
Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight

 , , , , ,
, , , , ,
Abstract

1. Introduction
1.1. Palmitic Acid and Oleic Acid in Pregnancy
1.2. The Influence of Maternal Glycemia, BMI and Fetal Sex on Maternal and Placental Lipids
1.3. Aims and Hypotheses
2. Results
2.1. Placental Production of Fresh 13C-PA- and 13C-OA-Containing Lipids Quantified in Explant Tissue and in Conditioned Media
2.2. Explants from Female Placentas Incorporate More 13C Lipids than Those from Male Placentas
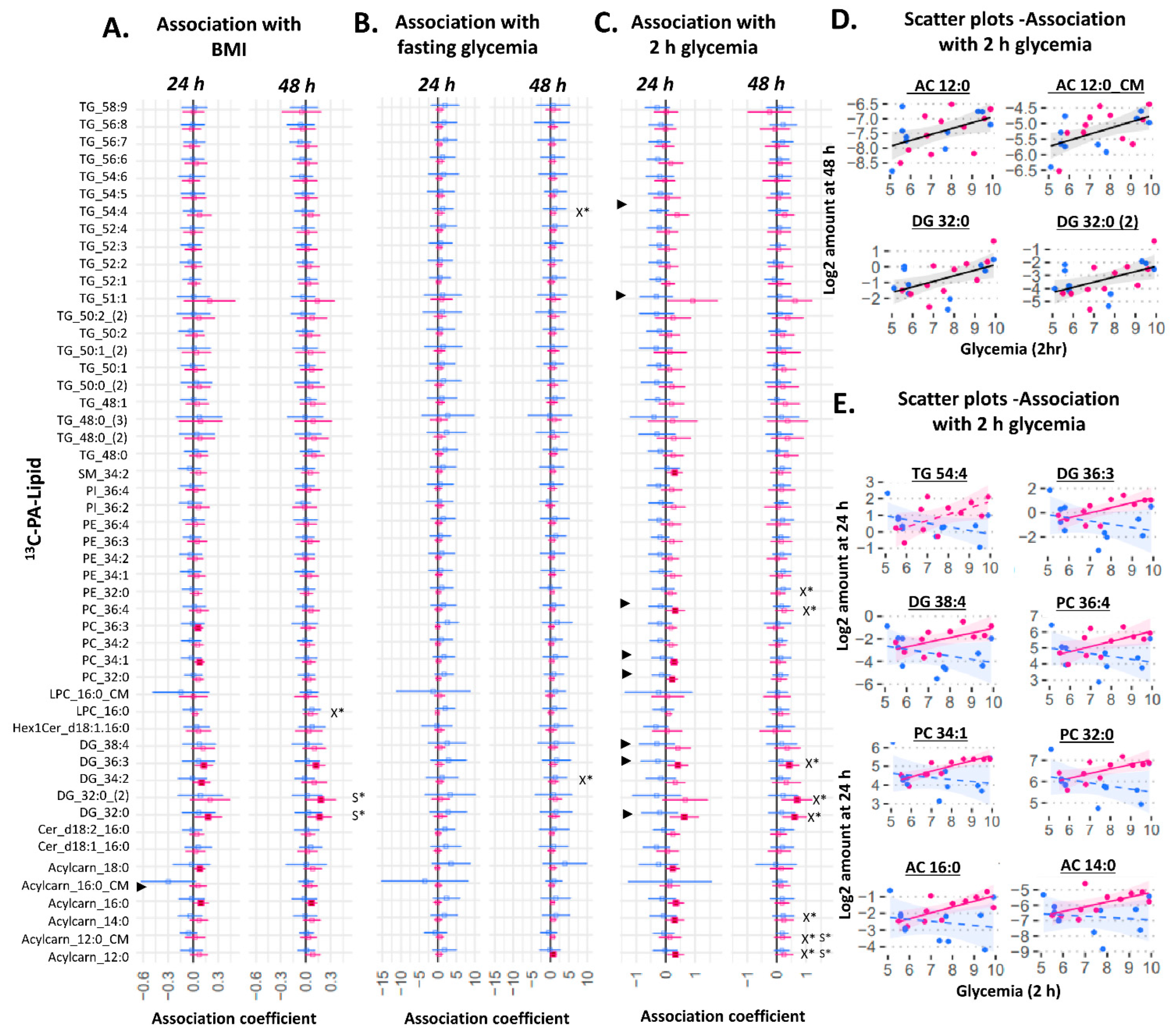
2.3. Placental Production of 13C Lipids Associated with Maternal Characteristics
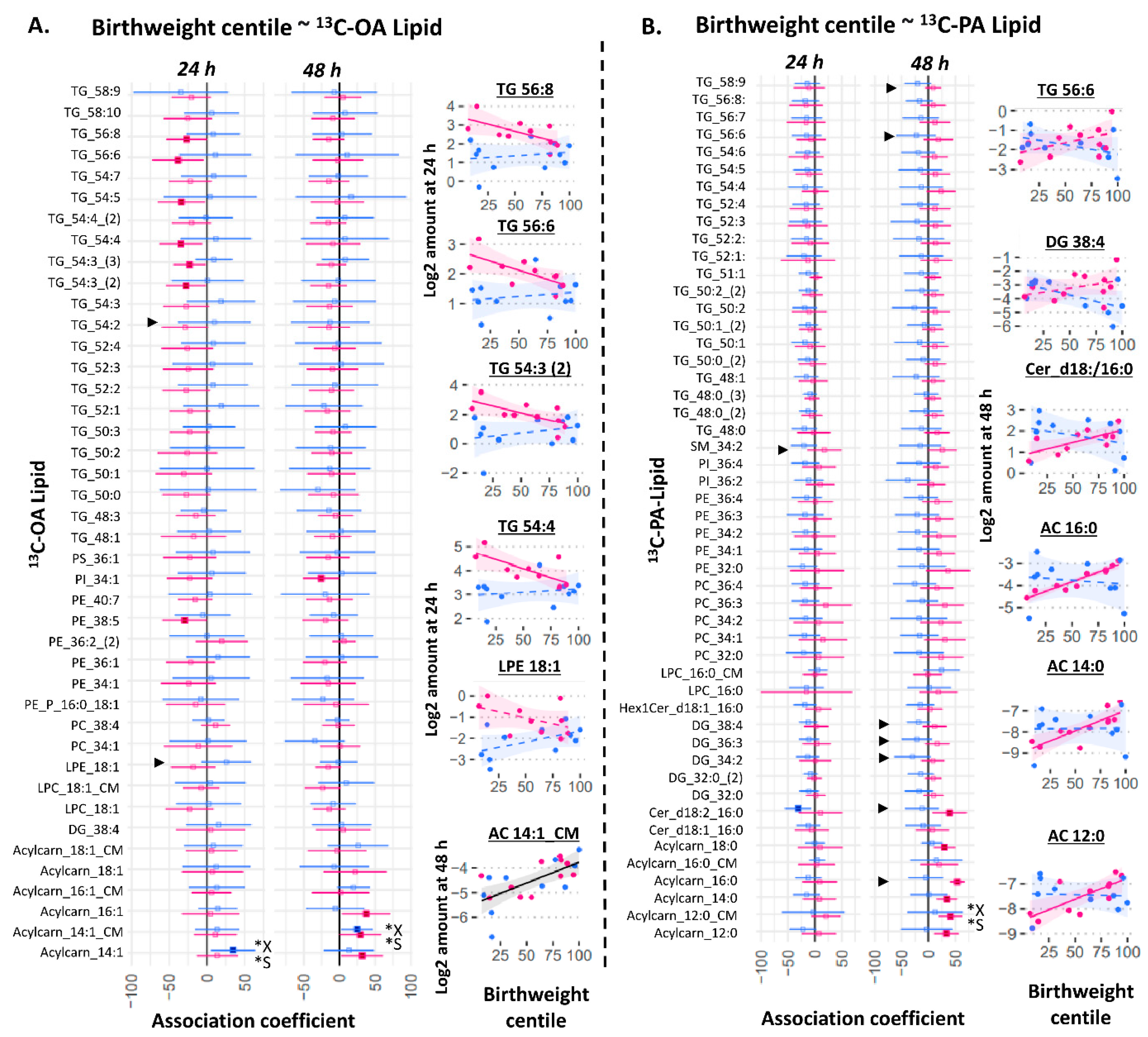
2.4. Birthweight Centile Is Associated with Placental 13C-OA Lipids and 13C-PA Lipids in Female Cases
2.5. The Relationship between mRNA Expression of Placental Fatty Acid Uptake Genes with Sex and Placental OA and PA Lipid Metabolism
3. Discussion
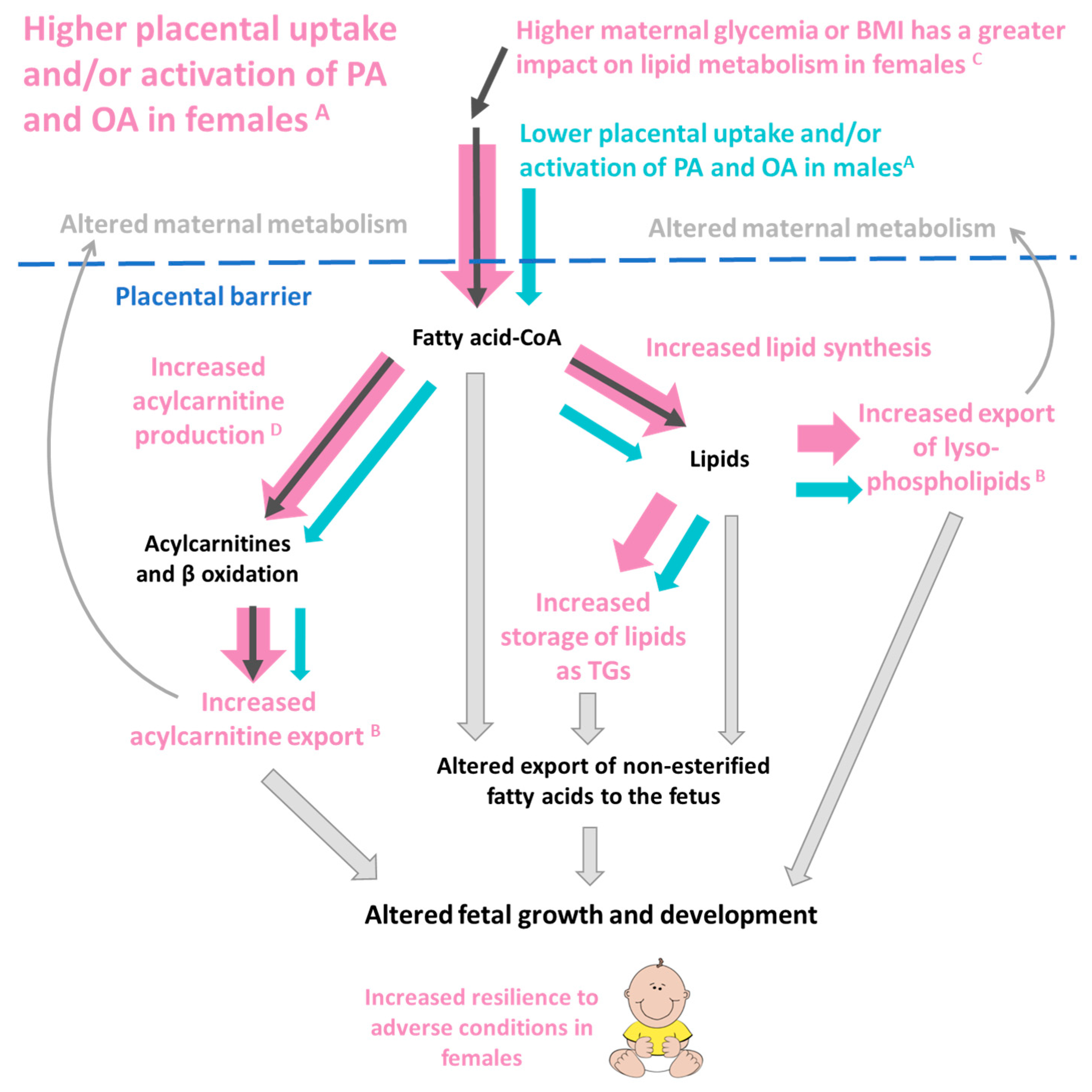
3.1. Main Findings
3.2. Sex Dependent Differences in Placental Lipid Metabolism
3.3. The Influence of Maternal Glycemia and BMI on Placental OA and PA Lipid Production
3.4. Placental Lipids and Birthweight
3.5. Strengths and Limitations
4. Materials and Methods
4.1. Placental Collection
4.2. Placenta Collection and Placental Explant Culture
4.3. RNA Extraction, cDNA Synthesis and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.4. Lipid Quantification by LCMS and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Voldner, N.; Frey Frøslie, K.; Godang, K.; Bollerslev, J.; Henriksen, T. Determinants of birth weight in boys and girls. Hum. Ontogenet. Int. J. Interdiscip. Dev. Res. 2009, 3, 7–12. [Google Scholar] [CrossRef]
- Roland, M.C.P.; Friis, C.M.; Godang, K.; Bollerslev, J.; Haugen, G.; Henriksen, T. Maternal factors associated with fetal growth and birthweight are independent determinants of placental weight and exhibit differential effects by fetal sex. PLoS ONE 2014, 9, e87303. [Google Scholar] [CrossRef] [PubMed]
- Group, H.S.C.R. Hyperglycaemia and Adverse Pregnancy Outcome (HAPO) Study: Associations with maternal body mass index. BJOG Int. J. Obstetr. Gynaecol. 2010, 117, 575–584. [Google Scholar]
- Clifton, V.L. Review: Sex and the Human Placenta: Mediating Differential Strategies of Fetal Growth and Survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.G.; Kajantie, E.; Osmond, C.; Thornburg, K.; Barker, D.J. Boys live dangerously in the womb. Am. J. Hum. Biol. 2010, 22, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Villa, P.M.; Laivuori, H.; Kajantie, E.; Kaaja, R. Free fatty acid profiles in preeclampsia. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 17–21. [Google Scholar] [CrossRef]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic acid: Physiological role, metabolism and nutritional implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef]
- Eastman, A.J.; Moore, R.E.; Townsend, S.D.; Gaddy, J.A.; Aronoff, D.M. The Influence of Obesity and Associated Fatty Acids on Placental Inflammation. Clin. Therap. 2021, 43, 265–278. [Google Scholar] [CrossRef]
