
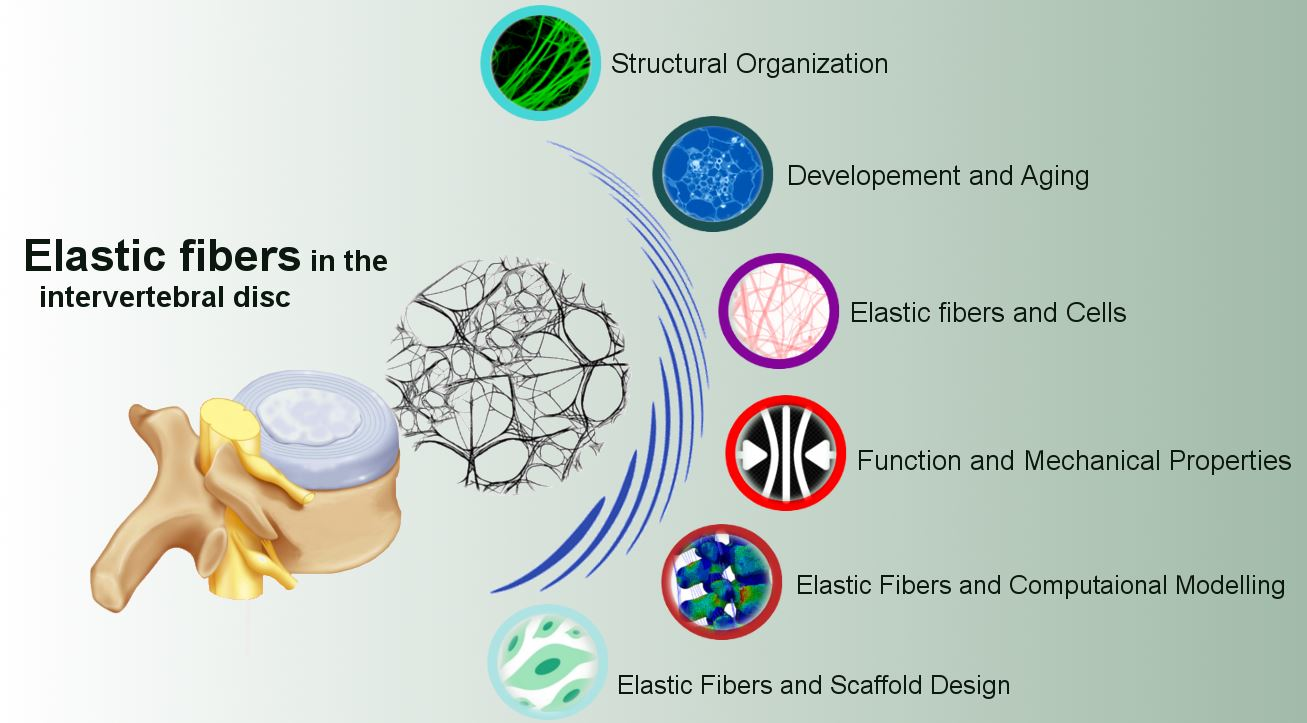
Elastic Fibers in the Intervertebral Disc: From Form to Function and toward Regeneration


 and
and 
Abstract

1. Introduction
2. Review Methodology
3. Multi-Scale Hierarchical Structure of the IVD Elastic Fibers
4. IVD Elastin Content
5. Development and Aging
6. IVD Elastic Fibers and Cells
7. Function and Mechanical Properties of IVD Elastic Fibers
7.1. Structural Analysis under Micromechanical Loading
7.2. Elastin Enzymatic Digestion and Micromechanical Loading
7.3. ECM Alkali Digestion and Micromechanical Loading
8. Future Outlook
8.1. Elastic Fibers and Full IVD Scaffold Design
8.2. Refined Computational Modeling
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Humzah, M.D.; Soames, R.W. Human intervertebral disc: Structure and function. Anat. Rec. 1988, 220, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, J.; Hiltner, A.; Baer, E. Hierarchical structure of the intervertebral disc. Connect. Tissue Res. 1989, 23, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Elliott, D.M.; Costi, J.J. Structure and mechanical function of the inter-lamellar matrix of the annulus fibrosus in the disc. J. Orthop. Res. 2016, 34, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Costi, J.J. Ultrastructural organization of elastic fibres in the partition boundaries of the annulus fibrosus within the intervertebral disc. Acta Biomater. 2018, 68, 67–77. [Google Scholar] [CrossRef]
- Schollum, M.L.; Robertson, P.A.; Broom, N.D. ISSLS prize winner: Microstructure and mechanical disruption of the lumbar disc annulus: Part I: A microscopic investigation of the translamellar bridging network. Spine 2008, 33, 2702–2710. [Google Scholar] [CrossRef]
- Yu, J.; Tirlapur, U.; Fairbank, J.; Handford, P.; Roberts, S.; Winlove, C.P.; Cui, Z.; Urban, J. Microfibrils, elastin fibres and collagen fibres in the human intervertebral disc and bovine tail disc. J. Anat. 2007, 210, 460–471. [Google Scholar] [CrossRef]
- Iatridis, J.C.; Weidenbaum, M.; Setton, L.A.; Mow, V.C. Is the nucleus pulposus a solid or a fluid? Mechanical behaviors of the nucleus pulposus of the human intervertebral disc. Spine 1996, 21, 1174–1184. [Google Scholar] [CrossRef]
- Eyre, D.R.; Muir, H. Types I and II collagens in intervertebral disc. Interchanging radial distributions in annulus fibrosus. Biochem. J. 1976, 157, 267. [Google Scholar] [CrossRef]
- Eyre, D.; Matsui, Y.; Wu, J.-J. Collagen polymorphisms of the intervertebral disc. Biochem. Soc. Trans. 2002, 30, 844–848. [Google Scholar] [CrossRef]
- Sivan, S.S.; Hayes, A.J.; Wachtel, E.; Caterson, B.; Merkher, Y.; Maroudas, A.; Brown, S.; Roberts, S. Biochemical composition and turnover of the extracellular matrix of the normal and degenerate intervertebral disc. Eur. Spine J. 2014, 23, 344–353. [Google Scholar] [CrossRef]
- Tavakoli, J.; Diwan, A.D.; Tipper, J.L. Advanced Strategies for the Regeneration of Lumbar Disc Annulus Fibrosus. Int. J. Mol. Sci. 2020, 21, 4889. [Google Scholar] [CrossRef] [PubMed]
- Vergroesen, P.P.A.; Kingma, I.; Emanuel, K.S.; Hoogendoorn, R.J.W.; Welting, T.J.; van Royen, B.J.; van Dieën, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cartil. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Luo, R.; Li, G.; Zhang, W.; Song, Y.; Yang, C. The Proteolysis of ECM in Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2022, 23, 1715. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; An, H.-J.; Yeo, H.; Jeong, Y.; Lee, H.; Lee, J.; Nam, K.; Lee, J.; Shin, D.-E.; Lee, S. Activation of Hypoxia-Inducible Factor-1α Signaling Pathway Has the Protective Effect of Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2021, 22, 11355. [Google Scholar] [CrossRef] [PubMed]
- Eckersley, A.; Ozols, M.; Chen, P.; Tam, V.; Hoyland, J.A.; Trafford, A.; Chan, D.; Sherratt, M.J. Peptide location fingerprinting reveals tissue region-specific differences in protein structures in an ageing human organ. Int. J. Mol. Sci. 2021, 22, 10408. [Google Scholar] [CrossRef] [PubMed]
- Takeoka, Y.; Paladugu, P.; Kang, J.D.; Mizuno, S. Augmented Chondroitin Sulfate Proteoglycan Has Therapeutic Potential for Intervertebral Disc Degeneration by Stimulating Anabolic Turnover in Bovine Nucleus Pulposus Cells under Changes in Hydrostatic Pressure. Int. J. Mol. Sci. 2021, 22, 6015. [Google Scholar] [CrossRef]
- Krupkova, O.; Hlavna, M.; Amir Tahmasseb, J.; Zvick, J.; Kunz, D.; Ito, K.; Ferguson, S.J.; Wuertz-Kozak, K. An inflammatory nucleus pulposus tissue culture model to test molecular regenerative therapies: Validation with epigallocatechin 3-gallate. Int. J. Mol. Sci. 2016, 17, 1640. [Google Scholar] [CrossRef]
- Li, W.; Liu, T.; Wu, L.; Chen, C.; Jia, Z.; Bai, X.; Ruan, D. Blocking the function of inflammatory cytokines and mediators by using IL-10 and TGF-β: A potential biological immunotherapy for intervertebral disc degeneration in a beagle model. Int. J. Mol. Sci. 2014, 15, 17270–17283. [Google Scholar] [CrossRef]
- Sharabi, M.; Wade, K.; Haj-Ali, R. Chapter 7—The Mechanical Role of Collagen Fibers in the Intervertebral Disc. In Biomechanics of the Spine; Galbusera, F., Wilke, H.-J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 105–123. [Google Scholar]
- Ushiki, T. Collagen fibers, reticular fibers and elastic fibers. A comprehensive understanding from a morphological viewpoint. Arch. Histol. Cytol. 2002, 65, 109–126. [Google Scholar] [CrossRef]
- Greenlee, T.K., Jr.; Ross, R.; Hartman, J.L. The fine structure of elastic fibers. J. Cell Biol. 1966, 30, 59–71. [Google Scholar] [CrossRef]
- Greenwald, J.; Riek, R. Biology of Amyloid: Structure, Function, and Regulation. Structure 2010, 18, 1244–1260. [Google Scholar] [CrossRef] [PubMed]
