
Lipolysis-Stimulated Lipoprotein Receptor Acts as Sensor to Regulate ApoE Release in Astrocytes

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
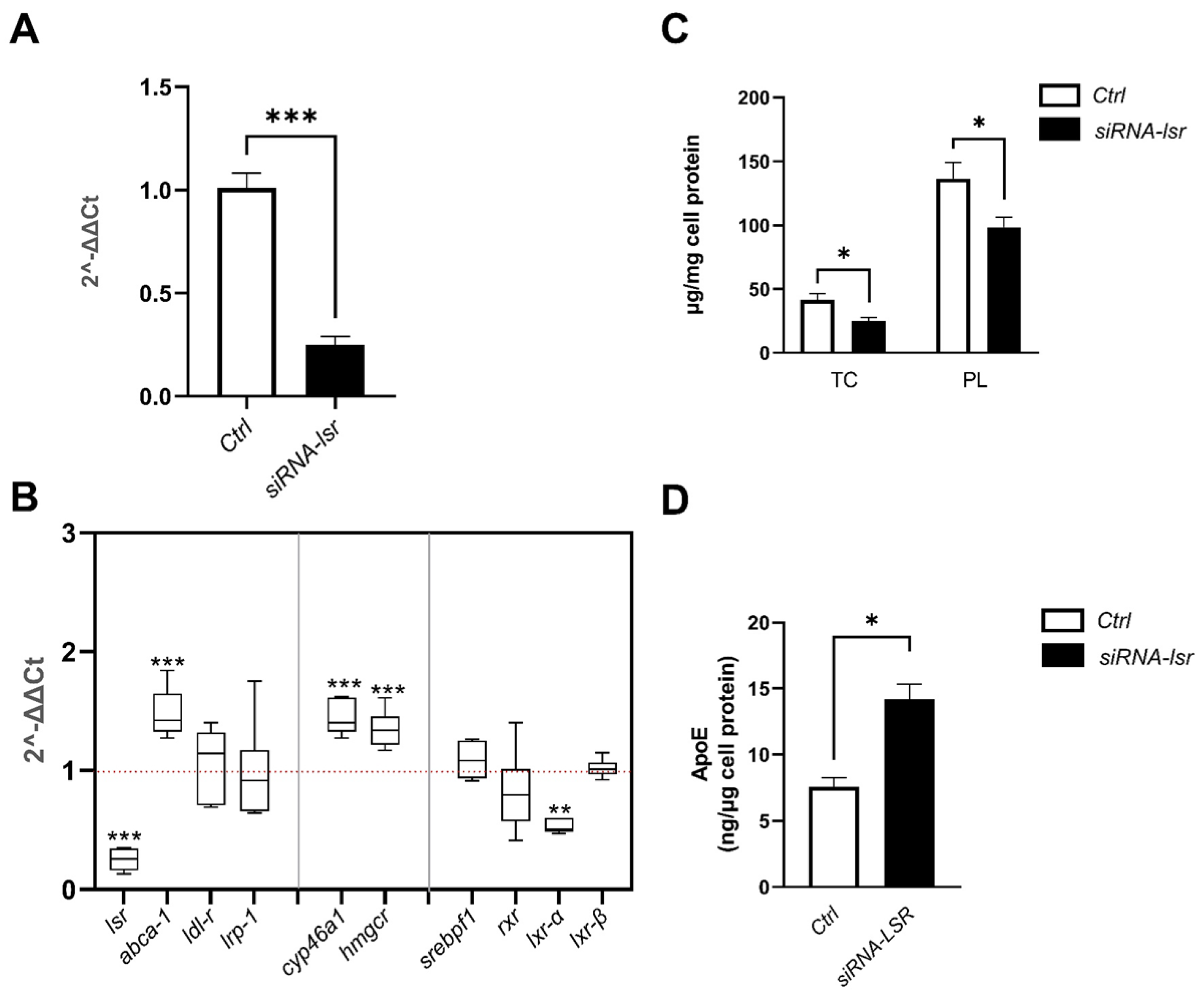
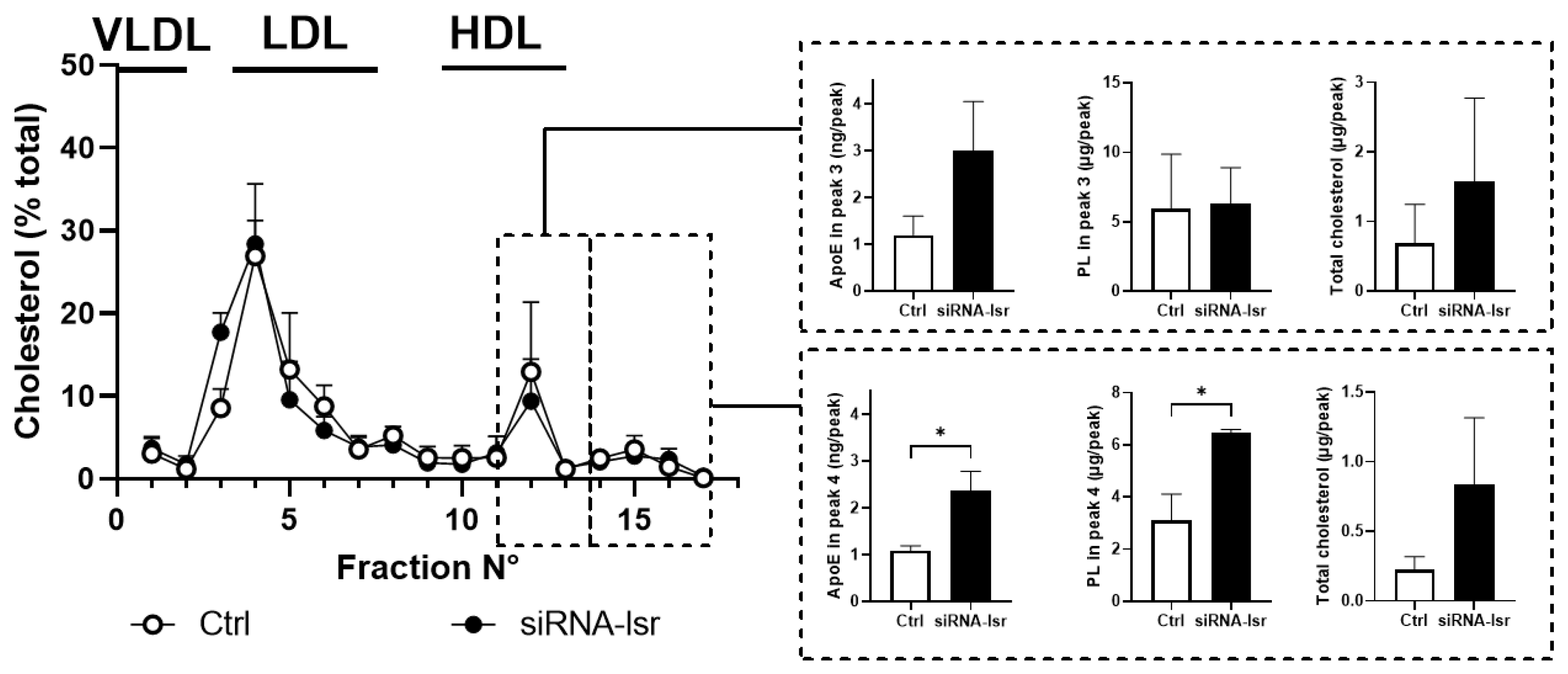
2.1. Effect of siRNA-Mediated lsr Knockdown on Astrocyte Cholesterol Homeostasis
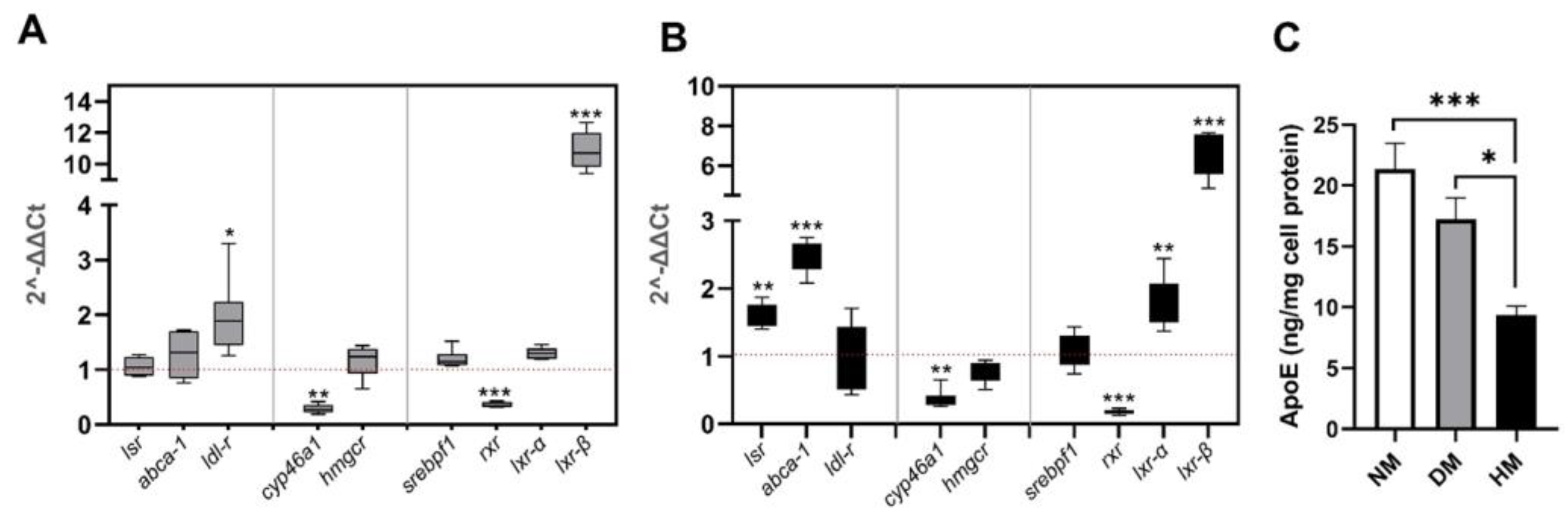
2.2. Effect of Culture Media Lipoprotein Content on lsr Expression in Astrocytes
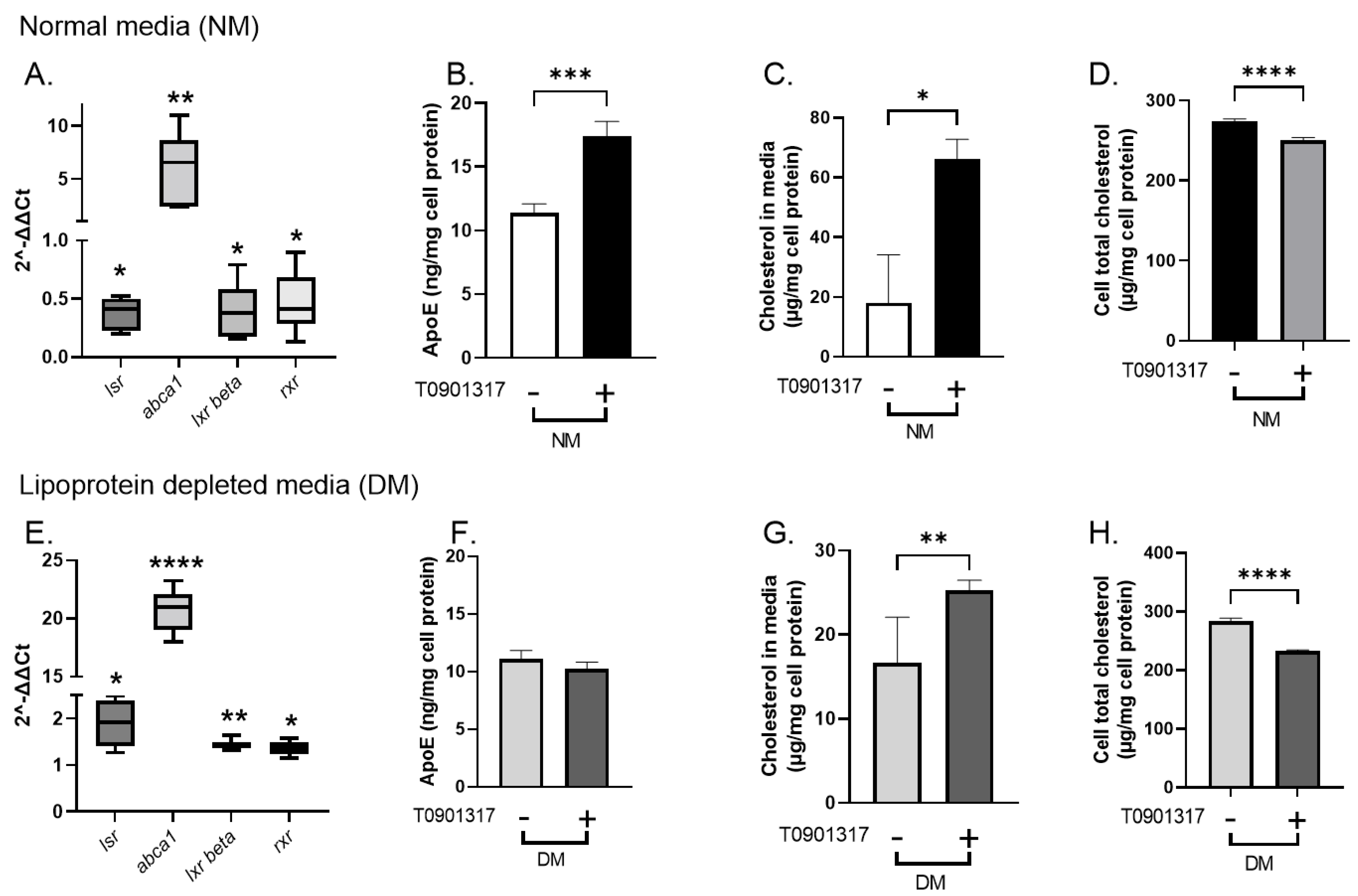
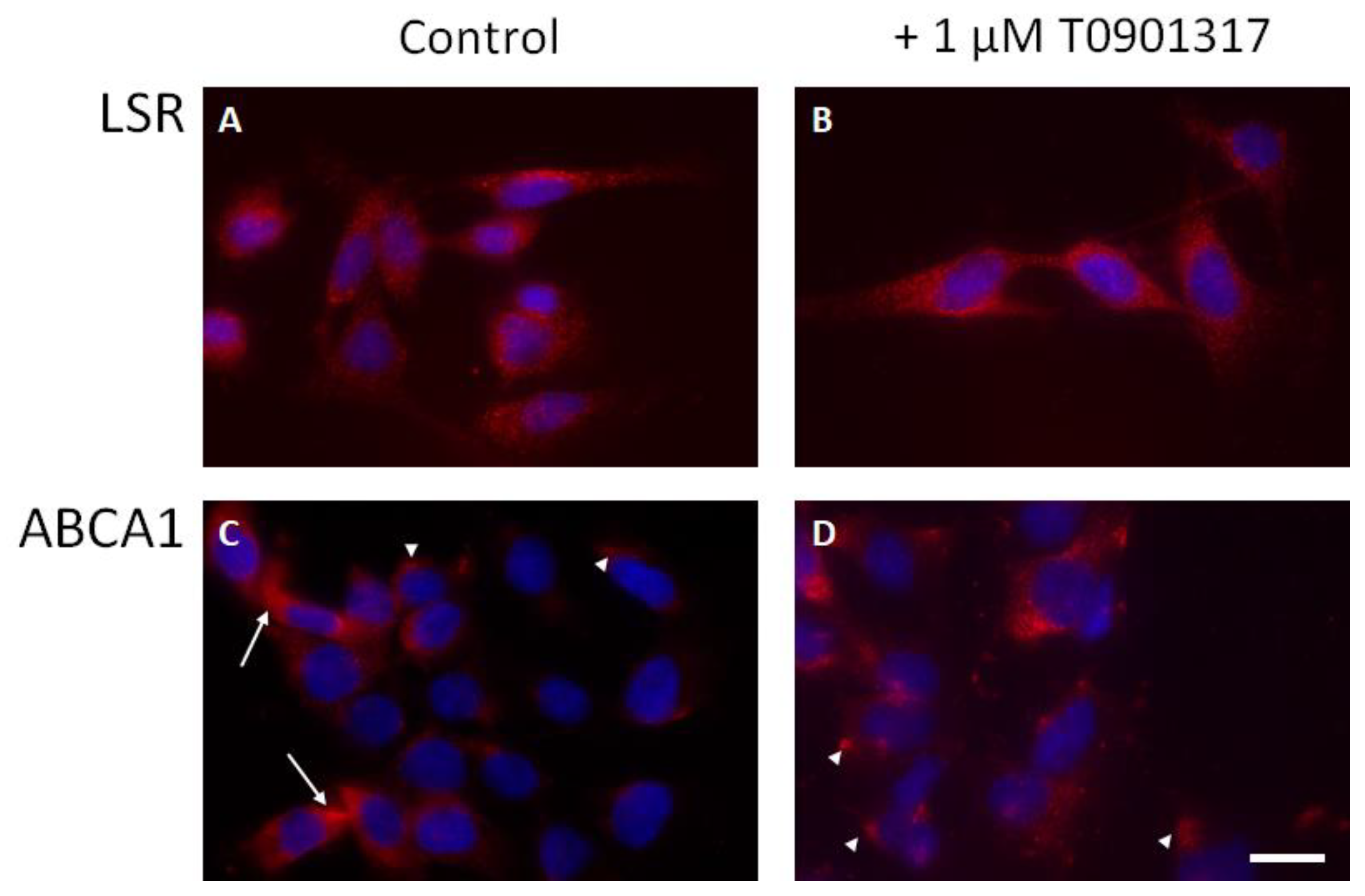
2.3. Effect of LXR Agonist T0901317 on lsr Expression in Astrocytes
2.4. LSR and ABCA1 Localization in Astrocytes
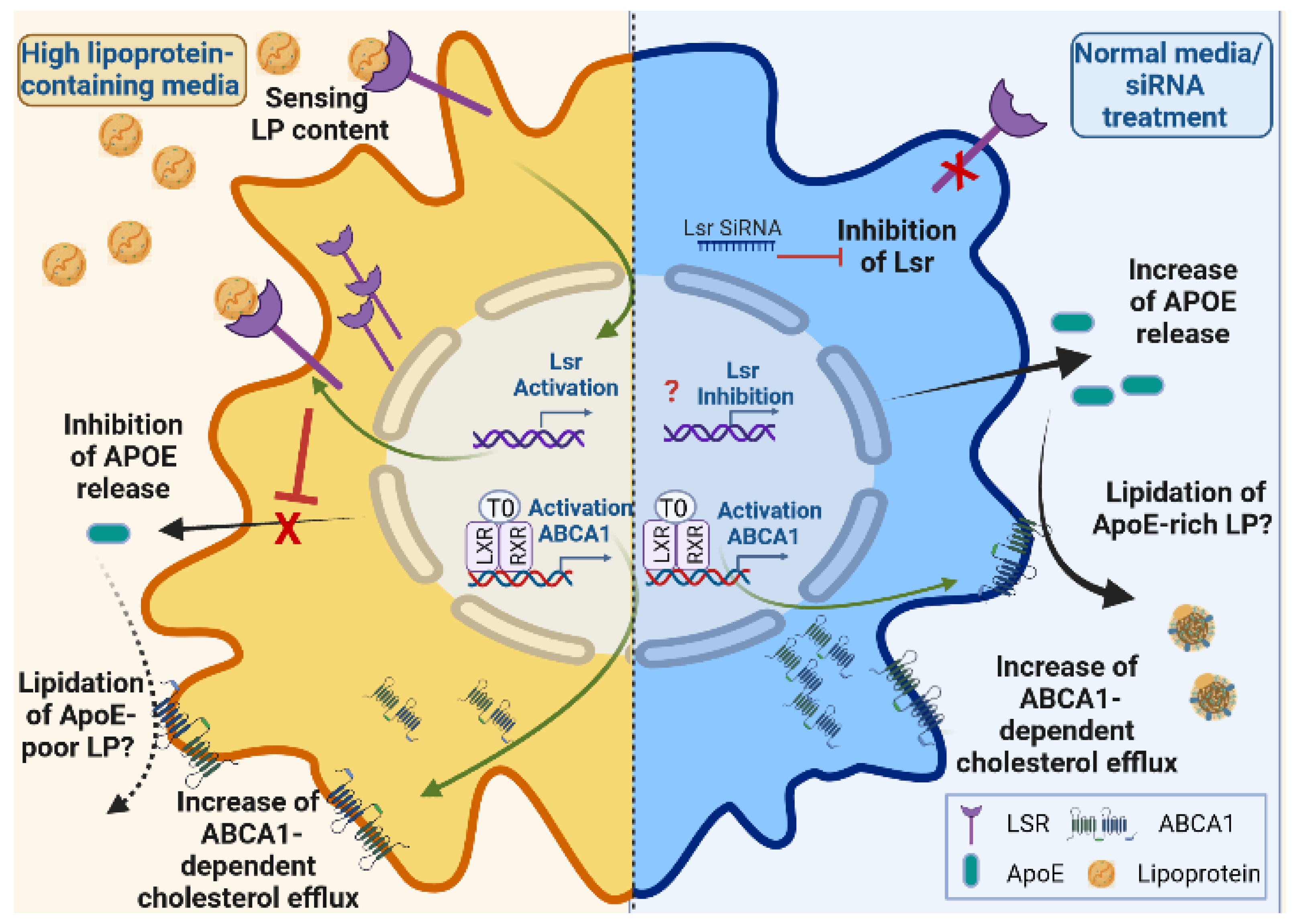
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.1.1. siRNA Treatment
4.1.2. Lipoprotein Enrichment (HM) or Depletion (DM) Treatment
4.1.3. LXR Agonist Treatment
4.1.4. Preparation of Samples for Analyses
4.2. qPCR (RNA Extraction, RT and Real-Time PCR)
4.3. ApoE and Lipid Assays
4.4. Immunocytochemistry
4.5. Lipoprotein Profiles
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, H.G.; Wheeler, M.A.; Quintana, F.J. Function and therapeutic value of astrocytes in neurological diseases. Nat. Rev. Drug Discov. 2022, 21, 339–358. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W.; Ungerer, N. Cholesterol metabolism in neurons and astrocytes. Prog. Lipid Res. 2011, 50, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, X.; Kusumo, H.; Costa, L.G.; Guizzetti, M. Cholesterol efflux is differentially regulated in neurons and astrocytes: Implications for brain cholesterol homeostasis. Biochim. Biophys. Acta 2013, 1831, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, C.N.; Raben, D.M. Lipid Metabolism Crosstalk in the Brain: Glia and Neurons. Front. Cell Neurosci. 