- Herrera, E. Metabolic adaptations in pregnancy and their implications for the availability of substrates to the fetus. Eur. J. Clin. Nutr. 2000, 54, S47–S51. [Google Scholar] [CrossRef]
- Colvin, B.N.; Longtine, M.S.; Chen, B.; Costa, M.L.; Nelson, D.M. Oleate attenuates palmitate-induced endoplasmic reticulum stress and apoptosis in placental trophoblasts. Reproduction 2017, 153, 369–380. [Google Scholar] [CrossRef]
- Yang, X.; Haghiac, M.; Glazebrook, P.; Minium, J.; Catalano, P.M.; Hauguel-de Mouzon, S. Saturated fatty acids enhance TLR4 immune pathways in human trophoblasts. Hum. Reprod. 2015, 30, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Lager, S.; Jansson, T.; Powell, T.L. Differential regulation of placental amino acid transport by saturated and unsaturated fatty acids. Am. J. Physiol. Cell Physiol. 2014, 307, C738–C744. [Google Scholar] [CrossRef] [PubMed]
- Ricchi, M.; Odoardi, M.R.; Carulli, L.; Anzivino, C.; Ballestri, S.; Pinetti, A.; Fantoni, L.I.; Marra, F.; Bertolotti, M.; Banni, S. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J. Gastroenterol. Hepatol. 2009, 24, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Pathmaperuma, A.; Mana, P.; Cheung, S.; Kugathas, K.; Josiah, A.; Koina, M.; Broomfield, A.; Delghingaro-Augusto, V.; Ellwood, D.; Dahlstrom, J. Fatty acids alter glycerolipid metabolism and induce lipid droplet formation, syncytialisation and cytokine production in human trophoblasts with minimal glucose effect or interaction. Placenta 2010, 31, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Ortega-Senovilla, H. Implications of Lipids in Neonatal Body Weight and Fat Mass in Gestational Diabetic Mothers and Non-Diabetic Controls. Curr. Diabetes Rep. 2018, 18, 7. [Google Scholar] [CrossRef]
- Lewis, R.M.; Wadsack, C.; Desoye, G. Placental fatty acid transfer. Curr. Opin. Clin. Nutr. Metabol. Care 2018, 21, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Brass, E.; Hanson, E.; O’Tierney-Ginn, P.F. Placental oleic acid uptake is lower in male offspring of obese women. Placenta 2013, 34, 503–509. [Google Scholar] [CrossRef]
- Watkins, O.C.; Islam, M.O.; Selvam, P.; Pillai, R.A.; Cazenave-Gassiot, A.; Bendt, A.K.; Karnani, N.; Godfrey, K.M.; Lewis, R.M.; Wenk, M.R.; et al. Metabolism of 13C-Labeled Fatty Acids in Term Human Placental Explants by Liquid Chromatography–Mass Spectrometry. Endocrinology 2019, 160, 1394–1408. [Google Scholar] [CrossRef]
- Aris, I.M.; Soh, S.E.; Tint, M.T.; Liang, S.; Chinnadurai, A.; Saw, S.M.; Rajadurai, V.S.; Kwek, K.; Meaney, M.J.; Godfrey, K.M. Effect of maternal glycemia on neonatal adiposity in a multiethnic Asian birth cohort. J. Clin. Endocrinol. Metab. 2014, 99, 240–247. [Google Scholar] [CrossRef]
- Chen, L.-W.; Soh, S.E.; Tint, M.-T.; Loy, S.L.; Yap, F.; Tan, K.H.; Lee, Y.S.; Shek, L.P.-C.; Godfrey, K.M.; Gluckman, P.D. Combined analysis of gestational diabetes and maternal weight status from pre-pregnancy through post-delivery in future development of type 2 diabetes. Sci. Rep. 2021, 11, 5021. [Google Scholar] [CrossRef]
