
Transgenic Mice for the Translational Study of Neuropathic Pain and Dystonia

 ,
,  , ,
, ,  and
and
Abstract
:1. Dystonia and Pain
2. TorsinA
3. Different Transgenic Models of Dystonia
4. Transgenic Modelling Is Fundamental for the Translational Study of Pain: Neuropathic Pain in hMT and hWT Transgenic Mice
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Schappert, S.M.; Burt, C.W. Ambulatory care visits to physician offices, hospital outpatient departments, and emergency departments: United States, 2001–2002. In Vital and Health Statistics; Series 13, Data from the National Health Survey; U.S. Department of Health and Human Services: Hyattsville, MD, USA, 2006; pp. 1–66. [Google Scholar]
- Abbott, C.A.; Malik, R.A.; van Ross, E.R.; Kulkarni, J.; Boulton, A.J. Prevalence and characteristics of painful diabetic neuropathy in a large community-based diabetic population in the U.K. Diabetes Care 2011, 34, 2220–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsden, C.D. The problem of adult-onset idiopathic torsion dystonia and other isolated dyskinesias in adult life (including blepharospasm, oromandibular dystonia, dystonic writer’s cramp, and torticollis, or axial dystonia). Adv. Neurol. 1976, 14, 259–276. [Google Scholar] [PubMed]
- Jinnah, H.A.; Hess, E.J. A new twist on the anatomy of dystonia: The basal ganglia and the cerebellum? Neurology 2006, 67, 1740–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scuteri, D.; Rombolà, L.; Natoli, S.; Pisani, A.; Bonsi, P.; Watanabe, C.; Bagetta, G.; Tonin, P.; Corasaniti, M.T. Effect of Gabapentin in a Neuropathic Pain Model in Mice Overexpressing Human Wild-Type or Human Mutated Torsin A. Life 2021, 11, 41. [Google Scholar] [CrossRef]
- Jiang, M.; Gold, M.S.; Boulay, G.; Spicher, K.; Peyton, M.; Brabet, P.; Srinivasan, Y.; Rudolph, U.; Ellison, G.; Birnbaumer, L. Multiple neurological abnormalities in mice deficient in the G protein Go. Proc. Natl. Acad. Sci. USA 1998, 95, 3269–3274. [Google Scholar] [CrossRef] [Green Version]
- Jahanshahi, M. Neuropsychological and Neuropsychiatric Features of Idiopathic and DYT1 Dystonia and the Impact of Medical and Surgical treatment. Arch. Clin. Neuropsychol. 2017, 32, 888–905. [Google Scholar] [CrossRef]
- Pekmezovic, T.; Svetel, M.; Ivanovic, N.; Dragasevic, N.; Petrovic, I.; Tepavcevic, D.K.; Kostic, V.S. Quality of life in patients with focal dystonia. Clin. Neurol. Neurosurg. 2009, 111, 161–164. [Google Scholar] [CrossRef]
- Soeder, A.; Kluger, B.M.; Okun, M.S.; Garvan, C.W.; Soeder, T.; Jacobson, C.E.; Rodriguez, R.L.; Turner, R.; Fernandez, H.H. Mood and energy determinants of quality of life in dystonia. J. Neurol. 2009, 256, 996–1001. [Google Scholar] [CrossRef]
- Lobbezoo, F.; Thu Thon, M.; Rémillard, G.; Montplaisir, J.Y.; Lavigne, G.J. Relationship between sleep, neck muscle activity, and pain in cervical dystonia. Can. J. Neurol. Sci. 1996, 23, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Tinazzi, M.; Squintani, G.M.; Bhatia, K.P.; Segatti, A.; Donato, F.; Valeriani, M.; Erro, R. Pain in cervical dystonia: Evidence of abnormal inhibitory control. Parkinsonism Relat. Disord. 2019, 65, 252–255. [Google Scholar] [CrossRef]
- Fung, V.S.C.; Jinnah, H.A.; Bhatia, K.; Vidailhet, M. Assessment of patients with isolated or combined dystonia: An update on dystonia syndromes. Mov. Disord. 2013, 28, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Kumar, K.R.; Watson, E.; Ahmad, K.; Sue, C.M.; Hayes, M.W. Dystonia Responsive to Dopamine: POLG Mutations Should Be Considered If Sensory Neuropathy Is Present. J. Mov. Disord. 2021, 14, 157–160. [Google Scholar] [CrossRef]
- Scuteri, D.; Corasaniti, M.T.; Tonin, P.; Bagetta, G. Eptinezumab for the treatment of migraine. Drugs Today 2019, 55, 695–703. [Google Scholar] [CrossRef]
- Patel, A.T.; Lew, M.F.; Dashtipour, K.; Isaacson, S.; Hauser, R.A.; Ondo, W.; Maisonobe, P.; Wietek, S.; Rubin, B.; Brashear, A. Sustained functional benefits after a single set of injections with abobotulinumtoxinA using a 2-mL injection volume in adults with cervical dystonia: 12-week results from a randomized, double-blind, placebo-controlled phase 3b study. PLoS ONE 2021, 16, e0245827. [Google Scholar] [CrossRef]
- Van Rijn, M.A.; Marinus, J.; Putter, H.; van Hilten, J.J. Onset and progression of dystonia in complex regional pain syndrome. Pain 2007, 130, 287–293. [Google Scholar] [CrossRef]
- Gosso, M.F.; de Rooij, A.M.; Alsina-Sanchis, E.; Kamphorst, J.T.; Marinus, J.; van Hilten, J.J.; van den Maagdenberg, A.M. Systematic mutation analysis of seven dystonia genes in complex regional pain syndrome with fixed dystonia. J. Neurol. 2010, 257, 820–824. [Google Scholar] [CrossRef]
