
Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis)

 , and
, and
Abstract
:1. Introduction
2. Results
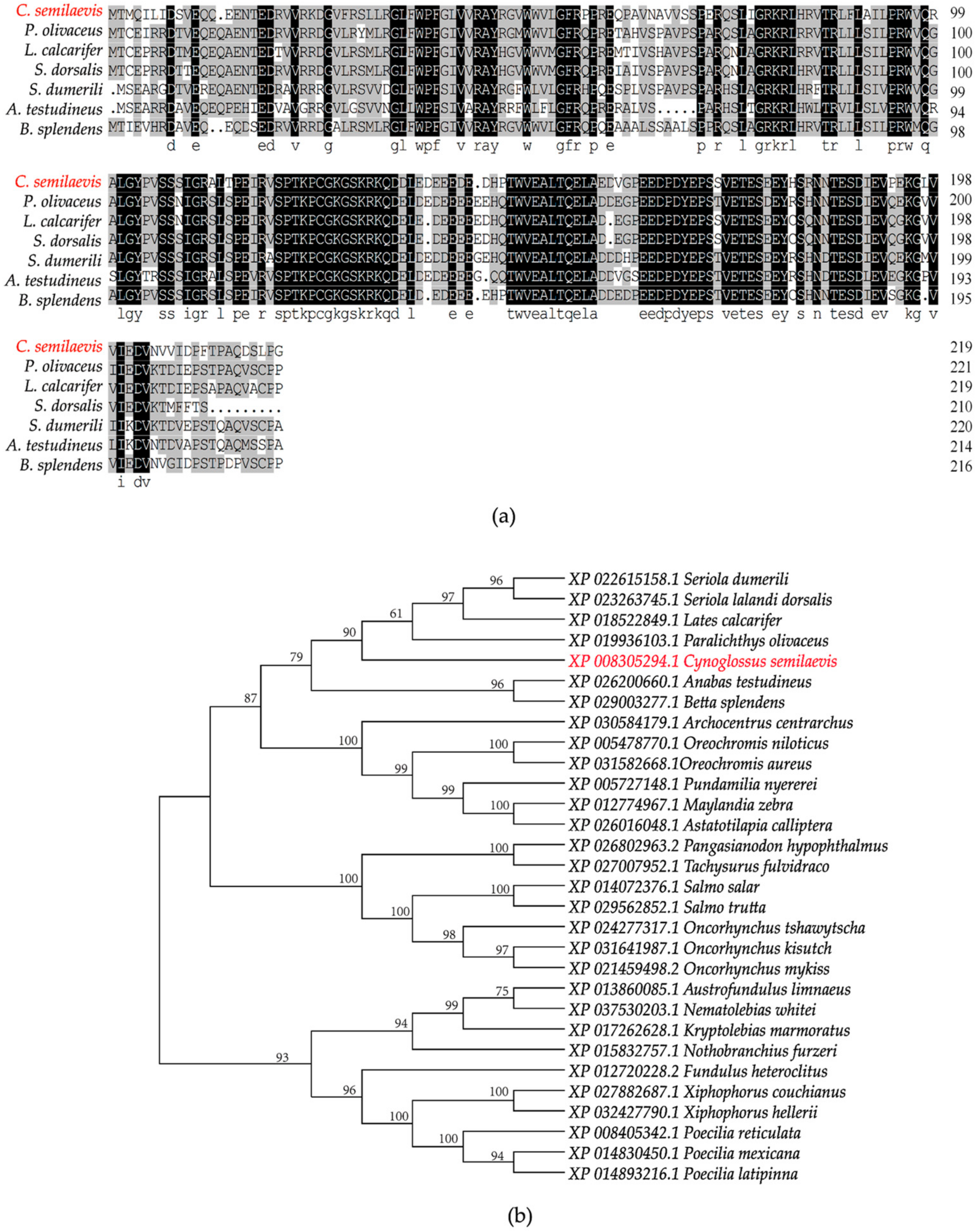
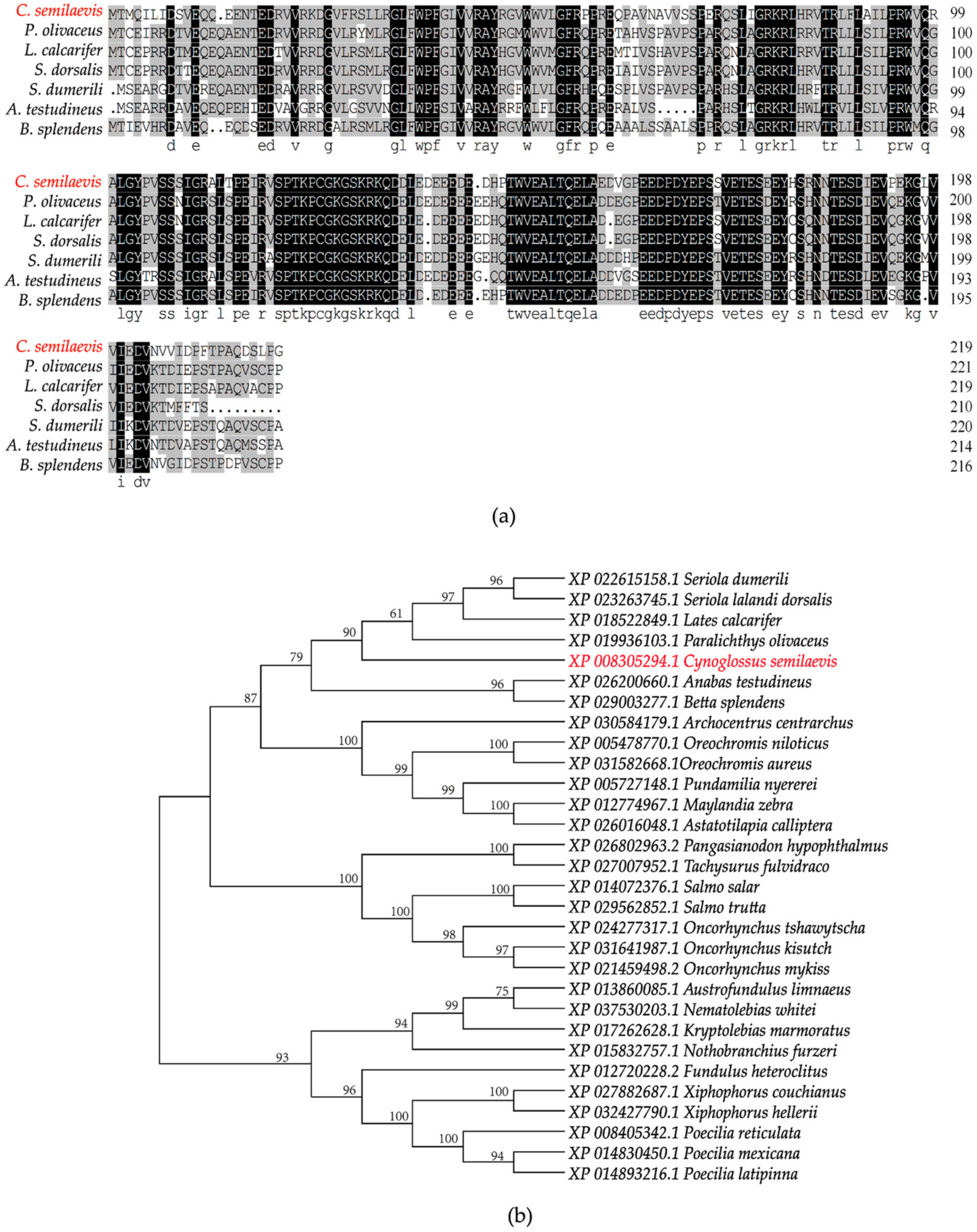
2.1. Cloning and Sequence Analysis of org
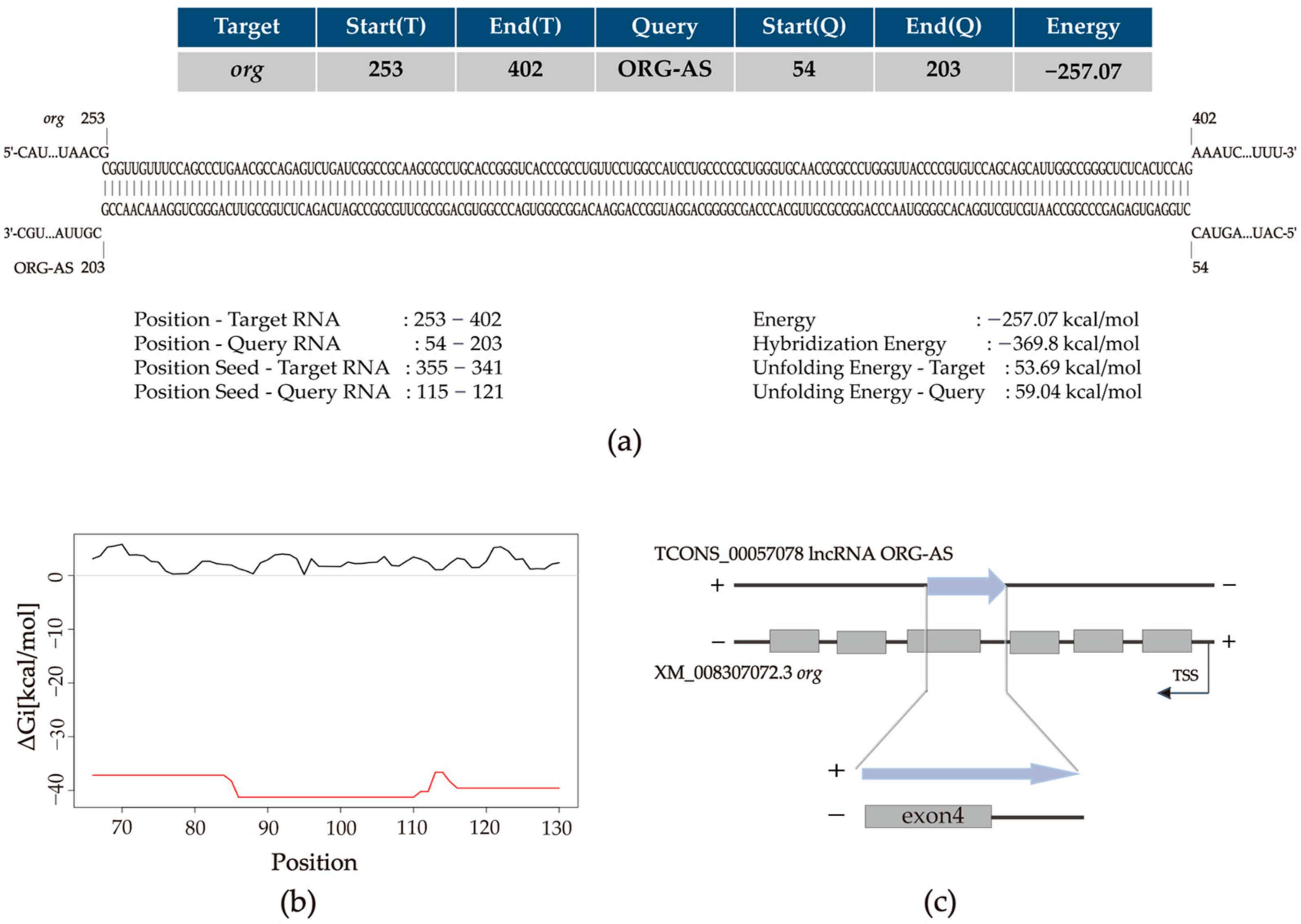
2.2. ORG-AS Is Predicted to Bind to org
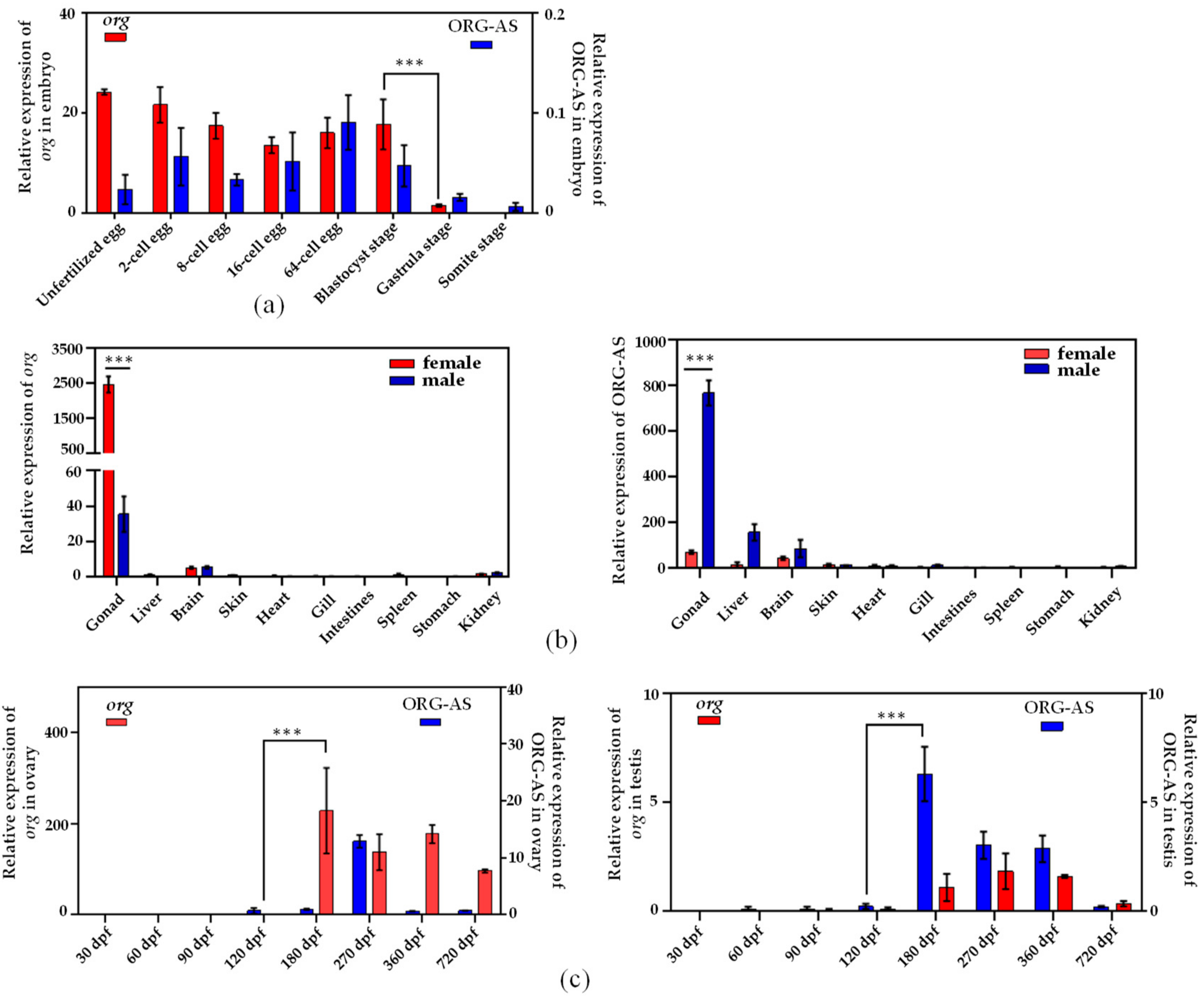
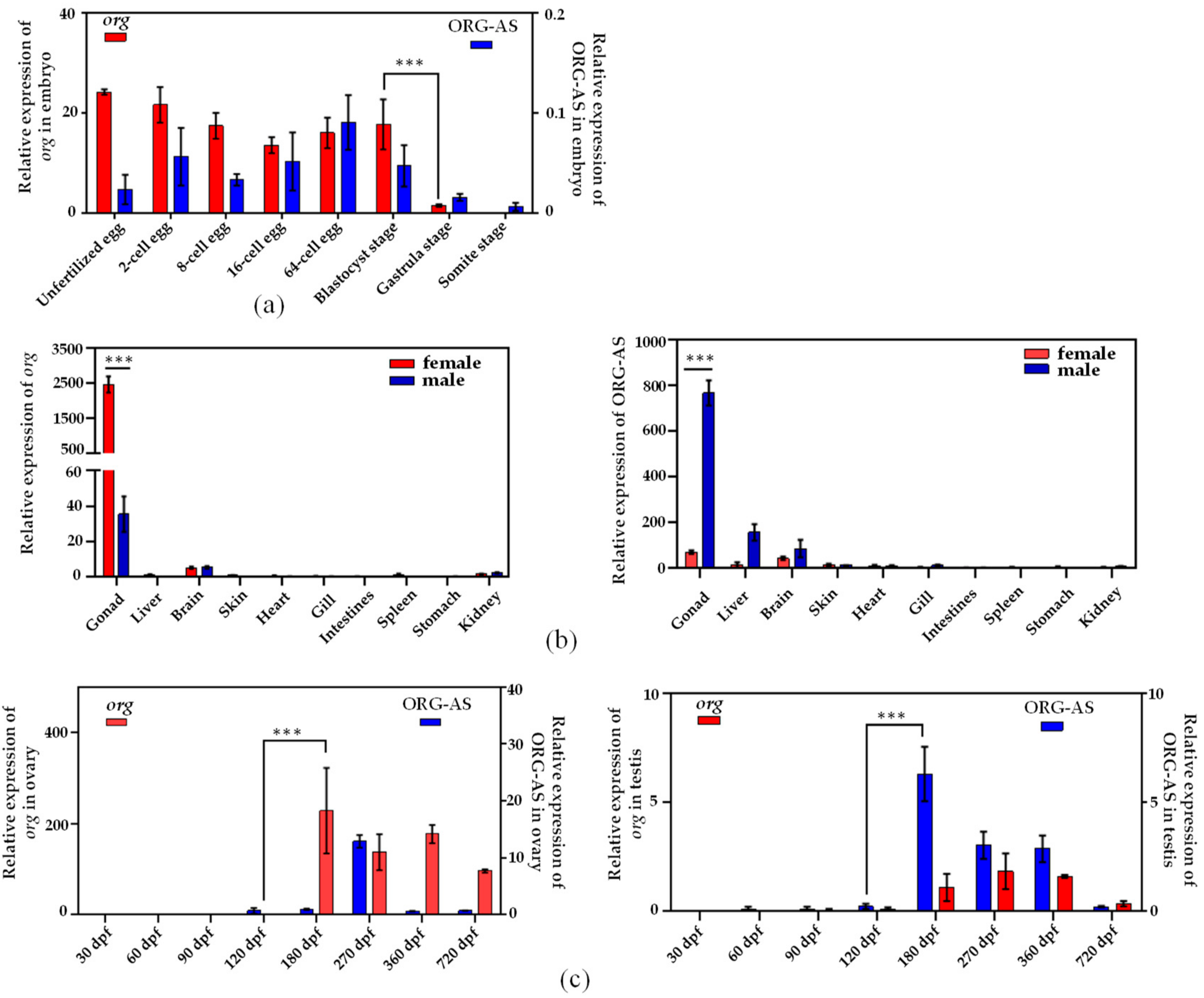
2.3. Expression Patterns of org and ORG-AS
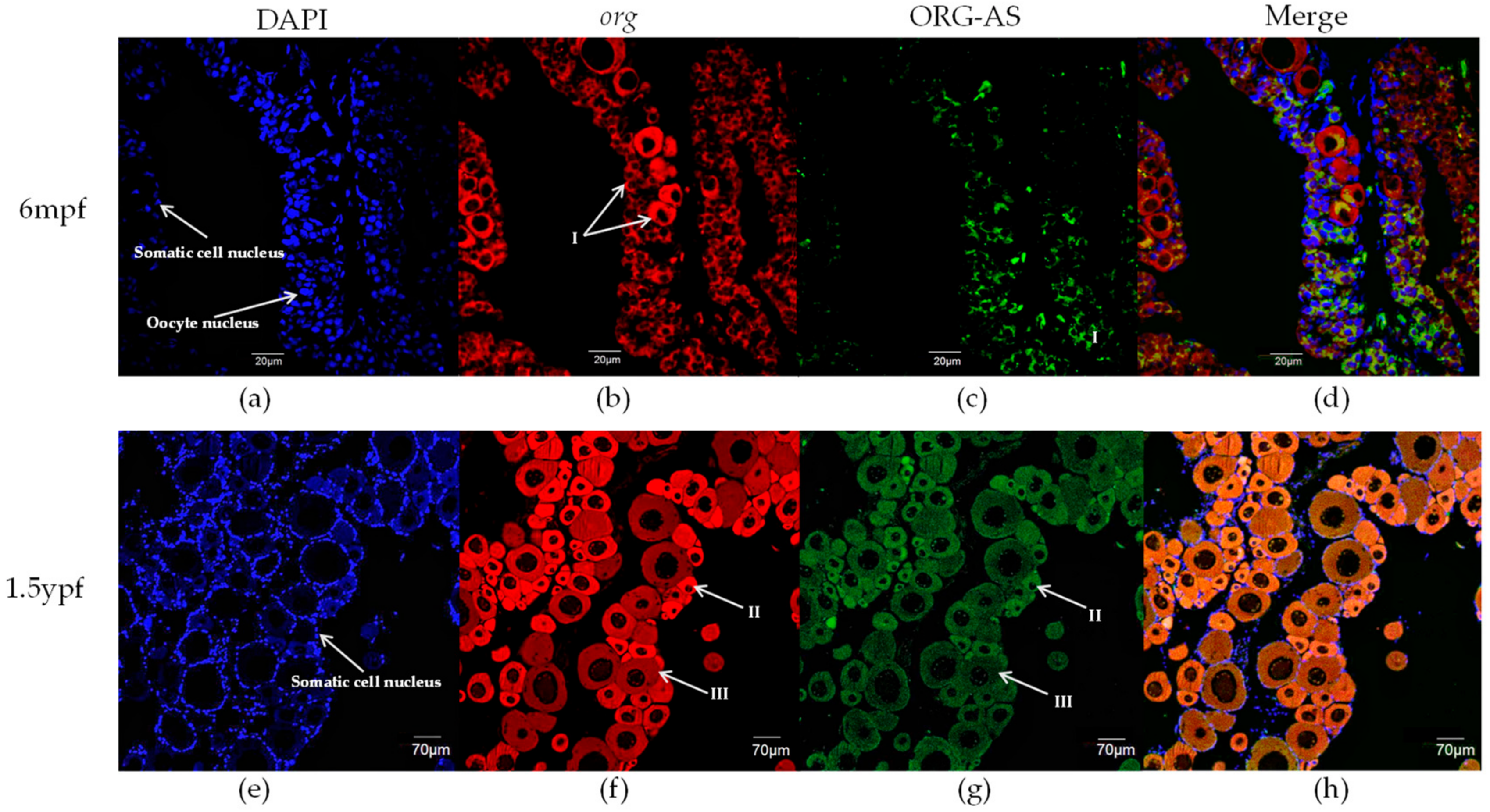
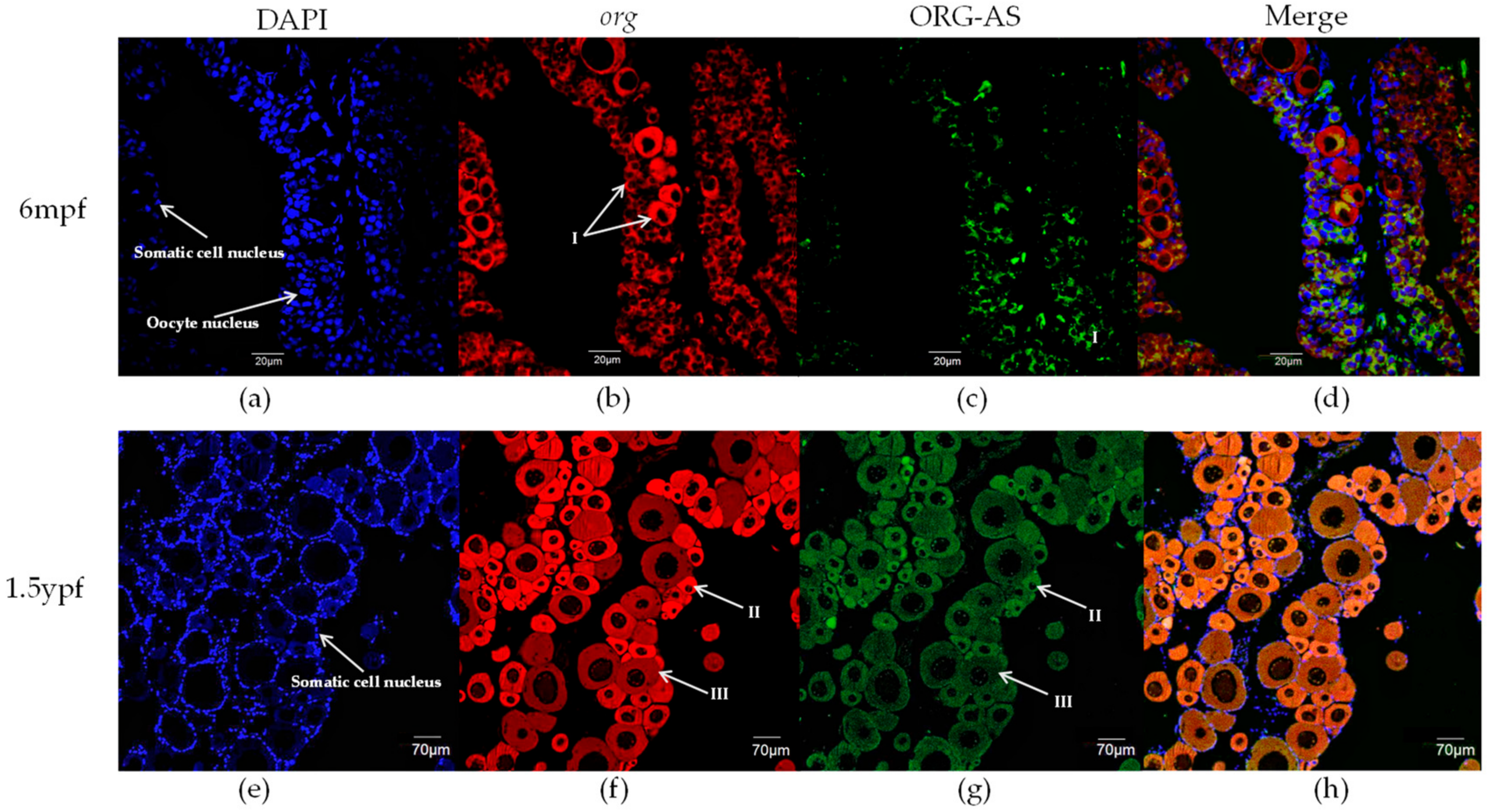
2.4. FISH of org and ORG-AS in Gonads
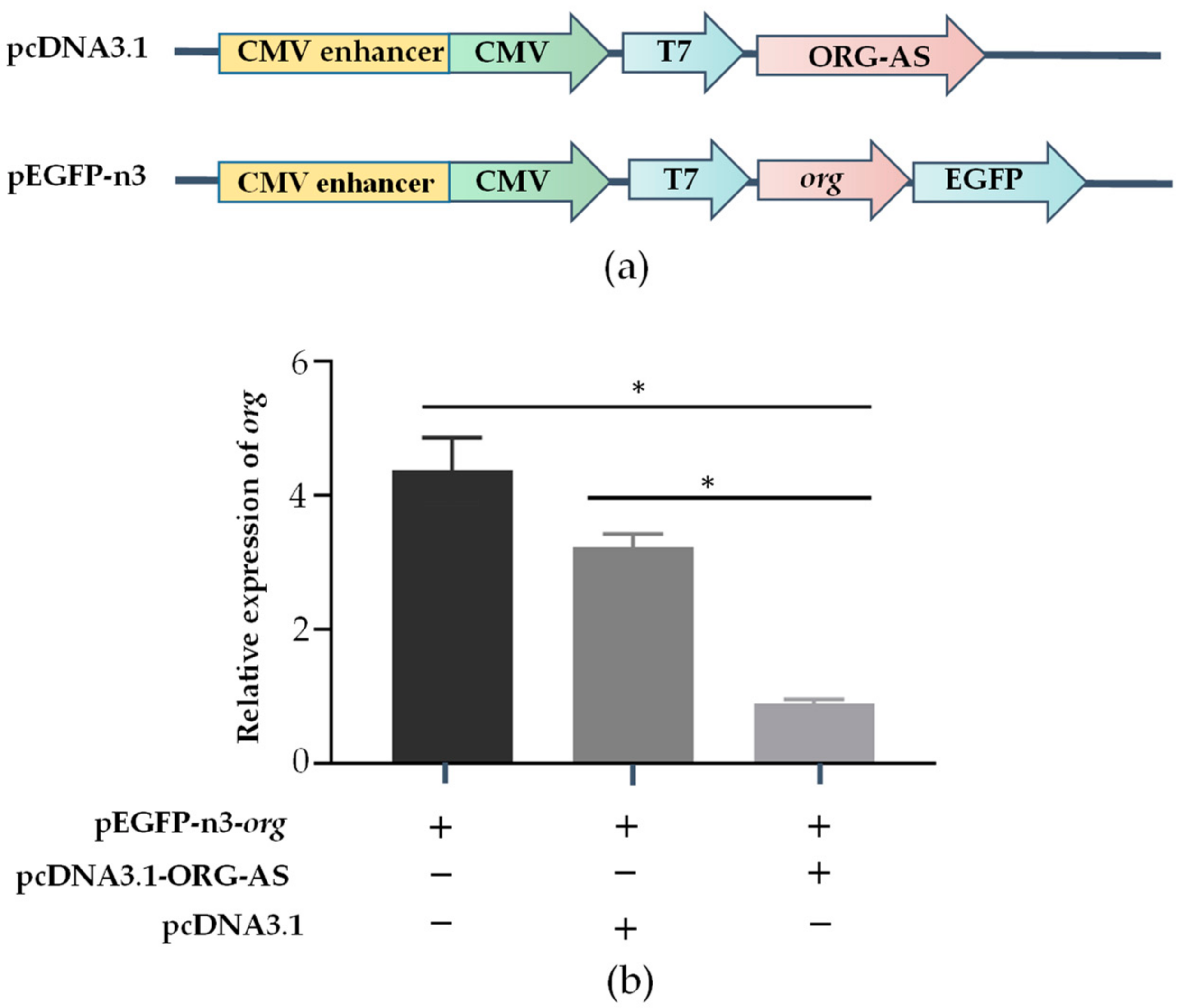
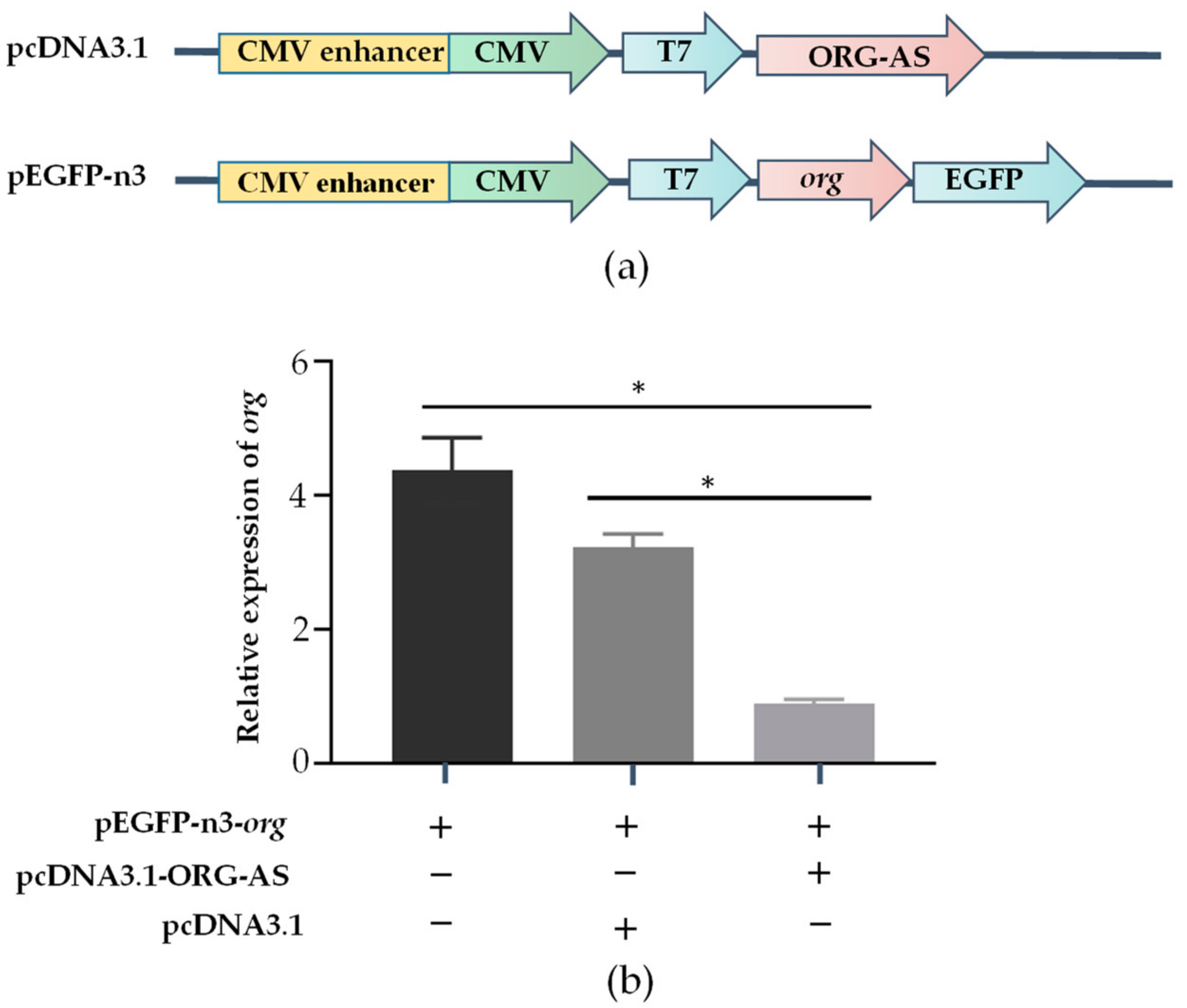
2.5. ORG-AS Negatively Regulates the Expression of org
3. Discussion
4. Materials and Methods
4.1. Experimental Fish and Tissue Sampling
4.2. Cloning of org
4.3. Real-Time Quantitative PCR
4.4. Fluorescence In Situ Hybridization of org and ORG-AS
4.5. Cell Culture, Vectors, and Transfection
4.6. Sequence Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gouw, J.W.; Pinkse, M.W.; Vos, H.R.; Moshkin, Y.; Verrijzer, C.P.; Heck, A.J.; Krijgsveld, J. In vivo stable isotope labeling of fruit flies reveals post-transcriptional regulation in the maternal-to-zygotic transition. Mol. Cell Proteom. 