
Simple to Complex: The Role of Actin and Microtubules in Mitochondrial Dynamics in Amoeba, Yeast, and Mammalian Cells


Abstract
:1. Introduction
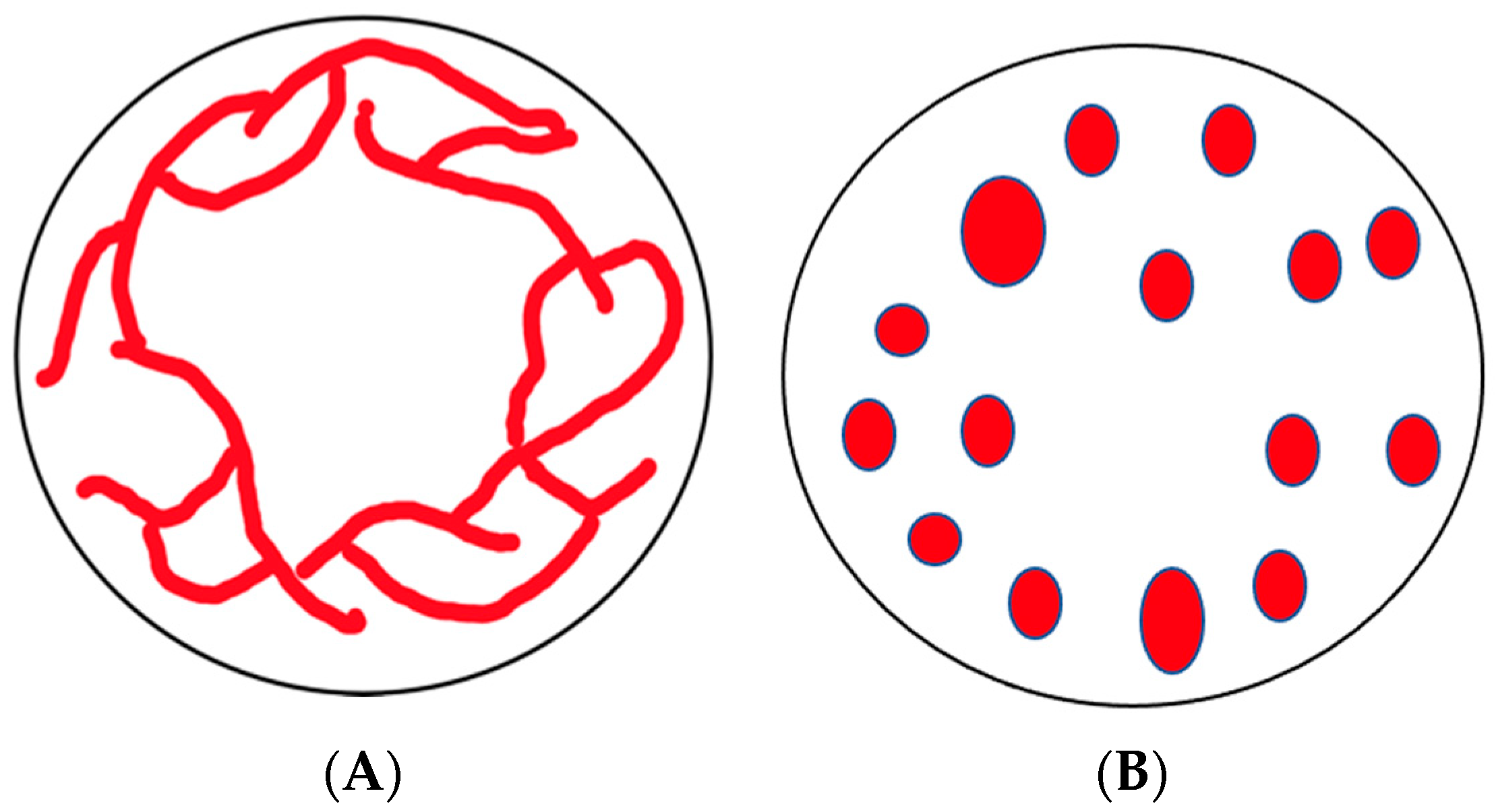
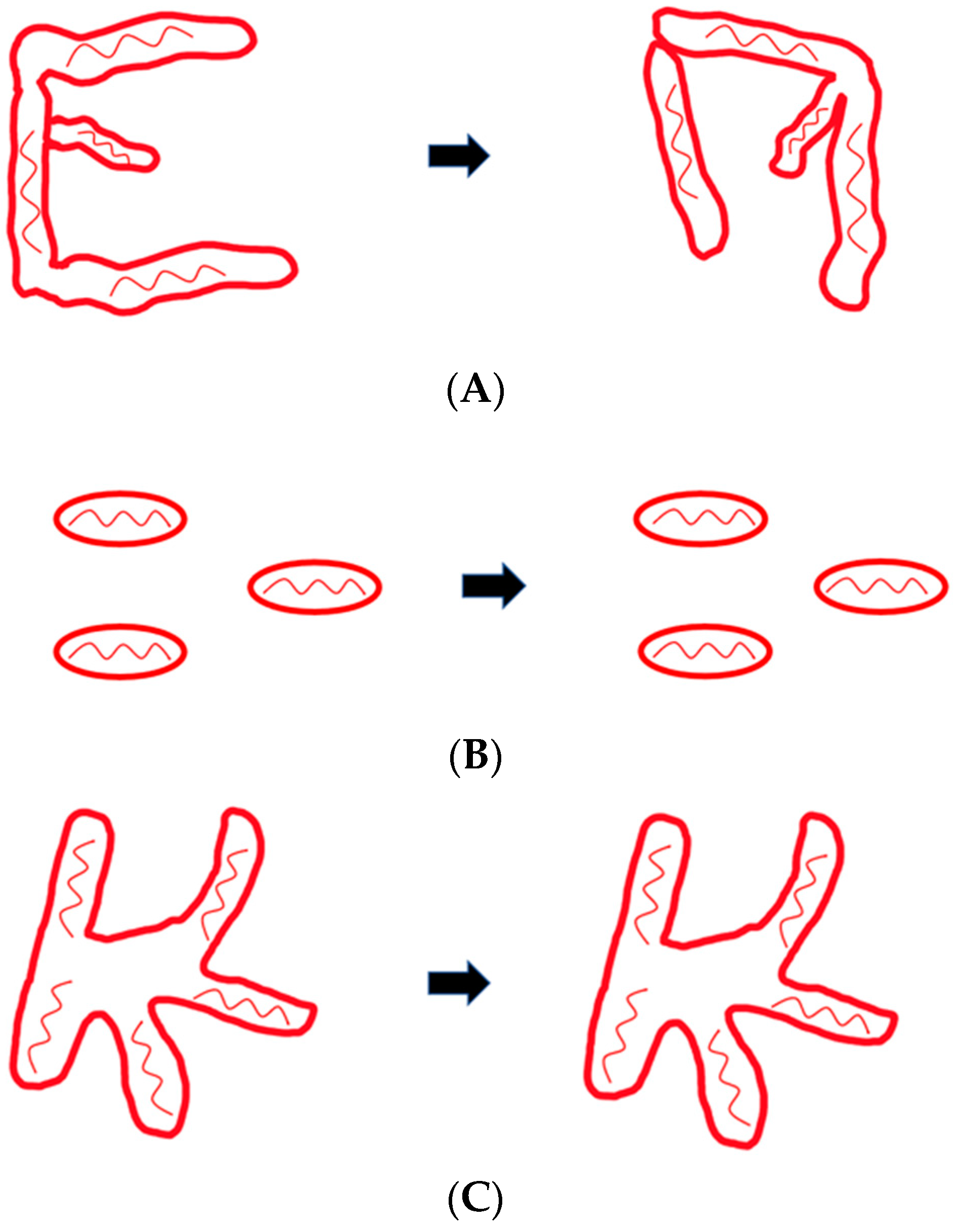
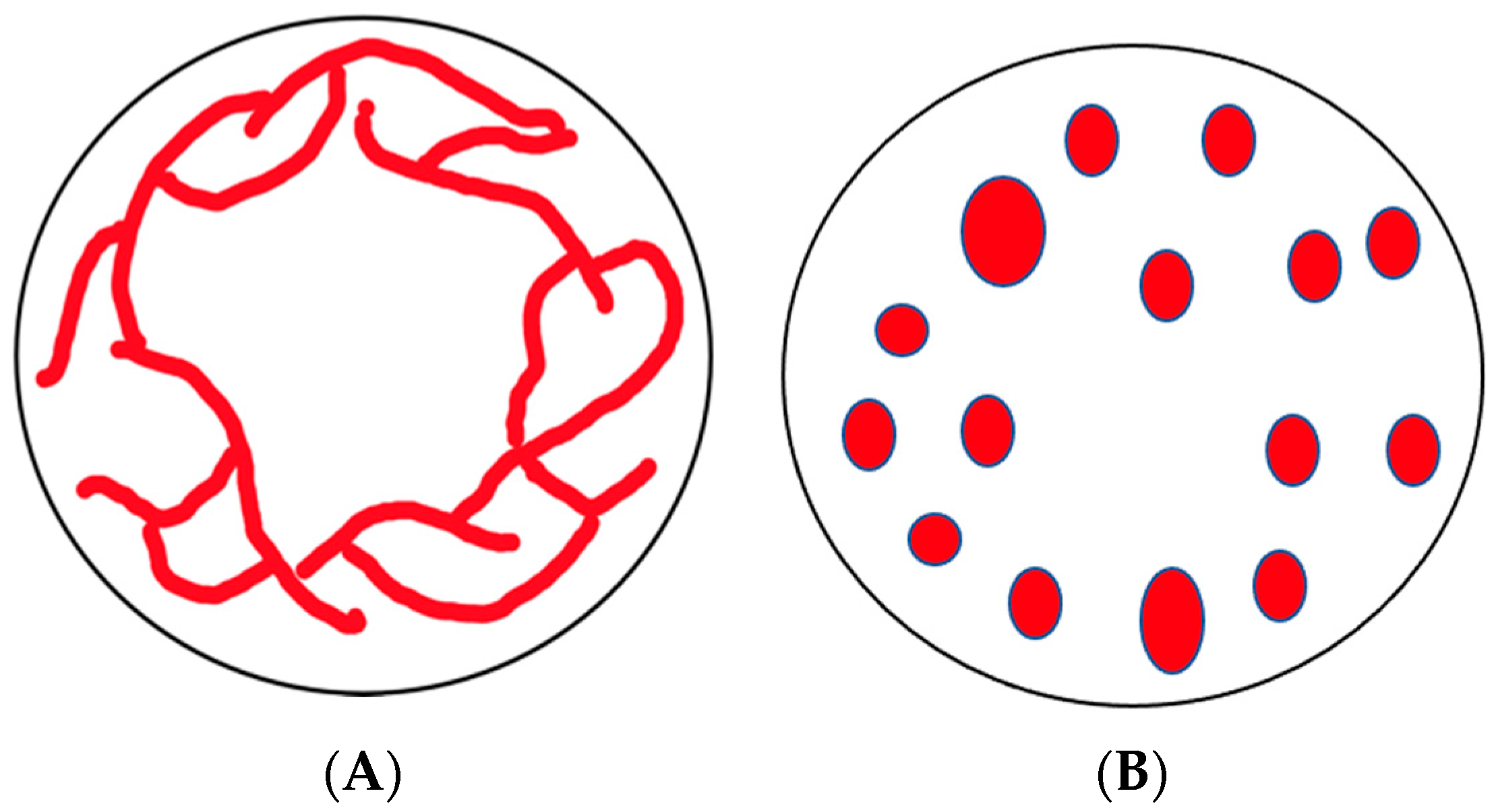
2. Mitochondrial Dynamics and the Cytoskeleton
3. Microtubules and Mitochondrial Dynamics in Neurons and Mammalian Cells
4. Microtubules and Mitochondrial Dynamics in Yeast
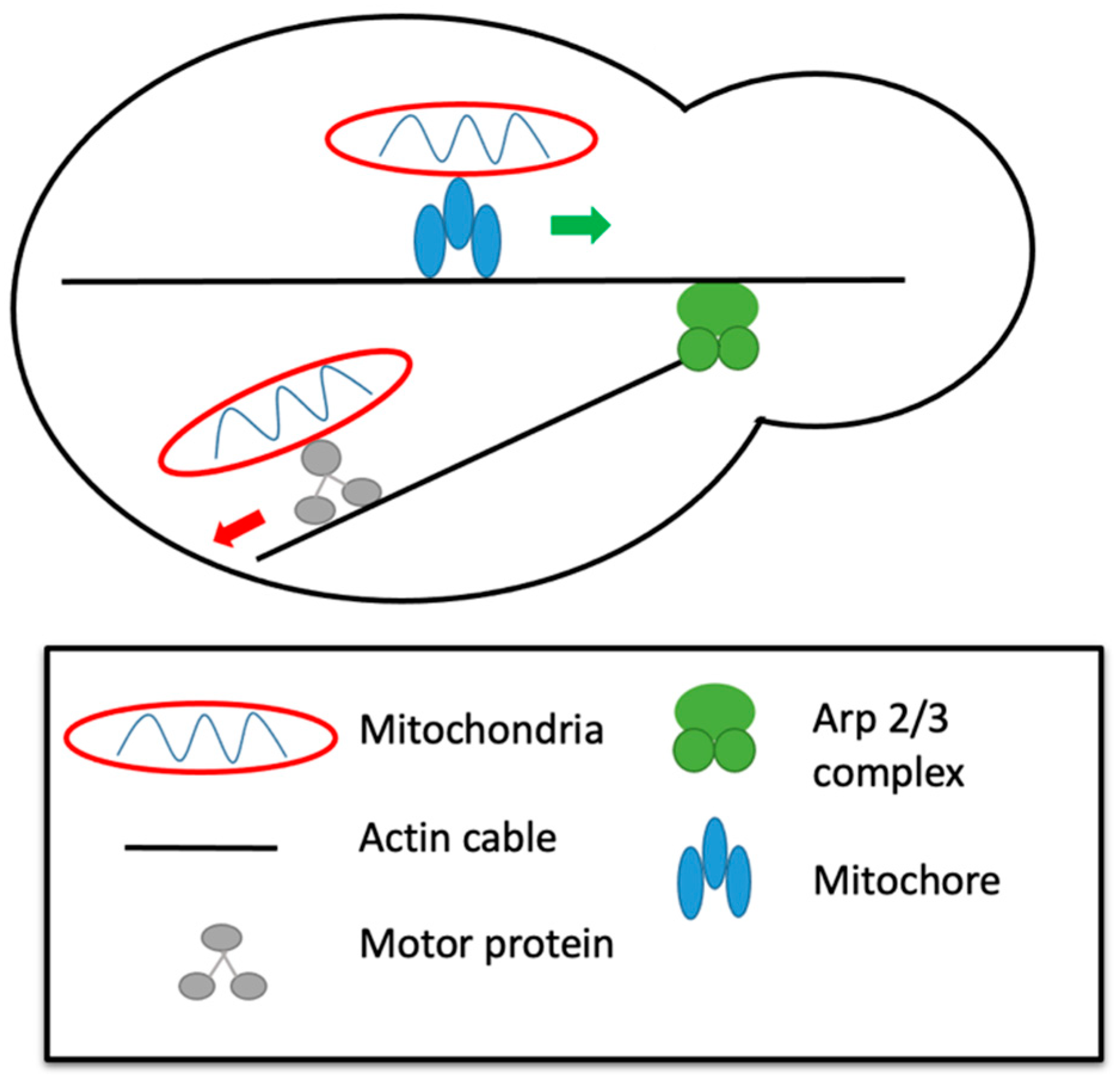
5. Actin and Mitochondrial Dynamics in Mammals
6. Actin and Mitochondrial Dynamics in Yeast
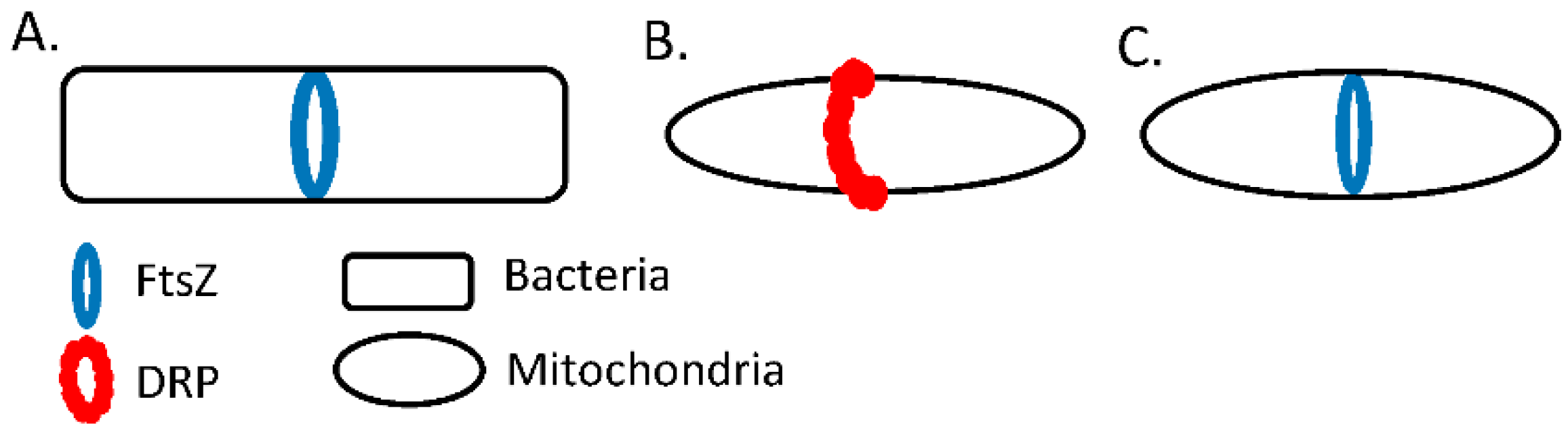
7. Dictyostelium discoideum as a Model
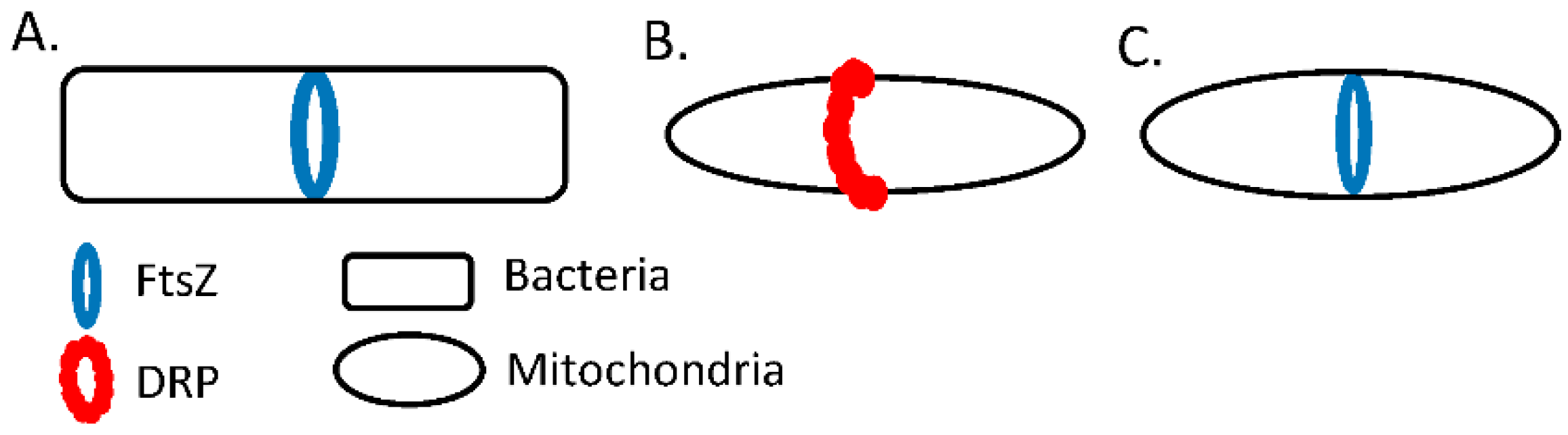
8. The Cytoskeleton and Mitochondrial Dynamics in Dictyostelium discoideum
9. The Cytoskeleton and Mitochondrial Dynamics in Neurodegenerative Diseases
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hatefi, Y. ATP Synthesis in Mitochondria. Eur. J. Biochem. 1993, 218, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, G.; Szabadkai, G.; Rizzuto, R. Mitochondrial Calcium Homeostasis: Mechanisms and Molecules. IUBMB Life 2001, 52, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Gulbins, E.; Dreschers, S.; Bock, J. Role of Mitochondria in Apoptosis. Exp. Physiol. 2003, 88, 85–90. [Google Scholar] [CrossRef]
- Maeda, Y.; Chida, J. Control of Cell Differentiation by Mitochondria, Typically Evidenced in Dictyostelium Development. Biomolecules 2013, 3, 943–966. [Google Scholar] [CrossRef]
- Nunnari, J.; Suomalainen, A. Mitochondria: In Sickness and in Health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [Green Version]
- Bartolák-Suki, E.; Imsirovic, J.; Nishibori, Y.; Krishnan, R.; Suki, B. Regulation of Mitochondrial Structure and Dynamics by the Cytoskeleton and Mechanical Factors. Int. J. Mol. Sci. 2017, 18, 1812. [Google Scholar] [CrossRef]
- Bereiter-Hahn, J. Behavior of Mitochondria in the Living Cell. Int. Rev. Cytol. 1990, 122, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.M.; Bak, I.J. The Fate of Mitochondria During Aging in Tetrahymena Pyriformis. J. Cell Biol. 1964, 20, 113–129. [Google Scholar] [CrossRef]
- Gilson, P.R.; Yu, X.-C.; Hereld, D.; Barth, C.; Savage, A.; Kiefel, B.R.; Lay, S.; Fisher, P.R.; Margolin, W.; Beech, P.L. Two Dictyostelium Orthologs of the Prokaryotic Cell Division Protein FtsZ Localize to Mitochondria and Are Required for the Maintenance of Normal Mitochondrial Morphology. Eukaryot. Cell 2003, 2, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Schimmel, B.G.; Berbusse, G.W.; Naylor, K. Mitochondrial Fission and Fusion in Dictyostelium Discoideum: A Search for Proteins Involved in Membrane Dynamics. BMC Res. Notes 2012, 5, 505. [Google Scholar] [CrossRef] [Green Version]