2019, 13, 212. [Google Scholar] [CrossRef] [Green Version]
- Reitz, C. Dyslipidemia and the risk of Alzheimer’s disease. Curr. Atheroscler. Rep. 2013, 15, 307. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.G.; Li, L.; Muller, W.E.; Eckert, G.P. Cholesterol as a causative factor in Alzheimer’s disease: A debatable hypothesis. J. Neurochem. 2014, 129, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Soderberg, M.; Edlund, C.; Kristensson, K.; Dallner, G. Lipid compositions of different regions of the human brain during aging. J. Neurochem. 1990, 54, 415–423. [Google Scholar] [CrossRef]
- Svennerholm, L.; Bostrom, K.; Jungbjer, B.; Olsson, L. Membrane lipids of adult human brain: Lipid composition of frontal and temporal lobe in subjects of age 20 to 100 years. J. Neurochem. 1994, 63, 1802–1811. [Google Scholar] [CrossRef]
- Jiang, Q.; Lee, C.Y.; Mandrekar, S.; Wilkinson, B.; Cramer, P.; Zelcer, N.; Mann, K.; Lamb, B.; Willson, T.M.; Collins, J.L.; et al. ApoE promotes the proteolytic degradation of Abeta. Neuron 2008, 58, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Fakhoury, M. Microglia and Astrocytes in Alzheimer’s Disease: Implications for Therapy. Curr. Neuropharmacol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Chang, T.Y.; Yamauchi, Y.; Hasan, M.T.; Chang, C. Cellular cholesterol homeostasis and Alzheimer’s disease. J. Lipid