
Lobelia chinensis Extract and Its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function through SPINK5/LEKTI Regulation


 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
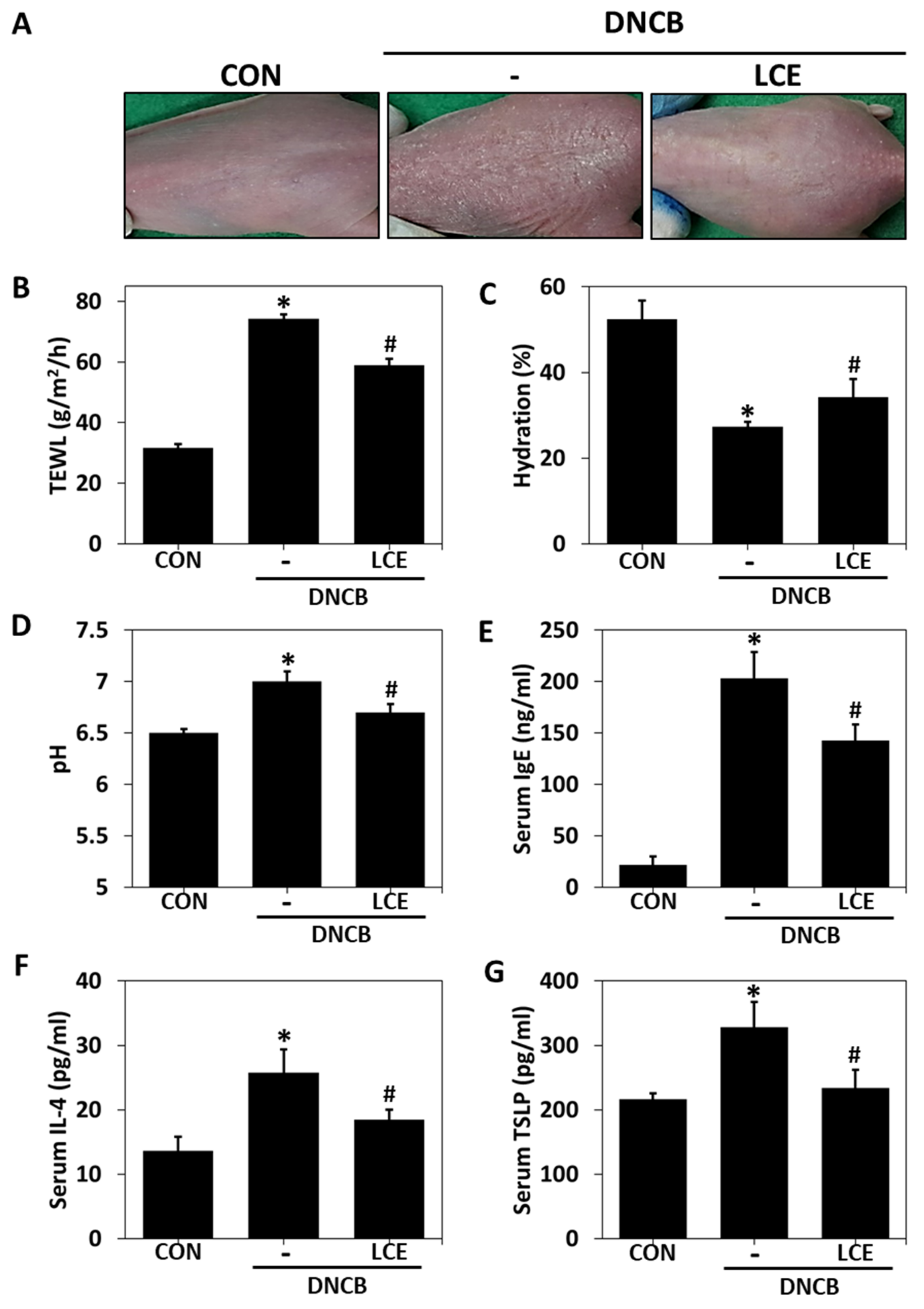
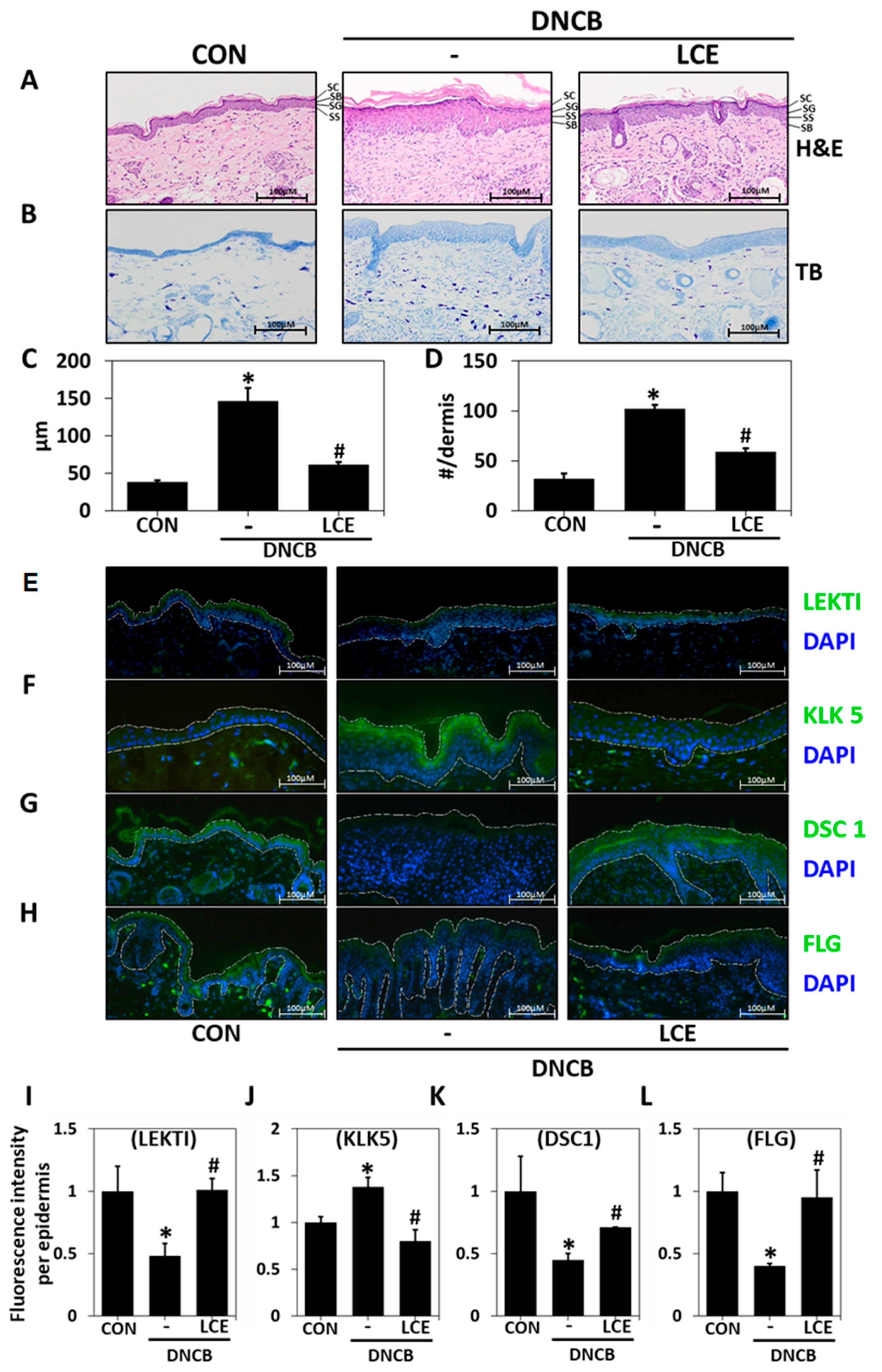
2.1. Effects of LCE on Impaired Skin Barrier Function and Serum Factors in the DNCB-Induced AD Model
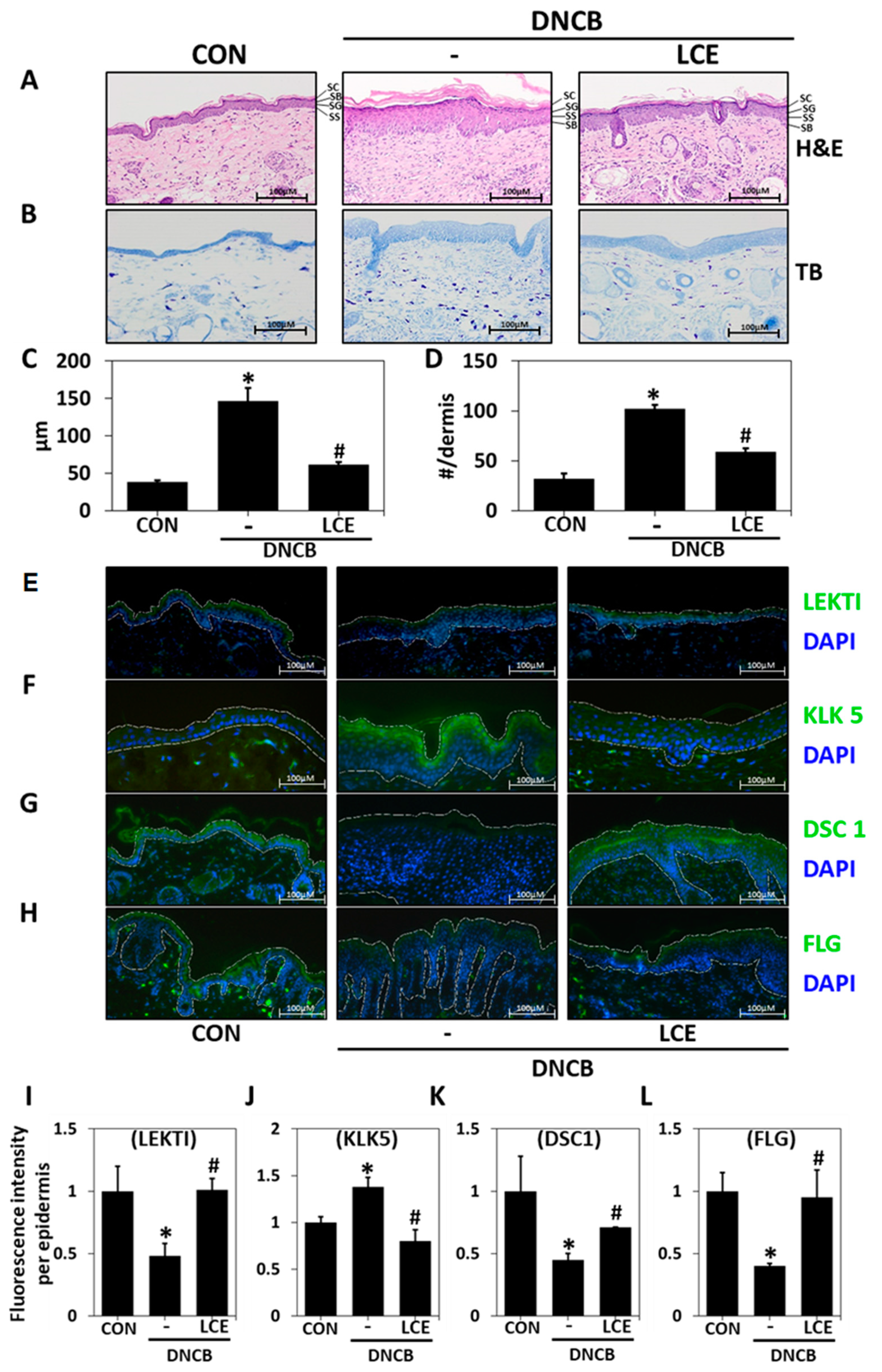
2.2. Effects of LCE on Histological Changes in DNCB-Treated Mice
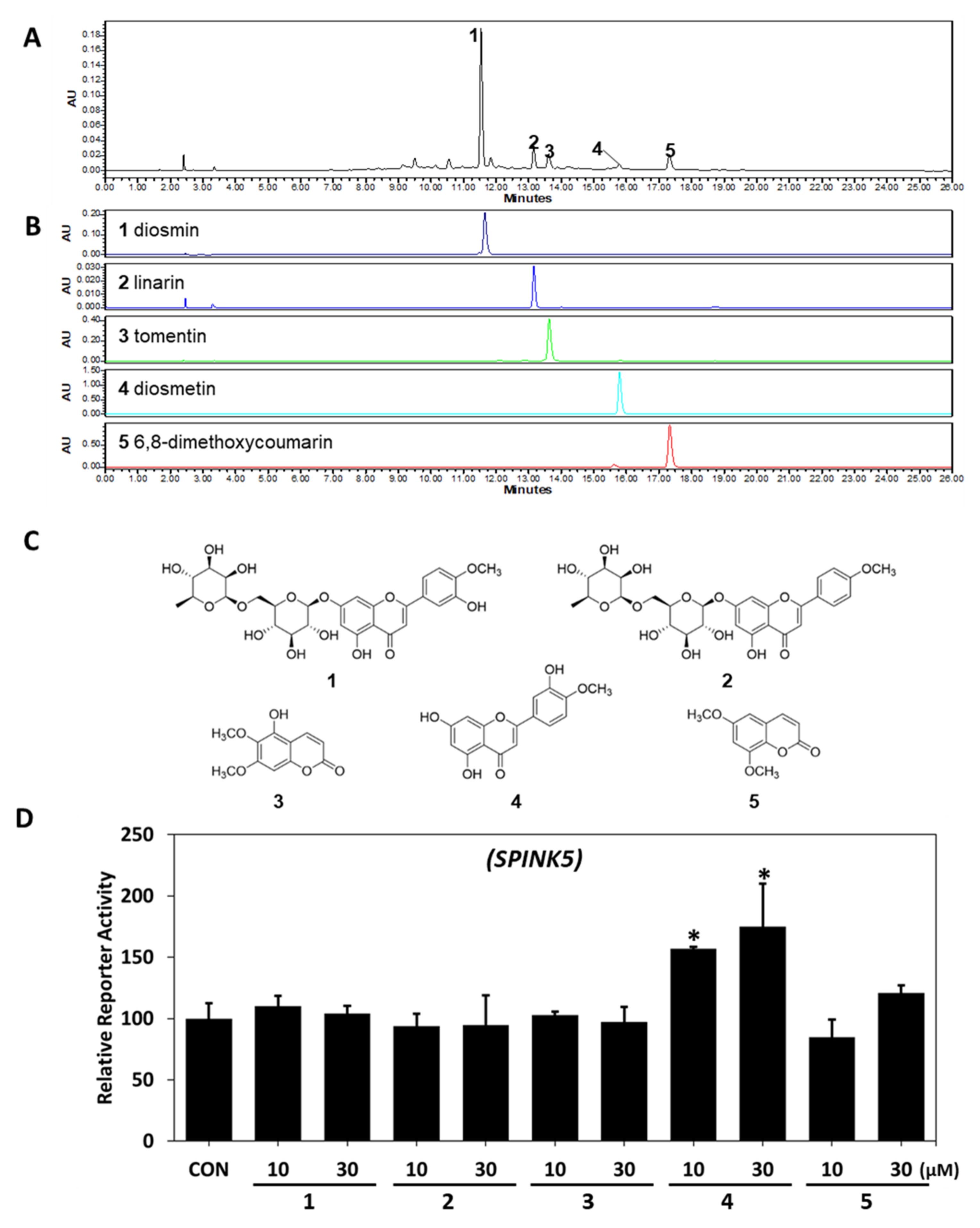
2.3. HPLC-PDA Analysis
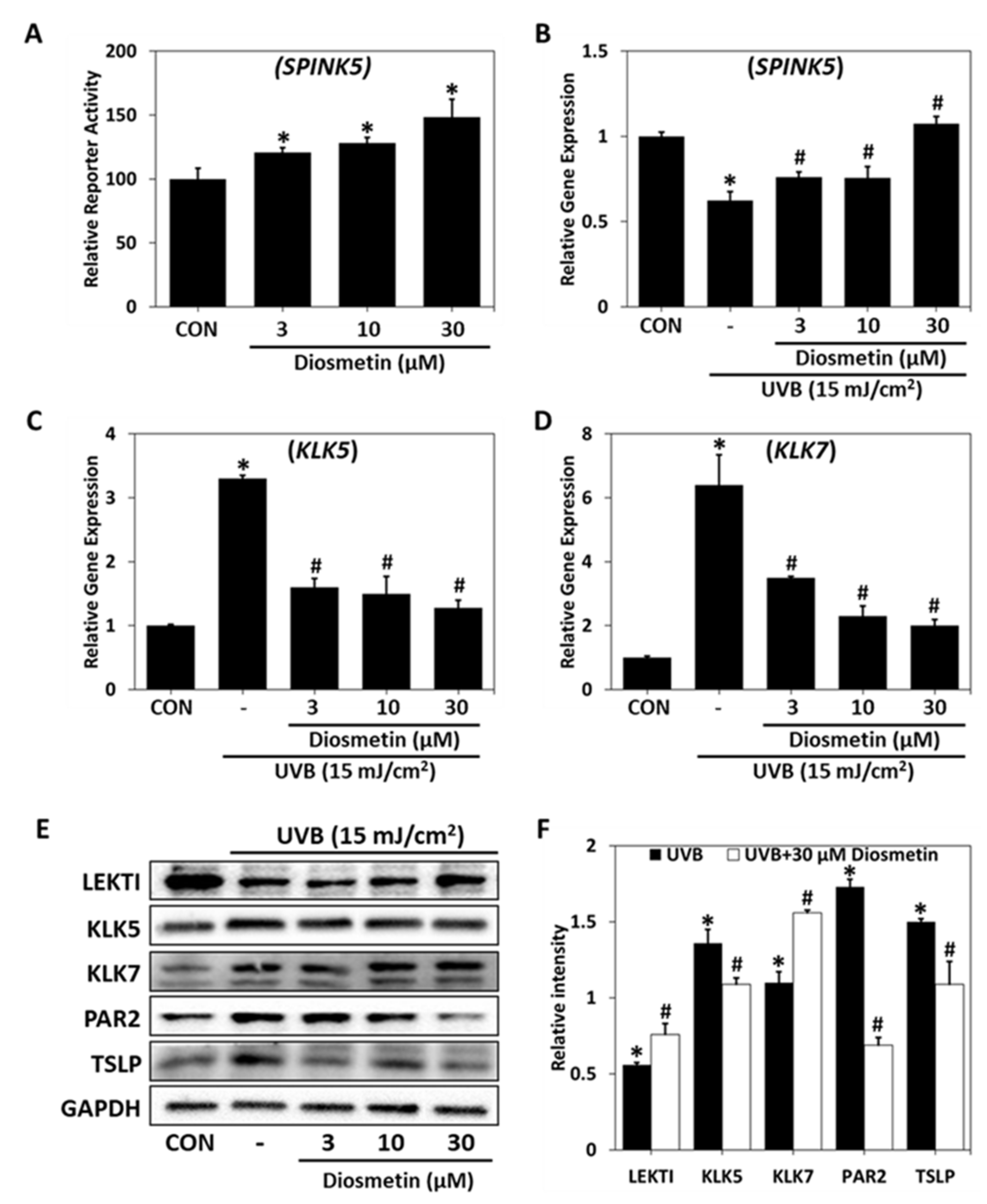
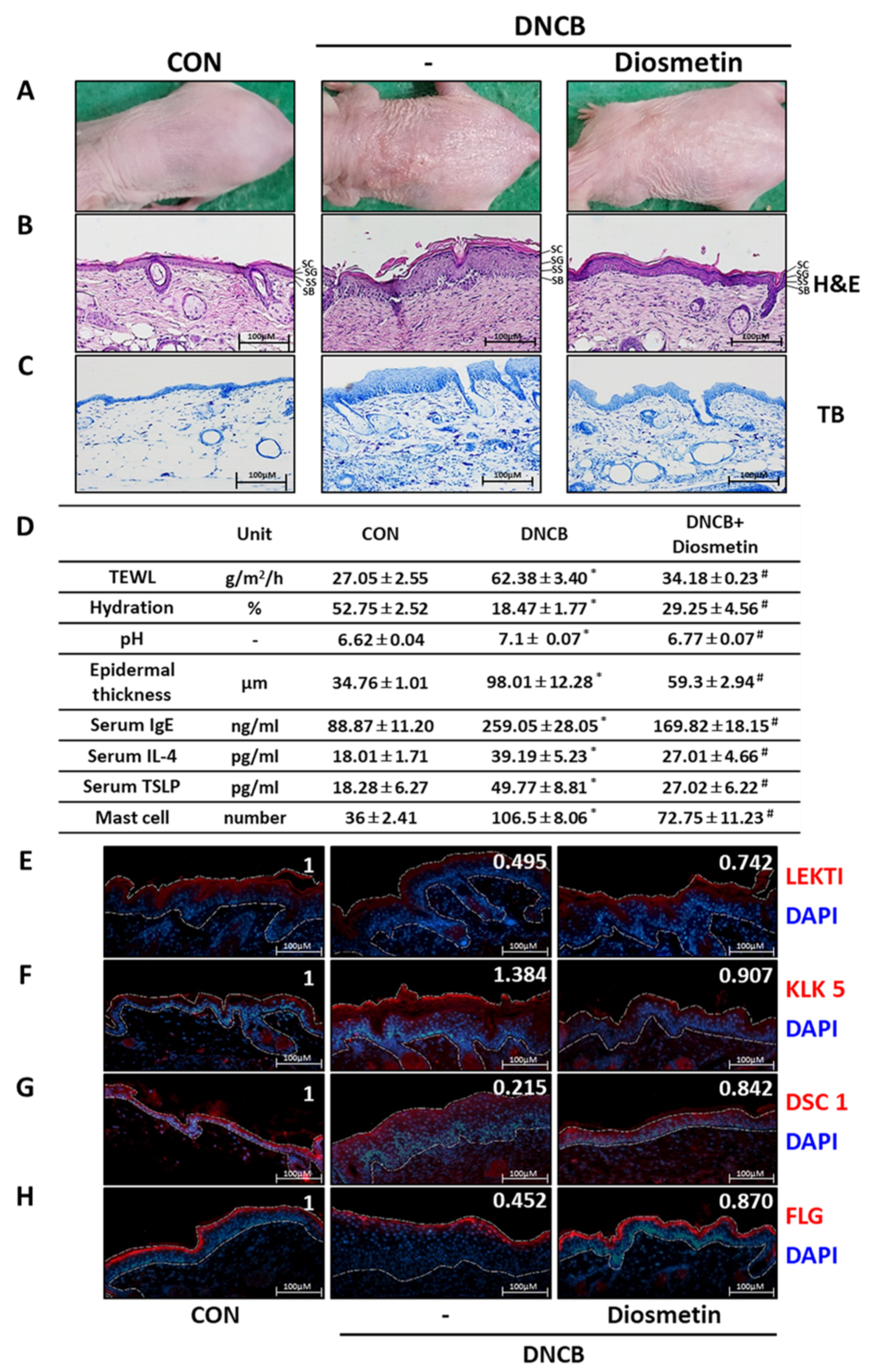
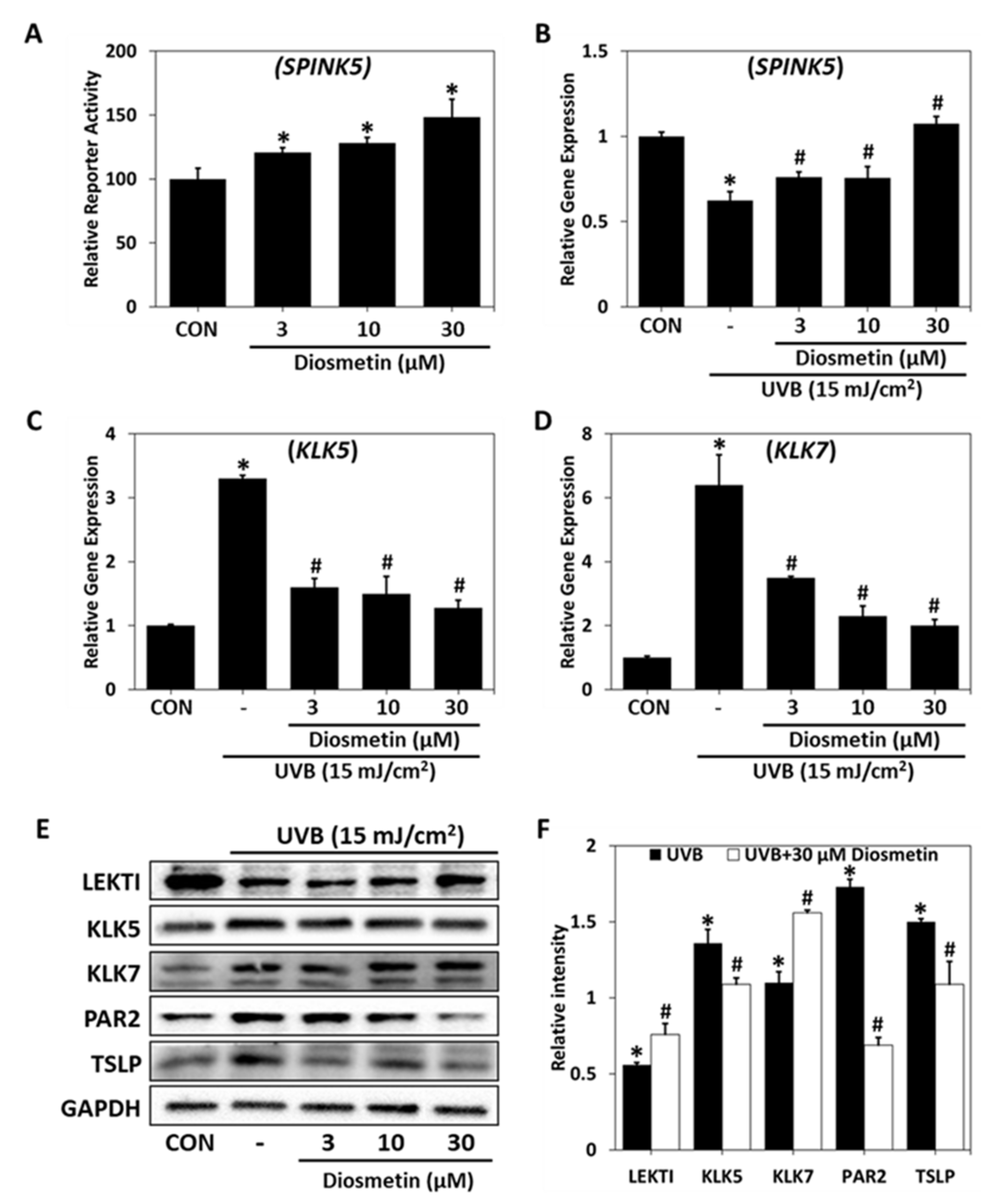
2.4. Regulatory Effect of Diosmetin on SPINK5/LEKTI Downstream Signaling
2.5. Effects of Diosmetin on the Skin Barrier in the DNCB Mouse Model
3. Discussion
4. Materials and Methods
4.1. Plant and Extraction
4.2. Chemicals
4.3. High-Performance Liquid Chromatography–Photodiode Array (HPLC–PDA) Conditions
4.4. Cell Culture
4.5. UVB Irradiation
4.6. Real-Time Quantitative PCR (Q-PCR)
4.7. Luciferase Reporter Gene Assay
4.8. Immunoblotting
4.9. Animal Experiments
4.10. Enzyme-Linked Immunosorbent Assay
4.11. Histological Examination
4.12. Immunohistochemistry
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ludriksone, L.; Bartels, N.G.; Kanti, V.; Blume-Peytavi, U.; Kottner, J. Skin barrier function in infancy: A systematic review. Arch. Dermatol. Res. 2014, 7, 591–599. [Google Scholar] [CrossRef]