2009, 8, 1566–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadros, W.; Lipshitz, H.D. The maternal-to-zygotic transition: A play in two acts. Development 2009, 136, 3033–3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Ji, S.-Y.; Sha, Q.-Q.; Dang, Y.; Zhou, J.-J.; Zhang, Y.-L.; Liu, Y.; Wang, Z.-W.; Hu, B.; Sun, Q.-Y. BTG4 is a meiotic cell cycle–coupled maternal-zygotic-transition licensing factor in oocytes. Nat. Struct. Mol. Biol. 2016, 23, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Nüsslein-Volhard, C.; Lohs-Schardin, M.; Sander, K.; Cremer, C. A dorso-ventral shift of embryonic primordia in a new maternal-effect mutant of Drosophila. Nature 1980, 283, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Bowerman, B. 3 Maternal Control of Pattern Formation in Early Caenorhabditis elegans Embryos. Curr. Top. Dev. Biol. 1998, 39, 73–117. [Google Scholar] [PubMed]
- Heasman, J. Maternal determinants of embryonic cell fate. Semin. Cell Dev. Biol. 2006, 17, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Viveiros, M.M.; Eppig, J.J.; Bai, Y.; Fitzpatrick, S.L.; Matzuk, M.M. Zygote arrest 1 (Zar1) is a novel maternal-effect gene critical for the oocyte-to-embryo transition. Nat. Genet. 2003, 33, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Ji, S.-Y.; Zhu, Y.-Z.; Wu, Y.-W.; Shen, L.; Fan, H.