- Berman, B.D.; Junker, J.; Shelton, E.; Sillau, S.H.; Jinnah, H.A.; Perlmutter, J.S.; Espay, A.J.; Jankovic, J.; Vidailhet, M.; Bonnet, C.; et al. Psychiatric associations of adult-onset focal dystonia phenotypes. J. Neurol. Neurosurg. Psychiatry 2017, 88, 595–602. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Rombola, L.; Morrone, L.A.; Bagetta, G.; Sakurada, S.; Sakurada, T.; Tonin, P.; Corasaniti, M.T. Neuropharmacology of the Neuropsychiatric Symptoms of Dementia and Role of Pain: Essential Oil of Bergamot as a Novel Therapeutic Approach. Int. J. Mol. Sci. 2019, 20, 3327. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Rombola, L.; Tridico, L.; Mizoguchi, H.; Watanabe, C.; Sakurada, T.; Sakurada, S.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Neuropharmacological Properties of the Essential Oil of Bergamot for the Clinical Management of Pain-Related BPSDs. Curr. Med. Chem. 2019, 26, 3764–3774. [Google Scholar] [CrossRef]
- Scuteri, D.; Morrone, L.A.; Rombola, L.; Avato, P.R.; Bilia, A.R.; Corasaniti, M.T.; Sakurada, S.; Sakurada, T.; Bagetta, G. Aromatherapy and Aromatic Plants for the Treatment of Behavioural and Psychological Symptoms of Dementia in Patients with Alzheimer’s Disease: Clinical Evidence and Possible Mechanisms. Evid. Based Complementary Altern. Med. 2017, 2017, 9416305. [Google Scholar] [CrossRef] [Green Version]
- Hewett, J.W.; Zeng, J.; Niland, B.P.; Bragg, D.C.; Breakefield, X.O. Dystonia-causing mutant torsinA inhibits cell adhesion and neurite extension through interference with cytoskeletal dynamics. Neurobiol. Dis. 2006, 22, 98–111. [Google Scholar] [CrossRef]
- Nery, F.C.; Armata, I.A.; Farley, J.E.; Cho, J.A.; Yaqub, U.; Chen, P.; da Hora, C.C.; Wang, Q.; Tagaya, M.; Klein, C.; et al. TorsinA participates in endoplasmic reticulum-associated degradation. Nat. Commun. 2011, 2, 393. [Google Scholar] [CrossRef] [Green Version]
- Kakazu, Y.; Koh, J.Y.; Iwabuchi, S.; Gonzalez-Alegre, P.; Harata, N.C. Miniature release events of glutamate from hippocampal neurons are influenced by the dystonia-associated protein torsinA. Synapse 2012, 66, 807–822. [Google Scholar] [CrossRef]
- Puglisi, F.; Vanni, V.; Ponterio, G.; Tassone, A.; Sciamanna, G.; Bonsi, P.; Pisani, A.; Mandolesi, G. Torsin A Localization in the Mouse Cerebellar Synaptic Circuitry. PLoS ONE 2013, 8, e68063. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, H.C.; Liu, Z.; Zacchi, L.F.; Brodsky, J.L.; Zolkiewski, M. Intracellular complexes of the early-onset torsion dystonia-associated AAA+ ATPase TorsinA. SpringerPlus 2014, 3, 743. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Brown, R.S.; Chase, A.R.; Eisele, M.R.; Schlieker, C. Regulation of Torsin ATPases by LAP1 and LULL1. Proc. Natl. Acad. Sci. USA 2013, 110, E1545–E1554. [Google Scholar] [CrossRef] [Green Version]
- Foisner, R.; Gerace, L. Integral membrane proteins of the nuclear envelope interact with lamins and chromosomes, and binding is modulated by mitotic phosphorylation. Cell 1993, 73, 1267–1279. [Google Scholar] [CrossRef]
- Goodchild, R.E.; Dauer, W.T. The AAA+ protein torsinA interacts with a conserved domain present in LAP1 and a novel ER protein. J. Cell Biol. 2005, 168, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Goodchild, R.E.; Buchwalter, A.L.; Naismith, T.V.; Holbrook, K.; Billion, K.; Dauer, W.T.; Liang, C.C.; Dear, M.L.; Hanson, P.I. Access of torsinA to the inner nuclear membrane is activity dependent and regulated in the endoplasmic reticulum. J. Cell Sci. 2015, 128, 2854–2865. [Google Scholar] [CrossRef] [Green Version]
- Laudermilch, E.; Schlieker, C. Torsin ATPases: Structural insights and functional perspectives. Curr. Opin. Cell Biol. 2016, 40, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Vander Heyden, A.B.; Naismith, T.V.; Snapp, E.L.; Hodzic, D.; Hanson, P.I. LULL1 retargets TorsinA to the nuclear envelope revealing an activity that is impaired by the DYT1 dystonia mutation. Mol. Biol. Cell 2009, 20, 2661–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nery, F.C.; Zeng, J.; Niland, B.P.; Hewett, J.; Farley, J.; Irimia, D.; Li, Y.; Wiche, G.; Sonnenberg, A.; Breakefield, X.O. TorsinA binds the KASH domain of nesprins and participates in linkage between nuclear envelope and cytoskeleton. J. Cell Sci. 2008, 121, 3476–3486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanGompel, M.J.; Nguyen, K.C.; Hall, D.H.; Dauer, W.T.; Rose, L.S. A novel function for the Caenorhabditis elegans torsin OOC-5 in nucleoporin localization and nuclear import. Mol. Biol. Cell 2015, 26, 1752–1763. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.A.; Harris, N.J.; Willey, P.T.; Woolums, B.M.; Wang, Y.; McQuown, A.J.; Schoenhofen, A.; Worman, H.J.; Dauer, W.T.; Gundersen, G.G.; et al. TorsinA controls TAN line assembly and the retrograde flow of dorsal perinuclear actin cables during rearward nuclear movement. J. Cell Biol. 2017, 216, 657–674. [Google Scholar] [CrossRef] [Green Version]