- Tanaka, R.J.; Ono, M.; Harrington, H.A. Skin barrier homeostasis in atopic dermatitis: Feedback regulation of kallikrein activity. PLoS ONE 2011, 5, e19895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cork, M.J.; Danby, S.; Vasilopoulos, Y.; Moustafa, M.; MacGowan, A.; Varghese, J.; Duff, G.W.; Tazi-Ahnini, R.; Ward, S.J. Epidermal barrier dysfunction in atopic dermatitis. J. Investig. Dermatol. 2009, 8, 1892–1908. [Google Scholar] [CrossRef] [PubMed]
- Milstone, L.M. Epidermal desquamation. J. Dermatol. Sci. 2004, 3, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.P.; Bornslaeger, E.A.; Norvell, S.M.; Palka, H.L.; Green, K.J. Desmosomes: Intercellular adhesive junctions specialized for attachment of intermediate filaments. Int. Rev. Cytol. 1998, 185, 237–302. [Google Scholar]
- King, I.A.; Tabiowo, A.; Fryer, P.R. Evidence that major 78-44-kD concanavalin A-binding glycopolypeptides in pig epidermis arise from the degradation of desmosomal glycoproteins during terminal differentiation. J. Cell Biol. 1987, 6, 3053–3063. [Google Scholar] [CrossRef]
- Long, S.; Banks, J.; Watkinson, A.; Harding, C.; Rawlings, A.V. Desmocollin 1: A key marker for desmosome processing in the stratum corneum. J. Investig. Dermatol. 1996, 106, 872. [Google Scholar]
- O’Regan, G.M.; Sandilands, A.; McLean, W.I.; Irvine, A.D. Filaggrin in atopic dermatitis. J. Allergy Clin. Immunol. 2008, 4, 689–693. [Google Scholar] [CrossRef]
- Netherton, E.W. A unique case of trichorrhexis nodosa—Bamboo hairs. AMA Arch. Dermatol. 1958, 4, 483–487. [Google Scholar] [CrossRef]
- Judge, M.; Morgan, G.; Harper, J. A clinical and immunological study of Netherton’s syndrome. Br. J. Dermatol. 1994, 5, 615–621. [Google Scholar] [CrossRef]