-Y. ZAR1 and ZAR2 are required for oocyte meiotic maturation by regulating the maternal transcriptome and mRNA translational activation. Nucleic Acids Res. 2019, 47, 11387–11402. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.; Kawakami, K.; Hopkins, N. Zebrafish vasa homologue RNA is localized to the cleavage planes of 2-and 4-cell-stage embryos and is expressed in the primordial germ cells. Development 1997, 124, 3157–3165. [Google Scholar] [CrossRef] [PubMed]
- Köprunner, M.; Thisse, C.; Thisse, B.; Raz, E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev. 2001, 15, 2877–2885. [Google Scholar] [CrossRef]
- Tan, C.-H.; Lee, T.-C.; Weeraratne, S.D.; Korzh, V.; Lim, T.-M.; Gong, Z. RETRACTED: Ziwi, the zebrafish homologue of the Drosophila piwi: Co-localization with vasa at the embryonic genital ridge and gonad-specific expression in the adults. Gene Expr. Patterns 2002, 2, 257–260. [Google Scholar] [CrossRef]
- Gosden, R. Oogenesis as a foundation for embryogenesis. Mol. Cell Endocrinol. 2002, 186, 149–153. [Google Scholar] [CrossRef]
- Rodrigues, P.; Limback, D.; McGinnis, L.K.; Plancha, C.E.; Albertini, D.F. Oogenesis: Prospects and challenges for the future. J. Cell Physiol. 