- Wagenseil, J.E.; Mecham, R.P. Vascular extracellular matrix and arterial mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef] [PubMed]
- Sherratt, M.J. Tissue elasticity and the ageing elastic fibre. Age 2009, 31, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Lacoste, J.-Y.; Chanez, P.; Vic, P.; Godard, P.; Michel, F.-B. Bronchial elastic fibers in normal subjects and asthmatic patients. Am. J. Respir. Crit. Care Med. 1996, 153, 1648–1654. [Google Scholar] [CrossRef]
- Hill, J.R.; Eekhoff, J.D.; Brophy, R.H.; Lake, S.P. Elastic fibers in orthopedics: Form and function in tendons and ligaments, clinical implications, and future directions. J. Orthop. Res. 2020, 38, 2305–2317. [Google Scholar] [CrossRef]
- Yanagisawa, H.; Wagenseil, J. Elastic fibers and biomechanics of the aorta: Insights from mouse studies. Matrix Biol. 2020, 85, 160–172. [Google Scholar] [CrossRef]
- Halabi, C.M.; Kozel, B.A. Vascular elastic fiber heterogeneity in health and disease. Curr. Opin. Hematol. 2020, 27, 190. [Google Scholar] [CrossRef]
- Langton, A.K.; Alessi, S.; Hann, M.; Chien, A.L.-L.; Kang, S.; Griffiths, C.E.M.; Watson, R.E.B. Aging in skin of color: Disruption to elastic fiber organization is detrimental to skin’s biomechanical function. J. Investig. Dermatol. 2019, 139, 779–788. [Google Scholar] [CrossRef]
- Schmelzer, C.E.H.; Duca, L. Elastic fibers: Formation, function, and fate during aging and disease. FEBS J. 2022, 289, 3704–3730. [Google Scholar] [CrossRef]
- Sharabi, M. Structural Mechanisms in Soft Fibrous Tissues: A Review. Front. Mater. 2022, 8, 793647. [Google Scholar] [CrossRef]
- Yu, J. Elastic tissues of the intervertebral disc. In Portland Press Ltd. Biochem. Soc. Trans. 2002, 30, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, C.; Schajowicz, F. Studies on structural changes in the lumbar annulus fibrosus. Acta Orthop. Scand. 1952, 22, 184–231. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.; Cooper, R.; Maynard, J. Elastic fibers in human intervertebral discs. J. Bone Jt. Surg. 1976, 58, 73–76. [Google Scholar] [CrossRef]
- Sylvest, J.; Hentzer, B.; Kobayasi, T. Ultrastructure of prolapsed disc. Acta Orthop. Scand. 1977, 48, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Hickey, D.S.; Hukins, D.W. Collagen fibril diameters and elastic fibres in the annulus fibrosus of human fetal intervertebral disc. J. Anat. 1981, 133, 351–357. [Google Scholar]
- Johnson, E.F.; Chetty, K.; Moore, I.M.; Stewart, A.; Jones, W. The distribution and arrangement of elastic fibres in the intervertebral disc of the adult human. J. Anat. 1982, 135, 301–309. [Google Scholar]
- Johnson, E.F.; Caldwell, R.W.; Berryman, H.E.; Miller, A.; Chetty, K. Elastic fibers in the anulus fibrosus of the dog intervertebral disc. Cells Tissues Organs 1984, 118, 238–242. [Google Scholar] [CrossRef]
- Johnson, E.F.; Berryman, H.; Mitchell, R.; Wood, W.B. Elastic fibres in the anulus fibrosus of the adult human lumbar intervertebral disc. A preliminary report. J. Anat. 1985, 143, 57–63. [Google Scholar]
- Mikawa, Y.; Hamagami, H.; Shikata, J.; Yamamuro, T. Elastin in the human intervertebral disk. Arch. Orthop. Trauma. Surg. 1986, 105, 343–349. [Google Scholar] [CrossRef]
- Yu, J.; Peter, C.; Roberts, S.; Urban, J.P. Elastic fibre organization in the intervertebral discs of the bovine tail. J. Anat. 2002, 201, 465–475. [Google Scholar] [CrossRef]
- Smith, L.J.; Fazzalari, N.L. Regional variations in the density and arrangement of elastic fibres in the anulus fibrosus of the human lumbar disc. J. Anat. 2006, 209, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Fairbank, J.C.T.; Roberts, S.; Urban, J.P.G. The Elastic Fiber Network of the Anulus Fibrosus of the Normal and Scoliotic Human Intervertebral Disc. Spine 2005, 30, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Davies, J.R.; Caterson, B. Ultrastructural Localization and Distribution of Proteoglycan in Normal and Scoliotic Lumbar Disc. Spine 2005, 30, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Davies, J.R.; Caterson, B. Ultrastructural Immunolocalization of α-Elastin and Keratan Sulfate Proteoglycan in Normal and Scoliotic Lumbar Disc. Spine 2005, 30, 1762–1769. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.J.; Fazzalari, N.L. The elastic fibre network of the human lumbar anulus fibrosus: Architecture, mechanical function and potential role in the progression of intervertebral disc degeneration. Eur. Spine J. 2009, 18, 439–448. [Google Scholar] [CrossRef]
- Yu, J.; Schollum, M.L.; Wade, K.R.; Broom, N.D.; Urban, J.P.G. ISSLS Prize Winner: A Detailed Examination of the Elastic Network Leads to a New Understanding of Annulus Fibrosus Organization. Spine 2015, 40, 1149–1157. [Google Scholar] [CrossRef]
- Tavakoli, J.; Costi, J.J. Development of a rapid matrix digestion technique for ultrastructural analysis of elastic fibers in the intervertebral disc. J. Mech. Behav. Biomed. Mater. 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Tavakoli, J.; Costi, J. A method for visualization and isolation of elastic fibres in annulus fibrosus of the disc. Mater. Sci. Eng. C 2018, 93, 299–304. [Google Scholar] [CrossRef]
- Tavakoli, J.; Elliott, D.; Costi, J. The ultra-structural organization of the elastic network in the intra-and inter-lamellar matrix of the intervertebral disc. Acta Biomater. 2017, 58, 269–277. [Google Scholar] [CrossRef]
- Tavakoli, J.; Diwan, A.D.; Tipper, J.L. Elastic fibers: The missing key to improve engineering concepts for reconstruction of the nucleus pulposus in the intervertebral disc. Acta Biomater. 2020, 113, 407–416. [Google Scholar] [CrossRef]