- Pârvu, M. Parasitic Fungi Sclerotiniaceae: Morphology and Ultrastructure. In Microscopy: Advances in Scientific Research and Education; Formatex Research Center: Badajoz, Spain, 2014; pp. 530–537. ISBN 978-849-421-343-4. [Google Scholar]
- Nunnari, J.; Marshall, W.F.; Straight, A.; Murray, A.; Sedat, J.W.; Walter, P. Mitochondrial Transmission during Mating in Saccharomyces Cerevisiae Is Determined by Mitochondrial Fusion and Fission and the Intramitochondrial Segregation of Mitochondrial DNA. Mol. Biol. Cell 1997, 8, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbowski, M.; Youle, R.J. Dynamics of Mitochondrial Morphology in Healthy Cells and during Apoptosis. Cell Death Differ. 2003, 10, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osteryoung, K.W.; Nunnari, J. The Division of Endosymbiotic Organelles. Science 2003, 302, 1698–1704. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.C. Mitochondrial Fusion and Fission in Mammals. Annu. Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trudeau, K.; Molina, A.J.A.; Roy, S. High Glucose Induces Mitochondrial Morphology and Metabolic Changes in Retinal Pericytes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8657. [Google Scholar] [CrossRef] [Green Version]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial Fission, Fusion, and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; McIntyre, R.L.; Janssens, G.E.; Houtkooper, R.H. Mitochondrial Fission and Fusion: A Dynamic Role in Aging and Potential Target for Age-Related Disease. Mech. Ageing Dev. 2020, 186, 111212. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.; Chang, X.; Zhang, X. Imbalance in Mitochondrial Dynamics Induced by Low PGC-1α Expression Contributes to Hepatocyte EMT and Liver Fibrosis. Cell Death Dis. 2020, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Downs, E.; Bottrell, A.D.; Naylor, K. Identifying the Effects of Reactive Oxygen Species on Mitochondrial Dynamics and Cytoskeleton Stability in Dictyostelium Discoideum. Cells 2021, 10, 2147. [Google Scholar] [CrossRef]
- Westermann, B. Mitochondrial Fusion and Fission in Cell Life and Death. Nat. Rev. Mol. Cell Biol. 2010, 11, 872–884. [Google Scholar] [CrossRef]
- Gomes, L.C.; Benedetto, G.D.; Scorrano, L. During Autophagy Mitochondria Elongate, Are Spared from Degradation and Sustain Cell Viability. Nat. Cell Biol. 2011, 13, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Mitra, K.; Sengupta, P.; Jarnik, M.; Lippincott-Schwartz, J.; Arias, I.M. Coordinated Elevation of Mitochondrial Oxidative Phosphorylation and Autophagy Help Drive Hepatocyte Polarization. Proc. Natl. Acad. Sci. USA 2013, 110, 7288–7293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, Y.-H.; Lin, S.-C.A.; Kuo, C.-H.; Li, C.-J. Molecular Machinery and Pathophysiology of Mitochondrial Dynamics. Front. Cell Dev. Biol. 2021, 9, 743892. [Google Scholar] [CrossRef] [PubMed]