2008, 216, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, F.; Smitz, J. Molecular control of oogenesis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Pan, H.; Montgomery, R.L.; Olson, E.N.; Schultz, R.M. Compensatory functions of histone deacetylase 1 (HDAC1) and HDAC2 regulate transcription and apoptosis during mouse oocyte development. Proc. Natl. Acad. Sci. USA 2012, 109, E481–E489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.-F.; Liu, Q.; Liu, K.-Q.; Wang, H.-Y.; Li, C.-H.; Wang, Q.; Shao, C.-W. Identification of global alternative splicing and sex-specific splicing via comparative transcriptome analysis of gonads of Chinese tongue sole (Cynoglossus semilaevis). Zool. Res. 2022, 43, 319. [Google Scholar] [CrossRef]
- Zhu, L.-Y.; Zhu, Y.-R.; Dai, D.-J.; Wang, X.; Jin, H.-C. Epigenetic regulation of alternative splicing. Am. J. Cancer Res. 2018, 8, 2346. [Google Scholar] [PubMed]
- Ulitsky, I.; Bartel, D.P. lincRNAs: Genomics, evolution, and mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [Green Version]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-gamma locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, B.; Li, S.; Wang, Q.; Tang, L.; Huang, F.; Zhang, Z.; Mahboobe, S.; Shao, C. lncRNA DMRT2-AS acts as a transcriptional regulator of dmrt2 involving in sex differentiation in the Chinese tongue sole (Cynoglossus semilaevis). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2021, 253, 110542. [Google Scholar] [CrossRef]