- Tavakoli, J.; Diwan, A.D.; Tipper, J.L. The ultrastructural organization of elastic fibers at the interface of the nucleus and annulus of the intervertebral disc. Acta Biomater. 2020, 114, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Tipper, J.L. Detailed mechanical characterization of the transition zone: New insight into the integration between the annulus and nucleus of the intervertebral disc. Acta Biomater. 2022, 143, 87–99. [Google Scholar] [CrossRef]
- Cloyd, J.M.; Elliott, D.M. Elastin content correlates with human disc degeneration in the anulus fibrosus and nucleus pulposus. Spine 2007, 32, 1826–1831. [Google Scholar] [CrossRef] [PubMed]
- Kobielarz, M.; Szotek, S.; Głowacki, M.; Dawidowicz, J.; Pezowicz, C. Qualitative and quantitative assessment of collagen and elastin in annulus fibrosus of the physiologic and scoliotic intervertebral discs. J. Mech. Behav. Biomed. Mater. 2016, 62, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Mecham, R.P. Elastic fiber structure and assembly. In Extracellular Matrix Assembly and Structure; Academic Press: Cambridge, MA, USA, 1994; pp. 218–310. [Google Scholar]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell Sci. 2002, 115, 2817–2828. [Google Scholar] [CrossRef]
- Kielty, C.M.; Baldock, C.; Lee, D.; Rock, M.J.; Ashworth, J.L.; Shuttleworth, C.A. Fibrillin: From microfibril assembly to biomechanical function. Philosophical Transactions of the Royal Society of London. Ser. B Biol. Sci. 2002, 357, 207–217. [Google Scholar] [CrossRef]
- Penner, A.S.; Rock, M.J.; Kielty, C.M.; Shipley, J.M. Microfibril-associated glycoprotein-2 interacts with fibrillin-1 and fibrillin-2 suggesting a role for MAGP-2 in elastic fiber assembly. J. Biol. Chem. 2002, 277, 35044–35049. [Google Scholar] [CrossRef]
- Kielty, C.M. Elastic fibres in health and disease. Expert Rev. Mol. Med. 2006, 8, 1. [Google Scholar] [CrossRef]
- Bressan, G.M.; Daga-Gordini, D.; Colombatti, A.; Castellani, I.; Marigo, V.; Volpin, D. Emilin, a component of elastic fibers preferentially located at the elastin-microfibrils interface. J. Cell Biol. 1993, 121, 201–212. [Google Scholar] [CrossRef]
- Hayes, A.J.; Lord, M.S.; Smith, S.M.; Smith, M.M.; Whitelock, J.M.; Weiss, A.S.; Melrose, J. Colocalization in vivo and association in vitro of perlecan and elastin. Histochem. Cell Biol. 2011, 136, 437. [Google Scholar] [CrossRef]
- Buckwalter, J.A. Do intervertebral discs deserve their bad reputation? Iowa Orthop. J. 1998, 18, 1–11. [Google Scholar] [PubMed]
- Postacchini, F.; Bellocci, M.; Massobrio, M. Morphologic Changes in Annulus Fibrosus During Aging An Ultrastructural Study in Rats. Spine 1984, 9, 596–603. [Google Scholar] [CrossRef]
- Olczyk, K. Age-related changes of elastin content in human intervertebral discs. Folia Histochem. et Cytobiol. 1994, 32, 41–44. [Google Scholar]
- Melrose, J.; Smith, S.M.; Appleyard, R.C.; Little, C.B. Aggrecan, versican and type VI collagen are components of annular translamellar crossbridges in the intervertebral disc. Eur. Spine J. 2008, 17, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Han, S.K.; Chen, C.-W.; Wierwille, J.; Chen, Y.; Hsieh, A.H. Three dimensional mesoscale analysis of translamellar cross-bridge morphologies in the annulus fibrosus using optical coherence tomography. J. Orthop. Res. 2015, 33, 304–311. [Google Scholar] [CrossRef]
- Sivan, S.-S.; van El, B.; Merkher, Y.; Schmelzer, C.E.H.; Zuurmond, A.-M.; Heinz, A.; Wachtel, E.; Varga, P.-P.; Lazary, A.; Brayda-Bruno, M.; et al. Longevity of elastin in human intervertebral disc as probed by the racemization of aspartic acid. Biochim. Biophys. Acta 2012, 1820, 1671–1677. [Google Scholar] [CrossRef]
- Fontes, R.B.; Baptista, J.S.; Rabbani, S.R.; Traynelis, V.C.; Liberti, E.A. Normal aging in human lumbar discs: An ultrastructural comparison. PLoS ONE 2019, 14, e0218121. [Google Scholar] [CrossRef]
- Midwood, K.S.; Schwarzbauer, J.E. Elastic Fibers: Building Bridges Between Cells and Their Matrix. Current Biology 2002, 12, R279–R281. [Google Scholar] [CrossRef]
- Jensen, S.A.; Robertson, I.B.; Handford, P.A. Dissecting the Fibrillin Microfibril: Structural Insights into Organization and Function. Structure 2012, 20, 215–225. [Google Scholar] [CrossRef]
- Whitelock, J.M.; Melrose, J.; Iozzo, R.V. Diverse cell signaling events modulated by perlecan. Biochemistry 2008, 47, 11174–11183. [Google Scholar] [CrossRef]
- Hayes, A.J.; Smith, S.M.; Melrose, J. Comparative immunolocalisation of fibrillin-1 and perlecan in the human foetal, and HS-deficient hspg2 exon 3 null mutant mouse intervertebral disc. Histochem. Cell Biol. 2013, 139, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Hayes, A.J.; Whitelock, J.M.; Little, C.B. Perlecan, the “jack of all trades” proteoglycan of cartilaginous weight-bearing connective tissues. Bioessays 2008, 30, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V. Basement membrane proteoglycans: From cellar to ceiling. Nat. Rev. Mol. Cell Biol. 2005, 6, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, S.S.; Cain, S.A.; Morgan, A.; Dallas, S.L.; Shuttleworth, C.A.; Kielty, C.M. Fibrillin-1 regulates the bioavailability of TGFβ1. J. Cell Biol. 2007, 176, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, A.H.; Twomey, J.D. Cellular mechanobiology of the intervertebral disc: New directions and approaches. J. Biomech. 2010, 43, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Bruehlmann, S.B.; Rattner, B.J.; Matyas, R.J.; Duncan, A.N. Regional variations in the cellular matrix of the annulus fibrosus of the intervertebral disc. J. Anat. 2002, 201, 159–171. [Google Scholar] [CrossRef]
- Bruehlmann, S.B.; Hulme, P.A.; Duncan, N.A. In situ intercellular mechanics of the bovine outer annulus fibrosus subjected to biaxial strains. J. Biomech. 2004, 37, 223–231. [Google Scholar] [CrossRef]