Res. 2017, 58, 2239–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane-Donovan, C.; Herz, J. The ApoE receptors Vldlr and Apoer2 in central nervous system function and disease. J. Lipid Res. 2017, 58, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, M.; Tachibana, M.; Kanekiyo, T.; Bu, G. Role of LRP1 in the pathogenesis of Alzheimer’s disease: Evidence from clinical and preclinical studies. J. Lipid Res. 2017, 58, 1267–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hajj, A.; Yen, F.T.; Oster, T.; Malaplate, C.; Pauron, L.; Corbier, C.; Lanhers, M.C.; Claudepierre, T. Age-related changes in regiospecific expression of Lipolysis Stimulated Receptor (LSR) in mice brain. PLoS ONE 2019, 14, e0218812. [Google Scholar] [CrossRef] [PubMed]
- Yen, F.T.; Masson, M.; Clossais-Besnard, N.; Andre, P.; Grosset, J.M.; Bougueleret, L.; Dumas, J.B.; Guerassimenko, O.; Bihain, B.E. Molecular cloning of a lipolysis-stimulated remnant receptor expressed in the liver. J. Biol. Chem. 1999, 274, 13390–13398. [Google Scholar] [CrossRef] [PubMed]
- Yen, F.T.; Mann, C.J.; Guermani, L.M.; Hannouche, N.F.; Hubert, N.; Hornick, C.A.; Bordeau, V.N.; Agnani, G.; Bihain, B.E. Identification of a lipolysis-stimulated receptor that is distinct from the LDL receptor and the LDL receptor-related protein. Biochemistry 1994, 33, 1172–1180. [Google Scholar] [CrossRef]
- Yen, F.T.; Roitel, O.; Bonnard, L.; Notet, V.; Pratte, D.; Stenger, C.; Magueur, E.; Bihain, B.E. Lipolysis stimulated lipoprotein receptor: A novel molecular link between hyperlipidemia, weight gain, and atherosclerosis in mice. J. Biol. Chem. 2008, 283, 25650–25659. [Google Scholar] [CrossRef] [Green Version]