- Kato, Y.; Perez, C.A.G.; Ishak, N.S.M.; Nong, Q.D.; Sudo, Y.; Matsuura, T.; Wada, T.; Watanabe, H. A 5′ UTR-Overlapping LncRNA Activates the Male-Determining Gene doublesex1 in the Crustacean Daphnia magna. Curr. Biol. 2018, 28, 1811–1817.e1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Ma, H.Y.; Xu, G.B.; Shao, C.W.; Tian, Y.S.; Ji, X.S.; Yang, J.F.; Chen, S.L. Construction of a genetic linkage map and mapping of a female-specific DNA marker in half-smooth tongue sole (Cynoglossus semilaevis). Mar. Biotechnol. 2009, 11, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Ma, W.; Li, J.; Xu, Y.; Li, W.; Zhao, Y.; Deng, F. Cloning and characterization of a novel oocyte-specific gene zorg in zebrafish. Theriogenology 2009, 71, 441–449. [Google Scholar] [CrossRef]
- Li, L.; Zheng, P.; Dean, J. Maternal control of early mouse development. Development 2010, 137, 859–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.T.; Bonneau, A.R.; Takacs, C.M.; Bazzini, A.A.; DiVito, K.R.; Fleming, E.S.; Giraldez, A.J. Nanog, Pou5f1 and SoxB1 activate zygotic gene expression during the maternal-to-zygotic transition. Nature 2013, 503, 360–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouleau, A.; Desvignes, T.; Traverso, J.M.; Nguyen, T.; Chesnel, F.; Fauvel, C.; Bobe, J. Maternally inherited npm2 mRNA is crucial for egg developmental competence in zebrafish. Biol. Reprod. 