- Smith, D.L.; Smith, J.G.; Wong, S.W.; De Shazo, R.D. Netherton’s syndrome. Br. J. Dermatol. 1995, 1, 153–154. [Google Scholar] [CrossRef]
- Hausser, I.; Anton-Lamprecht, I. Severe congenital generalized exfoliative erythroderma in newborns and infants: A possible sign of Netherton syndrome. Pediatric. Dermatol. 1996, 3, 183–199. [Google Scholar] [CrossRef]
- Suzuki, Y.; Nomura, J.; Koyama, J.; Horii, I. The role of proteases in stratum corneum: Involvement in stratum corneum desquamation. Arch. Dermatol. Res. 1994, 5, 249–253. [Google Scholar] [CrossRef]
- Komatsu, N.; Takata, M.; Otsuki, N.; Ohka, R.; Amano, O.; Takehara, K.; Saijoh, K. Elevated stratum corneum hydrolytic activity in Netherton syndrome suggests an inhibitory regulation of desquamation by SPINK5-derived peptides. J. Investig. Dermatol. 2002, 3, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Stoll, C.; Alembik, Y.; Tchomakov, D.; Messer, J.; Heid, E.; Boehm, N.; Calvas, P.; Hovnanian, A. Severe hypernatremic dehydration in an infant with Netherton syndrome. Genet. Couns. 2001, 3, 237–243. [Google Scholar]
- Kim, M.Y.; Lee, S.E.; Chang, J.Y.; Kim, S.C. Retinoid induces the degradation of corneodesmosomes and downregulation of corneodesmosomal cadherins: Implications on the mechanism of retinoid-induced desquamation. Ann. Dermatol. 2011, 4, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Deraison, C.; Bonnart, C.; Lopez, F.; Besson, C.; Robinson, R.; Jayakumar, A.; Wagberg, F.; Brattsand, M.; Hachem, J.P.; Leonardsson, G. LEKTI fragments specifically inhibit KLK5, KLK7, and KLK14 and control desquamation through a pH-dependent interaction. Mol. Biol. Cell 2007, 9, 3607–3619. [Google Scholar] [CrossRef] [Green Version]