- Stenger, C.; Pincon, A.; Hanse, M.; Royer, L.; Comte, A.; Koziel, V.; Olivier, J.L.; Pillot, T.; Yen, F.T. Brain region-specific immunolocalization of the lipolysis-stimulated lipoprotein receptor (LSR) and altered cholesterol distribution in aged LSR+/- mice. J. Neurochem. 2012, 123, 467–476. [Google Scholar] [CrossRef]
- Pincon, A.; Thomas, M.H.; Huguet, M.; Allouche, A.; Colin, J.C.; Georges, A.; Derrien, A.; Lanhers, M.C.; Malaplate-Armand, C.; Oster, T.; et al. Increased susceptibility of dyslipidemic LSR+/− mice to amyloid stress is associated with changes in cortical cholesterol levels. J. Alzheimers Dis. 2015, 45, 195–204. [Google Scholar] [CrossRef]
- El Hajj, A.; Herzine, A.; Calcagno, G.; Desor, F.; Djelti, F.; Bombail, V.; Denis, I.; Oster, T.; Malaplate, C.; Vigier, M.; et al. Targeted Suppression of Lipoprotein Receptor LSR in Astrocytes Leads to Olfactory and Memory Deficits in Mice. Int. J. Mol. Sci. 2022, 23, 2049. [Google Scholar] [CrossRef]
- Morikawa, M.; Fryer, J.D.; Sullivan, P.M.; Christopher, E.A.; Wahrle, S.E.; DeMattos, R.B.; O’Dell, M.A.; Fagan, A.M.; Lashuel, H.A.; Walz, T.; et al. Production and characterization of astrocyte-derived human apolipoprotein E isoforms from immortalized astrocytes and their interactions with amyloid-beta. NeuroBiol. Dis. 2005, 19, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, J.L.; Brown, M.S. The low-density lipoprotein pathway and its relation to atherosclerosis. Annu. Rev. Biochem. 