2014, 91, 43. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.T.; Patinote, A.; Guiguen, Y.; Bobe, J. foxr1 is a novel maternal-effect gene in fish that is required for early embryonic success. PeerJ 2018, 6, e5534. [Google Scholar] [CrossRef]
- Ma, X.; Liu, X.; Wen, H.; Xu, Y.; Zhang, L. Histological observation on gonadal sex differentiation in Cynoglossus semilaevis Günther. Mar. Fish Res. 2006, 27, 55–61. [Google Scholar]
- Liu, X.; Huang, Y.; Tan, F.; Wang, H.-Y.; Chen, J.-Y.; Zhang, X.; Zhao, X.; Liu, K.; Wang, Q.; Liu, S. Single-Cell Atlas of the Chinese Tongue Sole (Cynoglossus semilaevis) Ovary Reveals Transcriptional Programs of Oogenesis in Fish. Front. Cell Dev. Biol. 2022, 10, 828124. [Google Scholar] [CrossRef]
- Andrade, R.; Bazzoli, N.; Rizzo, E.; Sato, Y. Continuous gametogenesis in the neotropical freshwater teleost, Bryconops affinis (Pisces: Characidae). Tissue Cell 2001, 33, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Arun, G.; Mao, Y.S.; Lazar, Z.; Hung, G.; Bhattacharjee, G.; Xiao, X.; Booth, C.J.; Wu, J.; Zhang, C.; et al. The lncRNA Malat1 is dispensable for mouse development but