- Briot, A.; Deraison, C.; Lacroix, M.; Bonnart, C.; Robin, A.; Besson, C.; Dubus, P.; Hovnanian, A. Kallikrein 5 induces atopic dermatitis–like lesions through PAR2-mediated thymic stromal lymphopoietin expression in Netherton syndrome. J. Exp. Med. 2009, 5, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, S.; Derian, C.K.; Vergnolle, N.; Bunnett, N.W.; Nawroth, R.; Schmelz, M.; Von Der Weid, P.-Y.; Buddenkotte, J.; Sunderkötter, C.; Metze, D. Proinflammatory role of proteinase-activated receptor-2 in humans and mice during cutaneous inflammation in vivo. FASEB J. 2003, 13, 1871–1885. [Google Scholar] [CrossRef]
- Yoo, J.; Omori, M.; Gyarmati, D.; Zhou, B.; Aye, T.; Brewer, A.; Comeau, M.R.; Campbell, D.J.; Ziegler, S.F. Spontaneous atopic dermatitis in mice expressing an inducible thymic stromal lymphopoietin transgene specifically in the skin. J. Exp. Med. 2005, 4, 541–549. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Wang, R.; Guo, F.; Yan, C. Two-dimensional liquid chromatography coupled with mass spectrometry for the analysis of Lobelia chinensis Lour. using an ESI/APCI multimode ion source. J. Sep. Sci. 2008, 13, 2388–2394. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Shi, R.; Liu, B.; Wang, Q.; Dai, Y. Studies on chemical components of Lobelia chinensis. J. Chin. Mater. Med. 2009, 3, 294–297. [Google Scholar]