- Melrose, J.; Ghosh, P.; Taylor, T.K. A comparative analysis of the differential spatial and temporal distributions of the large (aggrecan, versican) and small (decorin, biglycan, fibromodulin) proteoglycans of the intervertebral disc. J. Anat. 2001, 198, 3–15. [Google Scholar] [CrossRef]
- Iatridis, J.C.; Gwynn, I. Mechanisms for mechanical damage in the intervertebral disc annulus fibrosus. J. Biomech. 2004, 37, 1165–1175. [Google Scholar] [CrossRef]
- Iatridis, J.C.; Kumar, S.; Foster, R.J.; Weidenbaum, M.; Mow, V.C. Shear mechanical properties of human lumbar annulus fibrosus. J. Orthop. Res. 1999, 17, 732–737. [Google Scholar] [CrossRef]
- Pezowicz, C.A.; Robertson, P.A.; Broom, N.D. The structural basis of interlamellar cohesion in the intervertebral disc wall. J. Anat. 2006, 208, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Pezowicz, C.A.; Robertson, P.A.; Broom, N.D. Intralamellar relationships within the collagenous architecture of the annulus fibrosus imaged in its fully hydrated state. J. Anat. 2005, 207, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Derby, R.; Chen, Y.; Seo, K.S.; Kim, M.J. In vitro measurement of pressure in intervertebral discs and annulus fibrosus with and without annular tears during discography. Spine J. 2004, 4, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Osti, O.; Vernon-Roberts, B.; Moore, R.; Fraser, R. Annular tears and disc degeneration in the lumbar spine. A post-mortem study of 135 discs. J. Bone Jt. Surg. 1992, 74, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Gregory, D.E.; Bae, W.C.; Sah, R.L.; Masuda, K. Anular delamination strength of human lumbar intervertebral disc. Eur. Spine J. 2012, 21, 1716–1723. [Google Scholar] [CrossRef]
- Gregory, D.E.; Bae, W.C.; Sah, R.L.; Masuda, K. Disc degeneration reduces the delamination strength of the annulus fibrosus in the rabbit annular disc puncture model. Spine J. 2014, 14, 1265–1271. [Google Scholar] [CrossRef]
- Mengoni, M.; Luxmoore, B.J.; Wijayathunga, V.N.; Jones, A.C.; Broom, N.D.; Wilcox, R.K. Derivation of inter-lamellar behaviour of the intervertebral disc annulus. J. Mech. Behav. Biomed. Mater. 2015, 48, 164–172. [Google Scholar] [CrossRef]
- Veres, S.P.; Robertson, P.A.; Broom, N.D. ISSLS prize winner: Microstructure and mechanical disruption of the lumbar disc annulus: Part II: How the annulus fails under hydrostatic pressure. Spine 2008, 33, 2711–2720. [Google Scholar] [CrossRef]
- Pezowicz, C.A.; Schechtman, H.; Robertson, P.A.; Broom, N.D. Mechanisms of anular failure resulting from excessive intradiscal pressure: A microstructural-micromechanical investigation. Spine 2006, 31, 2891–2903. [Google Scholar] [CrossRef]
- Wade, K.R.; Robertson, P.A.; Broom, N.D. On how nucleus-endplate integration is achieved at the fibrillar level in the ovine lumbar disc. J. Anat. 2012, 221, 39–46. [Google Scholar] [CrossRef]
- Wade, K.R.; Robertson, P.A.; Broom, N.D. A fresh look at the nucleus-endplate region: New evidence for significant structural integration. Eur. Spine J. 2011, 20, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Wade, K.R.; Robertson, P.A.; Broom, N.D. On the extent and nature of nucleus-annulus integration. Spine 2012, 37, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Vergari, C.; Mansfield, J.; Meakin, J.R.; Winlove, P.C. Lamellar and fibre bundle mechanics of the annulus fibrosus in bovine intervertebral disc. Acta Biomater. 2016, 37, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli, J.; Costi, J.J. New findings confirm the viscoelastic behaviour of the inter-lamellar matrix of the disc annulus fibrosus in radial and circumferential directions of loading. Acta Biomater. 2018, 71, 411–419. [Google Scholar] [CrossRef]
- Tavakoli, J.; Amin, D.B.; Freeman, B.J.C.; Costi, J.J. The Biomechanics of the Inter-Lamellar Matrix and the Lamellae During Progression to Lumbar Disc Herniation: Which is the Weakest Structure? Ann. Biomed. Eng. 2018, 46, 1280–1291. [Google Scholar] [CrossRef]
- Disney, C.M.; Eckersley, A.; McConnell, J.C.; Geng, H.; Bodey, A.J.; Hoyland, J.A.; Lee, P.D.; Sherratt, M.J.; Bay, B.K. Synchrotron tomography of intervertebral disc deformation quantified by digital volume correlation reveals microstructural influence on strain patterns. Acta Biomater. 2019, 92, 290–304. [Google Scholar] [CrossRef]
- Smith, L.J.; Byers, S.; Costi, J.J.; Fazzalari, N.L. Elastic fibers enhance the mechanical integrity of the human lumbar anulus fibrosus in the radial direction. Ann. Biomed. Eng. 2008, 36, 214–223. [Google Scholar] [CrossRef]
- Michalek, A.J.; Buckley, M.R.; Bonassar, L.J.; Cohen, I.; Iatridis, J.C. Measurement of local strains in intervertebral disc anulus fibrosus tissue under dynamic shear: Contributions of matrix fiber orientation and elastin content. J. Biomech. 2009, 42, 2279–2285. [Google Scholar] [CrossRef]
- Isaacs, J.L.; Vresilovic, E.; Sarkar, S.; Marcolongo, M. Role of biomolecules on annulus fibrosus micromechanics: Effect of enzymatic digestion on elastic and failure properties. J. Mech. Behav. Biomed. Mater. 2014, 40, 75–84. [Google Scholar] [CrossRef]
- Tavakoli, J.; Costi, J.J. New insights into the viscoelastic and failure mechanical properties of the elastic fiber network of the inter-lamellar matrix in the annulus fibrosus of the disc. Acta Biomater. 2018, 77, 292–300. [Google Scholar] [CrossRef]
- Silva-Correia, J.; Correia, S.I.; Oliveira, J.M.; Reis, R.L. Tissue engineering strategies applied in the regeneration of the human intervertebral disk. Biotechnol. Adv. 2013, 31, 1514–1531. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.; Clouet, J.; Le Bideau, J.; Le Visage, C.; Guicheux, J. Innovative strategies for intervertebral disc regenerative medicine: From cell therapies to multiscale delivery systems. Biotechnol. Adv. 2018, 36, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Sola, A.; Bellucci, D.; Cannillo, V. Functionally graded materials for orthopedic applications—An update on design and manufacturing. Biotechnol. Adv. 2016, 34, 504–531. [Google Scholar] [CrossRef] [PubMed]