- Beauvais, G.; Rodriguez-Losada, N.; Ying, L.; Zakirova, Z.; Watson, J.L.; Readhead, B.; Gadue, P.; French, D.L.; Ehrlich, M.E.; Gonzalez-Alegre, P. Exploring the Interaction Between eIF2α Dysregulation, Acute Endoplasmic Reticulum Stress and DYT1 Dystonia in the Mammalian Brain. Neuroscience 2018, 371, 455–468. [Google Scholar] [CrossRef]
- Maltese, M.; Stanic, J.; Tassone, A.; Sciamanna, G.; Ponterio, G.; Vanni, V.; Martella, G.; Imbriani, P.; Bonsi, P.; Mercuri, N.B.; et al. Early structural and functional plasticity alterations in a susceptibility period of DYT1 dystonia mouse striatum. eLife 2018, 7, e33331. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Ni, L.; Hu, X.; Ding, X. Inhibition of endoplasmic reticulum stress reverses synaptic plasticity deficits in striatum of DYT1 dystonia mice. Aging 2021, 13, 20319–20334. [Google Scholar] [CrossRef]
- Sciamanna, G.; Bonsi, P.; Tassone, A.; Cuomo, D.; Tscherter, A.; Viscomi, M.T.; Martella, G.; Sharma, N.; Bernardi, G.; Standaert, D.G.; et al. Impaired striatal D2 receptor function leads to enhanced GABA transmission in a mouse model of DYT1 dystonia. Neurobiol. Dis. 2009, 34, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Ponterio, G.; Tassone, A.; Sciamanna, G.; Vanni, V.; Meringolo, M.; Santoro, M.; Mercuri, N.B.; Bonsi, P.; Pisani, A. Enhanced mu opioid receptor-dependent opioidergic modulation of striatal cholinergic transmission in DYT1 dystonia. Mov. Disord. 2018, 33, 310–320. [Google Scholar] [CrossRef]
- Christoph, T.; Schröder, W.; Tallarida, R.J.; De Vry, J.; Tzschentke, T.M. Spinal-supraspinal and intrinsic μ-opioid receptor agonist-norepinephrine reuptake inhibitor (MOR-NRI) synergy of tapentadol in diabetic heat hyperalgesia in mice. J. Pharmacol. Exp. Ther. 2013, 347, 794–801. [Google Scholar] [CrossRef] [Green Version]
- Christoph, T.; De Vry, J.; Tzschentke, T.M. Tapentadol, but not morphine, selectively inhibits disease-related thermal hyperalgesia in a mouse model of diabetic neuropathic pain. Neurosci. Lett. 2010, 470, 91–94. [Google Scholar] [CrossRef]
- Khangura, R.K.; Bali, A.; Kaur, G.; Singh, N.; Jaggi, A.S. Neuropathic pain attenuating effects of perampanel in an experimental model of chronic constriction injury in rats. Biomed. Pharmacother. 2017, 94, 557–563. [Google Scholar] [CrossRef]
- Vanni, V.; Puglisi, F.; Bonsi, P.; Ponterio, G.; Maltese, M.; Pisani, A.; Mandolesi, G. Cerebellar synaptogenesis is compromised in mouse models of DYT1 dystonia. Exp. Neurol. 2015, 271, 457–467. [Google Scholar] [CrossRef]
- Pisani, A.; Martella, G.; Tscherter, A.; Bonsi, P.; Sharma, N.; Bernardi, G.; Standaert, D.G. Altered responses to dopaminergic D2 receptor activation and N-type calcium currents in striatal cholinergic interneurons in a mouse model of DYT1 dystonia. Neurobiol. Dis. 2006, 24, 318–325. [Google Scholar] [CrossRef]
- Zhang, L.; McCarthy, D.M.; Sharma, N.; Bhide, P.G. Dopamine receptor and Galpha(olf) expression in DYT1 dystonia mouse models during postnatal development. PLoS ONE 2015, 10, e0123104. [Google Scholar] [CrossRef]
- D’Angelo, V.; Castelli, V.; Giorgi, M.; Cardarelli, S.; Saverioni, I.; Palumbo, F.; Bonsi, P.; Pisani, A.; Giampa, C.; Sorge, R.; et al. Phosphodiesterase-10A Inverse Changes in Striatopallidal and Striatoentopeduncular Pathways of a Transgenic Mouse Model of DYT1 Dystonia. J. Neurosci. 2017, 37, 2112–2124. [Google Scholar] [CrossRef] [Green Version]
- Bonsi, P.; Ponterio, G.; Vanni, V.; Tassone, A.; Sciamanna, G.; Migliarini, S.; Martella, G.; Meringolo, M.; Dehay, B.; Doudnikoff, E.; et al. RGS9-2 rescues dopamine D2 receptor levels and signaling in DYT1 dystonia mouse models. EMBO Mol. Med. 2019, 11, e9283. [Google Scholar] [CrossRef]
- D’Angelo, V.; Paldino, E.; Cardarelli, S.; Sorge, R.; Fusco, F.R.; Biagioni, S.; Mercuri, N.B.; Giorgi, M.; Sancesario, G. Dystonia: Sparse Synapses for D2 Receptors in Striatum of a DYT1 Knock-out Mouse Model. Int. J. Mol. Sci. 2020, 21, 1073. [Google Scholar] [CrossRef] [Green Version]