- Madan, S.; Uttekar, B.; Chowdhary, S.; Rikhy, R. Mitochondria Lead the Way: Mitochondrial Dynamics and Function in Cellular Movements in Development and Disease. Front. Cell Dev. Biol. 2021, 9, 781933. [Google Scholar] [CrossRef]
- Fenton, A.R.; Jongens, T.A.; Holzbaur, E.L.F. Mitochondrial Dynamics: Shaping and Remodeling an Organelle Network. Curr. Opin. Cell Biol. 2021, 68, 28–36. [Google Scholar] [CrossRef]
- Fletcher, D.A.; Mullins, R.D. Cell Mechanics and the Cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Chernivec, E.; Cooper, J.; Naylor, K. Exploring the Effect of Rotenone—A Known Inducer of Parkinson’s Disease—On Mitochondrial Dynamics in Dictyostelium Discoideum. Cells 2018, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Knott, A.B.; Bossy-Wetzel, E. Impairing the Mitochondrial Fission and Fusion Balance: A New Mechanism of Neurodegeneration. Ann. N. Y. Acad. Sci. 2008, 1147, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chan, D.C. Mitochondrial Dynamics—Fusion, Fission, Movement, and Mitophagy--in Neurodegenerative Diseases. Hum. Mol. Genet. 2009, 18, R169–R176. [Google Scholar] [CrossRef]
- Reddy, P.H. Increased Mitochondrial Fission and Neuronal Dysfunction in Huntington’s Disease: Implications for Molecular Inhibitors of Excessive Mitochondrial Fission. Drug Discov. Today 2014, 19, 951–955. [Google Scholar] [CrossRef] [Green Version]
- Graef, M.; Nunnari, J. Mitochondria Regulate Autophagy by Conserved Signalling Pathways. EMBO J. 2011, 30, 2101–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 Coordinately Regulate Mitochondrial Fusion and Are Essential for Embryonic Development. J. Cell Biol. 2003, 160, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Iovine, J.C.; Claypool, S.M.; Alder, N.N. Mitochondrial Compartmentalization: Emerging Themes in Structure and Function. Trends Biochem. Sci. 2021, 46, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondrial Dynamics and Its Involvement in Disease. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 235–259. [Google Scholar] [CrossRef] [Green Version]
- McCarron, J.G.; Wilson, C.; Sandison, M.E.; Olson, M.L.; Girkin, J.M.; Saunter, C.; Chalmers, S. From Structure to Function: Mitochondrial Morphology, Motion and Shaping in Vascular Smooth Muscle. J. Vasc. Res. 2013, 50, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Westermann, B. Bioenergetic Role of Mitochondrial Fusion and Fission. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 1833–1838. [Google Scholar] [CrossRef] [Green Version]
- Twig, G.; Elorza, A.; Molina, A.J.A.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; Haigh, S.E.; Katz, S.; Las, G.; et al. Fission and Selective Fusion Govern Mitochondrial Segregation and Elimination by Autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Cagalinec, M.; Safiulina, D.; Liiv, M.; Liiv, J.; Choubey, V.; Wareski, P.; Veksler, V.; Kaasik, A. Principles of the Mitochondrial Fusion and Fission Cycle in Neurons. J. Cell Sci. 2013, 126, 2187–2197. [Google Scholar] [CrossRef] [Green Version]