- Skaggs, D.L.; Weidenbaum, M.; Latridis, J.C.; Ratcliffe, A.; Mow, V.C. Regional Variation in Tensile Properties and Biochemical Composition of the Human Lumbar Anulus Fibrosus. Spine 1994, 19, 1310–1319. [Google Scholar] [CrossRef] [PubMed]
- Trewhella, J. Recent advances in small-angle scattering and its expanding impact in structural biology. Structure 2022, 30, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Gronenborn, A.M. Small, but powerful and attractive: 19F in biomolecular NMR. Structure 2022, 30, 6–14. [Google Scholar] [CrossRef]
- Chiu, W.; Baker, M.L.; Jiang, W.; Dougherty, M.; Schmid, M.F. Electron Cryomicroscopy of Biological Machines at Subnanometer Resolution. Structure 2005, 13, 363–372. [Google Scholar] [CrossRef]
- Arnaout, M.A. The Structural Basis of Elasticity in Fibrillin-Based Microfibrils. Structure 2004, 12, 734–736. [Google Scholar] [CrossRef][Green Version]
- Morris, M.; Colleen, A.; Mervis, P.D. Carolyn B, Williams syndrome and related disorders. Annu. Rev. Genom. Hum. Genet. 2000, 1, 461–484. [Google Scholar] [CrossRef]
- Urbán, Z.; Zhang, J.; Davis, E.C.; Maeda, G.K.; Kumar, A.; Stalker, H.; Belmont, J.W.; Boyd, C.D.; Wallace, M.R. Supravalvular aortic stenosis: Genetic and molecular dissection of a complex mutation in the elastin gene. Hum. Genet. 2001, 109, 512–520. [Google Scholar] [CrossRef]
- Yee, A.; Lam, M.; Tam, V.; Chan, W.; Chu, I.; Cheah, K.; Cheung, K.; Chan, D. Fibrotic-like changes in degenerate human intervertebral discs revealed by quantitative proteomic analysis. Osteoarthr. Cartil. 2016, 24, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, M.; Liu, L.; Zhang, Y.; Zhu, Q.; Huang, C.; Wang, H.; Zhang, Y.; Li, H.; Li, C. Molecular basis of degenerative spinal disorders from a proteomic perspective. Mol. Med. Rep. 2020, 21, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, S.; Tangavel, C.; Soundararajan, D.C.R.; Nayagam, S.M.; Matchado, M.S.; Muthurajan, R.; Anand, K.; Rajendran, S.; Shetty, A.P.; Kanna, R.M.; et al. Proteomic Signatures of Healthy Intervertebral Discs From Organ Donors: A Comparison With Previous Studies on Discs From Scoliosis, Animals, and Trauma. Neurospine 2020, 17, 426–442. [Google Scholar] [CrossRef] [PubMed]
- Teodorescu, M.; Bercea, M.; Morariu, S. Biomaterials of PVA and PVP in medical and pharmaceutical applications: Perspectives and challenges. Biotechnol. Adv. 2019, 37, 109–131. [Google Scholar] [CrossRef]
- Bacakova, L.; Filova, E.; Parizek, M.; Ruml, T.; Svorcik, V. Modulation of cell adhesion, proliferation and differentiation on materials designed for body implants. Biotechnol. Adv. 2011, 29, 739–767. [Google Scholar] [CrossRef]
- Cheng, C.W.; Solorio, L.D.; Alsberg, E. Decellularized tissue and cell-derived extracellular matrices as scaffolds for orthopaedic tissue engineering. Biotechnol. Adv. 2014, 32, 462–484. [Google Scholar] [CrossRef]
- Tavakoli, J. Tissue Engineering of the Intervertebral Disc’s Annulus Fibrosus: A Scaffold-Based Review Study. Tissue Eng. Regen. Med. 2017, 14, 81–91. [Google Scholar] [CrossRef]
- Gullbrand, S.E.; Kim, D.H.; Bonnevie, E.; Ashinsky, B.G.; Smith, L.J.; Elliott, D.M.; Mauck, R.L.; Smith, H.E. Towards the scale up of tissue engineered intervertebral discs for clinical application. Acta Biomater. 2018, 70, 154–164. [Google Scholar] [CrossRef]
- D’Este, M.; Eglin, D.; Alini, M. Lessons to be learned and future directions for intervertebral disc biomaterials. Acta Biomater. 2018, 78, 13–22. [Google Scholar] [CrossRef]
- Gao, B.; Jiang, B.; Xing, W.; Xie, Z.; Luo, Z.; Zou, W. Discovery and Application of Postnatal Nucleus Pulposus Progenitors Essential for Intervertebral Disc Homeostasis and Degeneration. Adv. Sci. 2022, 9, 2104888. [Google Scholar] [CrossRef]
- Zhou, T.; Yang, X.; Chen, Z.; Yang, Y.; Wang, X.; Cao, X.; Chen, C.; Han, C.; Tian, H.; Qin, A.; et al. Prussian Blue Nanoparticles Stabilize SOD1 from Ubiquitination-Proteasome Degradation to Rescue Intervertebral Disc Degeneration. Adv. Sci. 2022, 9, 2105466. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Li, W.; Yang, S.; Yang, P.; Yan, Q.; Wang, S.; Lai, K.; Bai, X.; Wu, C.; Ding, W.; et al. Single-Cell Transcriptome Profiling Reveals Multicellular Ecosystem of Nucleus Pulposus during Degeneration Progression. Adv. Sci. 2022, 9, 2103631. [Google Scholar] [CrossRef] [PubMed]
- McHugh, J. Moving towards tissue-engineered disc replacement. Nat. Rev. Rheumatol. 2019, 15, 66. [Google Scholar] [CrossRef]
- Chen, W.; Chen, H.; Zheng, D.; Zhang, H.; Deng, L.; Cui, W.; Zhang, Y.; Santos, H.A.; Shen, H. Gene-Hydrogel Microenvironment Regulates Extracellular Matrix Metabolism Balance in Nucleus Pulposus. Adv. Sci. 2020, 7, 1902099. [Google Scholar] [CrossRef] [PubMed]
- Sharabi, M.; Wertheimer, S.; Wade, K.R.; Galbusera, F.; Benayahu, D.; Wilke, H.-J.; Haj-Ali, R. Towards intervertebral disc engineering: Bio-mimetics of form and function of the annulus fibrosus lamellae. J. Mech. Behav. Biomed. Mater. 2019, 94, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Sivan, S.S.; Roberts, S.; Urban, J.P.G.; Menage, J.; Bramhill, J.; Campbell, D.; Franklin, V.J.; Lydon, F.; Merkher, Y.; Maroudas, A.; et al. Injectable hydrogels with high fixed charge density and swelling pressure for nucleus pulposus repair: Biomimetic glycosaminoglycan analogues. Acta Biomater. 2014, 10, 1124–1133. [Google Scholar] [CrossRef]
- El-Husseiny, H.M.; Mady, E.A.; Hamabe, L.; Abugomaa, A.; Shimada, K.; Yoshida, T.; Tanaka, T.; Yokoi, A.; Elbadawy, M.; Tanaka, R. Smart/stimuli-responsive hydrogels: Cutting-edge platforms for tissue engineering and other biomedical applications. Mater. Today Bio 2022, 13, 100186. [Google Scholar] [CrossRef]