- Sciamanna, G.; Tassone, A.; Martella, G.; Mandolesi, G.; Puglisi, F.; Cuomo, D.; Madeo, G.; Ponterio, G.; Standaert, D.G.; Bonsi, P.; et al. Developmental profile of the aberrant dopamine D2 receptor response in striatal cholinergic interneurons in DYT1 dystonia. PLoS ONE 2011, 6, e24261. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, F.; Pasqualetti, M.; Usiello, A.; Santini, E.; Pacini, G.; Sciamanna, G.; Errico, F.; Tassone, A.; Di Dato, V.; Martella, G.; et al. Dopamine D2 receptor dysfunction is rescued by adenosine A2A receptor antagonism in a model of DYT1 dystonia. Neurobiol. Dis. 2010, 38, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Balcioglu, A.; Kim, M.O.; Sharma, N.; Cha, J.H.; Breakefield, X.O.; Standaert, D.G. Dopamine release is impaired in a mouse model of DYT1 dystonia. J. Neurochem. 2007, 102, 783–788. [Google Scholar] [CrossRef]
- Downs, A.M.; Fan, X.; Kadakia, R.F.; Donsante, Y.; Jinnah, H.A.; Hess, E.J. Cell-intrinsic effects of TorsinA(ΔE) disrupt dopamine release in a mouse model of TOR1A dystonia. Neurobiol. Dis. 2021, 155, 105369. [Google Scholar] [CrossRef]
- Martella, G.; Bonsi, P.; Imbriani, P.; Sciamanna, G.; Nguyen, H.; Yu-Taeger, L.; Schneider, M.; Poli, S.M.; Lütjens, R.; Pisani, A. Rescue of striatal long-term depression by chronic mGlu5 receptor negative allosteric modulation in distinct dystonia models. Neuropharmacology 2021, 192, 108608. [Google Scholar] [CrossRef]
- Ozelius, L.J.; Hewett, J.W.; Page, C.E.; Bressman, S.B.; Kramer, P.L.; Shalish, C.; de Leon, D.; Brin, M.F.; Raymond, D.; Corey, D.P.; et al. The early-onset torsion dystonia gene (DYT1) encodes an ATP-binding protein. Nat. Genet. 1997, 17, 40–48. [Google Scholar] [CrossRef]
- Ozelius, L.J.; Hewett, J.W.; Page, C.E.; Bressman, S.B.; Kramer, P.L.; Shalish, C.; de Leon, D.; Brin, M.F.; Raymond, D.; Jacoby, D.; et al. The gene (DYT1) for early-onset torsion dystonia encodes a novel protein related to the Clp protease/heat shock family. Adv. Neurol. 1998, 78, 93–105. [Google Scholar]
- Sharma, N.; Baxter, M.G.; Petravicz, J.; Bragg, D.C.; Schienda, A.; Standaert, D.G.; Breakefield, X.O. Impaired motor learning in mice expressing torsinA with the DYT1 dystonia mutation. J. Neurosci. 2005, 25, 5351–5355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiarini, L.; Sathasivam, K.; Seller, M.; Cozens, B.; Harper, A.; Hetherington, C.; Lawton, M.; Trottier, Y.; Lehrach, H.; Davies, S.W.; et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell 1996, 87, 493–506. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, F.; Chen, H.-X.; Oleas, J.; Dang, M.T.; Xing, H.; Dexter, K.M.; Li, Y. Characterization of the direct pathway in Dyt1 ΔGAG heterozygous knock-in mice and dopamine receptor 1-expressing-cell-specific Dyt1 conditional knockout mice. Behav. Brain Res. 2021, 411, 113381. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, V.; Giorgi, M.; Paldino, E.; Cardarelli, S.; Fusco, F.R.; Saverioni, I.; Sorge, R.; Martella, G.; Biagioni, S.; Mercuri, N.B.; et al. A2A Receptor Dysregulation in Dystonia DYT1 Knock-Out Mice. Int. J. Mol. Sci. 2021, 22, 2691. [Google Scholar] [CrossRef] [PubMed]
- Ungermann, C.; Sato, K.; Wickner, W. Defining the functions of trans-SNARE pairs. Nature 1998, 396, 543–548. [Google Scholar] [CrossRef]
- Ponterio, G.; Faustini, G.; El Atiallah, I.; Sciamanna, G.; Meringolo, M.; Tassone, A.; Imbriani, P.; Cerri, S.; Martella, G.; Bonsi, P.; et al. Alpha-Synuclein is Involved in DYT1 Dystonia Striatal Synaptic Dysfunction. Mov. Disord. 2022, 37, 949–961. [Google Scholar] [CrossRef]
- Lang, T.; Bruns, D.; Wenzel, D.; Riedel, D.; Holroyd, P.; Thiele, C.; Jahn, R. SNAREs are concentrated in cholesterol-dependent clusters that define docking and fusion sites for exocytosis. EMBO J. 2001, 20, 2202–2213. [Google Scholar] [CrossRef] [Green Version]
- Suh, Y.H.; Terashima, A.; Petralia, R.S.; Wenthold, R.J.; Isaac, J.T.; Roche, K.W.; Roche, P.A. A neuronal role for SNAP-23 in postsynaptic glutamate receptor trafficking. Nat. Neurosci. 2010, 13, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, F.; Oleas, J.; Xing, H.; Liu, Y.; Dexter, K.M.; Misztal, C.; Gerard, M.; Efimenko, I.; Lynch, P.; Villanueva, M.; et al. Decreased number of striatal cholinergic interneurons and motor deficits in dopamine receptor 2-expressing-cell-specific Dyt1 conditional knockout mice. Neurobiol. Dis. 2020, 134, 104638. [Google Scholar] [CrossRef]
- Wilkes, B.J.; DeSimone, J.C.; Liu, Y.; Chu, W.T.; Coombes, S.A.; Li, Y.; Vaillancourt, D.E. Cell-specific effects of Dyt1 knock-out on sensory processing, network-level connectivity, and motor deficits. Exp. Neurol. 2021, 343, 113783. [Google Scholar] [CrossRef]
- Yu-Taeger, L.; Ott, T.; Bonsi, P.; Tomczak, C.; Wassouf, Z.; Martella, G.; Sciamanna, G.; Imbriani, P.; Ponterio, G.; Tassone, A.; et al. Impaired dopamine- and adenosine-mediated signaling and plasticity in a novel rodent model for DYT25 dystonia. Neurobiol. Dis. 2020, 134, 104634. [Google Scholar] [CrossRef]