- Li, S.; Lin, Z.; Jiang, H.; Tong, L.; Wang, H.; Chen, S. Rapid identification and assignation of the active ingredients in fufang banbianlian injection using HPLC-DAD-ESI-IT-TOF-MS. J. Chromatogr. Sci. 2016, 7, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Li, Y.; Huang, Y.; Zhao, C.; Cheung, H.Y. Chemical profiling of Lobelia chinensis with high-performance liquid chromatography/quadrupole time-of-flight mass spectrometry (HPLC/Q-TOF MS) reveals absence of lobeline in the herb. Molecules 2018, 12, 3258. [Google Scholar] [CrossRef] [Green Version]
- Li, K.C.; Ho, Y.L.; Huang, G.J.; Chang, Y.S. Anti-oxidative and anti-inflammatory effects of Lobelia chinensis in vitro and in vivo. Am. J. Chin. Med. 2015, 02, 269–287. [Google Scholar] [CrossRef]
- Liang, J.; Li, D. A study of pure Chinese medicine preparation controlling the growth of tumor in mice. Chin. J. Mod. Drug Appl. 2009, 20, 37–38. [Google Scholar]
- Kobayashi, T.; Nakata, T.; Kuzumaki, T. Effect of flavonoids on cell cycle progression in prostate cancer cells. Cancer Lett. 2002, 1, 17–23. [Google Scholar] [CrossRef]
- Shi, R.-X.; Ong, C.-N.; Shen, H.-M. Luteolin sensitizes tumor necrosis factor-α-induced apoptosis in human tumor cells. Oncogene 2004, 46, 7712–7721. [Google Scholar] [CrossRef] [Green Version]
- Park, N.J.; Bong, S.K.; Lee, S.; Jung, Y.; Jegal, H.; Kim, J.; Kim, S.K.; Kim, Y.K.; Kim, S.N. Compound K improves skin barrier function by increasing SPINK5 expression. J. Ginseng Res. 2020, 6, 799–807. [Google Scholar] [CrossRef]
- Antonov, D.; Schliemann, S.; Elsner, P. Methods for the assessment of barrier function. Ski. Barrier Funct. 2016, 49, 61–70. [Google Scholar]
- Jo, B.G.; Park, Y.H.; Kim, K.H.; Kim, S.N.; Yang, M.H. Simultaneous Determination of Four Marker Compounds in Lobelia chinensis Lour. Extract by HPLC-PDA. Appl. Sci. 2021, 24, 12080. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, J.K.; Choi, J.; Jang, H.; Seol, J.W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef]