- Nerurkar, N.L.; Sen, S.; Huang, A.H.; Elliott, D.M.; Mauck, R.L. Engineered Disc-Like Angle-Ply Structures for Intervertebral Disc Replacement. Spine 2010, 35, 867–873. [Google Scholar] [CrossRef]
- Ashinsky, B.G.; Gullbrand, S.E.; Bonnevie, E.D.; Wang, C.; Kim, D.H.; Han, L.; Mauck, R.L.; Smith, H.E. Sacrificial Fibers Improve Matrix Distribution and Micromechanical Properties in a Tissue-Engineered Intervertebral Disc. Acta Biomater. 2020, 111, 232–241. [Google Scholar] [CrossRef]
- Martin, J.T.; Kim, D.H.; Milby, A.H.; Pfeifer, C.G.; Smith, L.J.; Elliott, D.M.; Smith, H.E.; Mauck, R.L. In vivo performance of an acellular disc-like angle ply structure (DAPS) for total disc replacement in a small animal model. J. Orthop. Res. 2017, 35, 23–31. [Google Scholar] [CrossRef]
- Wan, Y.; Feng, G.; Shen, F.H.; Laurencin, C.T.; Li, X. Biphasic scaffold for annulus fibrosus tissue regeneration. Biomaterials 2008, 29, 643–652. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.; Avegnon, K.L.M.; Holubeck, P.A.; Brown, D.; Karan, A.; Sharma, N.S.; John, J.V.; Weihs, S.; Ley, J.; Xie, J. Electrostatic flocking of salt-treated microfibers and nanofiber yarns for regenerative engineering. Mater. Today Bio 2021, 12, 100166. [Google Scholar] [CrossRef] [PubMed]
- Lazebnik, M.; Singh, M.; Glatt, P.; Friis, L.A.; Berkland, C.J.; Detamore, M.S. Biomimetic method for combining the nucleus pulposus and annulus fibrosus for intervertebral disc tissue engineering. J. Tissue Eng. Regen. Med. 2011, 5, e179–e187. [Google Scholar] [CrossRef] [PubMed]
- Nesti, L.J.; Li, W.-J.; Shanti, R.M.; Jiang, Y.J.; Jackson, W.; Freedman, B.A.; Kuklo, T.R.; Giuliani, J.R.; Tuan, R.S. Intervertebral disc tissue engineering using a novel hyaluronic acid–nanofibrous scaffold (HANFS) amalgam. Tissue Eng. Part A 2008, 14, 1527–1537. [Google Scholar] [CrossRef]
- Gullbrand, S.E.; Ashinsky, B.G.; Bonnevie, E.D.; Kim, D.H.; Engiles, J.B.; Smith, L.J.; Elliott, D.M.; Schaer, T.P.; Smith, H.E.; Mauck, R.L. Long-term mechanical function and integration of an implanted tissue-engineered intervertebral disc. Sci. Transl. Med. 2018, 10, eaau0670. [Google Scholar] [CrossRef]
- Choy, A.T.H.; Chan, B.P. A structurally and functionally biomimetic biphasic scaffold for intervertebral disc tissue engineering. PLoS ONE 2015, 10, e0131827. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, B.; Yang, Q.; Li, X.; Zhang, Y.; Ma, X.; Xia, Q.; Xu, H.; Zeng, C.; Zhang, Y. A novel natural ECM-derived biphasic scaffold for intervertebral disc tissue engineering. Mater. Lett. 2013, 105, 102–105. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Li, Y.; Zhou, Z.; Zhang, D.; Li, J.; Chu, H. Overall Structure Construction of an Intervertebral Disk Based on Highly Anisotropic Wood Hydrogel Composite Materials with Mechanical Matching and Buckling Buffering. ACS Appl. Mater. Interfaces 2021, 13, 15709–15719. [Google Scholar] [CrossRef]
- Sloan, S.R.; Galesso, D.; Secchieri, C.; Berlin, C.; Hartl, R.; Bonassar, L.J. Initial investigation of individual and combined annulus fibrosus and nucleus pulposus repair ex vivo. Acta Biomater. 2017, 59, 192–199. [Google Scholar] [CrossRef]
- Hudson, K.D.; Alimi, M.; Grunert, P.; Härtl, R.; Bonassar, L.J. Recent advances in biological therapies for disc degeneration: Tissue engineering of the annulus fibrosus, nucleus pulposus and whole intervertebral discs. Curr. Opin. Biotechnol. 2013, 24, 872–879. [Google Scholar] [CrossRef]
- Grunert, P.; Borde, B.H.; Towne, S.B.; Moriguchi, Y.; Hudson, K.D.; Bonassar, L.J.; Härtl, R. Riboflavin crosslinked high-density collagen gel for the repair of annular defects in intervertebral discs: An in vivo study. Acta Biomater. 2015, 26, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Borde, B.; Grunert, P.; Härtl, R.; Bonassar, L.J. Injectable, high-density collagen gels for annulus fibrosus repair: An in vitro rat tail model. J. Biomed. Mater. Res. Part A 2015, 103, 2571–2581. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Bonassar, L.J.; Grodzinsky, A.J. The role of cartilage streaming potential, fluid flow and pressure in the stimulation of chondrocyte biosynthesis during dynamic compression. J. Biomech. 1995, 28, 1055–1066. [Google Scholar] [CrossRef]
- Chan, L.K.; Leung, V.Y.; Tam, V.; Lu, W.W.; Sze, K.; Cheung, K.M. Decellularized bovine intervertebral disc as a natural scaffold for xenogenic cell studies. Acta Biomater. 2013, 9, 5262–5272. [Google Scholar] [CrossRef] [PubMed]
- Fiordalisi, M.; Silva, A.J.; Barbosa, M.; Gonçalves, R.; Caldeira, J. Decellularized Scaffolds for Intervertebral Disc Regeneration. Trends Biotechnol. 2020, 38, 947–951. [Google Scholar] [CrossRef]
- Xu, H.; Xu, B.; Yang, Q.; Li, X.; Ma, X.; Xia, Q.; Zhang, Y.; Zhang, C.; Wu, Y.; Zhang, Y. Comparison of decellularization protocols for preparing a decellularized porcine annulus fibrosus scaffold. PLoS ONE 2014, 9, e86723. [Google Scholar] [CrossRef]
- Wilke, H.-J.; Kettler, A.; Claes, L.E. Are sheep spines a valid biomechanical model for human spines? Spine 1997, 22, 2365–2374. [Google Scholar] [CrossRef]
- Vàzquez-Salat, N.; Salter, B.; Smets, G.; Houdebine, L.-M. The current state of GMO governance: Are we ready for GM animals? Biotechnol. Adv. 2012, 30, 1336–1343. [Google Scholar] [CrossRef]
- Moses, V. Biotechnology products and European consumers. Biotechnol. Adv. 1999, 17, 647–678. [Google Scholar] [CrossRef]
- Gloria, A.; Russo, T.; D’Amora, U.; Santin, M.; de Santis, R.; Ambrosio, L. Customised multiphasic nucleus/annulus scaffold for intervertebral disc repair/regeneration. Connect. Tissue Res. 2020, 61, 152–162. [Google Scholar] [CrossRef]