1977, 46, 897–930. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.Y.; Ou, X.; Hao, X.R.; Cao, D.L.; Tang, Y.L.; Hu, Y.W.; Li, X.X.; Tang, C.K. The effect of T0901317 on ATP-binding cassette transporter A1 and Niemann-Pick type C1 in apoE−/− mice. J. Cardiovasc. Pharmacol. 2008, 51, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zhao, R.Q.; Parro, C.; Zhao, W.; Chou, H.Y.; Robert, J.; Deeb, T.Z.; Raynoschek, C.; Barichievy, S.; Engkvist, O.; et al. Small molecule inducers of ABCA1 and apoE that act through indirect activation of the LXR pathway. J. Lipid Res. 2018, 59, 830–842. [Google Scholar] [CrossRef] [Green Version]
- Hirsch-Reinshagen, V.; Zhou, S.; Burgess, B.L.; Bernier, L.; McIsaac, S.A.; Chan, J.Y.; Tansley, G.H.; Cohn, J.S.; Hayden, M.R.; Wellington, C.L. Deficiency of ABCA1 impairs apolipoprotein E metabolism in brain. J. Biol. Chem. 2004, 279, 41197–41207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagan, A.M.; Holtzman, D.M.; Munson, G.; Mathur, T.; Schneider, D.; Chang, L.K.; Getz, G.S.; Reardon, C.A.; Lukens, J.; Shah, J.A.; et al. Unique lipoproteins secreted by primary astrocytes from wild type, apoE (−/−), and human apoE transgenic mice. J. Biol. Chem. 1999, 274, 30001–30007. [Google Scholar] [CrossRef] [Green Version]
- Vitali, C.; Wellington, C.L.; Calabresi, L. HDL and cholesterol handling in the brain. Cardiovasc. Res. 2014, 103, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast, N.; Lin, J.B.; Anderson, K.W.; Bjorkhem, I.; Pikuleva, I.A. Transcriptional and post-translational