- Sukhorukov, V.M.; Dikov, D.; Reichert, A.S.; Meyer-Hermann, M. Emergence of the Mitochondrial Reticulum from Fission and Fusion Dynamics. PLoS Comput. Biol. 2012, 8, e1002745. [Google Scholar] [CrossRef] [Green Version]
- Verstreken, P.; Ly, C.V.; Venken, K.J.T.; Koh, T.-W.; Zhou, Y.; Bellen, H.J. Synaptic Mitochondria Are Critical for Mobilization of Reserve Pool Vesicles at Drosophila Neuromuscular Junctions. Neuron 2005, 47, 365–378. [Google Scholar] [CrossRef] [Green Version]
- Seo, A.Y.; Joseph, A.-M.; Dutta, D.; Hwang, J.C.Y.; Aris, J.P.; Leeuwenburgh, C. New Insights into the Role of Mitochondria in Aging: Mitochondrial Dynamics and More. J. Cell Sci. 2010, 123, 2533–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakula, D.; Scheibye-Knudsen, M. MitophAging: Mitophagy in Aging and Disease. Front. Cell Dev. Biol. 2020, 8, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R.L.; Hollenbeck, P.J. Axonal Transport of Mitochondria along Microtubules and F-Actin in Living Vertebrate Neurons. J. Cell Biol. 1995, 131, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Saxton, W.M.; Hollenbeck, P.J. The Axonal Transport of Mitochondria. J. Cell Sci. 2012, 125, 2095–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melkov, A.; Abdu, U. Regulation of Long-Distance Transport of Mitochondria along Microtubules. Cell. Mol. Life Sci. 2018, 75, 163–176. [Google Scholar] [CrossRef]
- Martin, M.; Iyadurai, S.J.; Gassman, A.; Gindhart, J.G.; Hays, T.S.; Saxton, W.M. Cytoplasmic Dynein, the Dynactin Complex, and Kinesin Are Interdependent and Essential for Fast Axonal Transport. Mol. Biol. Cell 1999, 10, 3717–3728. [Google Scholar] [CrossRef] [Green Version]
- Hirokawa, N.; Takemura, R. Molecular Motors and Mechanisms of Directional Transport in Neurons. Nat. Rev. Neurosci. 2005, 6, 201–214. [Google Scholar] [CrossRef]
- Pilling, A.D.; Horiuchi, D.; Lively, C.M.; Saxton, W.M. Kinesin-1 and Dynein Are the Primary Motors for Fast Transport of Mitochondria in Drosophila Motor Axons. Mol. Biol. Cell 2006, 17, 2057–2068. [Google Scholar] [CrossRef] [Green Version]
- Kruppa, A.J.; Buss, F. Motor Proteins at the Mitochondria-Cytoskeleton Interface. J. Cell Sci. 2021, 134, jcs226084. [Google Scholar] [CrossRef]
- Fransson, S.; Ruusala, A.; Aspenström, P. The Atypical Rho GTPases Miro-1 and Miro-2 Have Essential Roles in Mitochondrial Trafficking. Biochem. Biophys. Res. Commun. 2006, 344, 500–510. [Google Scholar] [CrossRef]