- Catalano, P.M.; Kirwan, J.P. Maternal factors that determine neonatal size and body fat. Curr. Diabetes Rep. 2001, 1, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.L.; Barner, K.; Madi, L.; Armstrong, M.; Manke, J.; Uhlson, C.; Jansson, T.; Ferchaud-Roucher, V. Sex-specific responses in placental fatty acid oxidation, esterification and transfer capacity to maternal obesity. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2021, 1866, 158861. [Google Scholar] [CrossRef] [PubMed]
- Mansell, T.; Vlahos, A.; Collier, F.; Ponsonby, A.-L.; Vuillermin, P.; Ellul, S.; Tang, M.L.K.; Burgner, D.; Saffery, R.; Vuillermin, P.; et al. The newborn metabolome: Associations with gestational diabetes, sex, gestation, birth mode, and birth weight. Pediatr. Res. 2021, 91, 1864–1873. [Google Scholar] [CrossRef]
- Goldenberg, J.R.; Wang, X.; Lewandowski, E.D. Acyl CoA synthetase-1 links facilitated long chain fatty acid uptake to intracellular metabolic trafficking differently in hearts of male versus female mice. J. Mol. Cell. Cardiol. 2016, 94, 26995156. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, D.; Zhou, S.; Kokkotou, E.; Berk, P.D. Sex differences in hepatic fatty acid uptake reflect a greater affinity of the transport system in females. Am. J. Physiol. Gastrointest. Liver Physiol. 1992, 263, G380–G385. [Google Scholar] [CrossRef]
- Ockner, R.K.; Burnett, D.A.; Lysenko, N.; Manning, J.A. Sex differences in long chain fatty acid utilization and fatty acid binding protein concentration in rat liver. J. Clin. Investig. 1979, 64, 172–181. [Google Scholar] [CrossRef]
- Kushlan, M.C.; Gollan, J.L.; Ma, W.L.; Ockner, R.K. Sex differences in hepatic uptake of long chain fatty acids in single-pass perfused rat liver. J. Lipid Res. 1981, 22, 431–436. [Google Scholar] [CrossRef]
- Ockner, R.K.; Lysenko, N.; Manning, J.A.; Monroe, S.E.; Burnett, D.A. Sex steroid modulation of fatty acid utilization and fatty acid binding protein concentration in rat liver. J. Clin. Investig. 1980, 65, 1013–1023. [Google Scholar] [CrossRef]
- Luxon, B.A.; Weisiger, R.A. Sex differences in intracellular fatty acid transport: Role of cytoplasmic binding proteins. Am. J. Physiol. Gastrointest. Liver Physiol. 1993, 265, G831–G841. [Google Scholar] [CrossRef]
- Stahlberg, N.; Rico-Bautista, E.; Fisher, R.M.; Wu, X.; Cheung, L.; Flores-Morales, A.; Tybring, G.; Norstedt, G.; Tollet-Egnell, P. Female-Predominant Expression of Fatty Acid Translocase/CD36 in Rat and Human Liver. Endocrinology 2004, 145, 1972–1979. [Google Scholar] [CrossRef]