- Bogush, A.I.; McCarthy, L.E.; Tian, C.; Olm, V.; Gieringer, T.; Ivkovic, S.; Ehrlich, M.E. DARPP-32 genomic fragments drive Cre expression in postnatal striatum. Genesis 2005, 42, 37–46. [Google Scholar] [CrossRef]
- Gonzalez-Alegre, P.; Beauvais, G.; Martin, J.; Koch, R.J.; Walker, R.H.; Patel, J.C.; Rice, M.E.; Ehrlich, M.E. A Novel Transgenic Mouse Model to Investigate the Cell-Autonomous Effects of torsinA(ΔE) Expression in Striatal Output Neurons. Neuroscience 2019, 422, 1–11. [Google Scholar] [CrossRef]
- Liu, Y.; Xing, H.; Sheng, W.; Singh, K.N.; Korkmaz, A.G.; Comeau, C.; Anika, M.; Ernst, A.; Yokoi, F.; Vaillancourt, D.E.; et al. Alteration of the cholinergic system and motor deficits in cholinergic neuron-specific Dyt1 knockout mice. Neurobiol. Dis. 2021, 154, 105342. [Google Scholar] [CrossRef]
- Downs, A.M.; Donsante, Y.; Jinnah, H.A.; Hess, E.J. Blockade of M4 muscarinic receptors on striatal cholinergic interneurons normalizes striatal dopamine release in a mouse model of TOR1A dystonia. Neurobiol. Dis. 2022, 168, 105699. [Google Scholar] [CrossRef]
- Cascalho, A.; Foroozandeh, J.; Hennebel, L.; Swerts, J.; Klein, C.; Rous, S.; Dominguez Gonzalez, B.; Pisani, A.; Meringolo, M.; Gallego, S.F.; et al. Excess Lipin enzyme activity contributes to TOR1A recessive disease and DYT-TOR1A dystonia. Brain J. Neurol. 2020, 143, 1746–1765. [Google Scholar] [CrossRef]
- Pappas, S.S.; Liang, C.C.; Kim, S.; Rivera, C.O.; Dauer, W.T. TorsinA dysfunction causes persistent neuronal nuclear pore defects. Hum. Mol. Genet. 2018, 27, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Washburn, S.; Fremont, R.; Moreno-Escobar, M.C.; Angueyra, C.; Khodakhah, K. Acute cerebellar knockdown of Sgce reproduces salient features of myoclonus-dystonia (DYT11) in mice. eLife 2019, 8, e52101. [Google Scholar] [CrossRef]
- Li, J.; Liang, C.-C.; Pappas, S.S.; Dauer, W.T. TorsinB overexpression prevents abnormal twisting in DYT1 dystonia mouse models. eLife 2020, 9, e54285. [Google Scholar] [CrossRef]
- Frederick, N.M.; Pooler, M.M.; Shah, P.; Didonna, A.; Opal, P. Pharmacological perturbation reveals deficits in D2 receptor responses in Thap1 null mice. Ann. Clin. Transl. Neurol. 2021, 8, 2302–2308. [Google Scholar] [CrossRef]
- Zakirova, Z.; Fanutza, T.; Bonet, J.; Readhead, B.; Zhang, W.; Yi, Z.; Beauvais, G.; Zwaka, T.P.; Ozelius, L.J.; Blitzer, R.D.; et al. Mutations in THAP1/DYT6 reveal that diverse dystonia genes disrupt similar neuronal pathways and functions. PLoS Genet. 2018, 14, e1007169. [Google Scholar] [CrossRef]
- Sato, K.; Sumi-Ichinose, C.; Kaji, R.; Ikemoto, K.; Nomura, T.; Nagatsu, I.; Ichinose, H.; Ito, M.; Sako, W.; Nagahiro, S.; et al. Differential involvement of striosome and matrix dopamine systems in a transgenic model of dopa-responsive dystonia. Proc. Natl. Acad. Sci. USA 2008, 105, 12551–12556. [Google Scholar] [CrossRef] [Green Version]
- DeAndrade, M.P.; Yokoi, F.; van Groen, T.; Lingrel, J.B.; Li, Y. Characterization of Atp1a3 mutant mice as a model of rapid-onset dystonia with parkinsonism. Behav. Brain Res. 2011, 216, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Ledoux, M.S.; Dauer, W.T.; Warner, T.T. Emerging common molecular pathways for primary dystonia. Mov. Disord. 2013, 28, 968–981. [Google Scholar] [CrossRef] [Green Version]
- Quesada, C.; Kostenko, A.; Ho, I.; Leone, C.; Nochi, Z.; Stouffs, A.; Wittayer, M.; Caspani, O.; Finnerup, N.B.; Mouraux, A.; et al. Human surrogate models of central sensitization: A critical review and practical guide. Eur. J. Pain 2021, 25, 1389–1428. [Google Scholar] [CrossRef]
- Sharpless, N.E.; Depinho, R.A. The mighty mouse: Genetically engineered mouse models in cancer drug development. Nat. Rev. Drug Discov. 2006, 5, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.I.; Decker, S.; Zaharevitz, D.; Rubinstein, L.V.; Venditti, J.M.; Schepartz, S.; Kalyandrug, S.; Christian, M.; Arbuck, S.; Hollingshead, M.; et al. Relationships between drug activity in NCI preclinical in vitro and in vivo models and early clinical trials. Br. J. Cancer 2001, 84, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.S.C.; Morland, R.; Huang, W.; Currie, G.L.; Sena, E.S.; Macleod, M.R. Transparency in the reporting of in vivo pre-clinical pain research: The relevance and implications of the ARRIVE (Animal Research: Reporting In Vivo Experiments) guidelines. Scand. J. Pain 2013, 4, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Hamamura, K.; Sakurada, T.; Watanabe, C.; Sakurada, S.; Morrone, L.A.; Rombolà, L.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Efficacy of Essential Oils in Pain: A Systematic Review and Meta-Analysis of Preclinical Evidence. Front. Pharmacol. 2021, 12, 640128. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- Macleod, M.R.; O’Collins, T.; Howells, D.W.; Donnan, G.A. Pooling of Animal Experimental Data Reveals Influence of Study Design and Publication Bias. Stroke 2004, 35, 1203–1208. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ Clin. Res. Ed. 2021, 372, n71. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Savović, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.-Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A revised tool for assessing risk of bias in randomised trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [Green Version]