- Hassanzadeh, P.; Kharaziha, M.; Nikkhah, M.; Shin, S.R.; Jin, J.; He, S.; Sun, W.; Zhong, C.; Dokmeci, M.R.; Khademhosseini, A.; et al. Chitin nanofiber micropatterned flexible substrates for tissue engineering. J. Mater. Chem. B 2013, 1, 4217–4224. [Google Scholar] [CrossRef] [PubMed]
- Derrouiche, A.; Zaïri, F.; Zaïri, F. A chemo-mechanical model for osmo-inelastic effects in the annulus fibrosus. Biomech. Modeling Mechanobiol. 2019, 18, 1773–1790. [Google Scholar] [CrossRef] [PubMed]
- Ghezelbash, F.; Eskandari, A.H.; Shirazi-Adl, A.; Kazempour, M.; Tavakoli, J.; Baghani, M.; Costi, J.J. Modeling of human intervertebral disc annulus fibrosus with complex multi-fiber networks. Acta Biomater. 2021, 123, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Bashkuev, M.; Galbusera, F.; Wilke, H.-J.; Shirazi-Adl, A. Finite element study of human lumbar disc nucleus replacements. Comput. Methods Biomech. Biomed. Eng. 2014, 17, 1762–1776. [Google Scholar] [CrossRef]
- Klisch, S.M.; Lotz, J.C. Application of a fiber-reinforced continuum theory to multiple deformations of the annulus fibrosus. J. Biomech. 1999, 32, 1027–1036. [Google Scholar] [CrossRef]
- Elliott, D.M.; Setton, L.A. Anisotropic and inhomogeneous tensile behavior of the human anulus fibrosus: Experimental measurement and material model predictions. J. Biomech. Eng. 2001, 123, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, G.A.; Gasser, T.C. A viscoelastic model for fiber-reinforced composites at finite strains: Continuum basis, computational aspects and applications. Comput. Methods Appl. Mech. Eng. 2001, 190, 4379–4403. [Google Scholar] [CrossRef]
- Balzani, D.; Neff, P.; Schröder, J.; Holzapfel, G.A. A polyconvex framework for soft biological tissues. Adjustment to experimental data. Int. J. Solids Struct. 2006, 43, 6052–6070. [Google Scholar] [CrossRef]
- Zhou, M.; Werbner, B.; O’Connell, G.D. Fiber engagement accounts for geometry-dependent annulus fibrosus mechanics: A multiscale, Structure-Based Finite Element Study. J. Mech. Behav. Biomed. Mater. 2021, 115, 104292. [Google Scholar] [CrossRef]
- Sharabi, M.; Levi-Sasson, A.; Wolfson, R.; Wade, K.R.; Galbusera, F.; Benayahu, D.; Wilke, H.-J.; Haj-Ali, R. The Mechanical Role of the Radial Fiber Network Within the Annulus Fibrosus of the Lumbar Intervertebral Disc: A Finite Elements Study. J. Biomech. Eng. 2018, 141, 021006. [Google Scholar] [CrossRef]
- Noailly, J.; Lacroix, D.; Planell, J.A. Finite element study of a novel intervertebral disc substitute. Spine 2005, 30, 2257–2264. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.; Heuer, F.; Simon, U.; Kettler, A.; Rohlmann, A.; Claes, L.; Wilke, H.-J. Application of a new calibration method for a three-dimensional finite element model of a human lumbar annulus fibrosus. Clin. Biomech. 2006, 21, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Little, J.P.; Adam, C.J. Geometric sensitivity of patient-specific finite element models of the spine to variability in user-selected anatomical landmarks. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Nerurkar, N.L.; Mauck, R.L.; Elliott, D.M. Modeling interlamellar interactions in angle-ply biologic laminates for annulus fibrosus tissue engineering. Biomech. Modeling Mechanobiol. 2011, 10, 973–984. [Google Scholar] [CrossRef]
- Labus, K.M.; Han, S.K.; Hsieh, A.H.; Puttlitz, C.M. A computational model to describe the regional interlamellar shear of the annulus fibrosus. J. Biomech. Eng. 2014, 136, 051009. [Google Scholar] [CrossRef]
- Adam, C.; Rouch, P.; Skalli, W. Inter-lamellar shear resistance confers compressive stiffness in the intervertebral disc: An image-based modelling study on the bovine caudal disc. J. Biomech. 2015, 48, 4303–4308. [Google Scholar] [CrossRef]
- Kandil, K.; Zaïri, F.; Derrouiche, A.; Messager, T.; Zaïri, F. Interlamellar-induced time-dependent response of intervertebral disc annulus: A microstructure-based chemo-viscoelastic model. Acta Biomater. 2019, 100, 75–91. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Key Finding | Method | Ref. |
|---|---|---|---|
| Human IVD | Elastic fibers were found in the human IVD for the first time. | Transmission electron microscopy | [1] |
| Human IVD | The presence of elastic fibers at the AF and NP interface was reported. | Stained thin tissue (uranyl acetate and lead citrate) and transmission electron microscopy | [2] |
| Human fetal AF | Elastic fibers were found to be immature, consisting of a bundle of microfibrils with 20 nm in diameter. | Stained thin tissue (uranyl acetate and phosphotungstic acid) and electron microscopy | [3] |
| Human cervical IVD | The presence of a three-dimensional lattice of elastic fibers at the NP-endplate interface was reported. | Stained thin tissue (orcein, hematoxylin, and eosin) and transmission electron microscopy | [4] |
| Dog IVD | Elastic fibers were found in both the intra- and inter-lamellar regions of the AF. | Stained thin tissue (orcein, hematoxylin, and eosin) and transmission electron microscopy | [5] |
| Human lumbar IVD | Elastic fibers were found to occupy approximately 10% of the extracellular matrix of the AF (1985). | Stained thin tissue (orcein, hematoxylin, and eosin) and light microscopy | [6] |
| Human IVD | Elastin content was found to be less than 2% of the total IVD dry weight and elastic fiber in the intervertebral disk ere very sparse and irregular. | Immunohistochemistry and light microscopy for structural analysis and hot alkali method for elastin content measurement | [7] |
| Sample | Key Finding | Method | Ref. |
|---|---|---|---|
| Bovine IVD | Abundant and organized distribution of elastic fibers in different regions of IVD. | Stained thin tissue (orcein and immunostaining) and light microscopy | [8] |