- Xie, P.; Wang, X.-P.; Bu, Z.; Zou, X.-T. Differential expression of fatty acid transporters and fatty acid synthesis-related genes in crop tissues of male and female pigeons (Columba livia domestica) during incubation and chick rearing. Br. Poult. Sci. 2017, 58, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Yang, D.; Tang, B.; Yang, Y. Oleic acid induces smooth muscle foam cell formation and enhances atherosclerotic lesion development via CD36. Lipids Health Disease 2011, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Atshaves, B.P.; Storey, S.M.; Petrescu, A.; Greenberg, C.C.; Lyuksyutova, O.I.; Smith, R.; Schroeder, F. Expression of fatty acid binding proteins inhibits lipid accumulation and alters toxicity in L cell fibroblasts. Am. J. Physiol. Cell Physiol. 2002, 283, C688–C703. [Google Scholar] [CrossRef]
- Duttaroy, A.K. Transport of fatty acids across the human placenta: A review. Prog. Lipid Res. 2009, 48, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Baar, R.A.; Dingfelder, C.S.; Smith, L.A.; Bernlohr, D.A.; Wu, C.; Lange, A.J.; Parks, E.J. Investigation of in vivo fatty acid metabolism in AFABP/aP2−/− mice. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E187–E193. [Google Scholar] [CrossRef] [PubMed]
- Maatman, R.G.H.J.; van Moerkerk, H.T.B.; Nooren, I.M.A.; van Zoelen, E.J.J.; Veerkamp, J.H. Expression of human liver fatty acid-binding protein in Escherichia coli and comparative analysis of its binding characteristics with muscle fatty acid-binding protein. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1994, 1214, 8068722. [Google Scholar] [CrossRef]
- Thumser, A.E.; Voysey, J.E.; Wilton, D.C. The binding of lysophospholipids to rat liver fatty acid-binding protein and albumin. Biochem. J. 1994, 301, 801–806. [Google Scholar] [CrossRef]
- Prows, D.R.; Schroeder, F. Metallothionein-IIAPromoter Induction Alters Rat Intestinal Fatty Acid Binding Protein Expression, Fatty Acid Uptake, and Lipid Metabolism in Transfected L-Cells. Arch. Biochem. Biophys. 1997, 340, 135–143. [Google Scholar] [CrossRef]
- Coe, N.R.; Simpson, M.A.; Bernlohr, D.A. Targeted disruption of the adipocyte lipid-binding protein (aP2 protein) gene impairs fat cell lipolysis and increases cellular fatty acid levels. J. Lipid Res. 1999, 40, 967–972. [Google Scholar] [CrossRef]
- Larqué, E.; Pagán, A.; Prieto, M.T.; Blanco, J.E.; Gil-Sánchez, A.; Zornoza-Moreno, M.; Ruiz-Palacios, M.; Gázquez, A.; Demmelmair, H.; Parrilla, J.J. Placental fatty acid transfer: A key factor in fetal growth. Ann. Nutr. Metab. 2014, 64, 247–253. [Google Scholar] [CrossRef]
- Yang, Y.; Pan, Z.; Guo, F.; Wang, H.; Long, W.; Wang, H.; Yu, B. Placental metabolic profiling in gestational diabetes mellitus: An important role of fatty acids. J. Clin. Lab. Anal. 2021, 35, e24096. [Google Scholar] [CrossRef] [PubMed]
- Gázquez, A.; Prieto-Sánchez, M.T.; Blanco-Carnero, J.E.; Ruíz-Palacios, M.; Nieto, A.; van Harskamp, D.; Oosterink, J.E.; Schierbeek, H.; van Goudoever, J.B.; Demmelmair, H.; et al. Altered materno-fetal transfer of 13C-polyunsaturated fatty acids in obese pregnant women. Clin. Nutr. 2020, 39, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Shenai, J.P.; Borum, P.R.; Mohan, P.; Donlevy, S.C. Carnitine status at birth of newborn infants of varying gestation. Pediatr. Res. 1983, 17, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, N.; Hagerman, D.; Villee, C. Stimulation of fatty acid synthesis by oestradiol in vitro. Biochem. J. 1960, 76, 297. [Google Scholar] [CrossRef]