its transcription plays a cis-regulatory role in the adult. Cell Rep. 2012, 2, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, F.; Mendell, J.T. Functional classification and experimental dissection of long noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, S.; Gao, F.; Meng, L.; Hu, Q.; Song, W.; Shao, C.; Lv, W. SCAR-transformation of sex-specific SSR marker and its application in half-smooth tongue sole (Cynoglossus semiliaevis). J. Agric. Biotechnol. 2014, 22, 787–792. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) |
|---|---|
| org-F | CCAGCAGGCTTCAGAACCACCG |
| org-R | CAGTAACCAACATACGCAGGAA |
| org-3-GSP | CCCTGACACTGCCGTGTTTGCCCACTAGA |
| org-3-NGSP | ACTGGTCACTTGACACTTGCTCG |
| org-5-GSP | AGCCCAGCACCCACCAAACTCCACGGT |
| org-5-NGSP | CCACTCTGTCCTCCGTGTTCTC |
| org-Q-F | CAGCAGGAGGAGAACACGGA |
| org-Q-R | ACCAAACTCCACGGTATGCC |
| ORG-AS-Q-F | CACTTCGGAGAACACTGGCG |
| ORG-AS-Q-R | TGGGGCTCTTCTATTGGTCG |
| β-actin-Q-F | GCTGTGCTGTCCCTGTA |
| β-actin-Q-R | GAGTAGCCACGCTCTGTC |
| Cs-18S rRNA-F | GGTCTGTGATGCCCTTAGATGTC |
| Cs-18S rRNA-R | AGTGGGGTTCAGCGGGGTTAC |
| hACTINB-Q-F | GATGATATCGCCGCGCTCGT |
| hACTINB-Q-R | GTAGATGGGCACAGTGTGGGTG |
| org-EcoRI-F | CGGAATTCATGACCATGCAAATCCTC |
| org-KpnI-R | CGGGTACCAACACCAGGCAAAGAGT |
| ORG-AS-KpnI-F | CGGAATTCCTGGAGTGAGAGCCCGG |
