Curcumin Remedies Testicular Function and Spermatogenesis in Male Mice with Low-Carbohydrate-Diet-Induced Metabolic Dysfunction" International Journal of Molecular Sciences 23, no. 17: 10009. https://doi.org/10.3390/ijms231710009
APA StyleTsao, C.-W., Ke, P.-S., Yang, H.-Y., Chang, T.-C., & Liu, C.-Y. (2022). Curcumin Remedies Testicular Function and Spermatogenesis in Male Mice with Low-Carbohydrate-Diet-Induced Metabolic Dysfunction. International Journal of Molecular Sciences, 23(17), 10009. https://doi.org/10.3390/ijms231710009