- Shambaugh, G., 3rd; Koehler, R.; Freinkel, N. Fetal fuels II: Contributions of selected carbon fuels to oxidative metabolism in rat conceptus. Am. J. Physiol. Endocrinol. Metab. 1977, 233, E457. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E.; Murray, A.J. Oxygen and placental development; parallels and differences with tumour biology. Placenta 2017, 56, 14–18. [Google Scholar] [CrossRef]
- Oey, N.; Den Boer, M.; Ruiter, J.; Wanders, R.; Duran, M.; Waterham, H.; Boer, K.; van der Post, J.; Wijburg, F. High activity of fatty acid oxidation enzymes in human placenta: Implications for fetal-maternal disease. J. Inherit. Metab. Disease 2003, 26, 385–392. [Google Scholar] [CrossRef]
- Shekhawat, P.; Bennett, M.J.; Sadovsky, Y.; Nelson, D.M.; Rakheja, D.; Strauss, A.W. Human placenta metabolizes fatty acids: Implications for fetal fatty acid oxidation disorders and maternal liver diseases. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E1098–E1105. [Google Scholar] [CrossRef]
- Jones, L.L.; McDonald, D.A.; Borum, P.R. Acylcarnitines: Role in brain. Progr. Lipid Res. 2010, 49, 61–75. [Google Scholar] [CrossRef]
- Arduini, A.; Mancinelli, G.; Radatti, G.L.; Dottori, S.; Molajoni, F.; Ramsay, R.R. Role of carnitine and carnitine palmitoyltransferase as integral components of the pathway for membrane phospholipid fatty acid turnover in intact human erythrocytes. J. Biol. Chem. 1992, 267, 12673–12681. [Google Scholar] [CrossRef]
- Arduini, A.; Denisova, N.; Virmani, A.; Avrova, N.; Federici, G.; Arrigoni-Martelli, E. Evidence for the Involvement of Carnitine-Dependent Long-Chain Acyltransferases in Neuronal Triglyceride and Phospholipid Fatty Acid Turnover. J. Neurochem. 1994, 62, 1530–1538. [Google Scholar] [CrossRef]
- Batchuluun, B.; Al Rijjal, D.; Prentice, K.J.; Eversley, J.A.; Burdett, E.; Mohan, H.; Bhattacharjee, A.; Gunderson, E.P.; Liu, Y.; Wheeler, M.B. Elevated Medium-Chain Acylcarnitines Are Associated With Gestational Diabetes Mellitus and Early Progression to Type 2 Diabetes and Induce Pancreatic β-Cell Dysfunction. Diabetes 2018, 67, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 23258903. [Google Scholar] [CrossRef]
- Sánchez-Pintos, P.; de Castro, M.-J.; Roca, I.; Rite, S.; López, M.; Couce, M.-L. Similarities between acylcarnitine profiles in large for gestational age newborns and obesity. Sci. Rep. 2017, 7, 16267. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef]
- Gokina, N.I.; Bonev, A.D.; Gokin, A.P.; Goloman, G. Role of impaired endothelial cell Ca2+ signaling in uteroplacental vascular dysfunction during diabetic rat pregnancy. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H935–H945. [Google Scholar] [CrossRef][Green Version]