| ORG-AS-EcoRI-R | GCTCTAGAACGCTTGGTTTGAACTTGATG |
| org-5′Cy3 | TGGGCTCGTAGTCCGGATCTTCCTCTGGGCC |
| ORG-AS-5′6-FAM | CCTCATAGACTCTTCAGGACAGAGTGGTCAGGAAGG |
| sex-F | CCTAAATGATGGATGTAGATTCTGTC |
| sex-R | GATCCAGAGAAAATAAACCCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Feng, B.; Wang, Q.; Tang, L.; Liu, Q.; Ma, W.; Li, C.; Shao, C. Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis). Int. J. Mol. Sci. 2022, 23, 8605. https://doi.org/10.3390/ijms23158605
Zhao X, Feng B, Wang Q, Tang L, Liu Q, Ma W, Li C, Shao C. Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis). International Journal of Molecular Sciences. 2022; 23(15):8605. https://doi.org/10.3390/ijms23158605
Chicago/Turabian StyleZhao, Xiaona, Bo Feng, Qian Wang, Lili Tang, Qian Liu, Wenxiu Ma, Chenghua Li, and Changwei Shao. 2022. "Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis)" International Journal of Molecular Sciences 23, no. 15: 8605. https://doi.org/10.3390/ijms23158605
APA StyleZhao, X., Feng, B., Wang, Q., Tang, L., Liu, Q., Ma, W., Li, C., & Shao, C. (2022). Cloning of the Maternal Effector Gene org and Its Regulation by lncRNA ORG-AS in Chinese Tongue Sole (Cynoglossus semilaevis). International Journal of Molecular Sciences, 23(15), 8605. https://doi.org/10.3390/ijms23158605