- Wu, M.; Kalyanasundaram, A.; Zhu, J. Structural and Biomechanical Basis of Mitochondrial Movement in Eukaryotic Cells. Int. J. Nanomed. 2013, 8, 4033–4042. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Gerwin, C.; Sheng, Z.-H. Syntabulin-Mediated Anterograde Transport of Mitochondria along Neuronal Processes. J. Cell Biol. 2005, 170, 959–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligon, L.A.; Steward, O. Role of Microtubules and Actin Filaments in the Movement of Mitochondria in the Axons and Dendrites of Cultured Hippocampal Neurons. J. Comp. Neurol. 2000, 427, 351–361. [Google Scholar] [CrossRef]
- Berbusse, G.W.; Woods, L.C.; Vohra, B.P.S.; Naylor, K. Mitochondrial Dynamics Decrease Prior to Axon Degeneration Induced by Vincristine and Are Partially Rescued by Overexpressed CytNmnat1. Front. Cell. Neurosci. 2016, 10, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, Y.; Liu, Y.; Li, J.; Hao, Y.; Yin, P.; Liu, Z.; Chen, J.; Wang, Y.; Wang, N.; et al. Microtubule Associated Tumor Suppressor 1 Interacts with Mitofusins to Regulate Mitochondrial Morphology in Endothelial Cells. FASEB J. 2018, 32, 4504–4518. [Google Scholar] [CrossRef] [Green Version]
- Mattenberger, Y.; James, D.I.; Martinou, J.C. Fusion of Mitochondria in Mammalian Cells Is Dependent on the Mitochondrial Inner Membrane Potential and Independent of Microtubules or Actin. FEBS Lett. 2003, 538, 53–59. [Google Scholar] [CrossRef]
- Yaffe, M.P.; Harata, D.; Verde, F.; Eddison, M.; Toda, T.; Nurse, P. Microtubules Mediate Mitochondrial Distribution in Fission Yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 11664–11668. [Google Scholar] [CrossRef] [Green Version]
- Yaffe, M.P.; Stuurman, N.; Vale, R.D. Mitochondrial Positioning in Fission Yeast Is Driven by Association with Dynamic Microtubules and Mitotic Spindle Poles. Proc. Natl. Acad. Sci. USA 2003, 100, 11424–11428. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Jain, D.; Costa, J.; Velve-Casquillas, G.; Tran, P.T. Mmb1p Binds Mitochondria to Dynamic Microtubules. Curr. Biol. 2011, 21, 1431–1439. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Zheng, F.; Cheung, M.; Wang, F.; Fu, C. Fission Yeast Mitochondria Are Distributed by Dynamic Microtubules in a Motor-Independent Manner. Sci. Rep. 2015, 5, 11023. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, I.; Gachet, Y.; Hyams, J.S. The Dynamin Related Protein Dnm1 Fragments Mitochondria in a Microtubule-Dependent Manner during the Fission Yeast Cell Cycle. Cell Motil. Cytoskelet. 2009, 66, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Boldogh, I.R.; Pon, L.A. Mitochondria on the Move. Trends Cell Biol. 2007, 17, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Frederick, R.L.; McCaffery, J.M.; Cunningham, K.W.; Okamoto, K.; Shaw, J.M. Yeast Miro GTPase, Gem1p, Regulates Mitochondrial Morphology via a Novel Pathway. J. Cell Biol. 2004, 167, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frederick, R.L.; Okamoto, K.; Shaw, J.M. Multiple Pathways Influence Mitochondrial Inheritance in Budding Yeast. Genetics 2008, 178, 825–837. [Google Scholar] [CrossRef] [Green Version]