- Tóth, M. Attenuation of diacylglycerol signal in the primordial human placenta: Role of phosphatidylcholine formation. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1993, 1210, 105–112. [Google Scholar] [CrossRef]
- Eichmann, T.O.; Lass, A. DAG tales: The multiple faces of diacylglycerol—Stereochemistry, metabolism, and signaling. Cell. Mol. Life Sci. 2015, 72, 3931–3952. [Google Scholar] [CrossRef] [PubMed]
- Drzazga, A.; Sowińska, A.; Koziołkiewicz, M. Lysophosphatidylcholine and lysophosphatidylinosiol—Novel promissing signaling molecules and their possible therapeutic activity. Acta Poloniae Pharm. 2014, 71, 887–899. [Google Scholar]
- Iwashita, M.; Watanabe, M.; Setoyama, T.; Mimuro, T.; Nakayamab, S.; Adachi, T.; Takeda, Y.; Sakamoto, S. Effects of diacylglycerol and gonadotropin-releasing hormone on human chorionic gonadotropin release by cultured trophoblast cells. Placenta 1992, 13, 213–221. [Google Scholar] [CrossRef]
- Harman, I.; Zeitler, P.; Ganong, B.; Bell, R.M.; Handwerger, S. Sn-1,2-Diacylglycerols and Phorbol Esters Stimulate the Synthesis and Release of Human Placental Lactogen from Placental Cells: A Role for Protein Kinase C*. Endocrinology 1986, 119, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Sane, A.; Harman, I.; Quarfordt, S.; Costello, A.; Handwergera, S. Characterization of placental lactogen release from perifused human trophoblast cells. Placenta 1988, 9, 129–138. [Google Scholar] [CrossRef]
- Kato, H.; Kato, M.; Kasugai, M.; Mizutani, S.; Ninagawa, T.; Tomoda, Y. Sn-1, 2-diacylglycerols and phorbol ester stimulate the production of progesterone from the human placenta. Eur. J. Endocrinol. 1989, 121, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, S.; Yinon, Y.; Xu, J.; Ermini, L.; Sallais, J.; Tagliaferro, A.; Todros, T.; Post, M.; Caniggia, I. Aberrant TGFβ signalling contributes to dysregulation of sphingolipid metabolism in intrauterine growth restriction. J. Clin. Endocrinol. Metab. 2015, 100, E986–E996. [Google Scholar] [CrossRef] [PubMed]
- Guenther, G.G.; Peralta, E.R.; Rosales, K.R.; Wong, S.Y.; Siskind, L.J.; Edinger, A.L. Ceramide starves cells to death by downregulating nutrient transporter proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 17402–17407. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Griffiths, H.R. Ceramides reduce CD36 cell surface expression and oxidised LDL uptake by monocytes and macrophages. Arch. Biochem. Biophys. 2006, 450, 89–99. [Google Scholar] [CrossRef]
- Hyde, R.; Hajduch, E.; Powell, D.J.; Taylor, P.M.; Hundal, H.S. Ceramide down-regulates System A amino acid transport and protein synthesis in rat skeletal muscle cells. FASEB J. 2005, 19, 15611152. [Google Scholar] [CrossRef]
- Singh, A.T.; Dharmarajan, A.; Aye, I.L.M.H.; Keelan, J.A. Ceramide biosynthesis and metabolism in trophoblast syncytialization. Mol. Cell. Endocrinol. 2012, 362, 48–59. [Google Scholar] [CrossRef]