- Sterne, J.A.; Hernán, M.A.; Reeves, B.C.; Savović, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef] [Green Version]
- Mogil, J.S.; Grisel, J.E. Transgenic studies of pain. Pain 1998, 77, 107–128. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bráz, J.M. Transgenic Mouse Models for the Tracing of “Pain” Pathways. In Translational Pain Research: From Mouse to Man; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- McLeod, A.L.; Ritchie, J.; Cuello, A.C.; Julien, J.P.; Ribeiro-Da-Silva, A.; Henry, J.L. Transgenic mice over-expressing substance P exhibit allodynia and hyperalgesia which are reversed by substance P and N-methyl-D-aspartate receptor antagonists. Neuroscience 1999, 89, 891–899. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Davis, B.M.; Lewin, G.R.; Mendell, L.M.; Jones, M.E.; Albers, K.M. Altered expression of nerve growth factor in the skin of transgenic mice leads to changes in response to mechanical stimuli. Neuroscience 1993, 56, 789–792. [Google Scholar] [CrossRef]
- Aman, Y.; Pitcher, T.; Simeoli, R.; Ballard, C.; Malcangio, M. Reduced thermal sensitivity and increased opioidergic tone in the TASTPM mouse model of Alzheimer’s disease. Pain 2016, 157, 2285–2296. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Berliocchi, L.; Rombolà, L.; Morrone, L.A.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Effects of Aging on Formalin-Induced Pain Behavior and Analgesic Activity of Gabapentin in C57BL/6 Mice. Front. Pharmacol. 2020, 11, 663. [Google Scholar] [CrossRef]
- Hamm, R.J.; Knisely, J.S. Environmentally Induced Analgesia: An Age-Related Decline in an Endogenous Opioid System. J. Gerontol. 1985, 40, 268–274. [Google Scholar] [CrossRef]
- Hamm, R.J.; Knisely, J.S. Environmentally induced analgesia: Age-related decline in a neurally mediated, nonopioid system. Psychol. Aging 1986, 1, 195–201. [Google Scholar] [CrossRef]
- Jourdan, D.; Boghossian, S.; Alloui, A.; Veyrat-Durebex, C.; Coudore, M.A.; Eschalier, A.; Alliot, J. Age-related changes in nociception and effect of morphine in the Lou rat. Eur. J. Pain 2000, 4, 291–300. [Google Scholar] [CrossRef]
- Jourdan, D.; Pickering, G.; Marchand, F.; Gaulier, J.-M.; Alliot, J.; Eschalier, A. Impact of ageing on the antinociceptive effect of reference analgesics in the Lou/c rat. Br. J. Pharmacol. 2002, 137, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Bedrosian, T.A.; Herring, K.L.; Weil, Z.M.; Nelson, R.J. Altered temporal patterns of anxiety in aged and amyloid precursor protein (APP) transgenic mice. Proc. Natl. Acad. Sci. USA 2011, 108, 11686–11691. [Google Scholar] [CrossRef] [Green Version]
- Vigli, D.; Cosentino, L.; Pellas, M.; De Filippis, B. Chronic Treatment with Cannabidiolic Acid (CBDA) Reduces Thermal Pain Sensitivity in Male Mice and Rescues the Hyperalgesia in a Mouse Model of Rett Syndrome. Neuroscience 2021, 453, 113–123. [Google Scholar] [CrossRef]
- De Filippis, B.; Fabbri, A.; Simone, D.; Canese, R.; Ricceri, L.; Malchiodi-Albedi, F.; Laviola, G.; Fiorentini, C. Modulation of RhoGTPases improves the behavioral phenotype and reverses astrocytic deficits in a mouse model of Rett syndrome. Neuropsychopharmacol 2012, 37, 1152–1163. [Google Scholar] [CrossRef]
- Zimmer, A.; Zimmer, A.M.; Baffi, J.; Usdin, T.; Reynolds, K.; König, M.; Palkovits, M.; Mezey, E. Hypoalgesia in mice with a targeted deletion of the tachykinin 1 gene. Proc. Natl. Acad. Sci. USA 1998, 95, 2630–2635. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R.; Baños, J.E.; Cabañero, D. Usefulness of knockout mice to clarify the role of the opioid system in chronic pain. Br. J. Pharmacol. 2018, 175, 2791–2808. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Mantovani, E.; Tamburin, S.; Sandrini, G.; Corasaniti, M.T.; Bagetta, G.; Tonin, P. Opioids in Post-stroke Pain: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2020, 11, 587050. [Google Scholar] [CrossRef]
- Hain, H.S.; Belknap, J.K.; Mogil, J.S. Pharmacogenetic evidence for the involvement of 5-hydroxytryptamine (serotonin)-1B receptors in the mediation of morphine antinociceptive sensitivity. J. Pharmacol. Exp. Ther. 1999, 291, 444–449. [Google Scholar] [PubMed]