- Mehta, K.; Chacko, L.A.; Chug, M.K.; Jhunjhunwala, S.; Ananthanarayanan, V. Association of Mitochondria with Microtubules Inhibits Mitochondrial Fission by Precluding Assembly of the Fission Protein Dnm1. J. Biol. Chem. 2019, 294, 3385–3396. [Google Scholar] [CrossRef] [Green Version]
- Gutnick, A.; Banghart, M.R.; West, E.R.; Schwarz, T.L. The Light-Sensitive Dimerizer Zapalog Reveals Distinct Modes of Immobilization for Axonal Mitochondria. Nat. Cell Biol. 2019, 21, 768–777. [Google Scholar] [CrossRef]
- Straub, F.; Welz, T.; Alberico, H.; Brandão, R.O.; Huber, A.; Samol-Wolf, A.; Brakebusch, C.; Woods, D.; Kollmar, M.; Martin-Gonzalez, J.; et al. The SPIRE1 Actin Nucleator Coordinates Actin/Myosin Functions in the Regulation of Mitochondrial Motility. bioRxiv 2020. [Google Scholar] [CrossRef]
- López-Doménech, G.; Covill-Cooke, C.; Ivankovic, D.; Halff, E.F.; Sheehan, D.F.; Norkett, R.; Birsa, N.; Kittler, J.T. Miro Proteins Coordinate Microtubule- and Actin-dependent Mitochondrial Transport and Distribution. EMBO J. 2018, 37, 321–336. [Google Scholar] [CrossRef]
- Friedman, J.R.; Lackner, L.L.; West, M.; DiBenedetto, J.R.; Nunnari, J.; Voeltz, G.K. ER Tubules Mark Sites of Mitochondrial Division. Science 2011, 334, 358–362. [Google Scholar] [CrossRef] [Green Version]
- Korobova, F.; Ramabhadran, V.; Higgs, H.N. An Actin-Dependent Step in Mitochondrial Fission Mediated by the ER-Associated Formin INF2. Science 2013, 339, 464–467. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.S.; Wong, Y.C.; Simpson, C.L.; Holzbaur, E.L.F. Dynamic Actin Cycling through Mitochondrial Subpopulations Locally Regulates the Fission–Fusion Balance within Mitochondrial Networks. Nat. Commun. 2016, 7, 12886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vos, K.J.; Allan, V.J.; Grierson, A.J.; Sheetz, M.P. Mitochondrial Function and Actin Regulate Dynamin-Related Protein 1-Dependent Mitochondrial Fission. Curr. Biol. 2005, 15, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.R.; Swayne, T.C.; Pon, L.A. Actin-Dependent Mitochondrial Motility in Mitotic Yeast and Cell-Free Systems: Identification of a Motor Activity on the Mitochondrial Surface. J. Cell Biol. 1995, 130, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbacher, K.L.; Yang, H.-C.; Gay, A.C.; Huckaba, T.M.; Pon, L.A. Live Cell Imaging of Mitochondrial Movement along Actin Cables in Budding Yeast. Curr. Biol. 2004, 14, 1996–2004. [Google Scholar] [CrossRef] [Green Version]