- Signorelli, P.; Avagliano, L.; Reforgiato, M.R.; Toppi, N.; Casas, J.; Fabriàs, G.; Marconi, A.M.; Ghidoni, R.; Caretti, A. De novo ceramide synthesis is involved in acute inflammation during labor. Biol. Chem. 2016, 397, 147–155. [Google Scholar] [CrossRef]
- Jansson, N.; Rosario, F.J.; Gaccioli, F.; Lager, S.; Jones, H.N.; Roos, S.; Jansson, T.; Powell, T.L. Activation of Placental mTOR Signaling and Amino Acid Transporters in Obese Women Giving Birth to Large Babies. J. Clin. Endocrinol. Metab. 2013, 98, 105–113. [Google Scholar] [CrossRef]
- Chen, K.-Y.; Lin, S.-Y.; Lee, C.-N.; Wu, H.-T.; Kuo, C.-H.; Kuo, H.-C.; Chuang, C.-C.; Kuo, C.-H.; Chen, S.-C.; Fan, K.-C. Maternal Plasma Lipids During Pregnancy, Insulin-like Growth Factor-1 and Excess Foetal Growth. J. Clin. Endocrinol. Metab. 2021, 106, e3461–e3472. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Kawabata, T.; Kagawa, Y.; Shoji, K.; Kimura, F.; Miyazawa, T.; Tatsuta, N.; Arima, T.; Yaegashi, N.; Nakai, K. Associations of umbilical cord fatty acid profiles and desaturase enzyme indices with birth weight for gestational age in Japanese infants. Prostaglandins Leukot. Essent. Fat. Acids 2021, 165, 102233. [Google Scholar] [CrossRef] [PubMed]
- Giannacopoulou, C.; Evangeliou, A.; Matalliotakis, I.; Relakis, K.; Sbirakis, N.; Hatzidaki, E.; Koumandakis, E. Effects of gestation age and of birth weight in the concentration of carnitine in the umblical plasma. Clin. Exp. Obstet. Gynaecol. 1998, 25, 42–45. [Google Scholar]
- El-Wahed, M.A.; El-Farghali, O.; ElAbd, H.; El-Desouky, E.; Hassan, S. Metabolic derangements in IUGR neonates detected at birth using UPLC-MS. Egypt. J. Med. Hum. Genet. 2017, 18, 281–287. [Google Scholar] [CrossRef]
- Sánchez-Pintos, P.; Perez-Munuzuri, A.; Cocho, J.Á.; Fernández-Lorenzo, J.R.; Fraga, J.M.; Couce, M.L. Evaluation of carnitine deficit in very low birth weight preterm newborns small for their gestational age. J. Mater. Fetal Neonat. Med. 2016, 29, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Sommerfeld, E.; Penn, D.; Sodha, R.J.; Prögler, M.; Novak, M.; Schneider, H. Transfer and metabolism of carnitine and carnitine esters in the in vitro perfused human placenta. Pediatr. Res. 1985, 19, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.; Monkus, E.F.; Chung, D.; Buch, M. Carnitine in the perinatal metabolism of lipids I. Relationship between maternal and fetal plasma levels of carnitine and acylcarnitines. Pediatrics 1981, 67, 95–100. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, M.; González, R.S.; Maldonado, J.; López-Alarcón, M.; Bernabe-García, M. The effect of gestational age on expression of genes involved in uptake, trafficking and synthesis of fatty acids in the rat placenta. Gene 2016, 591, 403–410. [Google Scholar] [CrossRef]
- Watkins, O.C.; Selvam, P.; Pillai, R.A.; Cracknell-Hazra, V.; Yong, H.E.J.; Sharma, N.; Cazenave-Gassiot, A.; Bendt, A.K.; Godfrey, K.M.; Lewis, R.M.; et al. Placental 13C-DHA metabolism and relationship with maternal BMI, glycemia and birthweight. Mol. Med. 2021, 27, 84. [Google Scholar] [CrossRef]