- Scuteri, D.; Rombolà, L.; Natoli, S.; Pisani, A.; Bonsi, P.; Hamamura, K.; Bagetta, G.; Tonin, P.; Corasaniti, M.T. Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia. Life 2021, 11, 985. [Google Scholar] [CrossRef] [PubMed]
- Alles, S.R.A.; Smith, P.A. Etiology and Pharmacology of Neuropathic Pain. Pharmacol. Rev. 2018, 70, 315–347. [Google Scholar] [CrossRef] [PubMed]
- Goodchild, R.E.; Kim, C.E.; Dauer, W.T. Loss of the dystonia-associated protein torsinA selectively disrupts the neuronal nuclear envelope. Neuron 2005, 48, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Coull, J.A.; Beggs, S.; Boudreau, D.; Boivin, D.; Tsuda, M.; Inoue, K.; Gravel, C.; Salter, M.W.; De Koninck, Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 2005, 438, 1017–1021. [Google Scholar] [CrossRef]
- Dolphin, A.C. Calcium channel auxiliary alpha2delta and beta subunits: Trafficking and one step beyond. Nat. Rev. Neurosci. 2012, 13, 542–555. [Google Scholar] [CrossRef]
- Dolphin, A.C. Voltage-gated calcium channels and their auxiliary subunits: Physiology and pathophysiology and pharmacology. J. Physiol. 2016, 594, 5369–5390. [Google Scholar] [CrossRef] [Green Version]
- Deeks, E.D. Mirogabalin: First Global Approval. Drugs 2019, 79, 463–468. [Google Scholar] [CrossRef]
- Baba, M.; Kuroha, M.; Ohwada, S.; Murayama, E.; Matsui, N. Results of Mirogabalin Treatment for Diabetic Peripheral Neuropathic Pain in Asian Subjects: A Phase 2, Double-Blind, Randomized, Placebo-Controlled, Study. Pain Ther. 2020, 9, 261–278. [Google Scholar] [CrossRef] [Green Version]
- Scuteri, D.; Garreffa, M.R.; Esposito, S.; Bagetta, G.; Naturale, M.D.; Corasaniti, M.T. Evidence for accuracy of pain assessment and painkillers utilization in neuropsychiatric symptoms of dementia in Calabria region, Italy. Neural Regen. Res. 2018, 13, 1619–1621. [Google Scholar] [CrossRef]
- Scuteri, D.; Piro, B.; Morrone, L.A.; Corasaniti, M.T.; Vulnera, M.; Bagetta, G. The need for better access to pain treatment: Learning from drug consumption trends in the USA. Funct. Neurol. 2017, 22, 229–230. [Google Scholar] [CrossRef]
- Scuteri, D.; Vulnera, M.; Piro, B.; Bruno Bossio, R.; Morrone, L.A.; Sandrini, G.; Tamburin, S.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Pattern of treatment of behavioral and psychological symptoms of dementia and pain: Evidence on pharmacoutilization from a large real-world sample and from a center for cognitive disturbances and dementia. Eur. J. Clin. Pharm. 2021, 77, 241–249. [Google Scholar] [CrossRef]
- Scuteri, D.; Sandrini, G.; Tamburin, S.; Corasaniti, M.T.; Nicotera, P.; Tonin, P.; Bagetta, G. Bergamot rehabilitation AgaINst agitation in dementia (BRAINAID): Study protocol for a randomized, double-blind, placebo-controlled trial to assess the efficacy of furocoumarin-free bergamot loaded in a nanotechnology-based delivery system of the essential oil in the treatment of agitation in elderly affected by severe dementia. Phytother. Res. 2021, 35, 5333–5338. [Google Scholar] [CrossRef]
- Scuteri, D.; Cassano, R.; Trombino, S.; Russo, R.; Mizoguchi, H.; Watanabe, C.; Hamamura, K.; Katsuyama, S.; Komatsu, T.; Morrone, L.A.; et al. Development and Translation of NanoBEO, a Nanotechnology-Based Delivery System of Bergamot Essential Oil Deprived of Furocumarins, in the Control of Agitation in Severe Dementia. Pharmaceutics 2021, 13, 379. [Google Scholar] [CrossRef]
- Rombolà, L.; Scuteri, D.; Watanabe, C.; Sakurada, S.; Hamamura, K.; Sakurada, T.; Tonin, P.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Role of 5-HT1A Receptor in the Anxiolytic-Relaxant Effects of Bergamot Essential Oil in Rodent. Int. J. Mol. Sci. 2020, 21, 2597. [Google Scholar] [CrossRef] [Green Version]
- Rombolà, L.; Scuteri, D.; Marilisa, S.; Watanabe, C.; Morrone, L.A.; Bagetta, G.; Corasaniti, M.T. Pharmacokinetic Interactions between Herbal Medicines and Drugs: Their Mechanisms and Clinical Relevance. Life 2020, 10, 106. [Google Scholar] [CrossRef]
- Treede, R.D.; Meyer, R.A.; Raja, S.N.; Campbell, J.N. Peripheral and central mechanisms of cutaneous hyperalgesia. Prog. Neurobiol. 1992, 38, 397–421. [Google Scholar] [CrossRef]
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Treede, R.D.; Magerl, W. Multiple mechanisms of secondary hyperalgesia. Prog. Brain Res. 2000, 129, 331–341. [Google Scholar] [CrossRef]
- Tassone, A.; Martella, G.; Meringolo, M.; Vanni, V.; Sciamanna, G.; Ponterio, G.; Imbriani, P.; Bonsi, P.; Pisani, A. Vesicular Acetylcholine Transporter Alters Cholinergic Tone and Synaptic Plasticity in DYT1 Dystonia. Mov. Disord. 2021, 36, 2768–2779. [Google Scholar] [CrossRef]