changes in the brain of mice deficient in cholesterol removal mediated by cytochrome P450 46A1 (CYP46A1). PLoS ONE 2017, 12, e0187168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartagena, C.M.; Ahmed, F.; Burns, M.P.; Pajoohesh-Ganji, A.; Pak, D.T.; Faden, A.I.; Rebeck, G.W. Cortical injury increases cholesterol 24S hydroxylase (Cyp46) levels in the rat brain. J. Neurotrauma 2008, 25, 1087–1098. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Kong, Q.; Lai, L.; Ray-Chaudhury, A.; Lin, C.L. Increased expression of cholesterol 24S-hydroxylase results in disruption of glial glutamate transporter EAAT2 association with lipid rafts: A potential role in Alzheimer’s disease. J. Neurochem. 2010, 113, 978–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, R.; Landreth, G.E. LXR Regulation of Brain Cholesterol: From Development to Disease. Trends Endocrinol. Metab. 2016, 27, 404–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frechin, M.; Stoeger, T.; Daetwyler, S.; Gehin, C.; Battich, N.; Damm, E.M.; Stergiou, L.; Riezman, H.; Pelkmans, L. Cell-intrinsic adaptation of lipid composition to local crowding drives social behaviour. Nature 2015, 523, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Sano, O.; Ito, S.; Kato, R.; Shimizu, Y.; Kobayashi, A.; Kimura, Y.; Kioka, N.; Hanada, K.; Ueda, K.; Matsuo, M. ABCA1, ABCG1, and ABCG4 are distributed to distinct membrane meso-domains and disturb detergent-resistant domains on the plasma membrane. PLoS ONE 2014, 9, e109886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genaro-Mattos, T.C.; Anderson, A.; Allen, L.B.; Korade, Z.; Mirnics, K. Cholesterol Biosynthesis and Uptake in Developing Neurons. ACS Chem. Neurosci. 