- Park, S.A.; Bong, S.K.; Lee, J.W.; Park, N.J.; Choi, Y.S.; Kim, S.M.; Yang, M.H.; Kim, Y.K.; Kim, S.N. Diosmetin and its glycoside, diosmin, improve atopic dermatitis-like lesions in 2, 4-dinitrochlorobenzene-induced murine models. Biomol. Ther. 2020, 6, 542. [Google Scholar] [CrossRef]
- Borgono, C.A.; Michael, I.P.; Komatsu, N.; Jayakumar, A.; Kapadia, R.; Clayman, G.L.; Sotiropoulou, G.; Diamandis, E.P. A potential role for multiple tissue kallikrein serine proteases in epidermal desquamation. J. Biol. Chem. 2007, 6, 3640–3652. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G. Lipid abnormalities in atopic dermatitis. J. Am. Acad. Dermatol. 2001, 1, S29–S32. [Google Scholar] [CrossRef]
- Shirakawa, T.; Li, A.; Dubowitz, M.; Dekker, J.W.; Shaw, A.E.; Faux, J.A.; Ra, C.; Cookson, W.O.C.M.; Hopkin, J.M. Association between atopy and variants of the β subunit of the high–affinity immunoglobulin E receptor. Nat. Genet. 1994, 2, 125–130. [Google Scholar] [CrossRef]
- Sotiropoulou, G.; Pampalakis, G. Targeting the kallikrein-related peptidases for drug development. Trends Pharmacol. Sci. 2012, 12, 623–634. [Google Scholar] [CrossRef]
- Loden, M.; Olsson, H.; Axell, T.; Linde, Y.W. Friction, capacitance and transepidermal water loss (TEWL) in dry atopic and normal skin. Br. J. Dermatol. 1992, 2, 137–141. [Google Scholar] [CrossRef]