- Benassayag, C.; Mignot, T.; Haourigui, M.; Civel, C.; Hassid, J.; Carbonne, B.; Nunez, E.; Ferre, F. High polyunsaturated fatty acid, thromboxane A2, and alpha-fetoprotein concentrations at the human feto-maternal interface. J. Lipid Res. 1997, 38, 276–286. [Google Scholar] [CrossRef]
- Abdelmagid, S.A.; Clarke, S.E.; Nielsen, D.E.; Badawi, A.; El-Sohemy, A.; Mutch, D.M.; Ma, D.W. Comprehensive profiling of plasma fatty acid concentrations in young healthy Canadian adults. PLoS ONE 2015, 10, e0116195. [Google Scholar] [CrossRef] [PubMed]
- Pillai, R.A.; Islam, M.O.; Selvam, P.; Sharma, N.; Chu, A.H.; Watkins, O.C.; Godfrey, K.M.; Lewis, R.M.; Chan, S.Y. Placental Inositol Reduced in Gestational Diabetes as Glucose alters Inositol Transporters and IMPA1 enzyme expression. J. Clin. Endocrinol. Metab. 2020, 106, e875–e890. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Total | Male | Female |
|---|---|---|---|
| n | 22 | 11 | 11 |
| Maternal Age (years) | 33.2 (2.7) | 33.7 (2.4) | 32.6 (2.9) |
| Chinese: Indian ethnicity | 13:9 | 6:5 | 7:4 |
| Maternal BMI in first trimester (kg/m2) | 25.6 (5.0) | 25.4 (5.4) | 25.8 (4.7) |
| GDM # (%) | 50% | 45% | 54% |
| Fasting glycemia (mmol/L) # | 4.5 (0.4) | 4.4 (0.2) | 4.6 (0.5) |
| 2-h glycemia (mmol/L) # | 7.5 (1.6) | 7.2 (1.8) | 7.7 (1.5) |
| Gestational age at delivery (days) | 271.2 (5.3) | 270.5 (5.4) | 271.9 (5.4) |
| Birthweight (g) | 3215.7 (370.5) | 3205 (423.3) | 3226.4 (329.9) |
| Birthweight Centile (%) ^ | 57 (33.3) | 54.9 (37.3) | 59.2 (30.5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watkins, O.C.; Yong, H.E.J.; Mah, T.K.L.; Cracknell-Hazra, V.K.B.; Pillai, R.A.; Selvam, P.; Sharma, N.; Cazenave-Gassiot, A.; Bendt, A.K.; Godfrey, K.M.; et al. Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight. Int. J. Mol. Sci. 2022, 23, 8685. https://doi.org/10.3390/ijms23158685
Watkins OC, Yong HEJ, Mah TKL, Cracknell-Hazra VKB, Pillai RA, Selvam P, Sharma N, Cazenave-Gassiot A, Bendt AK, Godfrey KM, et al. Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight. International Journal of Molecular Sciences. 2022; 23(15):8685. https://doi.org/10.3390/ijms23158685
Chicago/Turabian StyleWatkins, Oliver C., Hannah E. J. Yong, Tania Ken Lin Mah, Victoria K. B. Cracknell-Hazra, Reshma Appukuttan Pillai, Preben Selvam, Neha Sharma, Amaury Cazenave-Gassiot, Anne K. Bendt, Keith M. Godfrey, and et al. 2022. "Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight" International Journal of Molecular Sciences 23, no. 15: 8685. https://doi.org/10.3390/ijms23158685
APA StyleWatkins, O. C., Yong, H. E. J., Mah, T. K. L., Cracknell-Hazra, V. K. B., Pillai, R. A., Selvam, P., Sharma, N., Cazenave-Gassiot, A., Bendt, A. K., Godfrey, K. M., Lewis, R. M., Wenk, M. R., & Chan, S.-Y. (2022). Sex-Dependent Regulation of Placental Oleic Acid and Palmitic Acid Metabolism by Maternal Glycemia and Associations with Birthweight. International Journal of Molecular Sciences, 23(15), 8685. https://doi.org/10.3390/ijms23158685