- Boldogh, I.R.; Pon, L.A. Interactions of Mitochondria with the Actin Cytoskeleton. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Boldogh, I.R.; Yang, H.-C.; Nowakowski, W.D.; Karmon, S.L.; Hays, L.G.; Yates, J.R.; Pon, L.A. Arp2/3 Complex and Actin Dynamics Are Required for Actin-Based Mitochondrial Motility in Yeast. Proc. Natl. Acad. Sci. USA 2001, 98, 3162–3167. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Cheng, T.-H.; Lin, W.-L.; Chen, C.-L.; Yang, W.Y.; Blackstone, C.; Chang, C.-R. Srv2 Is a Pro-Fission Factor That Modulates Yeast Mitochondrial Morphology and Respiration by Regulating Actin Assembly. iScience 2019, 11, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Anesti, V.; Scorrano, L. The Relationship between Mitochondrial Shape and Function and the Cytoskeleton. Biochim. Biophys. Acta 2006, 1757, 692–699. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Ji, W.-K.; Stan, R.V.; de Juan Sanz, J.; Ryan, T.A.; Higgs, H.N. INF2-Mediated Actin Polymerization at the ER Stimulates Mitochondrial Calcium Uptake, Inner Membrane Constriction, and Division. J. Cell Biol. 2018, 217, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.S.; Holzbaur, E.L. Mitochondrial-Cytoskeletal Interactions: Dynamic Associations That Facilitate Network Function and Remodeling. Curr. Opin. Physiol. 2018, 3, 94–100. [Google Scholar] [CrossRef]
- Lin, T.-K.; Lin, K.-J.; Lin, K.-L.; Liou, C.-W.; Chen, S.-D.; Chuang, Y.-C.; Wang, P.-W.; Chuang, J.-H.; Wang, T.-J. When Friendship Turns Sour: Effective Communication between Mitochondria and Intracellular Organelles in Parkinson’s Disease. Front. Cell Dev. Biol. 2020, 8, 607392. [Google Scholar] [CrossRef] [PubMed]
- Korobova, F.; Gauvin, T.J.; Higgs, H.N. A Role for Myosin II in Mammalian Mitochondrial Fission. Curr. Biol. 2014, 24, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Svitkina, T.M. Ultrastructure and Dynamics of the Actin-Myosin II Cytoskeleton during Mitochondrial Fission. Nat. Cell Biol. 2019, 21, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cheng, Q.; Liu, L.; Zhou, T.; Shan, C.; Hu, X.; Zhou, J.; Liu, E.; Li, P.; Gao, N. Mitochondrial Translocation of Cofilin Is Required for Allyl Isothiocyanate-Mediated Cell Death via ROCK1/PTEN/PI3K Signaling Pathway. Cell Commun. Signal. 2013, 11, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehklau, K.; Hoffmann, L.; Gurniak, C.B.; Ott, M.; Witke, W.; Scorrano, L.; Culmsee, C.; Rust, M.B. Cofilin1-Dependent Actin Dynamics Control DRP1-Mediated Mitochondrial Fission. Cell Death Dis. 2017, 8, e3063. [Google Scholar] [CrossRef]
- Annesley, S.J.; Fisher, P.R. Dictyostelium Discoideum—a Model for Many Reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef]
- Eichinger, L.; Pachebat, J.A.; Glöckner, G.; Rajandream, M.-A.; Sucgang, R.; Berriman, M.; Song, J.; Olsen, R.; Szafranski, K.; Xu, Q.; et al. The Genome of the Social Amoeba Dictyostelium Discoideum. Nature 2005, 435, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Fey, P.; Dodson, R.J.; Basu, S.; Chisholm, R.L. One Stop Shop for Everything Dictyostelium: DictyBase and the Dicty Stock Center in 2012. Methods Mol. Biol. 2013, 983, 59–92. [Google Scholar] [CrossRef] [Green Version]
- DictyBase Home. Available online: http://dictybase.org/ (accessed on 25 July 2022).