- Ishida-Yamamoto, A.; Deraison, C.; Bonnart, C.; Bitoun, E.; Robinson, R.; O’Brien, T.J.; Wakamatsu, K.; Ohtsubo, S.; Takahashi, H.; Hashimoto, Y. LEKTI is localized in lamellar granules, separated from KLK5 and KLK7, and is secreted in the extracellular spaces of the superficial stratum granulosum. J. Investig. Dermatol. 2005, 2, 360–366. [Google Scholar] [CrossRef]
- Le, N.A.; Katsuyama, M.; Demura, M.; Tanii, H.; Katsuyama, H.; Saijoh, K. Regulation of serine protease inhibitor Kazal type-5 (SPINK5) gene expression in the keratinocytes. Environ. Health Prev. Med. 2014, 4, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Corcuff, P.; Leveque, J. Corneocyte changes after acute UV irradiation and chronic solar exposure. Photodermatology 1988, 3, 110–115. [Google Scholar]
- Indra, A.K. Epidermal TSLP: A trigger factor for pathogenesis of atopic dermatitis. Expert Rev. Proteom. 2013, 4, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Kishibe, M. Physiological and pathological roles of kallikrein-related peptidases in the epidermis. J. Dermatol. Sci. 2019, 2, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Walley, A.J.; Chavanas, S.; Moffatt, M.F.; Esnouf, R.M.; Ubhi, B.; Lawrence, R.; Wong, K.; Abecasis, G.R.; Jones, E.Y.; Harper, J.I. Gene polymorphism in Netherton and common atopic disease. Nat. Genet. 2001, 2, 175–178. [Google Scholar] [CrossRef]
- Moffatt, M. SPINK5: A Gene for Atopic Dermatitis and Asthma. Clin. Exp. Allergy 2004, 34, 325–327. [Google Scholar] [CrossRef]
- Joseph-Nathan, P.; Dominguez, M.; Ortega, D.A. Shift reagent 1H NMR study of methoxycoumarins. J. Heterocycl. Chem. 1984, 4, 1141–1144. [Google Scholar] [CrossRef]
- Hammoda, H.M.; Ela, A.; El-Lakany, A.M.; El-Hanbali, O.; Zaki, C.S.; Ghazy, N.M. New constituents of Artemisia monosperma Del. Die Pharmazie 2008, 8, 611–614. [Google Scholar]