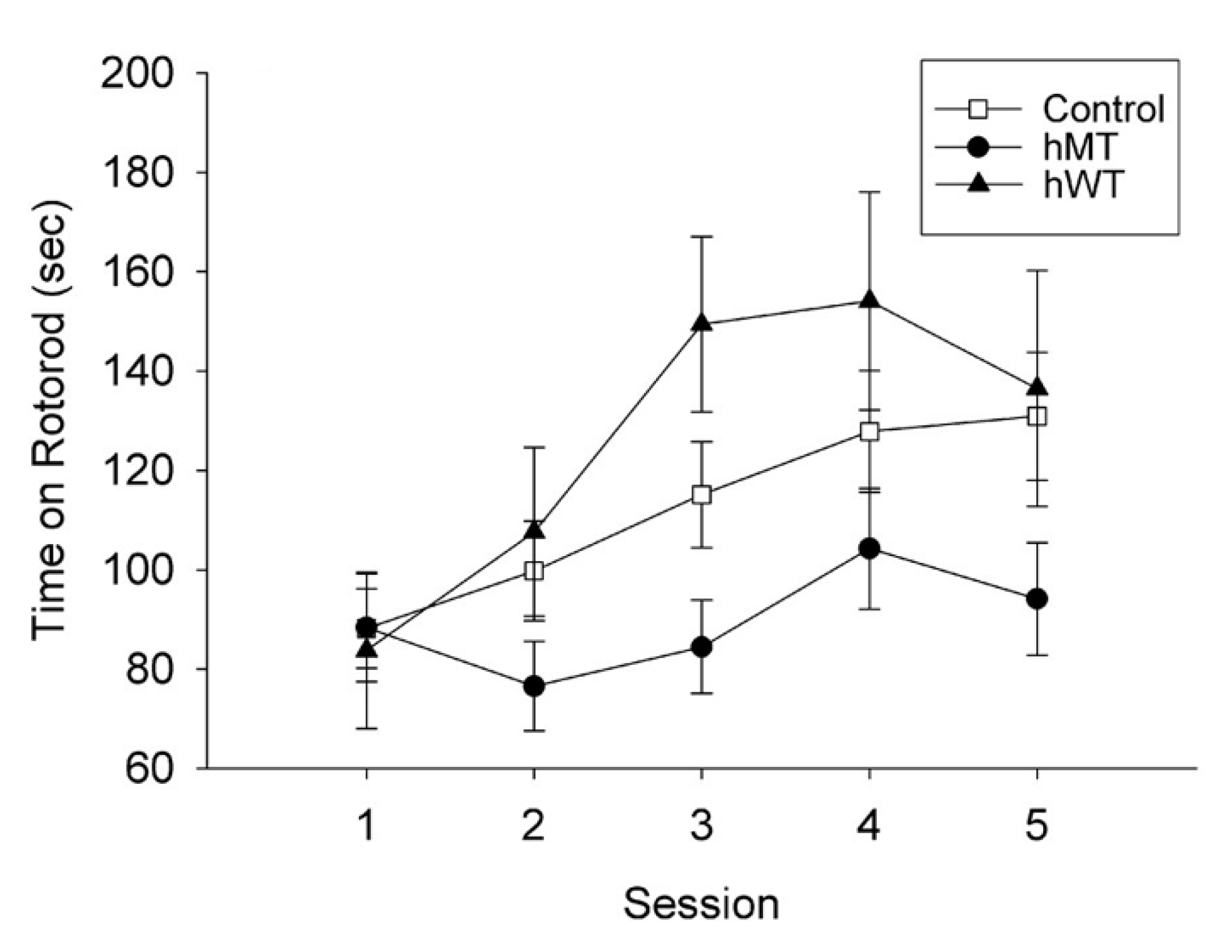

{kind=link}
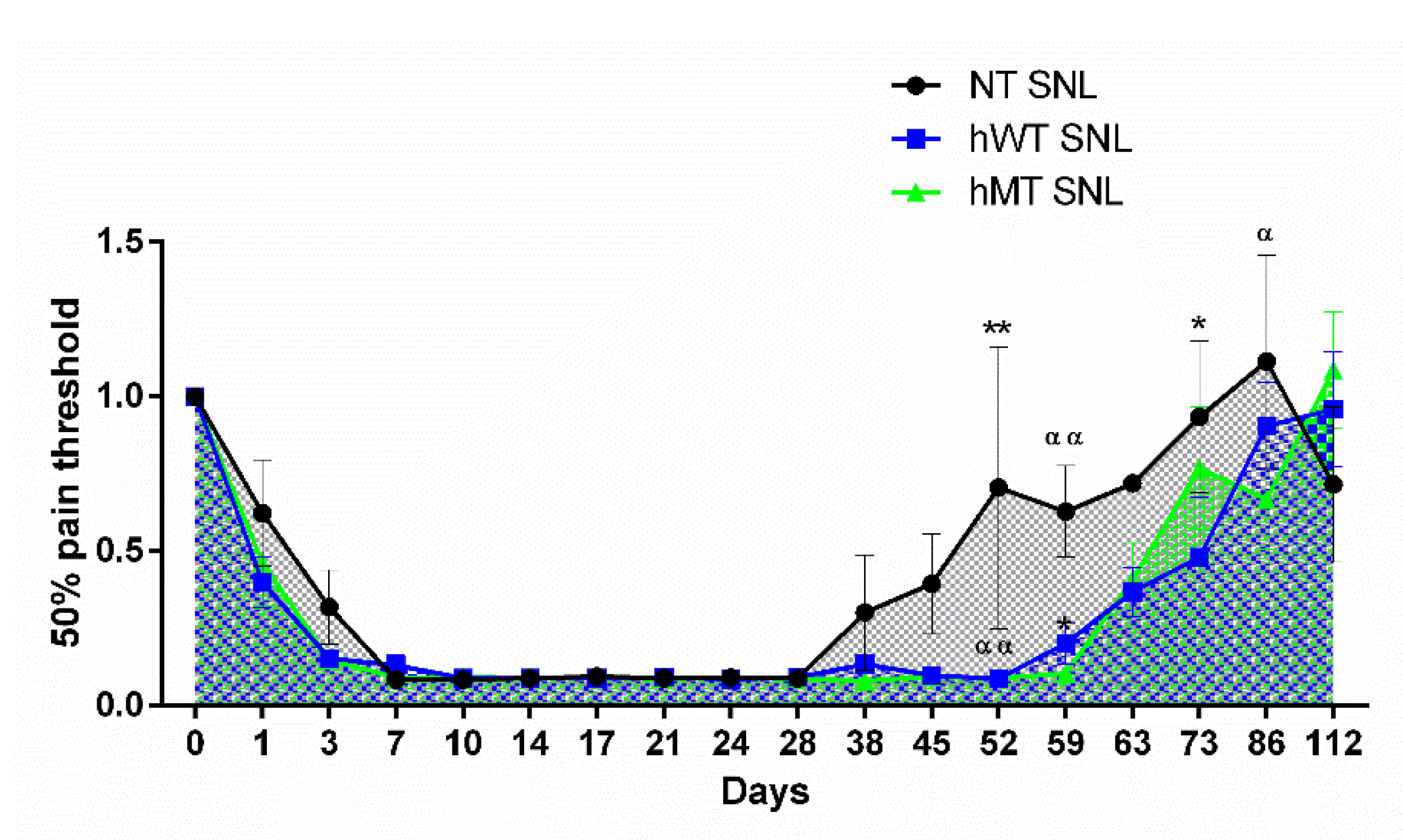
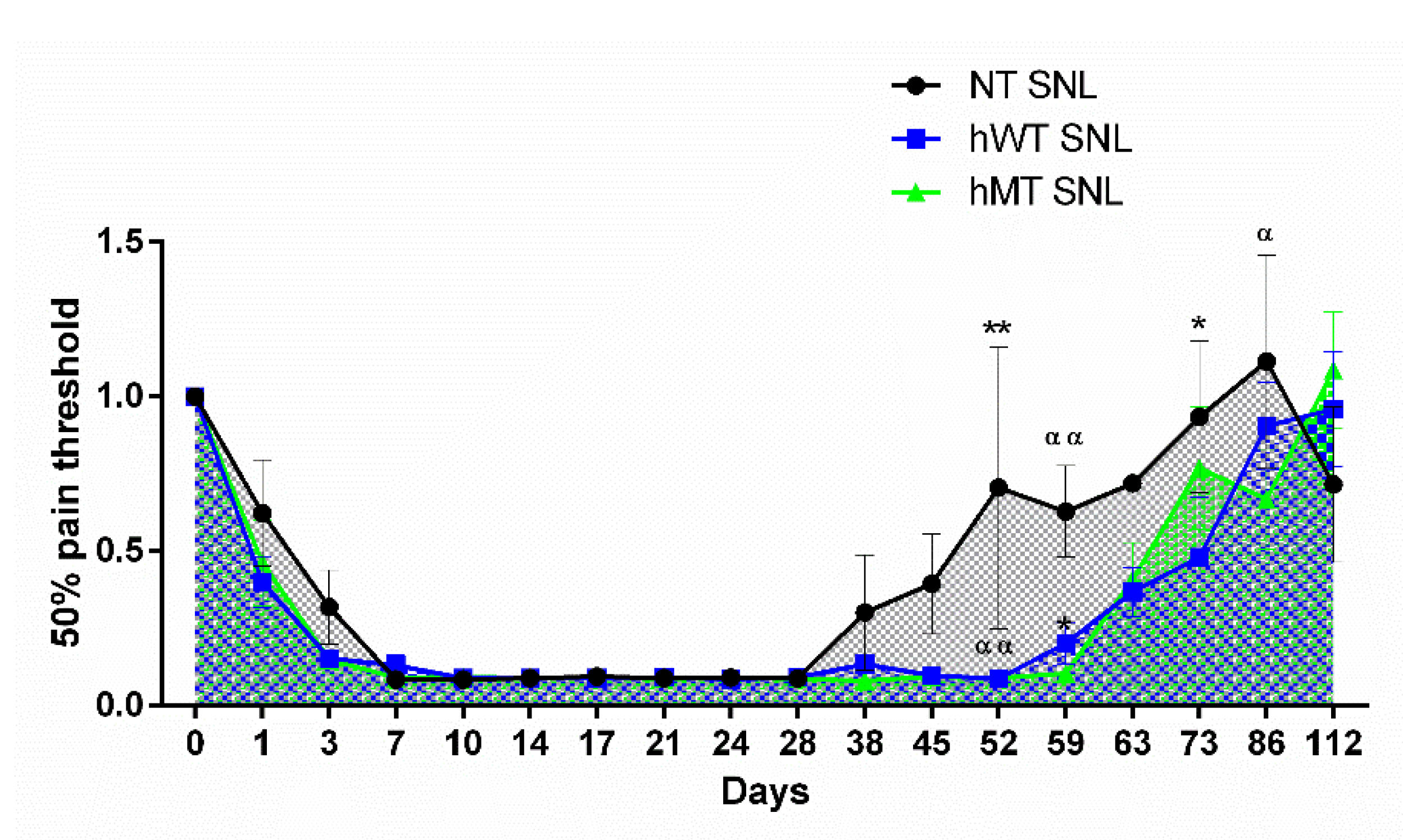{kind=link}
| Locus | Designation | Clinical Features | Gene/Inheritance | Protein | Putative Functions |
|---|---|---|---|---|---|
| Pure Dystonia | |||||
| DYT1 Chr9q34.11 | Early-onset primary dystonia | Childhood onset dystonia in limb with generalization | tor1A autosomal dominant | torsinA | AAA+ protein, nuclear envelope, endoplasmic reticulum secretory and stress response, regulation of synaptic function |
| DYT2 | Early-onset dystonia | Adolescent-onset segmental or generalized | Autosomal recessive | Unknown | |
| DYT4 Chr19p13.3 | Whispering dysphonia | Childhood-onset laryngeal abductor spasm with cervical dystonia | tubb4a autosomal dominant | beta-tubulin 4a | Structural cytoskeleton protein |
| DYT6 Chr8p11.21 | Autosomal dominant early-onset focal dystonia | Early-onset dystonia with prominent cervical and laryngeal involvement | thap1 autosomal dominant | Thanatos-associated domain-containing apoptosis associated protein 1 | Atypical zinc-finger protein; thap domain is chromatin-binding factor and regulates transcription |
| DYT7 Chr8p | Familial focal dystonia | Adult-onset focal dystonia | Unknown autosomal dominant | Unknown | |
| DYT13 Chr1p36.32– p36.13 | Familial craniocervical dystonia | Focal or segmental dystonia of the craniocervical region and upper limbs | Unknown autosomal dominant | Unknown | |
| DYT17 Chr20p11.2– 2q13.12 | Early-onset autosomal recessive dystonia | Early-onset focal dystonia progressing to generalized with dysphonia and dysarthria | Unknown autosomal recessive | Unknown | |
| DYT21 Chr2q14.3– q21.3 | Late-onset dystonia | Late-onset multifocal and generalized dystonia | Unknown autosomal dominant | Unknown | |
| DYT23 Chr9q34.11 | Cervical dystonia | Late-onset primary cervical dystonia | ciz1 autosomal dominant | cip1-interacting zinc finger protein 1 | Regulation of G1–S cell cycle and DNA replication |
| DYT24 Chr11p14.2 | Late-onset dystonia | Cranial and cervical dystonia | ano3 autosomal dominant | Anoctamin 3 | Calcium-gated chloride channel |
| DYT25 Chr18p | Cervical dystonia with local spread | Predominantly late-onset primary cervical dystonia with spread to face | gnal autosomal dominant | Alpha subunit of G protein | Probable interaction with D1 and adenosine 2A receptors. |
| Dystonia syndromes | |||||
| DYT3 Xq13.1 | X-linked dystonia (Lubag) | Segmental or generalized dystonia with parkinsonism | taf1 X-linked | TATA box-binding protein associated factor 1 | Regulation of transcription initiation and cell cycle |
| DYT5/14 Chr2q13.2 | DOPA (precursor of dopamine)-responsive dystonia | Dystonia with parkinsonism, diurnal variation, and very good response to L-dopa | gch1 autosomal dominant | GTP cyclohydrolase 1 | Rate-limiting enzyme in synthesis of tetrahydrobiopterin, key cofactor in monoamine synthesis; results in deficient dopamine synthesis |
| DYT11 Chr7q21.3 | Myoclonic dystonia syndrome | Upper body myoclonic jerks with dystonia; responsive to alcohol | sgce autosomal dominant | Epsilon-sarcoglycan | Cell membrane protein that may act as structural platform for other protein interactions |
| DYT12 Chr19q13.2 | Rapid-onset dystonia parkinsonism | Acute-onset generalized dystonia with parkinsonism; rostrocaudal gradient of symptoms | atp1a3aAutosomal dominant | Alpha 3 subunit of Na/K ATPase | Subunit of Na/K ATPase on neuronal membrane |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scuteri, D.; Hamamura, K.; Watanabe, C.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Transgenic Mice for the Translational Study of Neuropathic Pain and Dystonia. Int. J. Mol. Sci. 2022, 23, 8580. https://doi.org/10.3390/ijms23158580
Scuteri D, Hamamura K, Watanabe C, Tonin P, Bagetta G, Corasaniti MT. Transgenic Mice for the Translational Study of Neuropathic Pain and Dystonia. International Journal of Molecular Sciences. 2022; 23(15):8580. https://doi.org/10.3390/ijms23158580
Chicago/Turabian StyleScuteri, Damiana, Kengo Hamamura, Chizuko Watanabe, Paolo Tonin, Giacinto Bagetta, and Maria Tiziana Corasaniti. 2022. "Transgenic Mice for the Translational Study of Neuropathic Pain and Dystonia" International Journal of Molecular Sciences 23, no. 15: 8580. https://doi.org/10.3390/ijms23158580
APA StyleScuteri, D., Hamamura, K., Watanabe, C., Tonin, P., Bagetta, G., & Corasaniti, M. T. (2022). Transgenic Mice for the Translational Study of Neuropathic Pain and Dystonia. International Journal of Molecular Sciences, 23(15), 8580. https://doi.org/10.3390/ijms23158580