2019, 10, 3671–3681. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.S.; Kobayashi, M.; Hayashi, H.; Zou, K.; Sawamura, N.; Fujita, S.C.; Yanagisawa, K.; Michikawa, M. Apolipoprotein E (ApoE) isoform-dependent lipid release from astrocytes prepared from human ApoE3 and ApoE4 knock-in mice. J. Biol. Chem. 2002, 277, 29919–29926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slot, R.E.; Van Harten, A.C.; Kester, M.I.; Jongbloed, W.; Bouwman, F.H.; Teunissen, C.E.; Scheltens, P.; Veerhuis, R.; van der Flier, W.M. Apolipoprotein A1 in Cerebrospinal Fluid and Plasma and Progression to Alzheimer’s Disease in Non-Demented Elderly. J. Alzheimers Dis. 2017, 56, 687–697. [Google Scholar] [CrossRef]
- Endres, K. Apolipoprotein A1, the neglected relative of Apolipoprotein E and its potential role in Alzheimer’s disease. Neural Regen Res. 2021, 16, 2141–2148. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| abca1 | CAACCCCTGCTTCCGTTATCCAA | GAGAACAGGCGAGACACGATGGAC |
| cyp46a1 | GGCTAAGAAGT TGGTCCTGTTGTAAGA | GGTGGACATCAGGAACTTCTTGACT |
| hmgcr | CCCCACATTCACTCTTGACGCTCT | GCTGGCGGACGCCTGACAT |
| ldl-r | TGGCTATACCTACCCCTCAAGACAG | GATCCCGGAAAGAGACGGAT |
| lrp-1 | CGAGAGCCTTTGTGCTGGATGA | CGGATGTCCTCAATGAGGG |
| lsr (total) | AGTAATACACTCCACTGTCTCCCCAG | CAGGAGAATCACCATCACAGGAA |
| nr1h3 (lxr α) | AGGAGTGTCGACTTCGCAAA | CTCTTCTTGCCGCAGTTT |
| nr1h2 (lxr β) | GCTCTGCCTACATCGTGGTC | CTCATGGCCCAGCATCTT |