- Berbusse, G.W. Investigating the Importance of Mitochondrial Dynamics: Fission, Fusion, Trafficking and Fragmentation in the Single-Celled Organism Dictyostelium Discoideum and CD-1 Mice In Vitro. Master’s Thesis, University of Central Arkansas, Conway, AR, USA, 2014. [Google Scholar]
- West, N.W. Investigation into the Role of the Dynamin-Like Proteins in the Mitochondrial Dynamics of Dictyostelium Discoideum via Cytoskeletal Disruption. Master’s Thesis, University of Central Arkansas, Conway, AR, USA, 2016. [Google Scholar]
- Loomis, W.F.; Insall, R.H. A Cell for All Reasons. Nature 1999, 401, 440–441. [Google Scholar] [CrossRef]
- Erickson, H.P. Dynamin and Ftsz: Missing Links in Mitochondrial and Bacterial Division. J. Cell Biol. 2000, 148, 1103–1106. [Google Scholar] [CrossRef] [Green Version]
- de Duve, C. The Birth of Complex Cells. Sci. Am. 1996, 274, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Fields, S.D.; Arana, Q.; Heuser, J.; Clarke, M. Mitochondrial Membrane Dynamics Are Altered in CluA− Mutants of Dictyostelium. J. Muscle Res. Cell Motil. 2002, 23, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Pollock, N.; Koonce, M.P.; de Hostos, E.L.; Vale, R.D. In Vitro Microtubule-Based Organelle Transport in Wild-Type Dictyostelium and Cells Overexpressing a Truncated Dynein Heavy Chain. Cell Motil. Cytoskelet. 1998, 40, 304–314. [Google Scholar] [CrossRef]
- Woods, L.C.; Berbusse, G.W.; Naylor, K. Microtubules Are Essential for Mitochondrial Dynamics–Fission, Fusion, and Motility–in Dictyostelium Discoideum. Front. Cell Dev. Biol. 2016, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlahou, G.; Eliáš, M.; von Kleist-Retzow, J.-C.; Wiesner, R.J.; Rivero, F. The Ras Related GTPase Miro Is Not Required for Mitochondrial Transport in Dictyostelium Discoideum. Eur. J. Cell Biol. 2011, 90, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chisholm, R.L. Cytoplasmic Dynein-Associated Structures Move Bidirectionally In Vivo. J. Cell Sci. 2002, 115, 1453–1460. [Google Scholar] [CrossRef]
- Ma, S.; Fey, P.; Chisholm, R.L. Molecular Motors and Membrane Traffic in Dictyostelium. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2001, 1525, 234–244. [Google Scholar] [CrossRef]
- Kollmar, M. Thirteen Is Enough: The Myosins of Dictyostelium Discoideum and Their Light Chains. BMC Genom. 2006, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- McInnes, J. Insights on Altered Mitochondrial Function and Dynamics in the Pathogenesis of Neurodegeneration. Transl. Neurodegener. 2013, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Francione, L.M.; Annesley, S.J.; Carilla-Latorre, S.; Escalante, R.; Fisher, P.R. The Dictyostelium Model for Mitochondrial Disease. Semin. Cell Dev. Biol. 2011, 22, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chan, D.C. Critical Dependence of Neurons on Mitochondrial Dynamics. Curr. Opin. Cell Biol. 2006, 18, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Whitley, B.N.; Engelhart, E.A.; Hoppins, S. Mitochondrial Dynamics and Their Potential as a Therapeutic Target. Mitochondrion 2019, 49, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Esteves, A.R.; Gozes, I.; Cardoso, S.M. The Rescue of Microtubule-Dependent Traffic Recovers Mitochondrial Function in Parkinson’s Disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 7–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Function |
|---|---|
| Kinesin-1 | Microtubule motor protein |
| Kinesin-3 | Microtubule motor protein |
| Cytoplasmic dynein | Microtubule motor protein |
| TRAK1/TRAK2 | Adaptors linking microtubule motors to mitochondria |
| Miro (Mitochondrial Rho GTPase) | Adaptor that connects some TRAK1/2 adaptors to the mitochondria |
| Syntabulin | Adaptor between microtubules and kinesin-1 |
| KIFBP (Kinesin Binding Protein) | Kinesin-3 adaptor linking motor to microtubules |
| Mitofusins (Mfn1/2) | GTPase responsible for mitochondrial fusion |
| Mtus1 (Microtubule-associated tumor suppressor 1) | Interacts with mitofusins to mediate fusion |
| Drp1 (Dynamin related protein 1) | GTPase responsible for mitochondrial fission |
| Model | Motility | Fission | Fusion | Citations |
|---|---|---|---|---|
| Mammals | Microtubules used as tracks primarily, but actin is also sometimes used as tracks in neurons for short distances. Actin may also anchor mitochondria in place. Unclear if actin-based motility is based on motors or actin dynamics. | Microtubules also affect rate of fission, mechanism unclear. Actin directly involved in fission initiation at sites of ER-mitochondria contact. Actin polymerization via INF2 but not ring formation drives mitochondrial constriction and then recruits Drp1 to complete scission of the organelle. | Microtubules affect rate of fusion, mechanism unclear; Actin attachment to mitochondria prohibits fusion. | [44,45,46,50,54,55,56,61,67,68,69,70,71,72,73] |
| Fission yeast | Microtubules used as tracks, Microtubule dynamics move mitochondria. No actin-based motility. | Microtubules block Dnm1-mediated fission; unclear if actin is involved. | Currently unknown | [58,59,60,61,62,66] |
| Budding yeast | No microtubule-based motility. Actin used as tracks, dynamics are the predominant motility mechanism. | Actin likely plays a role as evidenced by pro-fission actin regulatory protein-Srv2. | Currently unknown | [65,74,75,76,77,78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, M.D.; Naylor, K. Simple to Complex: The Role of Actin and Microtubules in Mitochondrial Dynamics in Amoeba, Yeast, and Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 9402. https://doi.org/10.3390/ijms23169402
Jones MD, Naylor K. Simple to Complex: The Role of Actin and Microtubules in Mitochondrial Dynamics in Amoeba, Yeast, and Mammalian Cells. International Journal of Molecular Sciences. 2022; 23(16):9402. https://doi.org/10.3390/ijms23169402
Chicago/Turabian StyleJones, Meghan D., and Kari Naylor. 2022. "Simple to Complex: The Role of Actin and Microtubules in Mitochondrial Dynamics in Amoeba, Yeast, and Mammalian Cells" International Journal of Molecular Sciences 23, no. 16: 9402. https://doi.org/10.3390/ijms23169402
APA StyleJones, M. D., & Naylor, K. (2022). Simple to Complex: The Role of Actin and Microtubules in Mitochondrial Dynamics in Amoeba, Yeast, and Mammalian Cells. International Journal of Molecular Sciences, 23(16), 9402. https://doi.org/10.3390/ijms23169402