- Yan, Y.; Liu, X.; Gao, J.; Wu, Y.; Li, Y. Inhibition of TGF-β Signaling in Gliomas by the Flavonoid Diosmetin Isolated from Dracocephalum peregrinum L. Molecules 2020, 1, 192. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, N.-J.; Jo, B.-G.; Bong, S.-K.; Park, S.-a.; Lee, S.; Kim, Y.K.; Yang, M.H.; Kim, S.-N. Lobelia chinensis Extract and Its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function through SPINK5/LEKTI Regulation. Int. J. Mol. Sci. 2022, 23, 8687. https://doi.org/10.3390/ijms23158687
Park N-J, Jo B-G, Bong S-K, Park S-a, Lee S, Kim YK, Yang MH, Kim S-N. Lobelia chinensis Extract and Its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function through SPINK5/LEKTI Regulation. International Journal of Molecular Sciences. 2022; 23(15):8687. https://doi.org/10.3390/ijms23158687
Chicago/Turabian StylePark, No-June, Beom-Geun Jo, Sim-Kyu Bong, Sang-a Park, Sullim Lee, Yong Kee Kim, Min Hye Yang, and Su-Nam Kim. 2022. "Lobelia chinensis Extract and Its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function through SPINK5/LEKTI Regulation" International Journal of Molecular Sciences 23, no. 15: 8687. https://doi.org/10.3390/ijms23158687
APA StylePark, N.-J., Jo, B.-G., Bong, S.-K., Park, S.-a., Lee, S., Kim, Y. K., Yang, M. H., & Kim, S.-N. (2022). Lobelia chinensis Extract and Its Active Compound, Diosmetin, Improve Atopic Dermatitis by Reinforcing Skin Barrier Function through SPINK5/LEKTI Regulation. International Journal of Molecular Sciences, 23(15), 8687. https://doi.org/10.3390/ijms23158687