| pgk1 | GAGCCTCACTGTCCAAACTA | CTTTAGCGCCTCCCAAGATA |
| rxr | CAAACATGGGGCTGAACC | GCCCAC-CCACAAGAGTGA |
| srebpf1 | GGTCCAGCAGGTCCCAGTTGT | CTGCAGTCTTCACGGTGGCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herzine, A.; Sekkat, G.; Kaminski, S.; Calcagno, G.; Boschi-Muller, S.; Safi, H.; Corbier, C.; Siest, S.; Claudepierre, T.; Yen, F.T. Lipolysis-Stimulated Lipoprotein Receptor Acts as Sensor to Regulate ApoE Release in Astrocytes. Int. J. Mol. Sci. 2022, 23, 8630. https://doi.org/10.3390/ijms23158630
Herzine A, Sekkat G, Kaminski S, Calcagno G, Boschi-Muller S, Safi H, Corbier C, Siest S, Claudepierre T, Yen FT. Lipolysis-Stimulated Lipoprotein Receptor Acts as Sensor to Regulate ApoE Release in Astrocytes. International Journal of Molecular Sciences. 2022; 23(15):8630. https://doi.org/10.3390/ijms23158630
Chicago/Turabian StyleHerzine, Ameziane, Ghita Sekkat, Sandra Kaminski, Gaetano Calcagno, Sandrine Boschi-Muller, Hela Safi, Catherine Corbier, Sophie Siest, Thomas Claudepierre, and Frances T. Yen. 2022. "Lipolysis-Stimulated Lipoprotein Receptor Acts as Sensor to Regulate ApoE Release in Astrocytes" International Journal of Molecular Sciences 23, no. 15: 8630. https://doi.org/10.3390/ijms23158630
APA StyleHerzine, A., Sekkat, G., Kaminski, S., Calcagno, G., Boschi-Muller, S., Safi, H., Corbier, C., Siest, S., Claudepierre, T., & Yen, F. T. (2022). Lipolysis-Stimulated Lipoprotein Receptor Acts as Sensor to Regulate ApoE Release in Astrocytes. International Journal of Molecular Sciences, 23(15), 8630. https://doi.org/10.3390/ijms23158630