| Human IVD | High-density and long radially oriented elastic fibers in the ILM and NP, respectively. | Stained thin tissue (orcein and immunostaining) and polarized light microscopy | [9] |
| Human lumbar IVD | Disorganized elastic fibers in the AF lamellae of scoliotic compared to normal IVDs. | Stained thin tissue (orcein and immunostaining) and light microscopy | [9,10] |
| Human lumbar IVD | Regional variation of elastic fibers (density and arrangement) in the AF. | Stained thin tissue (resorcin-fuchsin) and light microscopy | [11] |
| Human and bovine IVD | In the AF, the alignment of microfibrils and elastin fibres were similar to that of the collagen fibres. In the NP, microfibrils were mostly arranged around the cells and elastin fibers were rarely detected. | Stained thin tissue (dual immunostaining) and light microscopy | [12] |
| Bovine caudal IVD | Collagen compartments in the AF lamellae are enclosed by an integrated network of elastic fibers. | Immunohistology and confocal microscopy using unloaded or radially stretched tissues | [13] |
| Mechanical Properties | Strain Rate (%s−1) | Load Direction | ILM | ILM Elastic Network |
|---|---|---|---|---|
| Phase angle (°) | 0.1 | R | 13.5 ± 3.6 | 6.5 ± 3.4 |
| C | 12.5 ± 2.9 | 15.3 ± 6.7 | ||
| 1 | R | 11.5 ± 3.1 | 6 ± 2.3 | |
| C | 8.9 ± 2.7 | 10.5 ± 3.6 | ||
| 10 | R | 5.1 ± 3.2 | 1.6 ± 0.9 | |
| C | 2.5 ± 1.2 | 2.1 ± 0.8 | ||
| Extensibility (%) | 0.1 | R | 26.2 ± 6.3 | 30.5 ± 20.1 |
| C | 30.4 ± 11 | 31.6 ± 10 | ||
| 1 | R | 27.6 ± 4.2 | 27.7 ± 16 | |
| C | 22.4 ± 4.6 | 25.3± 12 | ||
| 10 | R | 27.4 ± 6.5 | 19.4 ± 9 | |
| C | 21.8 ± 5.1 | 20.4 ± 8.9 | ||
| Modulus (kPa) | 0.1 | R | 1.3 ± 0.5 | 0.8 ± 0.3 |
| C | 1.0 ± 0.4 | 0.5 ± 0.3 | ||
| 1 | R | 1.8 ± 0.6 | 0.8 ± 0.4 | |
| C | 1.1 ± 0.3 | 0.5 ± 0.3 | ||
| 10 | R | 1.8 ± 0.5 | 0.6 ± 0.3 | |
| C | 1.2 ± 0.5 | 0.6 ± 0.2 | ||
| Failure stress (kPa) | 10 | R | 287 ± 58 | 225 ±66 |
| C | 302 ± 46 | 149 ± 47 | ||
| Failure strain (%) | 10 | R | 256 ± 80 | 308 ± 78 |
| C | 343 ± 154 | 473 ± 256 |
| Sample | Key Finding | Ref. |
|---|---|---|
| Strategy 1—Structural analysis in the presence of ECM under micromechanical loading | ||
| Rat tail | Mechanical properties of elastic fibers in the ILM were likely associated with the propagation of circumferential tears in the AF | [14] |
| Ox tail | The ILM provides structural connectivity between the AF layers | [15] |
| Elastic fibers may contribute to the structural integrity of the AF lamella with a peak stress of 0.3 MPa | [16] | |
| Human | The annular delamination strength of the AF was measured | [17] |
| Rabbit | IVD degeneration affects the ILM (elastic fibers) and reduces the AF deamination strength. | [18] |
| Ovine | A high level of connectivity between the ILM and PB was found in the AF | [19] |
| A higher tensile stiffness was found for the ILM compared to that of the AF lamella (the contribution of the ILM’s elastic fibers to the structural integrity of the AF) | [20] | |
| The ILM exhibited weak connectivity in the posterior region of the AF, creating a pathway for IVD herniation while pressurizing the NP | [21,22] | |
| The NP microfibrils penetrate the endplate | [23] | |
| NP-endplate interface supporting an average tensile force of 20 N before failure | [24] | |
| The AF-NP interface tensile force in the radial direction was measured (7.7 N) | [25] | |
| The ILM was fully characterized mechanically in both shear and tension directions of loading | [26] | |
| The ILM mechanical properties, compared to the lamella, were significantly reduced during progression to herniation. | [27] | |
| Bovine | A well-organized network of elastic fibers to provide a mechanical linkage across the AF | [13] |
| A higher linear strain (3 times) for the ILM was found compared to the lamellae | [28] | |
| Strategy 2—Elastin enzymatic digestion and micromechanical loading | ||
| Human | The elimination of elastic fibers from the AF was shown to decrease modulus while increasing the AF extensibility. | [29] |
| The impact of elastin and collagen digestion on elastic and failure properties of the IVD was examined | [30] | |
| Bovine | The mechanical properties of the ILM were mostly controlled by collagen fibers and fibrillin rather than elastic fibers | [31] |
| Strategy 3—ECM alkali digestion and micromechanical loading | ||
| Ovine | The viscoelastic and failure mechanical properties of elastic fibers in the ILM were characterized. | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cyril, D.; Giugni, A.; Bangar, S.S.; Mirzaeipoueinak, M.; Shrivastav, D.; Sharabi, M.; Tipper, J.L.; Tavakoli, J. Elastic Fibers in the Intervertebral Disc: From Form to Function and toward Regeneration. Int. J. Mol. Sci. 2022, 23, 8931. https://doi.org/10.3390/ijms23168931
Cyril D, Giugni A, Bangar SS, Mirzaeipoueinak M, Shrivastav D, Sharabi M, Tipper JL, Tavakoli J. Elastic Fibers in the Intervertebral Disc: From Form to Function and toward Regeneration. International Journal of Molecular Sciences. 2022; 23(16):8931. https://doi.org/10.3390/ijms23168931
Chicago/Turabian StyleCyril, Divya, Amelia Giugni, Saie Sunil Bangar, Melika Mirzaeipoueinak, Dipika Shrivastav, Mirit Sharabi, Joanne L. Tipper, and Javad Tavakoli. 2022. "Elastic Fibers in the Intervertebral Disc: From Form to Function and toward Regeneration" International Journal of Molecular Sciences 23, no. 16: 8931. https://doi.org/10.3390/ijms23168931
APA StyleCyril, D., Giugni, A., Bangar, S. S., Mirzaeipoueinak, M., Shrivastav, D., Sharabi, M., Tipper, J. L., & Tavakoli, J. (2022). Elastic Fibers in the Intervertebral Disc: From Form to Function and toward Regeneration. International Journal of Molecular Sciences, 23(16), 8931. https://doi.org/10.3390/ijms23168931